- Системы фоторецепторов. Эффекты фитохромов

Содержание
- 2. Системы фоторецепторов тесно взаимодействуют
- 3. Фитохром. Принцип действия. (Bortwick H., Hendriks S. PNAS, 1952).
- 4. Фотообращаемые эффекты фитохромов. Покрытосеменные салат-латук семена Стимуляция прорастания овес этиолированные Стимуляция деэтиоляции проростки горчица проростки Стимуляция
- 5. Эффекты фитохромов можно разделить по их зависимости от интенсивности света VLFR (very low fluence responses) 0,1
- 6. Фитохромов несколько. Спектры поглощения фитохромов А и В Соотношение красного и дальнего красного света в разных
- 7. Фитохромы - структура спектр поглощения NB – фотостационарное состояние фитохрома Апопротеин: димер, мономеры ~ 120 kDa
- 8. Фотоконверсия фитохрома – цис-транс изомерия под действием света. Форма 660 (неактивная) Форма 730 (активная) Фитохром А:
- 9. Фитохромы – серин-треониновые протеинкиназы, но… Свойства фитохромов модифицируются светом. А. Спектр поглощения phyA овса в Pr
- 10. Фитохромов несколько, их активность регулируется фосфорилированием Вслед за активированием красным светом, активность фитохромов регулируется в зависимости
- 11. Фитохром А регулирует около 10% генов арабидопсиса Всего исследовали активность 8 200 генов – около трети
- 12. Откуда взялся фитохром? А. Консервативные участки фитохромов цианобактерий (Cph1) и Arabidopsis. Консервативный остаток цистеина для связывания
- 13. Фитохромы высших растений – серин-треониновая киназа, цианобактерий – гистидинкиназа… 1. Хромофор синтезируется в пластидах, апопротеин –
- 14. Схема сигналинга бактериального и эукариотического фитохромов Бактериальный фитохром типичная двухкомпонентная гистидин-киназа, которая под действием Fr переходит
- 15. Сигналинг фитохромов. Основные «игроки» и общие положения Два типа ответов – быстрые (секунды) и «глобалные». Быстрые
- 16. Как фитохром А перемещается в ядро? Быстро (минуты), свет – красный, дальний красный, синий, NB -
- 17. Перемещение фитохрома В в ядро и схема его сигналинга. Транслокация в ядро медленно (часы) и только
- 18. В сигналинге фитохромов важную роль играет убиквитирование трансфакторов и их развал в протеасоме. COP1 – это
- 19. Упрощенная схема работы фитохромов
- 20. Схема возможных путей трансдукции сигнала от фитохромов Фитохромы А или В в ядре могут регулировать активность
- 21. И обобщающая картинка PKS1
- 22. Рецепторы синего света отвечают за многое… Реакция фототропизма Движение хлоропластов Циркадные ритмы: открытие устьиц «сонные» движения
- 23. Структура фототропина и криптохрома. Фототропины: мембранносвязанные Ser-Thr- киназы. 120 kDa. В плазмалемме. Криптохромы: Cry1 – 190
- 24. Фототропин-Phy3 – «гибридный» фоторецетор Adiantum.
- 25. Схема строения фоторецепторов и их взаимодействие
- 26. Фототропины участвуют в регуляции разных фотодвижений Изгибание колеоптиля Движение хлоропластов Открывание устьиц У Arabidopsis работают два
- 27. Фототропины: строение и локализация Каждый LOV-домен нековалентно связывает хромофор ― FMN, — а также обеспечивает белок-белковые
- 28. Фототропины: сигналинг СС/УФ-А зависимое образование тиоаддукта FMN с цистеином LOV 2 домена приводит к изменению конформации
- 29. Криптохромы: фотопревращения и передача сигнала У криптохромов MTHF (птерин) поглощает квант УФ/СС и передаёт энергию на
- 30. Криптохромы: фотопревращения, передача сигнала Криптохромы – ядерные белки На свету cry1 выходит из ядра; cry2 –
- 31. Криптохромы: возможный механизм фосфорилирования и трансдукции сигнала Два возможных варинта передачи сигнала: 1.Через активацию транс-факкторов CIBs
- 32. Роль «биологических часов» в жизни растения а. Экспрессия некоторых генов подчинена циркадным ритмам. Гены, кодирующие хлорофилл-а/в-связывающие
- 33. Компоненты системы «биологических часов»
- 34. Молекулярная модель эукариотического циркадного осциллятора a. Общая схема, показывающая главную петлю обратной связи - основу для
- 35. Упрощенная модель циркадного осциллятора Arabidopsis . Модель основана на обратной связи между LHY, CCA1 и TOC1.
- 36. Модель циркадной системы Arabidopsis Входной сигнал - Сry1 и Сry2 – криптохромы, PhyA, PhyB - фитохромы
- 37. То же , но в другом виде и с некоторыми вариациями… Вход светового сигнала осуществляется через
- 38. Стратегии развития животных и растений различны. У животных: развитие – серия сложных преобразований, которые ведут к
- 39. Этап 1. От зиготы до семени. Эмбриогенез у арабидопсиса Установление оси «корень – побег». Зигота делится
- 40. Особенности и механизмы эмбрионального развития растений Формирование зародыша у растений происходит за счет изменения скорости и
- 41. Главную роль в регуляции эмбриогенеза играет ауксин Мутанты по эмбриогенезу у Arabidopsis GURK – отсутствуют апикальная
- 42. Наиболее «ранние» регуляторы эмбриогенеза: гены, контролирующие полярный транспорт ауксинов полярность развития зародыша со стадии 2х клеток
- 43. Гены-регуляторы упорядоченного деления клеток в эмбриогенезе gnom knolle keule hinkel tsd1,2 эндо-1,4-β-D-глюканаза синтаксин кинезин белок SNARE
- 44. GNOM (GN) – регулятор локализации PIN белков PIN Направление полярного транспорта ауксинов Направление растяжения клеток Направление
- 45. Гены- регуляторы развития центрального домена зародыша WT fk (fackel), hyd (hydra), cephalopod (cph) редукция гипокотиля +
- 46. Соматический эмбриогенез – модель для изучения механизмов эмбриогенеза in vivo
- 47. Соматический эмбриогенез этапы и факторы
- 48. Этап 2. Покой и прорастание семян Покой семян: АБК, этилен. В состоянии покоя могут выдерживать –
- 49. Гравитропизм и фототропизм - неравномерное распределение транспортеров ИУК Основа действия - изменения латерального транспорта ауксинов. Но
- 50. Что важно при формировании тканей корня – «история» или «позиция»? Ткани корня формируются из 4 типов
- 51. И все же главное – «позиция»… И все же основную роль в морфогенезе корня играет позиционный
- 52. Позиционный контроль – основной механизм развития корня «Убийство» лазером покоящегося центра и его последующее восстановление за
- 53. Роль ауксина и покоящегося центра в развитии корня Клетки покоящегося центра ингибируют дифференцировку окружающих клеток. Стволовые
- 54. PZ - периферическая зона - латеральная или органогенная меристема - Дочерние клетки CZ. Скорость и направление
- 55. Генетический контроль структуры и функции АМП («классическая генетика») Выявлены два классов мутантов, имеющих альтернативный фенотип: мутанты
- 56. Ауксин играет ключевую роль и в формирование побеговой апикальной меристемы и билатеральной симметрии CUP-SHAPED COTYLEDON («чашевидные
- 57. Развитие побега и работа апикальной меристемы STM – поддержание стволовых клеток SAM в недифференцированном cсостоянии WUS
- 58. Развитие SAM регулируется по типу отрицательной обратной связи WUSCHEL (WUS) – дурачок, юродивый – кодирует трансфактор
- 59. Система WOX - CLE Wang, Fiers 2010. CLE peptide signaling during plant development. Protoplasma 240: 33-43
- 60. Схема регулирования развития меристемы побега. Экспрессия гена WUS в организующем центре негативно регулируется пептидом CLV3 при
- 61. CLE - пептиды участвуют в определении судьбы клеток меристем Wang, Fiers 2010. CLE peptide signaling during
- 62. Схема регулирования развития меристемы корня: Экспрессия гена WOX5 в организующем центре негативно регулируется пептидом CLE40 при
- 63. Распределение факторов транскрипции между клетками определяет радиальное строение корня SHR – фактор транскрипции. Регулирует активность SCR
- 64. Схема регулирования развития камбиальной меристемы и меристемы бококвого корня В прокамбии а затем камбии экспрессия гена
- 65. Системы регулирования развития разных типов меристем
- 66. Этап 3. Ювенильный этап развития – «самоускорение» ростовых процессов С появлением апексов корня и побега и
- 67. Подпрограммы развития: закладка и развитие листа. Основные события в развитии листа: 1. Закладка листового примордия 2.
- 68. Подпрограммы развития: закладка и развитие листа. Разметка закладки – knotted–гены с гомеобоксом у кукурузы, stm –
- 69. Очаги деления клеток при развитии листа Arabidopsis: Трансгенное растение с cyclin1At:GUS репортерным геном. Циклин1At – маркер
- 70. Ауксины – ключевые регуляторы инициации листа Белки PIN Синтез ауксинов в молодых листьях Транспорт ауксинов в
- 71. Ауксины стимулируют закладку листьев через активацию мембранных Н-АТФаз и экспансинов Локальная индукция экспрессии гена EXP1 Закладка
- 72. Гомеозисные гены Гомеозисные гены — гены, определяющие процессы роста и дифференцировки в организме. Кодируют транскрипционные факторы,
- 73. KNOX (Knotted-like homeobox) гены, активирующиеся при развитии листа и форма листьев при их эктопической (смещенной) экспрессии
- 74. Форма розеточных листьев у мутантов по KNOX - генам
- 75. PIN транспорт ауксинов локальный максимум концентрации ауксинов закладка лопасти активация экспансинов, рост клеток, изменение направления деления
- 76. Подпрограммы развития: закладка пазушных почек и ветвление После образования листа (но никогда одновременно с листовым примордием!)
- 77. Подпрограммы развития: рост междоузлия Формирование листа → синтез ГК → транспорт в междоузлие → ГК активирует
- 78. Этап 4. Переход к генеративной стадии развития. Фотопериодизм Индукция цветения Эвокация цветения Формирование цветка.
- 79. Флоральный стимул. Гипотеза М.Х.Чайлахяна Гипотеза: У ДД растений нет гббереллинов, но есть антезины. У КД растений
- 80. Регуляция индукции цветения. «Генетическая модель». GI – Gigantea. Кодирует большой белок неизвестной функции, локализованный в ядре.
- 81. COP1 – специальная Е3-убиквитин лигаза. опосредует убиквитин-зависимую деградацию белка CO Как фоторецепторы могут влиять на индукцию
- 82. Общая схема регуляции цветения ДДР на длинном дне: фоторецепторы + часы - экспрессия СО-трансфактора в клетках-спутницах
- 83. Как совместить «биохимическую» и «молекулярно-генетическую» модели индукции цветения?
