- Гормоны. Классификация гормонов

Содержание
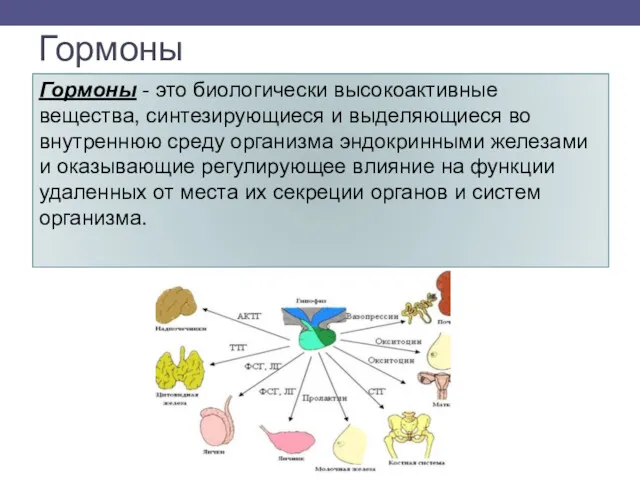
- 2. Гормоны Гормоны - это биологически высокоактивные вещества, синтезирующиеся и выделяющиеся во внутреннюю среду организма эндокринными железами
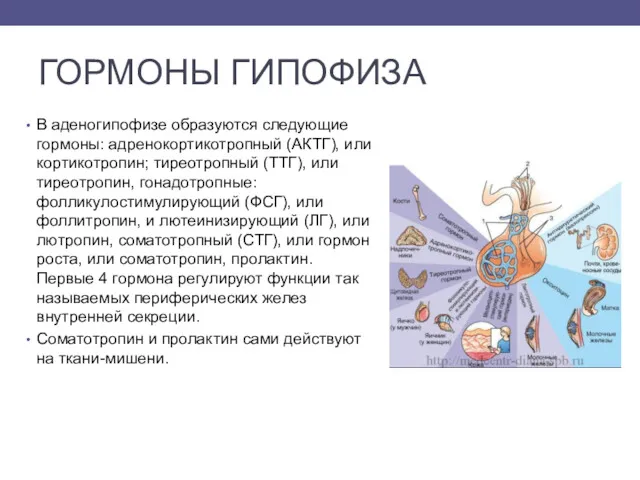
- 3. ГОРМОНЫ ГИПОФИЗА В аденогипофизе образуются следующие гормоны: адренокортикотропный (АКТГ), или кортикотропин; тиреотропный (ТТГ), или тиреотропин, гонадотропные:
- 4. СОМАТОТРОПНЫЙ ГОРМОН Строение Представляет собой пептид, включающий 191 аминокислоту с молекулярной массой 22 кД и периодом
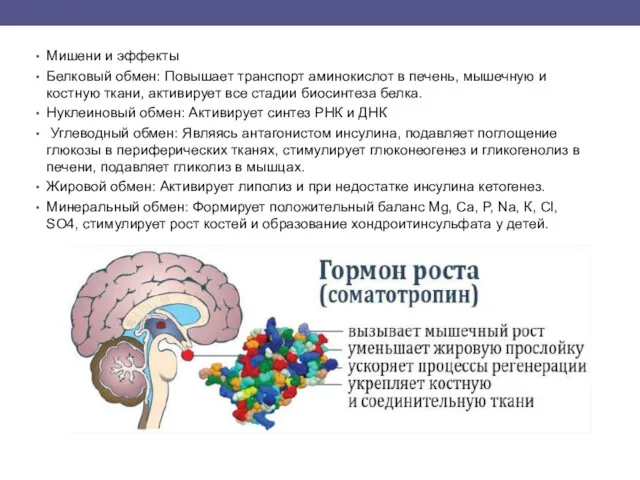
- 5. Мишени и эффекты Белковый обмен: Повышает транспорт аминокислот в печень, мышечную и костную ткани, активирует все
- 6. Адренокортикотропный гормон (АКТГ) Кортикотропин синтезируется в базофильных клетках аденогипофиза, и продукция его стимулируется кортиколиберином гипоталамуса. По
- 7. Вненадпочечниковые эффекты АКТГ заключаются в стимуляции липолиза (мобилизует жиры из жировых депо и способствует окислению жиров),
- 8. Тиреотропный гормон (ТТГ), Тиреотропный гормон (ТТГ), или тиреотропин – это димерный белок, состоящий из двух неодинаковых
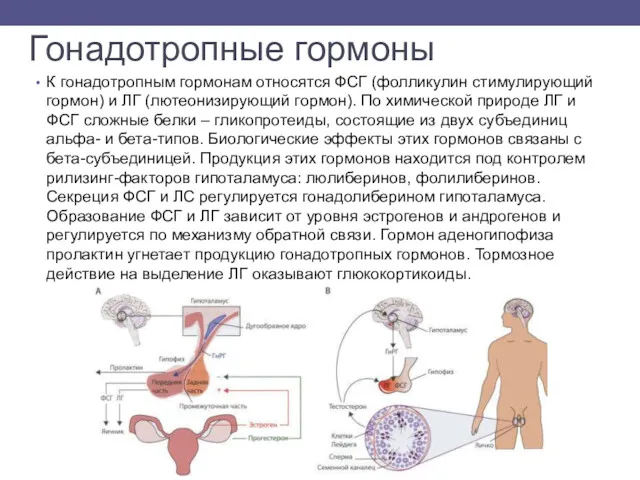
- 9. Гонадотропные гормоны К гонадотропным гормонам относятся ФСГ (фолликулин стимулирующий гормон) и ЛГ (лютеонизирующий гормон). По химической
- 10. Лютропин содержит альфа-субъединицу, состоящую из 88 и бета-субъединицу из 119 аминокислотных остатков. На ранних стадиях эмбриогенеза
- 11. ЛГ – главный индуктор формирования желтых тел и синтеза ими прогестерона. Отсюда второе название пролактина –
- 12. Фолликулостимулирующий гормон Фоллитропин, как и ЛГ, состоит из двух субъединиц гликопротеидной природы. ФСГ вызывает рост и
- 13. Пролактин Пролактин – лактогенный гормон – является мономерным белком, состоящим из 199 аминокислотных остатков, сходным по
- 14. Образование пролактина регулируется пролактолиберином и пролактостатином гипоталамуса, а также в меньшей степени другими пептидами гипоталамуса .
- 15. Гормоны задней доли гипофиза Эти гормоны образуются в гипоталамусе. В нейрогипофизе происходит их накопление. В клетках
- 16. Антидиуретический гормон (АДГ), или вазопрессин, осуществляет в организме антидиуретическое действие, которое выражается в стимуляции реабсорбции воды
- 17. Окситоцин Представляет собой пептид, состоящий из 9 аминокислот и периодом полураспада 5 минут. Окситоцин имеет рецепторы
- 18. Щитовидная железа К гормонам щитовидной железы относятся йодтиронины – тироксин (Т4) и трийодтиронин (Т3), являющиеся производными
- 19. Представляют собой йодированные производные аминокислоты тирозина. Синтез оуществляется в фолликулярных клетках щитовидной железы. Йодиды, поступающие из
