- Регуляция экспрессии генов

Содержание
- 3. Тема лекции Регуляция экспрессии генов
- 4. Зачем необходима регуляция экспрессии генов? Позволяет приспособить организм к изменяющимся условиям среды. В клетке в каждый
- 5. Особенности экспрессии генов (1)
- 6. Особенности экспрессии генов (2)
- 7. Типы генов (1) Конститутивные («гены домашнего хозяйства») – постоянно включены (гены тРНК, рРНК, ДНК-полимеразы, гистонов, рибосомальных
- 8. Типы генов (2) Структурные гены – несут информацию о белках. Функциональные гены: гены, регулирующие работу структурных
- 9. Теория регуляции активности генов Жакоба и Моно (1961 г)
- 10. Транскрипционная единица у прокариот
- 11. Разберемся в терминах… Гены-операторы включают и выключают структурные гены для транскрипции (активны непостоянно). Гены-регуляторы кодируют синтез
- 12. Типы регуляции
- 13. Пример негативной регуляции https://ppt-online.org/280599
- 14. Механизм негативной регуляции При отсутствии индуктора белок-репрессор блокирует оператор и оперон неактивен. Если в клетку поступает
- 15. Общая теория регуляции активности генов (Жакоб и Моно, 1961г.) https://en.ppt-online.org/382209
- 16. Схема работы лактозного оперона https://en.ppt-online.org/382209
- 17. Механизм регуляции на примере триптофанового оперона (1) Синтез триптофановых ферментов контролируются содержанием триптофана в клетке. Белок-репрессор
- 18. Механизм регуляции на примере триптофанового оперона (2)
- 19. Регуляция активности генов у эукариот Схема регуляции была разработана Г.П.Георгиевым (1972). Принцип регуляции (по типу обратной
- 20. Регуляция активности генов у эукариот происходит на уровнях: Инактивированная
- 21. Факторы контроля экспрессии генов цис-генетические (последовательности ДНК) Боксы для инициации транскрипции Тканеспецифичные боксы Индуцируемые боксы Энхансеры,
- 22. Уровни контроля экспрессии генов Претранскрипционный – деконденсация ДНК, изменения гистонов, деметилирование ДНК (эпигенетический, обратимый, генетически обусловленный)
- 23. Претранскрипционный контроль 1. Изменение гистонов: метилирование H3 (Lys4) – активация генов метилирование H3 (Lys9) – уменьшение
- 26. Претранскрипционный контроль 2. Метилирование ДНК (островки 5MeCpG, обычно в промоторе) – инактивация транскрипции. !!! Метилирование ДНК
- 27. Активные гены расположены в гиперацетилированном хроматине, неактивные – в гипоацетилированном -При онкологических заболеваниях и вирусных инфекциях
- 28. Транскрипционный контроль один из основных уровней, осуществляется при участии факторов транскрипции и модуляторных последовательностей, характеризуется двумя
- 29. Контроль транскрипции !!! Образование комплекса инициации !!! Контроль уровня транскрипции !!! Активация генов при помощи специфических
- 30. https://ppt-online.org/280599
- 31. Схема транскриптона
- 32. Регуляции транскрипции у эукариот Работу транскриптона регулируют несколько генов-регуляторов Индукторами могут быть сложные белки, например гормоны
- 33. Усиление и ослабление транскрипции Энхансеры – участки ДНК вне промотора, которые связываются с определенными факторами транскрипции
- 34. Адаптивная регуляция транскрипции у эукариот
- 36. Посттранскрипционный контроль осуществляется после транскрипции; характеризуется следующими механизмами: - процессинг и сплайсинг, - инактивация мРНК, -
- 40. Посттранскрипционные изменения мРНК (редактирование мРНК)
- 41. Посттранскрипционные изменения мРНК
- 42. Трансляционный контроль Выбор мРНК для трансляции Локализация мРНК в цитоплазме Количество рибосом Нетранслируемый участок Факторы трансляции
- 43. Факторы трансляции Инициация eIF-2 - активирует комплекс Met-ARNtmet eIF-3 – обеспечивает присоединение 40S к мРНК eIF-4A
- 44. Элонгация eEF-1α – лрисоединение аминоацит-тРНК к рибосоме eEF-1βγ – гидролиз ГТФ eEF-2 – транслокация рибосомы Tерминация
- 45. Продолжительность жизни мРНК
- 46. Механизмы деградации мРНК
- 47. Механизмы реализации генетической информации у эукариот ДНК-мРНК-белок- биохимическая реакция-признак Другие гены Факторы среды Признак
- 48. Представим себе возможный механизм дифференцировки стволовых клеток… Недифференцированные клетки имеют в цитоплазме разные молекулы-индукторы Молекулы индукторы
- 49. Следовательно… Главный механизм дифференцировки клеток – это активация и подавление различных транскриптонов на определенных этапах развития
- 50. Изменение активности генов, не затрагивающее первичную структуру ДНК, но влияющее на проявление тех или иных свойств
- 51. Эпигенетика и старение человека При старении уменьшается число генов, «работающих» в молодости, и активируются гены, которые
- 52. «Генетика предполагает, а эпигенетика располагает» Peter B.Medawar, английский биолог
- 54. Скачать презентацию