- 84. Четыре пути регулирования цветения… Фотопериодический путь. Начинается в листьях. Участвуют фитохромы и криптохромы. Различно для ДДР
- 85. «Круговое» строение цветка Иоганн Вольфганг Гете – фолиарная теория морфогенеза цветка. Эссе «Опыт объяснения метаморфоза растений»
- 86. Общая схема индукции и эвокации цветения. Ген LEAFY (LFY) – интегратор информации от разных путей индукции
- 87. Фенотип мутации lfy у арабидопсиса lfy Вместо цветков в пазухах листьев формируются боковые побеги. «Трансгенное» подтверждение
- 88. АВС-схема генетики развития цветка (теория «войны позиций) Ap1, Ap2 (4-я хромосома) – вместо чашелистиков – карпелы,
- 89. Мутанты арабидопсиса с точки зрения АВС-системы и гены. AP1, AP2 – функция А, трансфакторы, AP1 с
- 90. Сильно упрощенная схема иерархии генов флорального морфогенеза Гены с MADS-боксом изображены в квадратиках. AG - AGAMOUS;
- 91. То же, но чуть подробнее. В квадратиках – гены с MADS-бокслм
- 92. Роль miRNAs в развитии растений развитие цветка развитие листа развитие корня miR170, miR171 (GRAS транскрипционные факторы)
- 93. A. wt B. ap2-9 C. 35S::MIR172a-1 Фенотип трансгенных растений 35S::MIR172a-1 повторяет фенотипическое проявление мутации ap2-9 miR172
- 94. Дополненная модель развития цветка SHATTERPROOF 1, 2, SEEDSTICK SEPALLATA 1,2,3,4
- 95. «Модель квартета»: продукты генов идентичности органов цветка функционируют в виде гетеротетрамерных белковых комплексов (Гюнтер Тайссен, 2001г)
- 96. «Модель квартета»: продукты генов идентичности органов цветка функционируют в виде гетеротетрамерных белковых комплексов (Гюнтер Тайссен, 2001г)
- 97. Мутанты по структуре цветка сходны у разных растений
- 98. Предполагаемая схема эволюции АВС-системы у растений
- 99. Детерминация пола у растений Определение (детерминация) пола у растений - формирование признаков пола у клеток, органов
- 100. Аллосомы или sex-хромосомы - хромосомы отличающиеся по морфологии и количеству в мужской и женской особи и
- 101. Хромосомное определение пола у растений Большинство видов растений, у которых определение пола определяется хромосомами, могут быть
- 102. Факторы, опеделяющие фенотипическое определение пола
- 106. Скачать презентацию
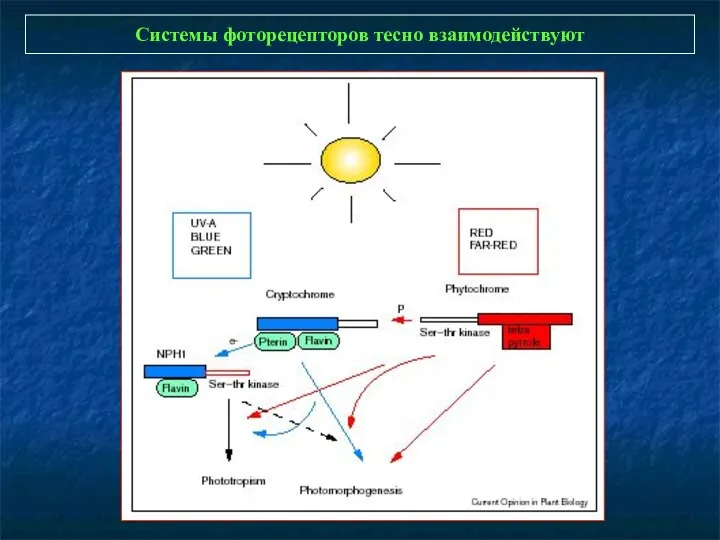
Системы фоторецепторов тесно взаимодействуют
Системы фоторецепторов тесно взаимодействуют
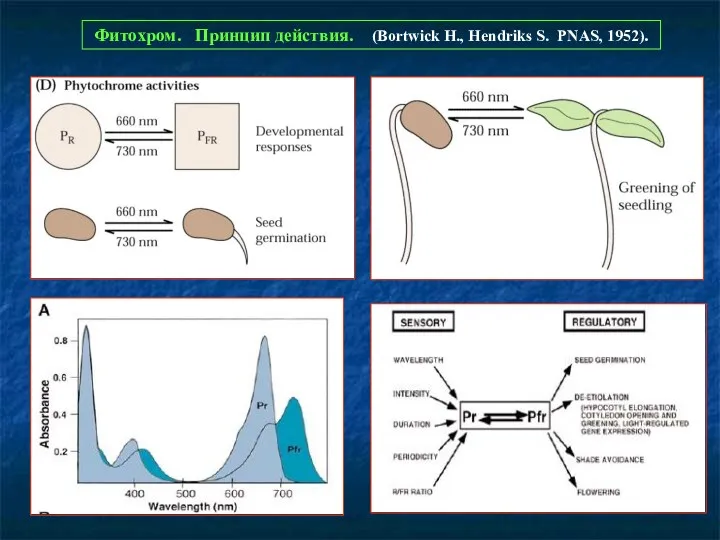
Фитохром. Принцип действия. (Bortwick H., Hendriks S. PNAS, 1952).
Фитохром. Принцип действия. (Bortwick H., Hendriks S. PNAS, 1952).

Фотообращаемые эффекты фитохромов.
Покрытосеменные
салат-латук семена Стимуляция прорастания
овес этиолированные Стимуляция деэтиоляции
проростки
горчица
Фотообращаемые эффекты фитохромов.
Покрытосеменные
салат-латук семена Стимуляция прорастания
овес этиолированные Стимуляция деэтиоляции
проростки
горчица
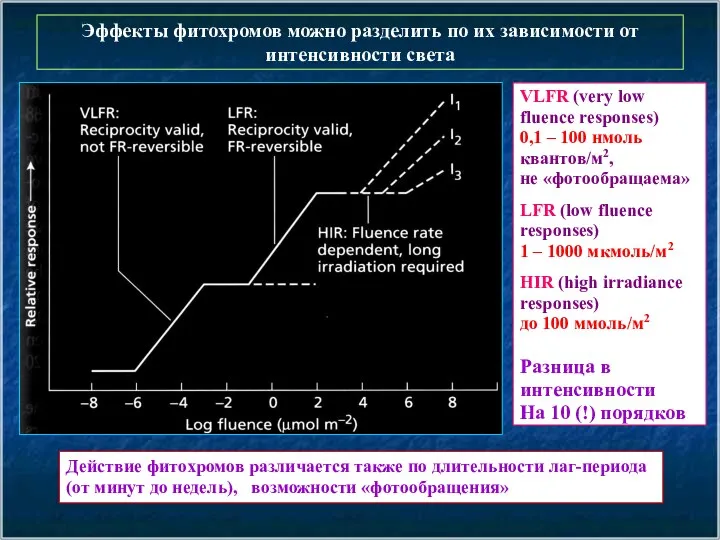
Эффекты фитохромов можно разделить по их зависимости от интенсивности света
VLFR (very
Эффекты фитохромов можно разделить по их зависимости от интенсивности света
VLFR (very
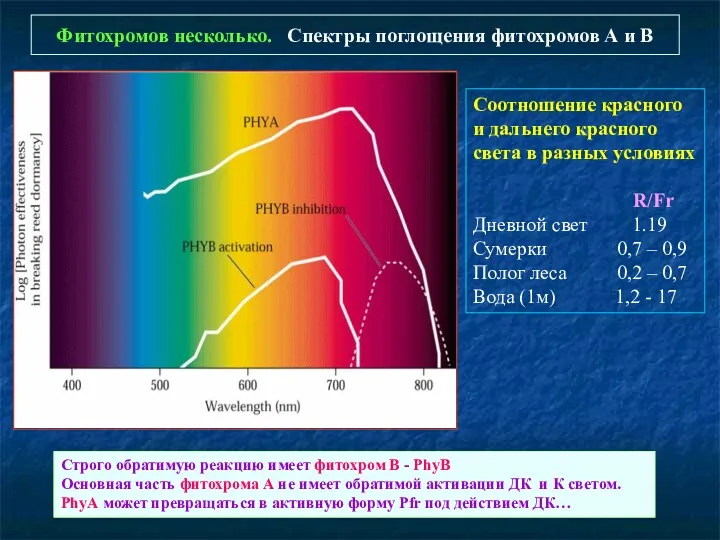
Фитохромов несколько. Спектры поглощения фитохромов А и В
Соотношение красного
и дальнего
Фитохромов несколько. Спектры поглощения фитохромов А и В
Соотношение красного
и дальнего
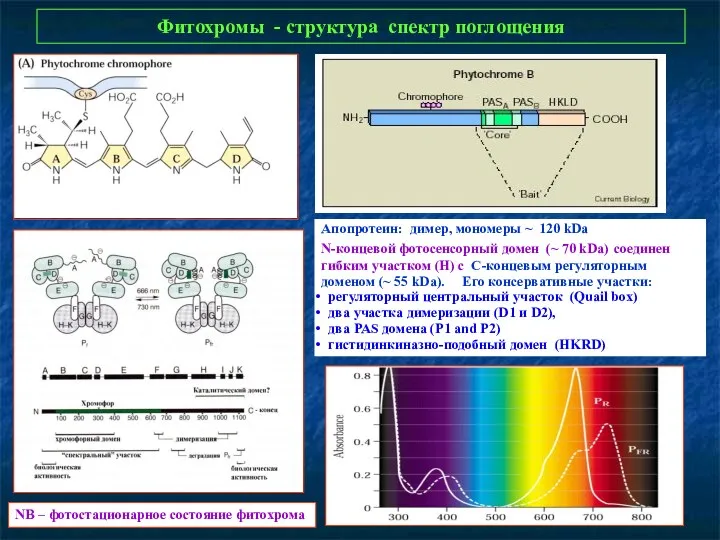
Фитохромы - структура спектр поглощения
NB – фотостационарное состояние фитохрома
Апопротеин: димер, мономеры
Фитохромы - структура спектр поглощения
NB – фотостационарное состояние фитохрома
Апопротеин: димер, мономеры
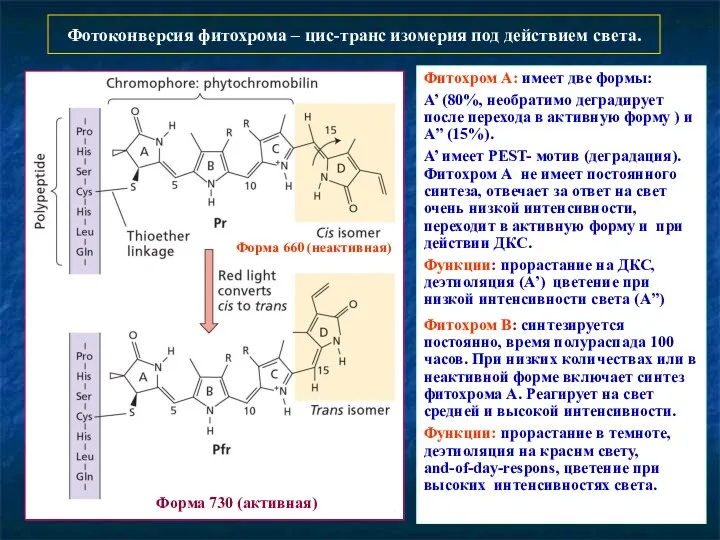
Фотоконверсия фитохрома – цис-транс изомерия под действием света.
Форма 660 (неактивная)
Фотоконверсия фитохрома – цис-транс изомерия под действием света.
Форма 660 (неактивная)
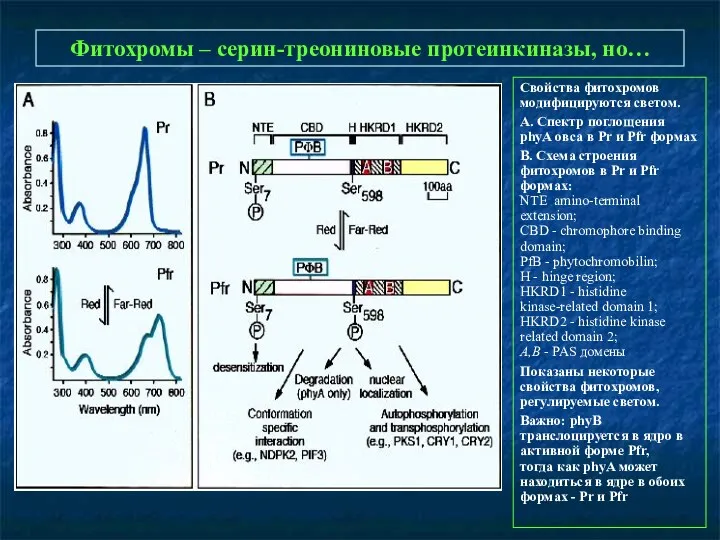
Фитохромы – серин-треониновые протеинкиназы, но…
Свойства фитохромов модифицируются светом.
А. Спектр поглощения phyA
Фитохромы – серин-треониновые протеинкиназы, но…
Свойства фитохромов модифицируются светом.