- 20. Активирует синтез тиреотропный гормон. Уменьшают синтез по механизму обратной отрицательной связи тироксин и трийодтиронин, высокие концентрации
- 21. Кальцитонин или тиреокальцитонин, вместе с паратгормоном околощитовидных желез участвует в регуляции кальциевого обмена. Под его влиянием
- 22. Околощитовидные (паращитовидные) железы Человек имеет 2 пары околощитовидных желез, расположенных на задней поверхности или погруженных внутри
- 23. ПОЛОВЫЕ ГОРМОНЫ Строение Являются производными холестерола – стероиды. Синтез Женские гормоны: эстрогены синтезируются в фолликулах яичников,
- 24. Эстрогены Синтез эстрогенов последовательно активируется ЛГ и ФСГ, прогестерона – ЛГ. Подавляется синтез по механизму обратной
- 25. Мишени и эффекты Эстрогены активируют синтез белка и нуклеиновых кислот в органах половой сферы 1. При
- 26. МУЖСКИЕ ПОЛОВЫЕ ГОРМОНЫ Синтез Мужские гормоны: на 95% тестостерон синтезируется в клетках Лейдига семенников и на
- 27. Кора надпочечников Клубочковая зона В клубочковой зоне образуются гормоны, называемые минералокортикоидами. К ним относятся: Альдостерон Кортикостерон
- 28. МИНЕРАЛОКОРТИКОИДЫ Производные холестерола – стероиды. Основным гормоном у человека является альдостерон. Синтез Осуществляется в клубочковой зоне
- 29. Г ЛЮКОКОРТИКОИДЫ Производные холестерола – стероиды. Основным гормоном у человека является кортизол. Синтез Осуществляется в сетчатой
- 30. Углеводный обмен Стимуляция глюконеогенеза посредством увеличения синтеза фосфоенолпируват– карбоксикиназы (совместно с глюкагоном) и синтеза аминотрансфераз, обеспечивающих
- 31. Гормоны мозгового вещества надпочечников В хромаффинных клетках мозгового вещества надпочечников из тирозина через стадию образования дофамина
- 32. Адреналин – типичный гормон мозгового вещества надпочечников, очень лабильное соединение с выраженными оксидо-редукторными свойствами, повышающий тонус
- 33. ИНСУЛИН Представляет собой полипептид, состоящий из двух цепей А и В, связанных между собой дисульфидными мостиками,
- 34. Мишени и эффекты Печень • активация гликолиза и гликогеногенеза • подавление глюконеогенеза . • усиление синтеза
- 36. Скачать презентацию
Гормоны
Гормоны - это биологически высокоактивные вещества, синтезирующиеся и выделяющиеся во внутреннюю
Гормоны
Гормоны - это биологически высокоактивные вещества, синтезирующиеся и выделяющиеся во внутреннюю
ГОРМОНЫ ГИПОФИЗА
В аденогипофизе образуются следующие гормоны: адренокортикотропный (АКТГ), или кортикотропин; тиреотропный
ГОРМОНЫ ГИПОФИЗА
В аденогипофизе образуются следующие гормоны: адренокортикотропный (АКТГ), или кортикотропин; тиреотропный