Тема лекции
Регуляция экспрессии генов
Тема лекции
Регуляция экспрессии генов

Зачем необходима регуляция экспрессии генов?
Позволяет приспособить организм к изменяющимся условиям среды.
В
Зачем необходима регуляция экспрессии генов?
Позволяет приспособить организм к изменяющимся условиям среды.
В

Особенности экспрессии генов (1)
Особенности экспрессии генов (1)

Особенности экспрессии генов (2)
Особенности экспрессии генов (2)

Типы генов (1)
Конститутивные («гены домашнего хозяйства») – постоянно включены (гены тРНК,
Типы генов (1)
Конститутивные («гены домашнего хозяйства») – постоянно включены (гены тРНК,

Типы генов (2)
Структурные гены – несут информацию о белках.
Функциональные гены:
Типы генов (2)
Структурные гены – несут информацию о белках.
Функциональные гены:

Теория регуляции активности генов Жакоба и Моно (1961 г)
Теория регуляции активности генов Жакоба и Моно (1961 г)

Транскрипционная единица у прокариот
Транскрипционная единица у прокариот

Разберемся в терминах…
Гены-операторы включают и выключают структурные гены для транскрипции (активны
Разберемся в терминах…
Гены-операторы включают и выключают структурные гены для транскрипции (активны

Типы регуляции
Типы регуляции

Пример негативной регуляции
https://ppt-online.org/280599
Пример негативной регуляции
https://ppt-online.org/280599

Механизм негативной регуляции
При отсутствии индуктора белок-репрессор блокирует оператор и оперон
Механизм негативной регуляции
При отсутствии индуктора белок-репрессор блокирует оператор и оперон

Общая теория регуляции активности генов (Жакоб и Моно, 1961г.)
https://en.ppt-online.org/382209
Общая теория регуляции активности генов (Жакоб и Моно, 1961г.)
https://en.ppt-online.org/382209

Схема работы лактозного оперона
https://en.ppt-online.org/382209
Схема работы лактозного оперона
https://en.ppt-online.org/382209

Механизм регуляции
на примере триптофанового оперона (1)
Синтез триптофановых ферментов контролируются содержанием
Механизм регуляции
на примере триптофанового оперона (1)
Синтез триптофановых ферментов контролируются содержанием

Механизм регуляции
на примере триптофанового оперона (2)
Механизм регуляции
на примере триптофанового оперона (2)

Регуляция активности генов у эукариот
Схема регуляции была разработана Г.П.Георгиевым (1972).
Принцип
Регуляция активности генов у эукариот
Схема регуляции была разработана Г.П.Георгиевым (1972).
Принцип

Регуляция активности генов у эукариот
происходит на уровнях:
Инактивированная
Регуляция активности генов у эукариот
происходит на уровнях:
Инактивированная

Факторы контроля экспрессии генов
цис-генетические
(последовательности ДНК)
Боксы для инициации транскрипции
Тканеспецифичные боксы
Индуцируемые боксы
Энхансеры,
Факторы контроля экспрессии генов
цис-генетические
(последовательности ДНК)
Боксы для инициации транскрипции
Тканеспецифичные боксы
Индуцируемые боксы
Энхансеры,

Уровни контроля экспрессии генов
Претранскрипционный – деконденсация ДНК, изменения гистонов, деметилирование ДНК
Уровни контроля экспрессии генов
Претранскрипционный – деконденсация ДНК, изменения гистонов, деметилирование ДНК

Претранскрипционный контроль
1. Изменение гистонов:
метилирование H3 (Lys4) – активация генов
метилирование H3
Претранскрипционный контроль
1. Изменение гистонов:
метилирование H3 (Lys4) – активация генов
метилирование H3



Претранскрипционный контроль
2. Метилирование ДНК (островки 5MeCpG, обычно в промоторе) – инактивация
Претранскрипционный контроль
2. Метилирование ДНК (островки 5MeCpG, обычно в промоторе) – инактивация