А. Спектр поглощения phyA
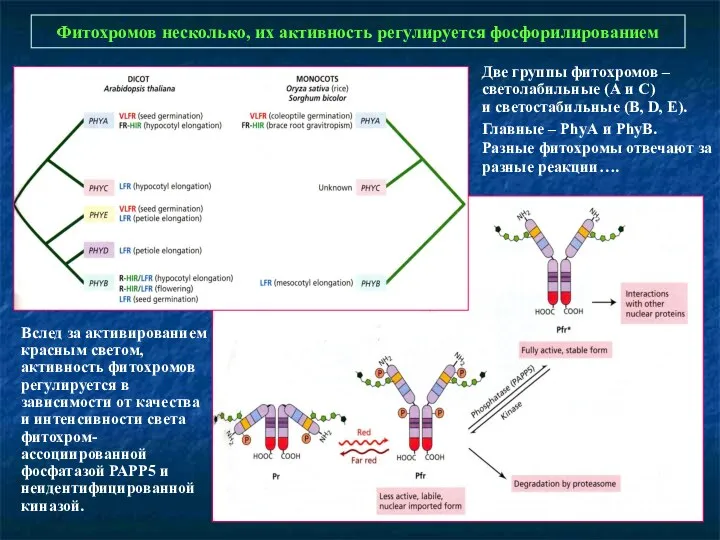
Фитохромов несколько, их активность регулируется фосфорилированием
Вслед за активированием красным светом, активность
Фитохромов несколько, их активность регулируется фосфорилированием
Вслед за активированием красным светом, активность
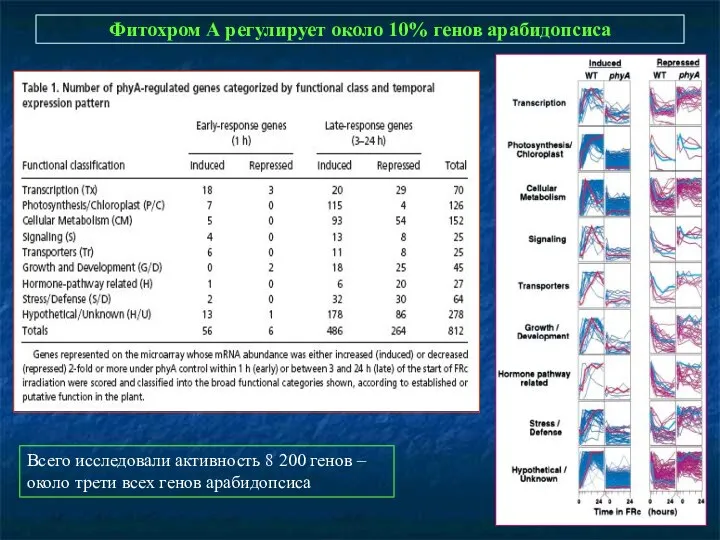
Фитохром А регулирует около 10% генов арабидопсиса
Всего исследовали активность 8
Фитохром А регулирует около 10% генов арабидопсиса
Всего исследовали активность 8
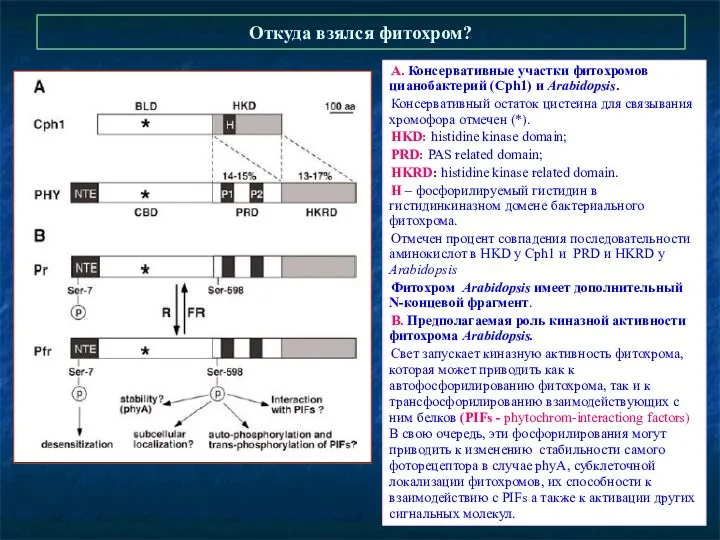
Откуда взялся фитохром?
А. Консервативные участки фитохромов цианобактерий (Cph1) и Arabidopsis.
Консервативный
Откуда взялся фитохром?
А. Консервативные участки фитохромов цианобактерий (Cph1) и Arabidopsis.
Консервативный
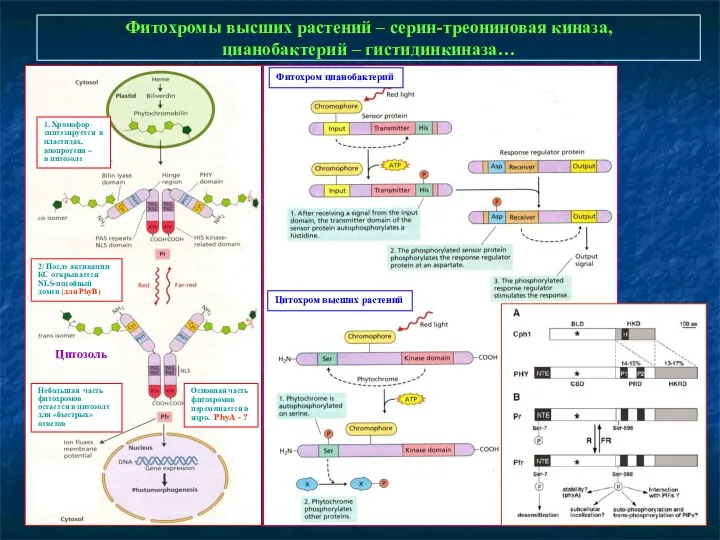
Фитохромы высших растений – серин-треониновая киназа,
цианобактерий – гистидинкиназа…
1. Хромофор синтезируется
Фитохромы высших растений – серин-треониновая киназа,
цианобактерий – гистидинкиназа…
1. Хромофор синтезируется
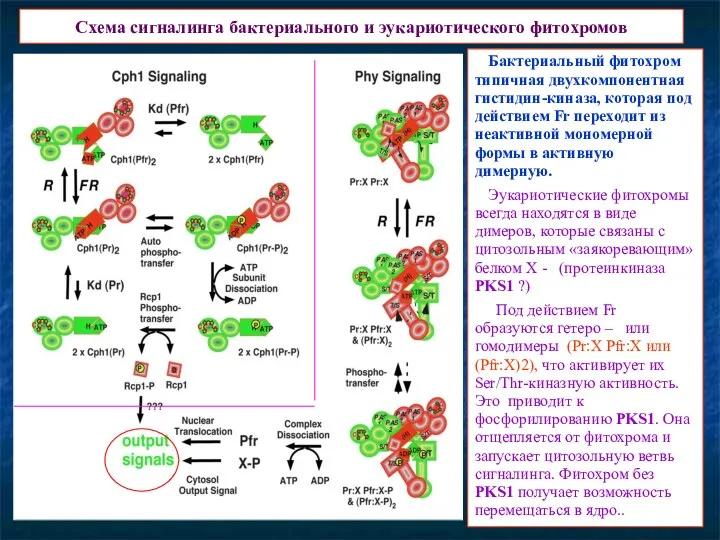
Схема сигналинга бактериального и эукариотического фитохромов
Бактериальный фитохром типичная двухкомпонентная гистидин-киназа, которая
Схема сигналинга бактериального и эукариотического фитохромов
Бактериальный фитохром типичная двухкомпонентная гистидин-киназа, которая

Сигналинг фитохромов. Основные «игроки» и общие положения
Два типа ответов – быстрые
Сигналинг фитохромов. Основные «игроки» и общие положения
Два типа ответов – быстрые
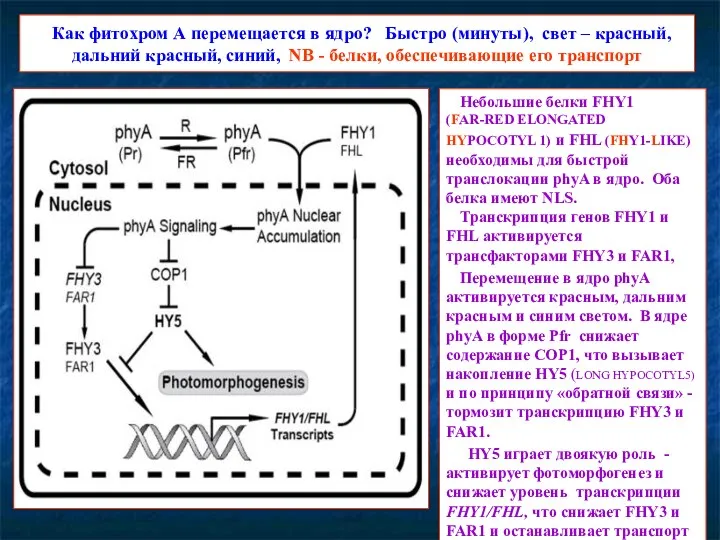
Как фитохром А перемещается в ядро? Быстро (минуты), свет –
Как фитохром А перемещается в ядро? Быстро (минуты), свет –
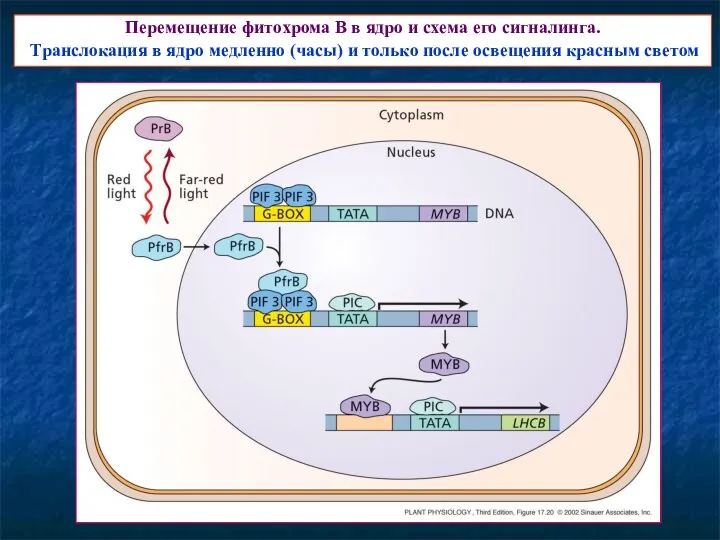
Перемещение фитохрома В в ядро и схема его сигналинга.
Транслокация
Перемещение фитохрома В в ядро и схема его сигналинга.