СОМАТОТРОПНЫЙ ГОРМОН
Строение Представляет собой пептид, включающий 191 аминокислоту с молекулярной
СОМАТОТРОПНЫЙ ГОРМОН
Строение Представляет собой пептид, включающий 191 аминокислоту с молекулярной
Мишени и эффекты
Белковый обмен: Повышает транспорт аминокислот в печень, мышечную
Мишени и эффекты
Белковый обмен: Повышает транспорт аминокислот в печень, мышечную

Адренокортикотропный гормон (АКТГ)
Кортикотропин синтезируется в базофильных клетках аденогипофиза, и продукция его
Адренокортикотропный гормон (АКТГ)
Кортикотропин синтезируется в базофильных клетках аденогипофиза, и продукция его

Вненадпочечниковые эффекты АКТГ заключаются в стимуляции липолиза (мобилизует жиры из жировых
Вненадпочечниковые эффекты АКТГ заключаются в стимуляции липолиза (мобилизует жиры из жировых

Тиреотропный гормон (ТТГ),
Тиреотропный гормон (ТТГ), или тиреотропин – это димерный
Тиреотропный гормон (ТТГ),
Тиреотропный гормон (ТТГ), или тиреотропин – это димерный
Гонадотропные гормоны
К гонадотропным гормонам относятся ФСГ (фолликулин стимулирующий гормон) и ЛГ
Гонадотропные гормоны
К гонадотропным гормонам относятся ФСГ (фолликулин стимулирующий гормон) и ЛГ

Лютропин содержит альфа-субъединицу, состоящую из 88 и бета-субъединицу из 119 аминокислотных
Лютропин содержит альфа-субъединицу, состоящую из 88 и бета-субъединицу из 119 аминокислотных
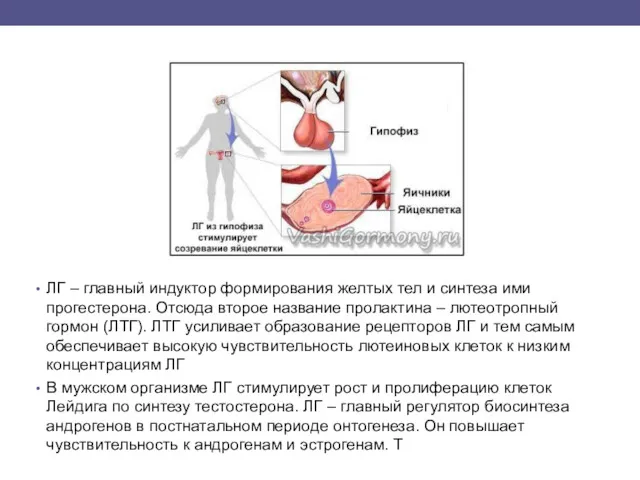
ЛГ – главный индуктор формирования желтых тел и синтеза ими прогестерона.
ЛГ – главный индуктор формирования желтых тел и синтеза ими прогестерона.