Активные гены расположены в гиперацетилированном хроматине, неактивные – в гипоацетилированном
-При онкологических
Активные гены расположены в гиперацетилированном хроматине, неактивные – в гипоацетилированном
-При онкологических

Транскрипционный контроль
один из основных уровней,
осуществляется при участии факторов транскрипции и модуляторных
Транскрипционный контроль
один из основных уровней,
осуществляется при участии факторов транскрипции и модуляторных

Контроль транскрипции
!!! Образование комплекса инициации
!!! Контроль уровня транскрипции
!!! Активация генов при
Контроль транскрипции
!!! Образование комплекса инициации
!!! Контроль уровня транскрипции
!!! Активация генов при

https://ppt-online.org/280599
https://ppt-online.org/280599

Схема транскриптона
Схема транскриптона

Регуляции транскрипции у эукариот
Работу транскриптона регулируют несколько генов-регуляторов
Индукторами могут быть сложные
Регуляции транскрипции у эукариот
Работу транскриптона регулируют несколько генов-регуляторов
Индукторами могут быть сложные

Усиление и ослабление транскрипции
Энхансеры – участки ДНК вне промотора, которые связываются
Усиление и ослабление транскрипции
Энхансеры – участки ДНК вне промотора, которые связываются

Адаптивная регуляция транскрипции у эукариот
Адаптивная регуляция транскрипции у эукариот


Посттранскрипционный контроль
осуществляется после транскрипции;
характеризуется следующими механизмами:
- процессинг и сплайсинг,
-
Посттранскрипционный контроль
осуществляется после транскрипции;
характеризуется следующими механизмами:
- процессинг и сплайсинг,
-




Посттранскрипционные изменения мРНК (редактирование мРНК)
Посттранскрипционные изменения мРНК (редактирование мРНК)

Посттранскрипционные изменения мРНК
Посттранскрипционные изменения мРНК

Трансляционный контроль
Выбор мРНК для трансляции
Локализация мРНК в цитоплазме
Количество рибосом
Нетранслируемый участок
Факторы трансляции
Трансляционный контроль
Выбор мРНК для трансляции
Локализация мРНК в цитоплазме
Количество рибосом
Нетранслируемый участок
Факторы трансляции

Факторы трансляции
Инициация
eIF-2 - активирует комплекс Met-ARNtmet
eIF-3 – обеспечивает присоединение 40S к
Факторы трансляции
Инициация
eIF-2 - активирует комплекс Met-ARNtmet
eIF-3 – обеспечивает присоединение 40S к

Элонгация
eEF-1α – лрисоединение аминоацит-тРНК к рибосоме
eEF-1βγ – гидролиз ГТФ
eEF-2 – транслокация
Элонгация
eEF-1α – лрисоединение аминоацит-тРНК к рибосоме
eEF-1βγ – гидролиз ГТФ
eEF-2 – транслокация

Продолжительность жизни мРНК
Продолжительность жизни мРНК

Механизмы деградации мРНК
Механизмы деградации мРНК

Механизмы реализации генетической информации у эукариот
ДНК-мРНК-белок- биохимическая реакция-признак
Другие гены
Факторы среды
Признак
Механизмы реализации генетической информации у эукариот
ДНК-мРНК-белок- биохимическая реакция-признак
Другие гены
Факторы среды
Признак

Представим себе возможный механизм дифференцировки стволовых клеток…
Недифференцированные клетки имеют в
Представим себе возможный механизм дифференцировки стволовых клеток…
Недифференцированные клетки имеют в

Следовательно…
Главный механизм дифференцировки клеток – это активация и подавление различных транскриптонов
Следовательно…
Главный механизм дифференцировки клеток – это активация и подавление различных транскриптонов

Изменение активности генов, не затрагивающее первичную структуру ДНК, но влияющее на проявление тех или
Изменение активности генов, не затрагивающее первичную структуру ДНК, но влияющее на проявление тех или

Эпигенетика и старение человека
При старении уменьшается число генов, «работающих» в молодости, и активируются
Эпигенетика и старение человека
При старении уменьшается число генов, «работающих» в молодости, и активируются