Транслокация
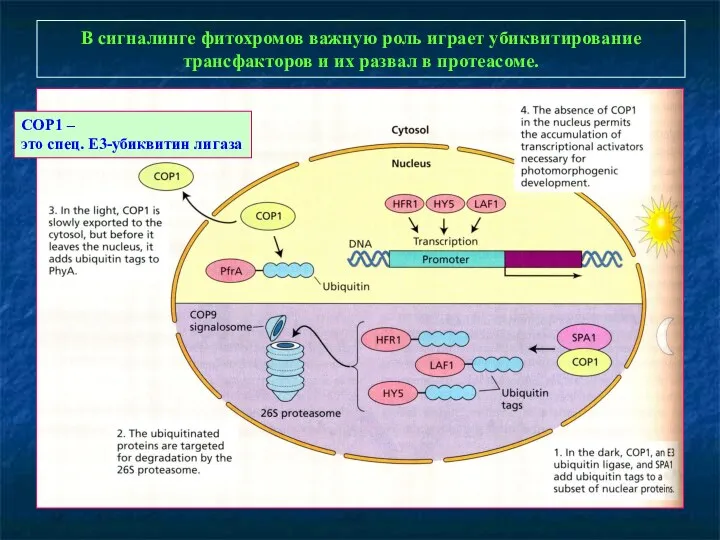
В сигналинге фитохромов важную роль играет убиквитирование трансфакторов и их развал
В сигналинге фитохромов важную роль играет убиквитирование трансфакторов и их развал
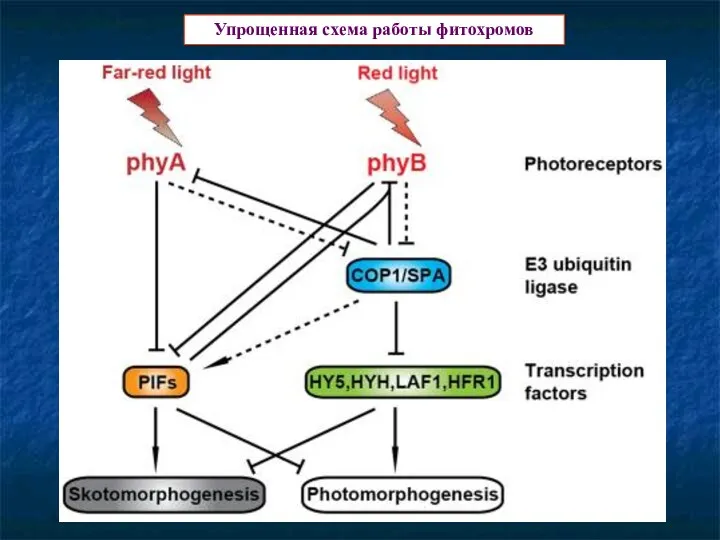
Упрощенная схема работы фитохромов
Упрощенная схема работы фитохромов
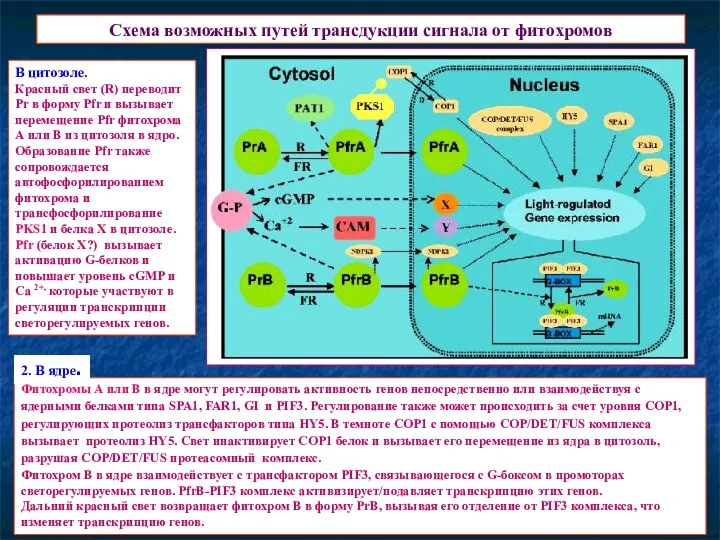
Схема возможных путей трансдукции сигнала от фитохромов
Фитохромы А или В в
Схема возможных путей трансдукции сигнала от фитохромов
Фитохромы А или В в
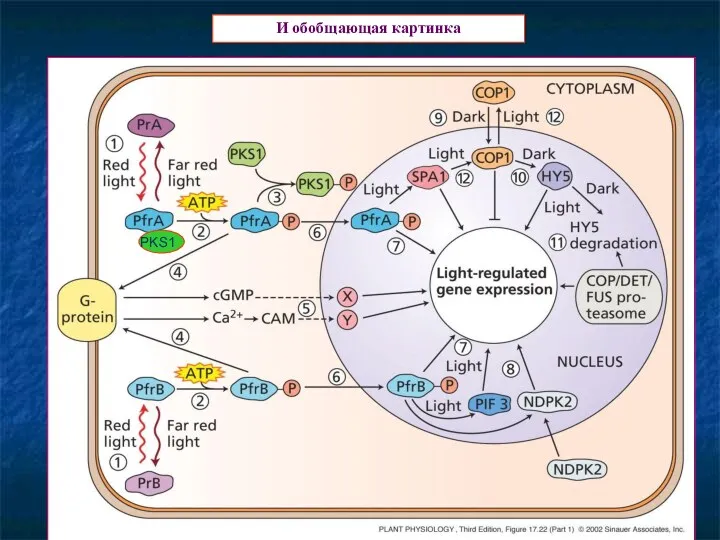
И обобщающая картинка
PKS1
И обобщающая картинка
PKS1

Рецепторы синего света отвечают за многое…
Реакция фототропизма
Движение хлоропластов
Циркадные ритмы:
открытие устьиц
«сонные» движения
Рецепторы синего света отвечают за многое…
Реакция фототропизма
Движение хлоропластов
Циркадные ритмы:
открытие устьиц
«сонные» движения
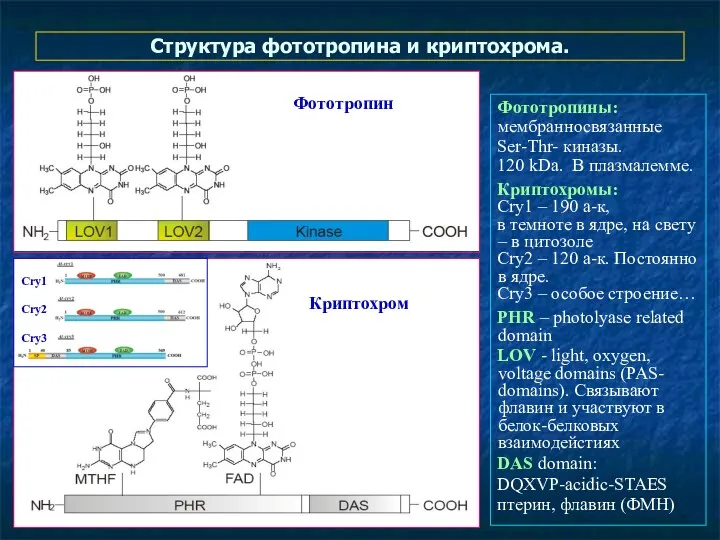
Структура фототропина и криптохрома.
Фототропины: мембранносвязанные
Ser-Thr- киназы.
120 kDa. В плазмалемме.
Криптохромы:
Структура фототропина и криптохрома.
Фототропины: мембранносвязанные
Ser-Thr- киназы.
120 kDa. В плазмалемме.
Криптохромы:
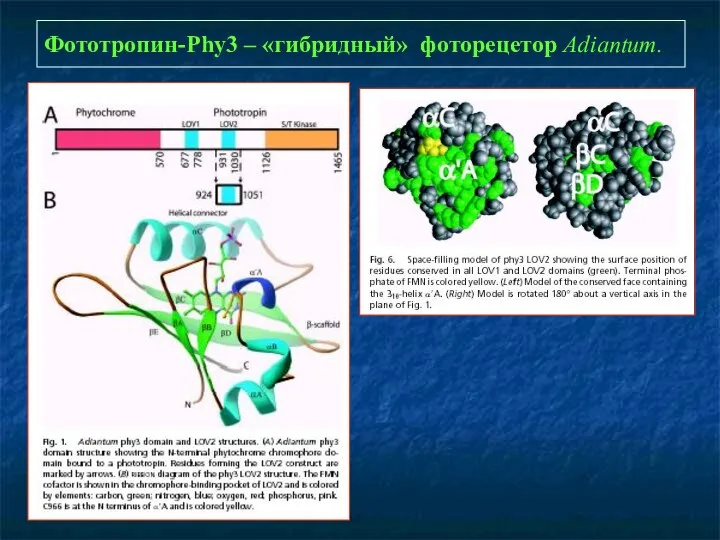
Фототропин-Phy3 – «гибридный» фоторецетор Adiantum.
Фототропин-Phy3 – «гибридный» фоторецетор Adiantum.

Схема строения фоторецепторов и их взаимодействие
Схема строения фоторецепторов и их взаимодействие

Фототропины участвуют в регуляции разных фотодвижений
Изгибание колеоптиля
Движение хлоропластов
Открывание устьиц
У Arabidopsis работают
Фототропины участвуют в регуляции разных фотодвижений
Изгибание колеоптиля
Движение хлоропластов
Открывание устьиц
У Arabidopsis работают
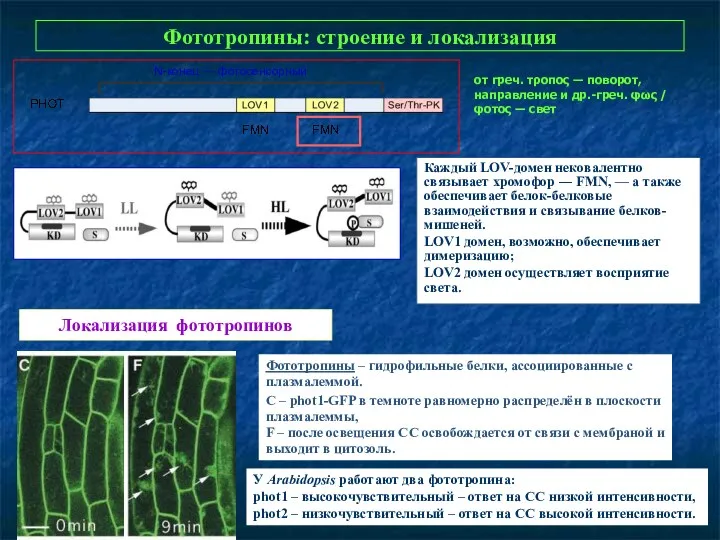
Фототропины: строение и локализация
Каждый LOV-домен нековалентно связывает хромофор ― FMN, —
Фототропины: строение и локализация
Каждый LOV-домен нековалентно связывает хромофор ― FMN, —
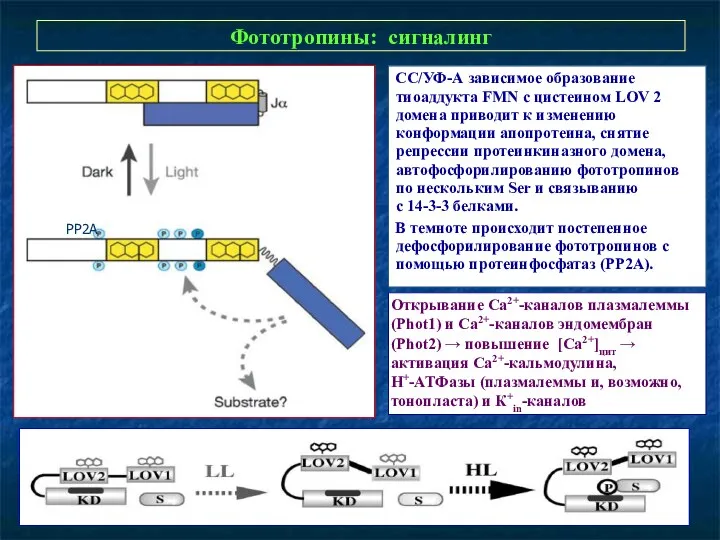
Фототропины: сигналинг
СС/УФ-А зависимое образование тиоаддукта FMN с цистеином LOV 2 домена