Фолликулостимулирующий гормон
Фоллитропин, как и ЛГ, состоит из двух субъединиц гликопротеидной природы.
ФСГ
Фолликулостимулирующий гормон
Фоллитропин, как и ЛГ, состоит из двух субъединиц гликопротеидной природы.
ФСГ

Пролактин
Пролактин – лактогенный гормон – является мономерным белком, состоящим из 199
Пролактин
Пролактин – лактогенный гормон – является мономерным белком, состоящим из 199
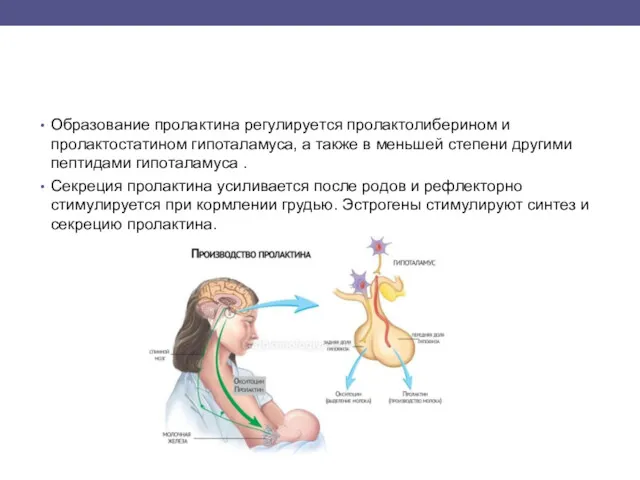
Образование пролактина регулируется пролактолиберином и пролактостатином гипоталамуса, а также в меньшей
Образование пролактина регулируется пролактолиберином и пролактостатином гипоталамуса, а также в меньшей
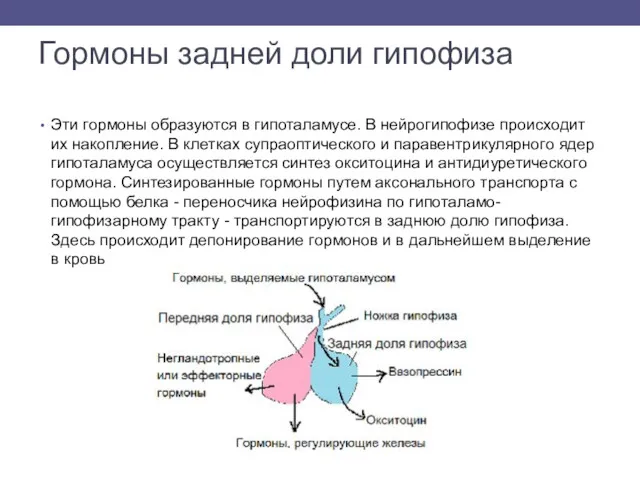
Гормоны задней доли гипофиза
Эти гормоны образуются в гипоталамусе. В нейрогипофизе происходит
Гормоны задней доли гипофиза
Эти гормоны образуются в гипоталамусе. В нейрогипофизе происходит
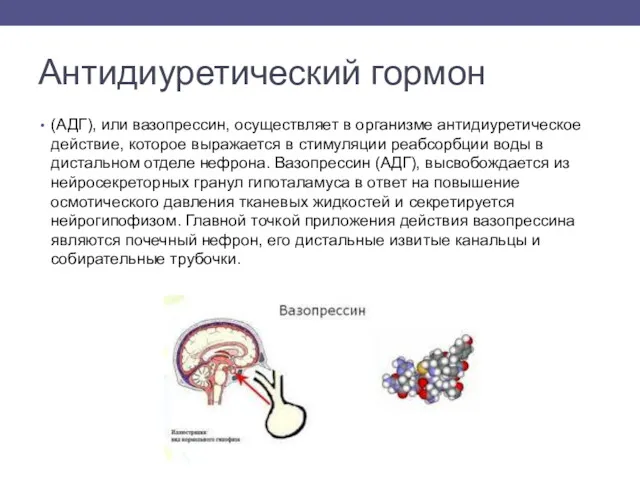
Антидиуретический гормон
(АДГ), или вазопрессин, осуществляет в организме антидиуретическое действие, которое
Антидиуретический гормон
(АДГ), или вазопрессин, осуществляет в организме антидиуретическое действие, которое