«Генетика предполагает, а эпигенетика располагает»
Peter B.Medawar, английский биолог
«Генетика предполагает, а эпигенетика располагает»
Peter B.Medawar, английский биолог
 Фізіологія крові
Фізіологія крові Синюха голубая - Polemonium caeruleum
Синюха голубая - Polemonium caeruleum Подкласс гамамелидиды
Подкласс гамамелидиды Эволюция. Доказательства эволюции (часть 6)
Эволюция. Доказательства эволюции (часть 6) Фундаментальные свойства живого
Фундаментальные свойства живого Різноманітність плазунів
Різноманітність плазунів Использование фитонцидоносных растений для очистки воздуха
Использование фитонцидоносных растений для очистки воздуха Анатомия и физиология нервной системы
Анатомия и физиология нервной системы Морфология побега растений. Разнообразие побегов
Морфология побега растений. Разнообразие побегов Кто что даёт. Домашние животные. Для детей 2-4 лет
Кто что даёт. Домашние животные. Для детей 2-4 лет Строение стеблей однодольных растений
Строение стеблей однодольных растений Биология – наука о жизни
Биология – наука о жизни 20231204_35.obobshchenie_po_teme_teoriya_evolyutsii
20231204_35.obobshchenie_po_teme_teoriya_evolyutsii Neisseria. General Characteristics of Neisseria spp
Neisseria. General Characteristics of Neisseria spp Жизнь в океане
Жизнь в океане Сезонные изменения в жизни организмов
Сезонные изменения в жизни организмов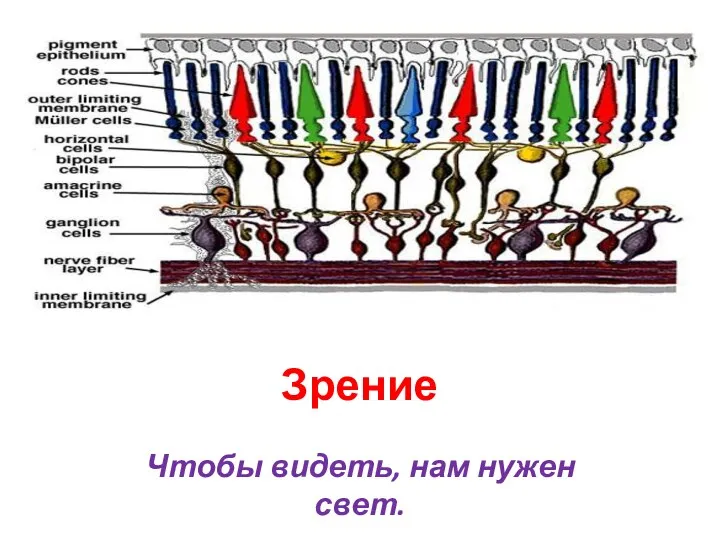 Зрение. Чтобы видеть, нам нужен свет
Зрение. Чтобы видеть, нам нужен свет Антропогенез. Положение человека в системе органического мира. Этапы эволюции человека
Антропогенез. Положение человека в системе органического мира. Этапы эволюции человека Пищеварение беспозвоночных
Пищеварение беспозвоночных Иглокожие
Иглокожие Генная инженерия
Генная инженерия Жасушаның негізгі компаненттері
Жасушаның негізгі компаненттері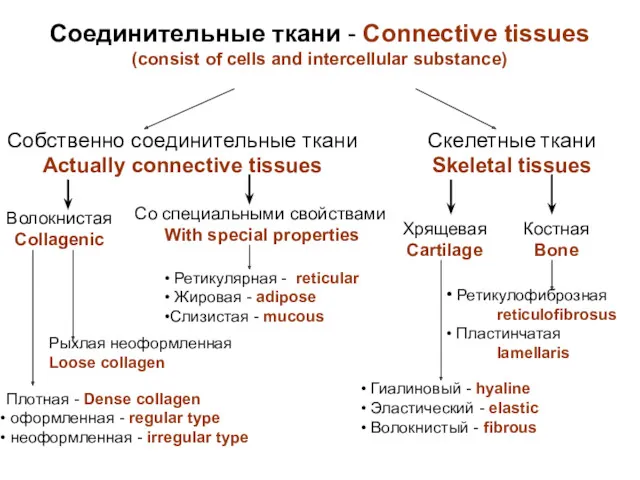 Соединительныя ткани
Соединительныя ткани Обмен веществ и энергии в клетке
Обмен веществ и энергии в клетке Гормоны. Классификация гормонов
Гормоны. Классификация гормонов Вкусовые ощущения
Вкусовые ощущения Циклы споровых растений. Задачи на моногибридное скрещивание
Циклы споровых растений. Задачи на моногибридное скрещивание Особо охраняемые территории Ленинградской области. Заказник Раковые озера
Особо охраняемые территории Ленинградской области. Заказник Раковые озера