Фототропины: сигналинг
СС/УФ-А зависимое образование тиоаддукта FMN с цистеином LOV 2 домена
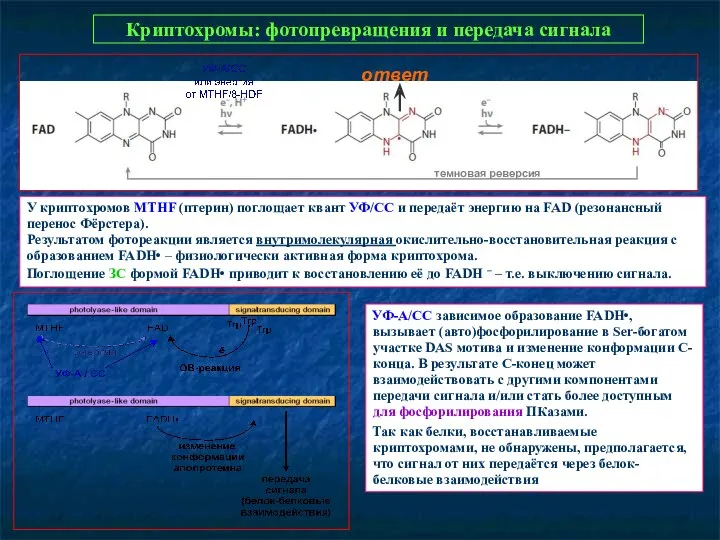
Криптохромы: фотопревращения и передача сигнала
У криптохромов MTHF (птерин) поглощает квант УФ/СС
Криптохромы: фотопревращения и передача сигнала
У криптохромов MTHF (птерин) поглощает квант УФ/СС
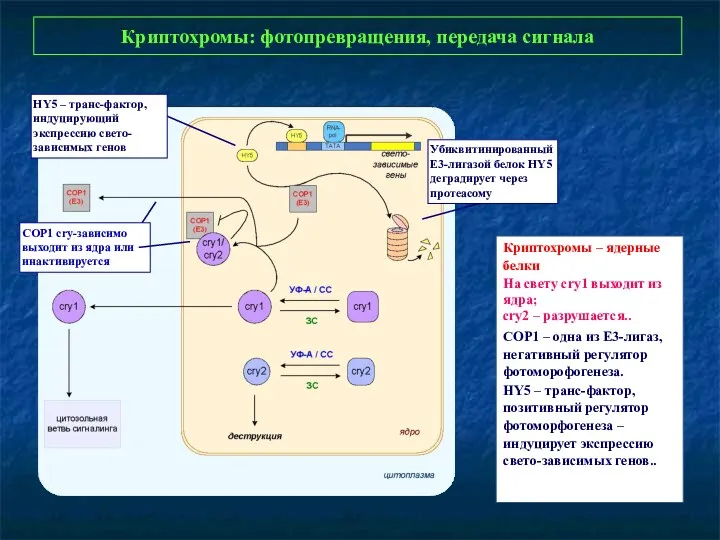
Криптохромы: фотопревращения, передача сигнала
Криптохромы – ядерные белки
На свету cry1 выходит
Криптохромы: фотопревращения, передача сигнала
Криптохромы – ядерные белки
На свету cry1 выходит
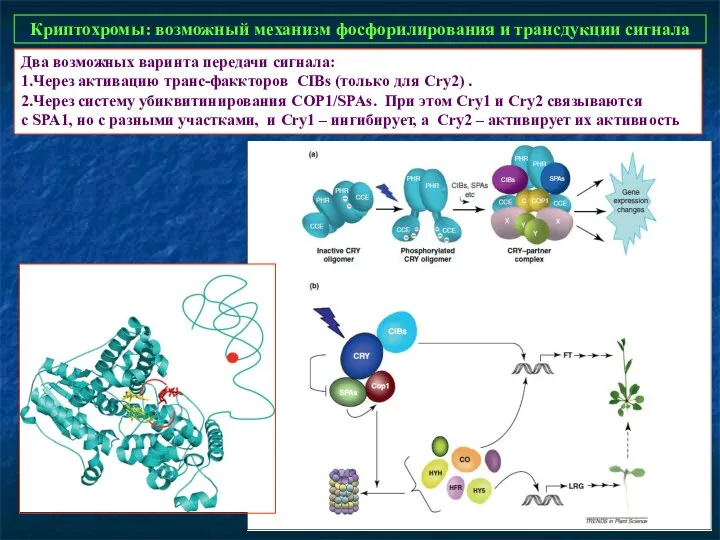
Криптохромы: возможный механизм фосфорилирования и трансдукции сигнала
Два возможных варинта передачи сигнала:
Криптохромы: возможный механизм фосфорилирования и трансдукции сигнала
Два возможных варинта передачи сигнала:
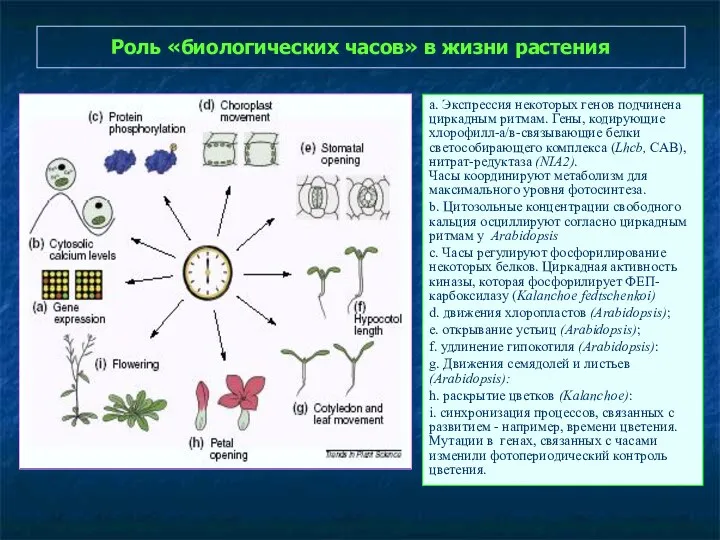
Роль «биологических часов» в жизни растения
а. Экспрессия некоторых генов подчинена циркадным
Роль «биологических часов» в жизни растения
а. Экспрессия некоторых генов подчинена циркадным
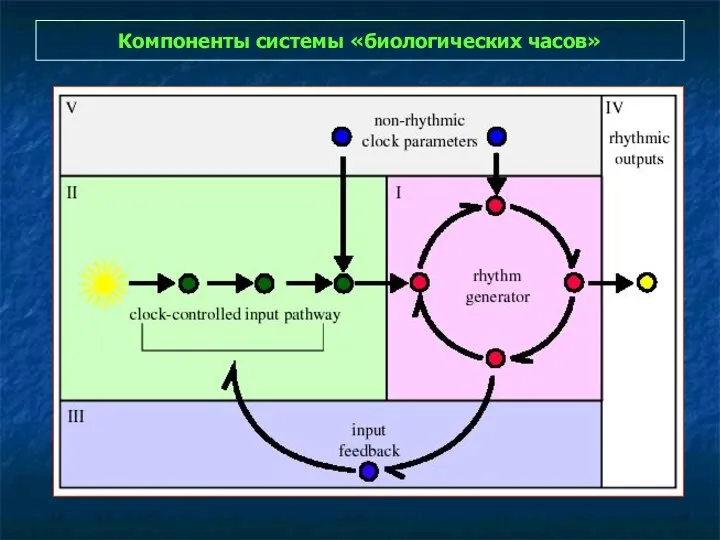
Компоненты системы «биологических часов»
Компоненты системы «биологических часов»
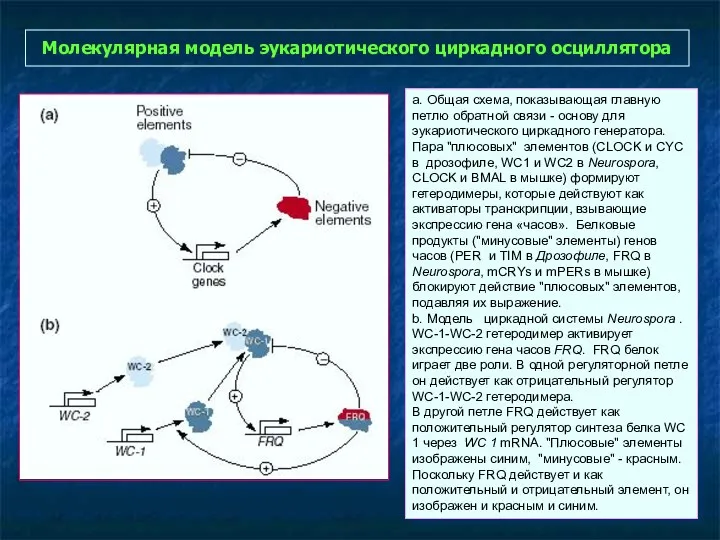
Молекулярная модель эукариотического циркадного осциллятора
a. Общая схема, показывающая главную петлю обратной
Молекулярная модель эукариотического циркадного осциллятора
a. Общая схема, показывающая главную петлю обратной
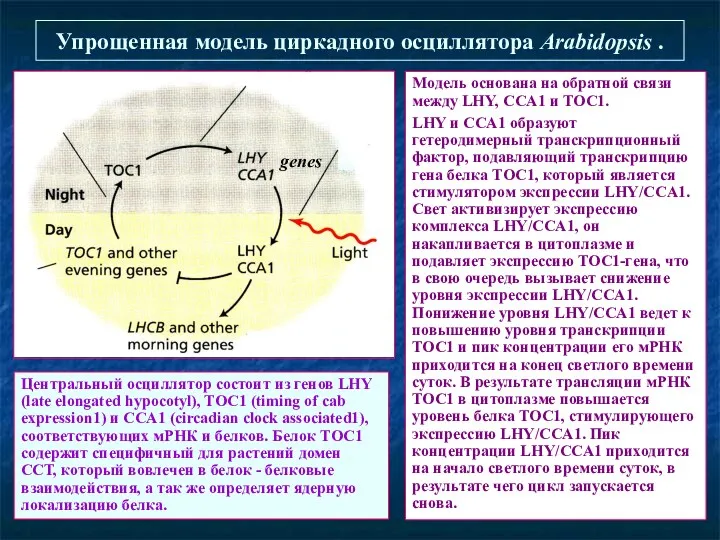
Упрощенная модель циркадного осциллятора Arabidopsis .
Модель основана на обратной связи между
Упрощенная модель циркадного осциллятора Arabidopsis .
Модель основана на обратной связи между
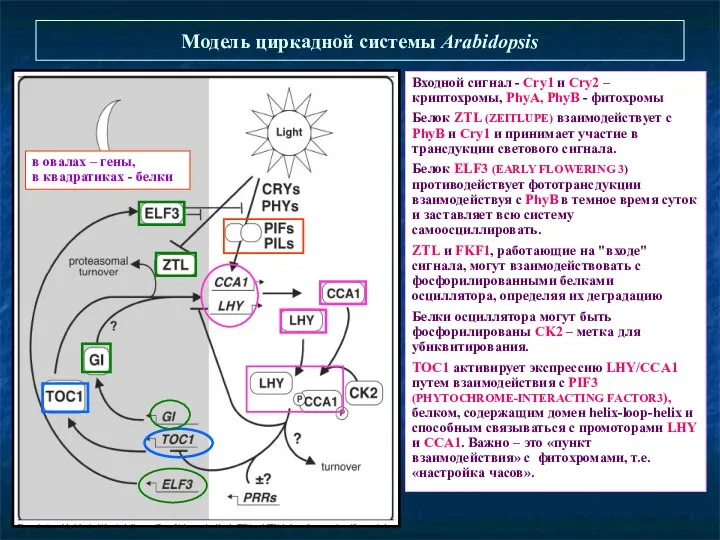
Модель циркадной системы Arabidopsis
Входной сигнал - Сry1 и Сry2 –
Модель циркадной системы Arabidopsis
Входной сигнал - Сry1 и Сry2 –
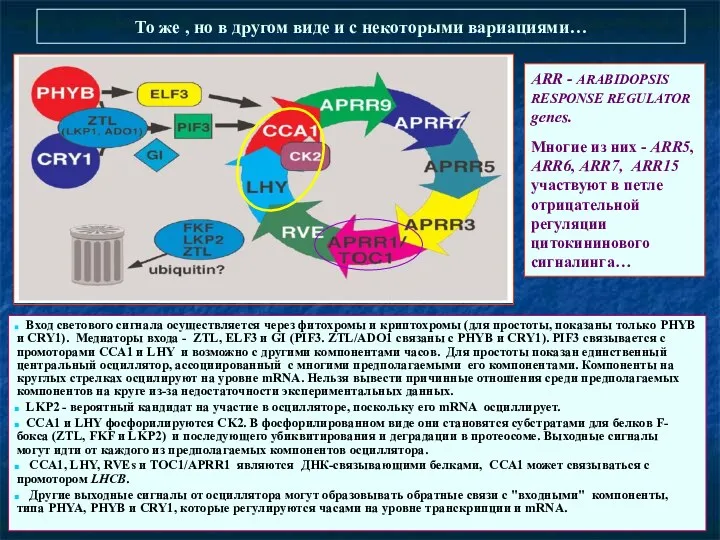
То же , но в другом виде и с некоторыми вариациями…
То же , но в другом виде и с некоторыми вариациями…

Стратегии развития животных и растений различны.