Окситоцин
Представляет собой пептид, состоящий из 9 аминокислот и периодом полураспада 5
Окситоцин
Представляет собой пептид, состоящий из 9 аминокислот и периодом полураспада 5

Щитовидная железа
К гормонам щитовидной железы относятся йодтиронины – тироксин (Т4) и
Щитовидная железа
К гормонам щитовидной железы относятся йодтиронины – тироксин (Т4) и

Представляют собой йодированные производные аминокислоты тирозина.
Синтез оуществляется в фолликулярных клетках щитовидной
Представляют собой йодированные производные аминокислоты тирозина.
Синтез оуществляется в фолликулярных клетках щитовидной

Активирует синтез тиреотропный гормон. Уменьшают синтез по механизму обратной отрицательной связи
Активирует синтез тиреотропный гормон. Уменьшают синтез по механизму обратной отрицательной связи
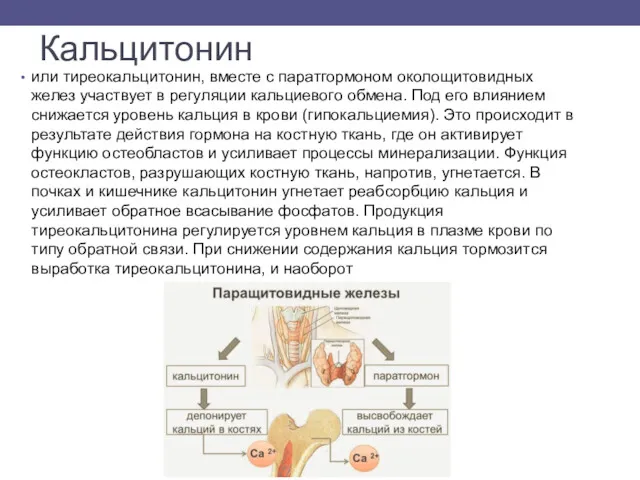
Кальцитонин
или тиреокальцитонин, вместе с паратгормоном околощитовидных желез участвует в регуляции кальциевого
Кальцитонин
или тиреокальцитонин, вместе с паратгормоном околощитовидных желез участвует в регуляции кальциевого

Околощитовидные (паращитовидные) железы
Человек имеет 2 пары околощитовидных желез, расположенных на задней
Околощитовидные (паращитовидные) железы
Человек имеет 2 пары околощитовидных желез, расположенных на задней

ПОЛОВЫЕ ГОРМОНЫ
Строение Являются производными холестерола – стероиды.
Синтез Женские гормоны:
ПОЛОВЫЕ ГОРМОНЫ
Строение Являются производными холестерола – стероиды.
Синтез Женские гормоны:

Эстрогены
Синтез эстрогенов последовательно активируется ЛГ и ФСГ, прогестерона – ЛГ.
Подавляется синтез
Эстрогены
Синтез эстрогенов последовательно активируется ЛГ и ФСГ, прогестерона – ЛГ.
Подавляется синтез

Мишени и эффекты
Эстрогены активируют синтез белка и нуклеиновых кислот в
Мишени и эффекты
Эстрогены активируют синтез белка и нуклеиновых кислот в

МУЖСКИЕ ПОЛОВЫЕ ГОРМОНЫ
Синтез Мужские гормоны: на 95% тестостерон синтезируется в
МУЖСКИЕ ПОЛОВЫЕ ГОРМОНЫ
Синтез Мужские гормоны: на 95% тестостерон синтезируется в

Кора надпочечников
Клубочковая зона
В клубочковой зоне образуются гормоны, называемые минералокортикоидами. К ним относятся:
Альдостерон
Кортикостерон —
Кора надпочечников
Клубочковая зона
В клубочковой зоне образуются гормоны, называемые минералокортикоидами. К ним относятся:
Альдостерон
Кортикостерон —

МИНЕРАЛОКОРТИКОИДЫ
Производные холестерола – стероиды. Основным гормоном у человека является альдостерон. Синтез
МИНЕРАЛОКОРТИКОИДЫ
Производные холестерола – стероиды. Основным гормоном у человека является альдостерон. Синтез

Г ЛЮКОКОРТИКОИДЫ
Производные холестерола – стероиды. Основным гормоном у человека является
Г ЛЮКОКОРТИКОИДЫ
Производные холестерола – стероиды. Основным гормоном у человека является