У животных: развитие – серия сложных
Стратегии развития животных и растений различны.
У животных: развитие – серия сложных
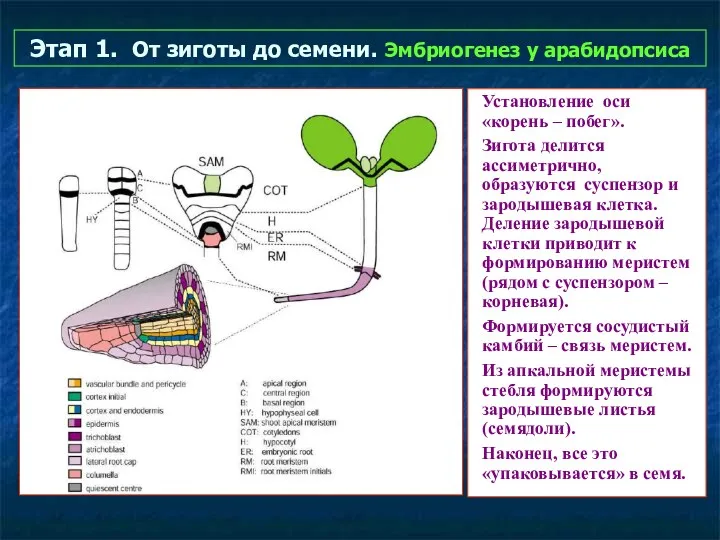
Этап 1. От зиготы до семени. Эмбриогенез у арабидопсиса
Установление оси «корень
Этап 1. От зиготы до семени. Эмбриогенез у арабидопсиса
Установление оси «корень

Особенности и механизмы эмбрионального развития растений
Формирование зародыша у растений происходит
Особенности и механизмы эмбрионального развития растений
Формирование зародыша у растений происходит
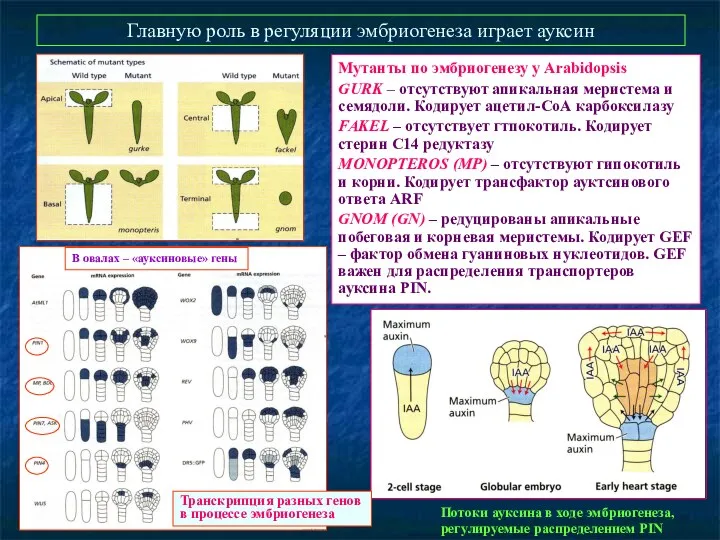
Главную роль в регуляции эмбриогенеза играет ауксин
Мутанты по эмбриогенезу у Arabidopsis
GURK
Главную роль в регуляции эмбриогенеза играет ауксин
Мутанты по эмбриогенезу у Arabidopsis
GURK
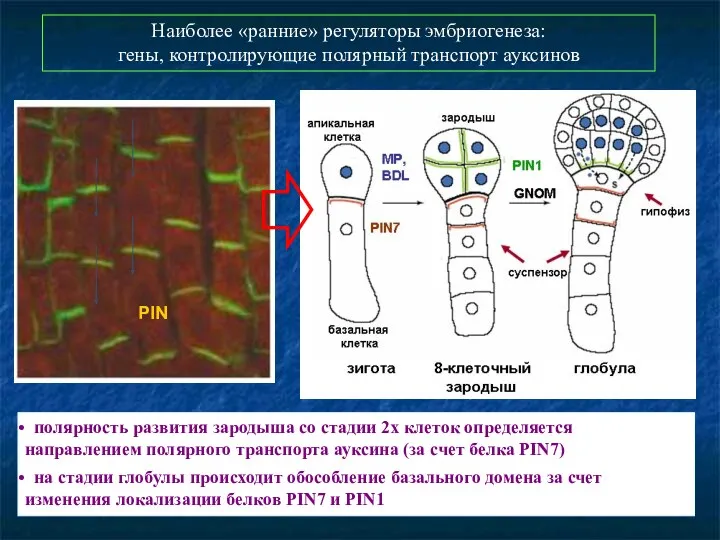
Наиболее «ранние» регуляторы эмбриогенеза:
гены, контролирующие полярный транспорт ауксинов
полярность развития зародыша
Наиболее «ранние» регуляторы эмбриогенеза:
гены, контролирующие полярный транспорт ауксинов
полярность развития зародыша
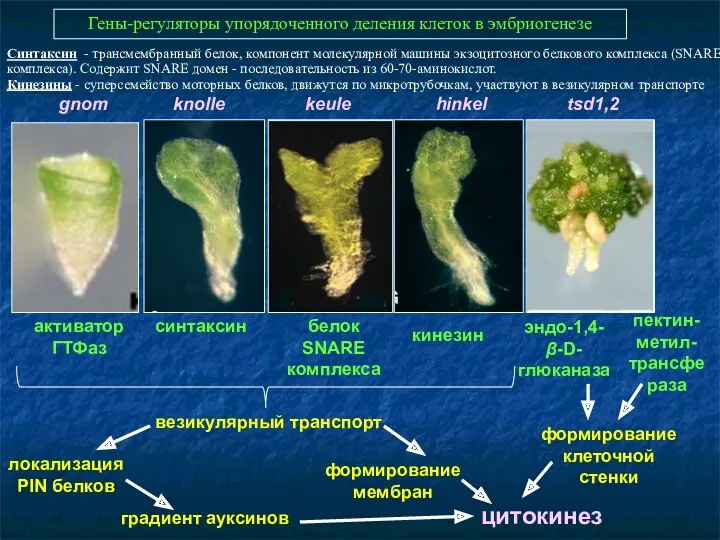
Гены-регуляторы упорядоченного деления клеток в эмбриогенезе
gnom knolle keule hinkel tsd1,2
эндо-1,4-β-D-глюканаза
синтаксин
кинезин
белок
Гены-регуляторы упорядоченного деления клеток в эмбриогенезе
gnom knolle keule hinkel tsd1,2
эндо-1,4-β-D-глюканаза
синтаксин
кинезин
белок
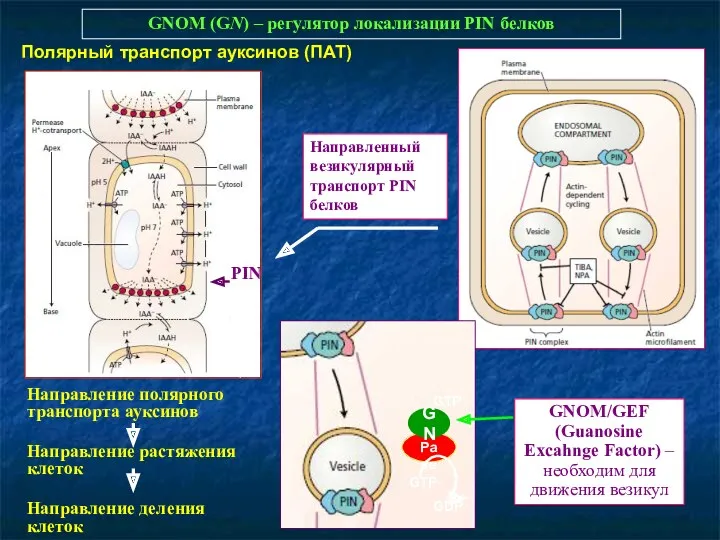
GNOM (GN) – регулятор локализации PIN белков
PIN
Направление полярного
транспорта ауксинов
Направление растяжения
GNOM (GN) – регулятор локализации PIN белков
PIN
Направление полярного
транспорта ауксинов
Направление растяжения
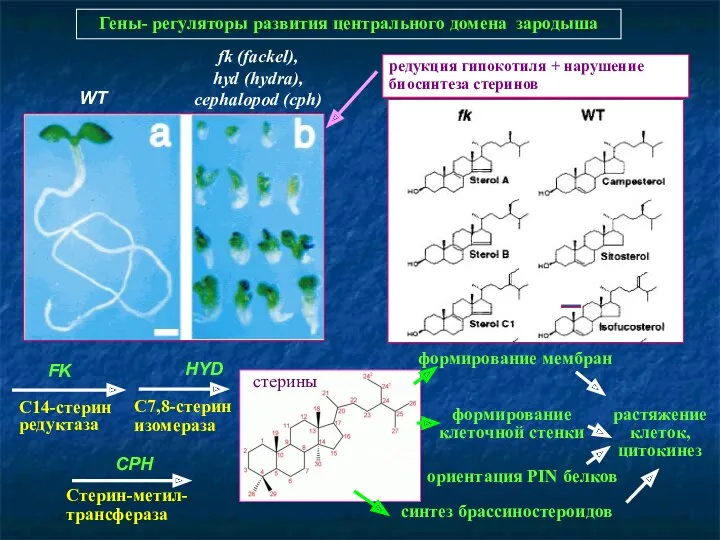
Гены- регуляторы развития центрального домена зародыша
WT
fk (fackel),
hyd (hydra), cephalopod (cph)
редукция
Гены- регуляторы развития центрального домена зародыша
WT
fk (fackel),
hyd (hydra), cephalopod (cph)
редукция
Соматический эмбриогенез – модель для изучения механизмов эмбриогенеза in vivo
Соматический эмбриогенез – модель для изучения механизмов эмбриогенеза in vivo
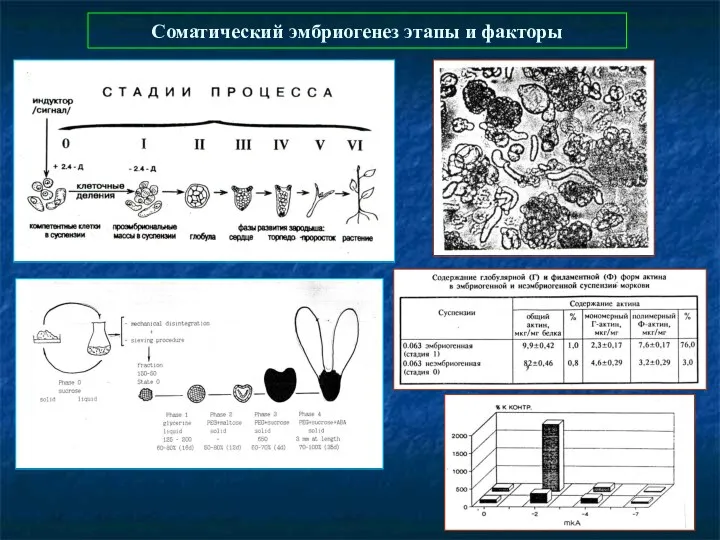
Соматический эмбриогенез этапы и факторы
Соматический эмбриогенез этапы и факторы
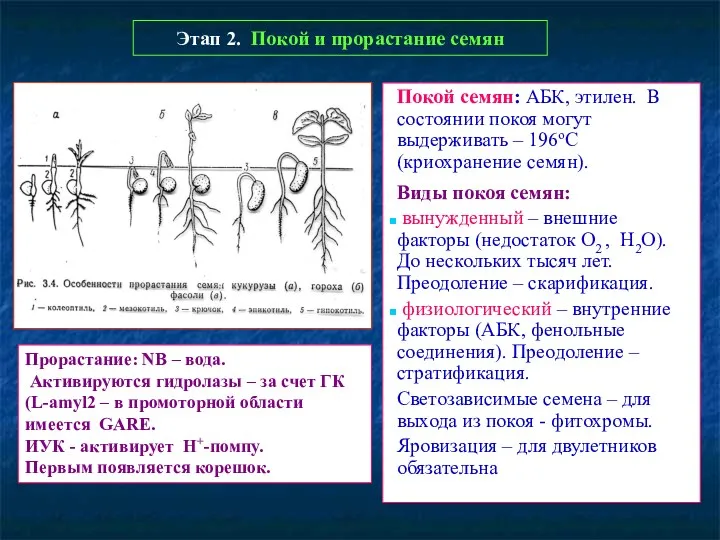
Этап 2. Покой и прорастание семян
Покой семян: АБК, этилен. В состоянии
Этап 2. Покой и прорастание семян
Покой семян: АБК, этилен. В состоянии
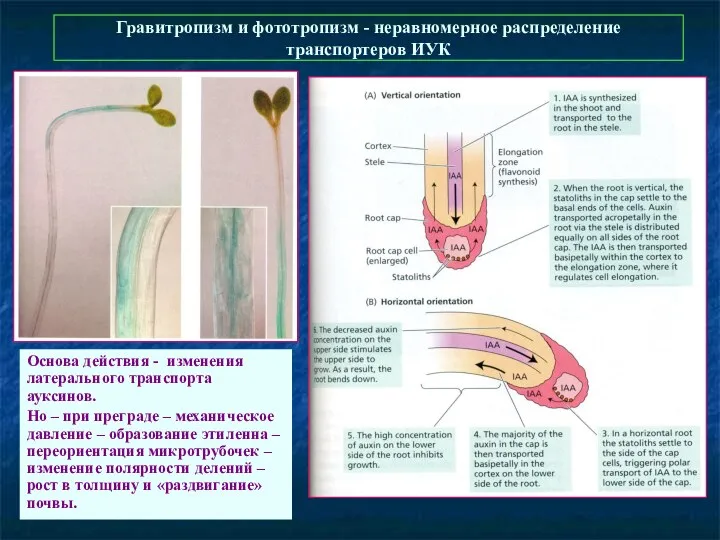
Гравитропизм и фототропизм - неравномерное распределение транспортеров ИУК
Основа действия - изменения
Гравитропизм и фототропизм - неравномерное распределение транспортеров ИУК
Основа действия - изменения
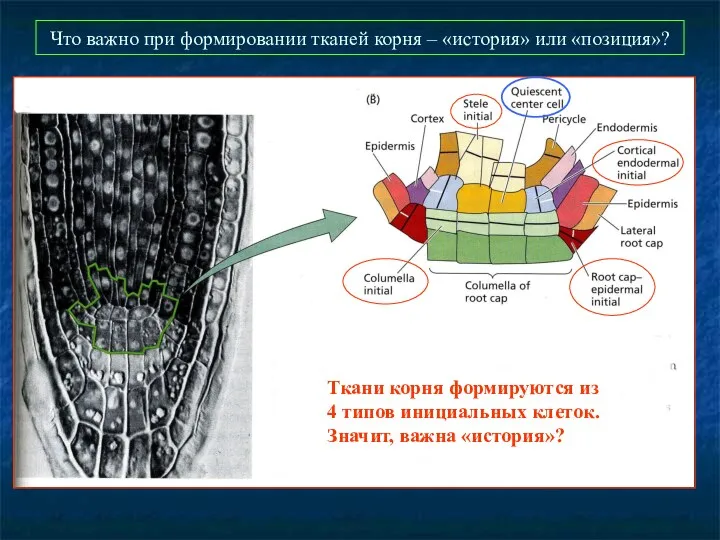
Что важно при формировании тканей корня – «история» или «позиция»?
Ткани корня
Что важно при формировании тканей корня – «история» или «позиция»?
Ткани корня
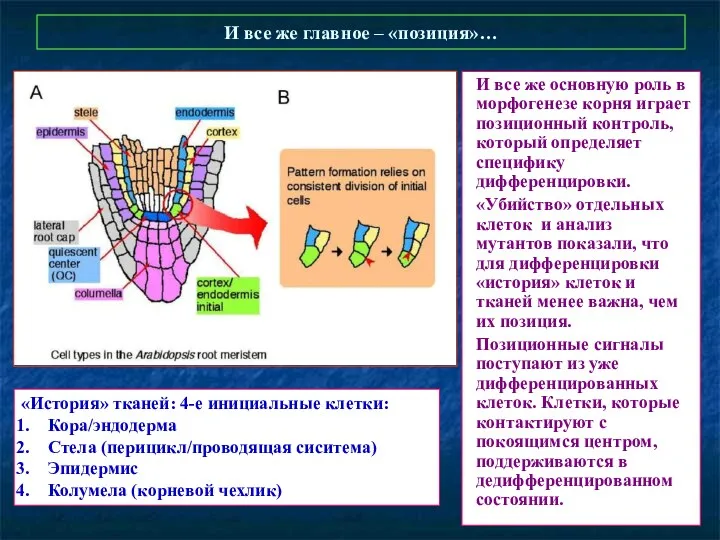
И все же главное – «позиция»…
И все же основную роль в
И все же главное – «позиция»…
И все же основную роль в
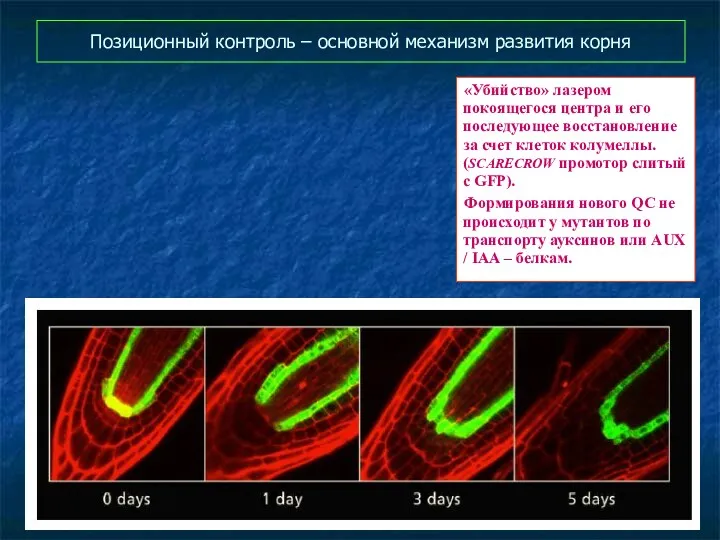
Позиционный контроль – основной механизм развития корня
«Убийство» лазером покоящегося центра и
Позиционный контроль – основной механизм развития корня
«Убийство» лазером покоящегося центра и
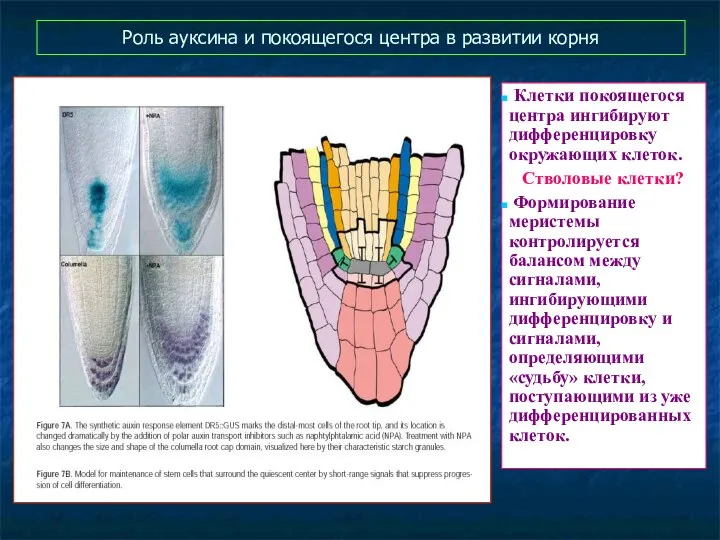
Роль ауксина и покоящегося центра в развитии корня
Клетки покоящегося центра
Роль ауксина и покоящегося центра в развитии корня
Клетки покоящегося центра
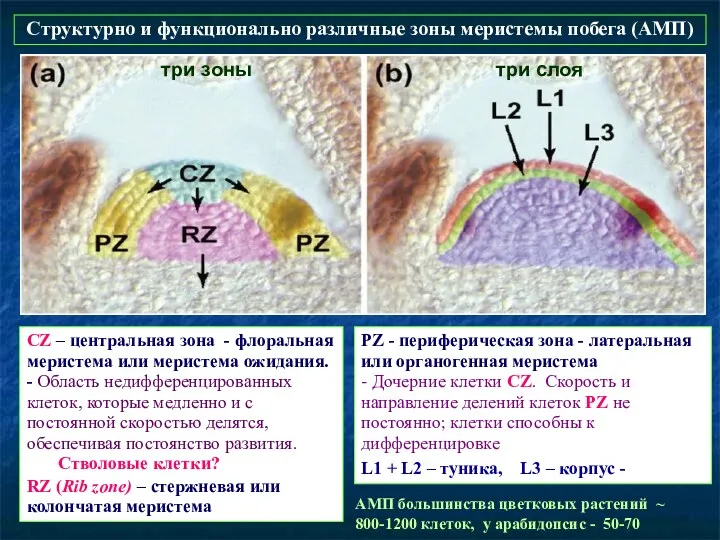
PZ - периферическая зона - латеральная или органогенная меристема
- Дочерние
PZ - периферическая зона - латеральная или органогенная меристема
- Дочерние
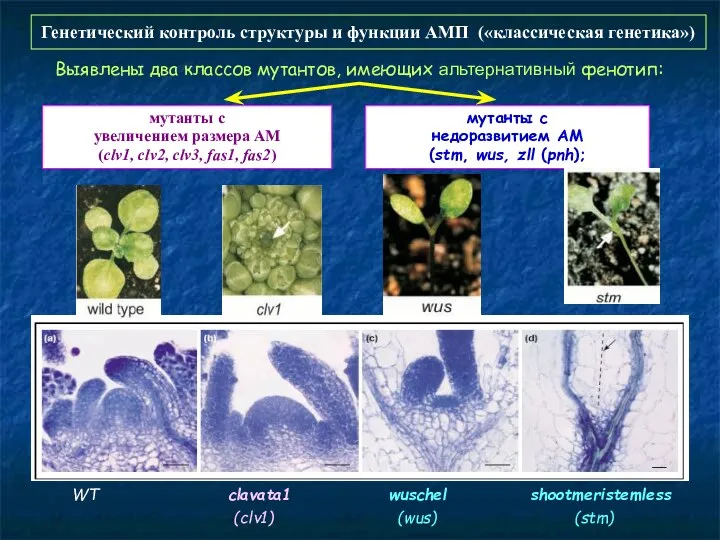
Генетический контроль структуры и функции АМП («классическая генетика»)
Выявлены два классов мутантов,
Генетический контроль структуры и функции АМП («классическая генетика»)
Выявлены два классов мутантов,
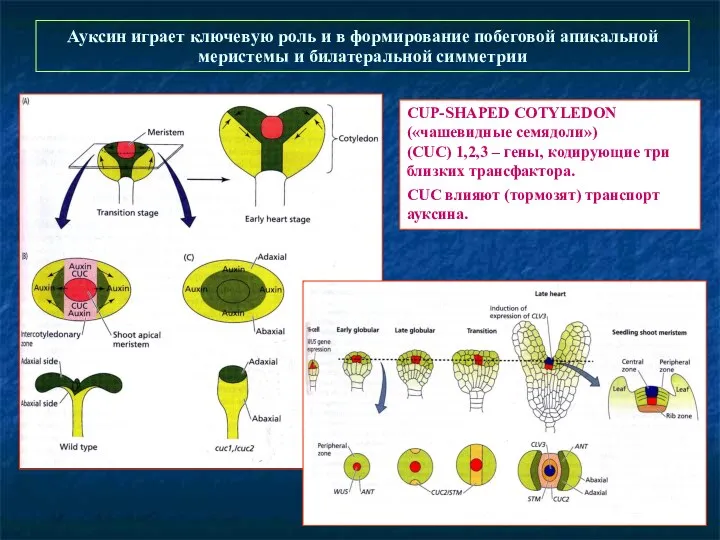
Ауксин играет ключевую роль и в формирование побеговой апикальной меристемы и
Ауксин играет ключевую роль и в формирование побеговой апикальной меристемы и
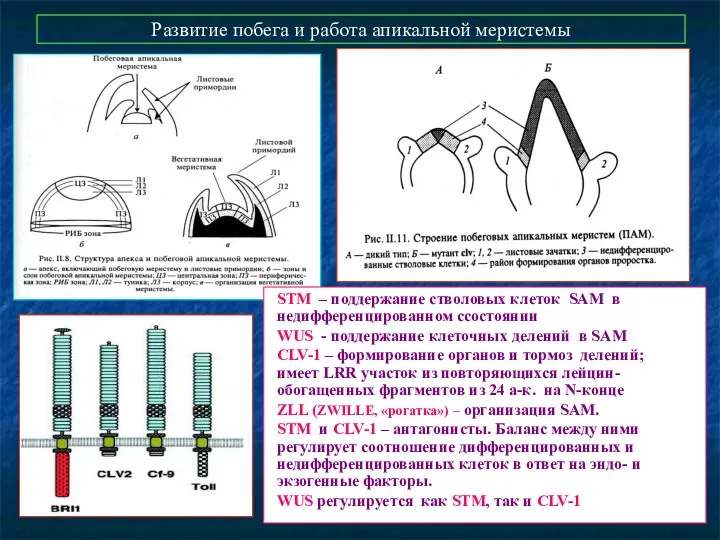
Развитие побега и работа апикальной меристемы
STM – поддержание стволовых клеток
Развитие побега и работа апикальной меристемы
STM – поддержание стволовых клеток
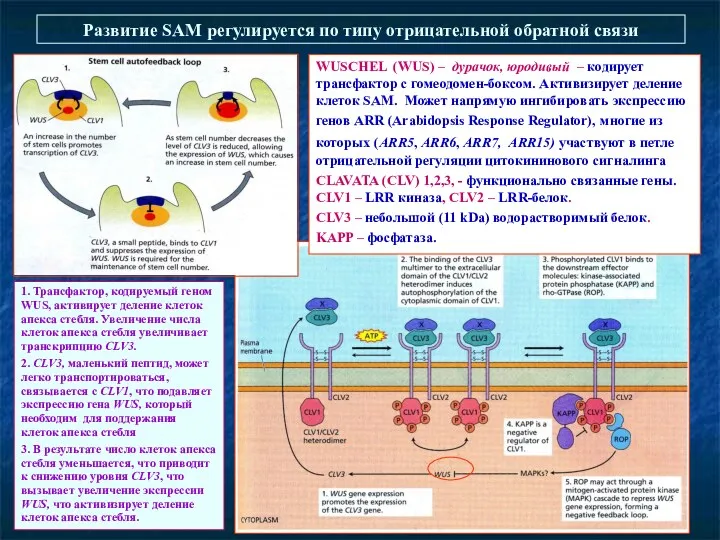
Развитие SAM регулируется по типу отрицательной обратной связи
WUSCHEL (WUS) – дурачок,