Углеводный обмен Стимуляция глюконеогенеза посредством увеличения синтеза фосфоенолпируват– карбоксикиназы (совместно с
Углеводный обмен Стимуляция глюконеогенеза посредством увеличения синтеза фосфоенолпируват– карбоксикиназы (совместно с

Гормоны мозгового вещества надпочечников
В хромаффинных клетках мозгового вещества надпочечников из тирозина
Гормоны мозгового вещества надпочечников
В хромаффинных клетках мозгового вещества надпочечников из тирозина

Адреналин – типичный гормон мозгового вещества надпочечников, очень лабильное соединение с
Адреналин – типичный гормон мозгового вещества надпочечников, очень лабильное соединение с

ИНСУЛИН
Представляет собой полипептид, состоящий из двух цепей А и В,
ИНСУЛИН
Представляет собой полипептид, состоящий из двух цепей А и В,
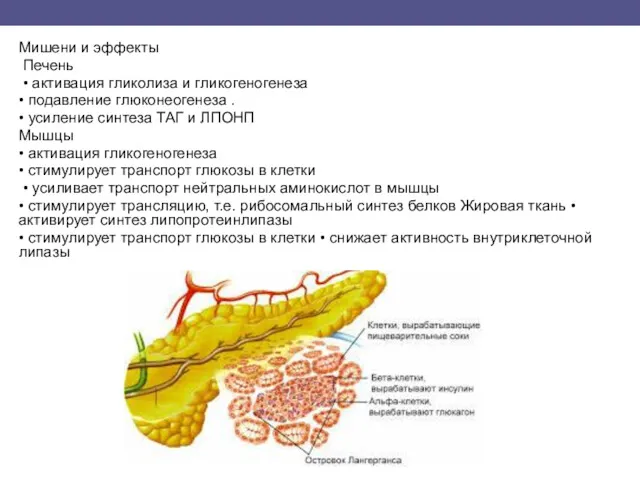
Мишени и эффекты
Печень
• активация гликолиза и гликогеногенеза
• подавление
Мишени и эффекты
Печень
• активация гликолиза и гликогеногенеза
• подавление
 Перелетные птицы весной
Перелетные птицы весной Генетика микроорганизмов
Генетика микроорганизмов Модификационная изменчивость. Построение диаграмм
Модификационная изменчивость. Построение диаграмм Вестники весны
Вестники весны Предки человека
Предки человека Лесная фитопатология. Болезни питомников, лесных культур и молодняков. Инфекционное полегание всходов
Лесная фитопатология. Болезни питомников, лесных культур и молодняков. Инфекционное полегание всходов Физиология дыхания
Физиология дыхания Ткани. Мышечная, нервная ткань
Ткани. Мышечная, нервная ткань Морские обитатели
Морские обитатели Человек и природа
Человек и природа Биология 5 класс
Биология 5 класс Будова і функції лейкоцитів
Будова і функції лейкоцитів Выделение. Процесс выделения у растений и животных
Выделение. Процесс выделения у растений и животных Фотоальбом. Собаки
Фотоальбом. Собаки Основные этапы антропогенеза. 9 класс
Основные этапы антропогенеза. 9 класс The First Three Weeks of Human Embryogenesis
The First Three Weeks of Human Embryogenesis Высшие споровые растения
Высшие споровые растения Желудочно-кишечный тракт. Пищеварительная система
Желудочно-кишечный тракт. Пищеварительная система Осень и здоровье человека
Осень и здоровье человека Физиология слухового и вестибулярного анализаторов
Физиология слухового и вестибулярного анализаторов Введение в зоологию. Подцарство Одноклеточные (Protozoa)
Введение в зоологию. Подцарство Одноклеточные (Protozoa) Мир и человек. Живые обитатели планеты
Мир и человек. Живые обитатели планеты Биохимия на региональном этапе ВОШ
Биохимия на региональном этапе ВОШ Общая вирусология
Общая вирусология Наследственная информация и ее реализация в клетке
Наследственная информация и ее реализация в клетке Рослини і тварини Сумщини занесені до Червоної книги України
Рослини і тварини Сумщини занесені до Червоної книги України Растительный мир Алтайского края
Растительный мир Алтайского края Молекулярно-генетический уровень организации живого
Молекулярно-генетический уровень организации живого