Развитие SAM регулируется по типу отрицательной обратной связи
WUSCHEL (WUS) – дурачок,

Система WOX - CLE
Wang, Fiers 2010. CLE peptide signaling during plant
Система WOX - CLE
Wang, Fiers 2010. CLE peptide signaling during plant
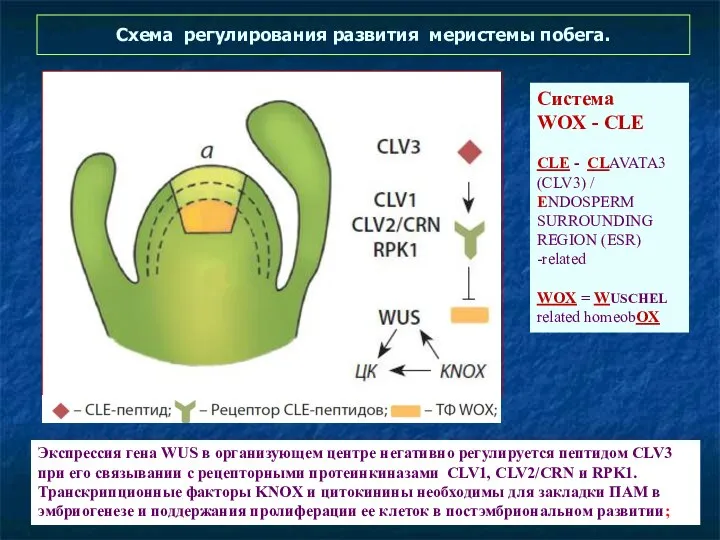
Схема регулирования развития меристемы побега.
Экспрессия гена WUS в организующем центре
Схема регулирования развития меристемы побега.
Экспрессия гена WUS в организующем центре

CLE - пептиды участвуют в определении судьбы клеток меристем
Wang, Fiers 2010.
CLE - пептиды участвуют в определении судьбы клеток меристем
Wang, Fiers 2010.
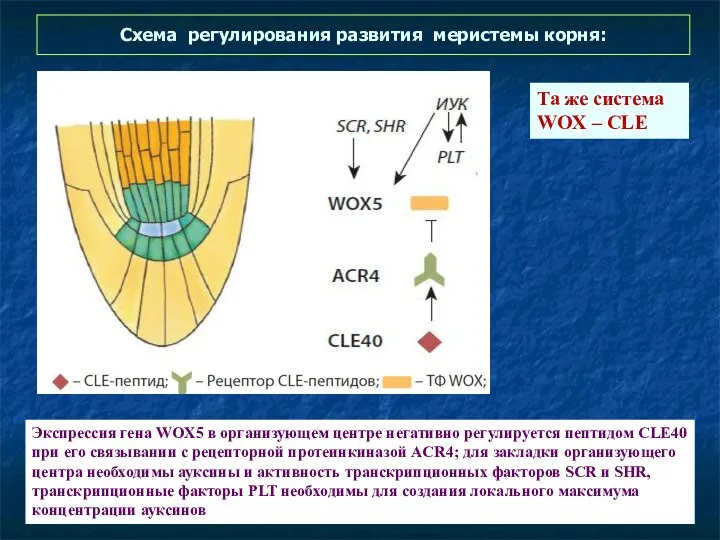
Схема регулирования развития меристемы корня:
Экспрессия гена WOX5 в организующем центре
Схема регулирования развития меристемы корня:
Экспрессия гена WOX5 в организующем центре
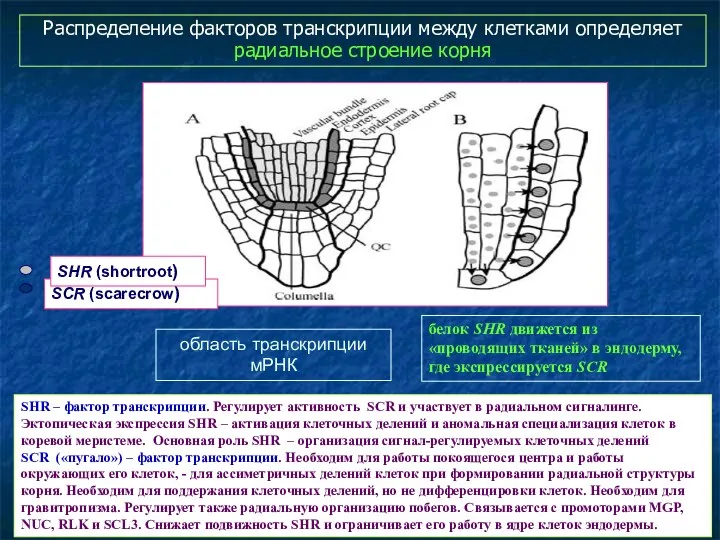
Распределение факторов транскрипции между клетками определяет радиальное строение корня
SHR – фактор
Распределение факторов транскрипции между клетками определяет радиальное строение корня
SHR – фактор
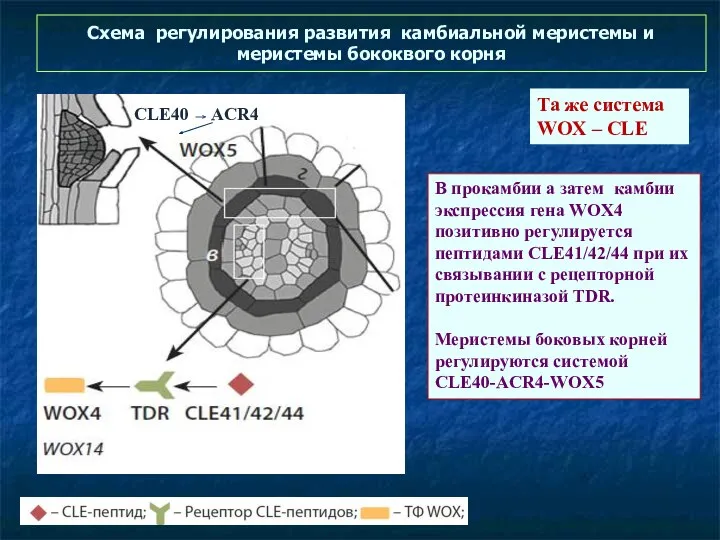
Схема регулирования развития камбиальной меристемы и меристемы бококвого корня
В прокамбии а
Схема регулирования развития камбиальной меристемы и меристемы бококвого корня
В прокамбии а
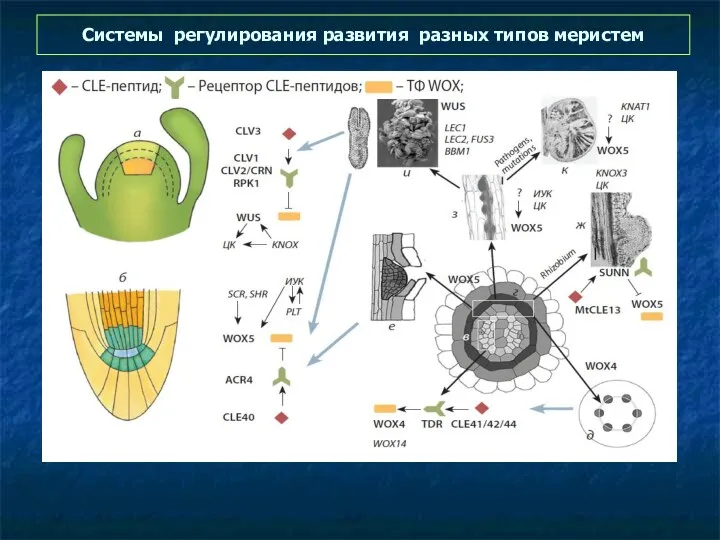
Системы регулирования развития разных типов меристем
Системы регулирования развития разных типов меристем
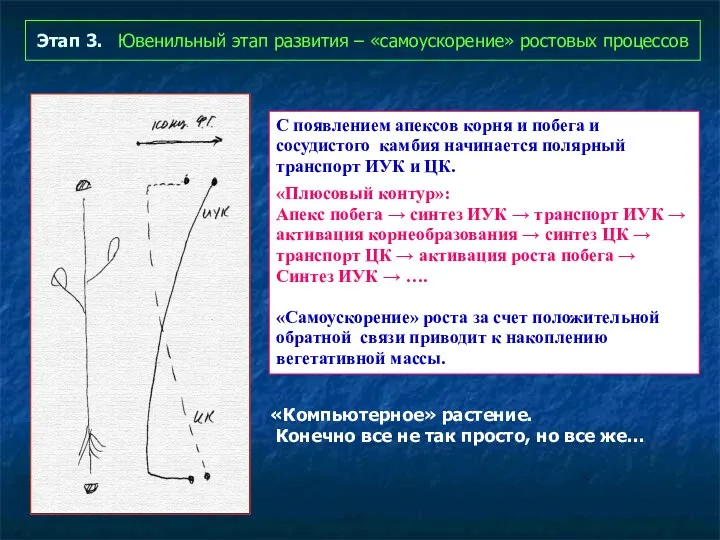
Этап 3. Ювенильный этап развития – «самоускорение» ростовых процессов
С появлением апексов
Этап 3. Ювенильный этап развития – «самоускорение» ростовых процессов
С появлением апексов
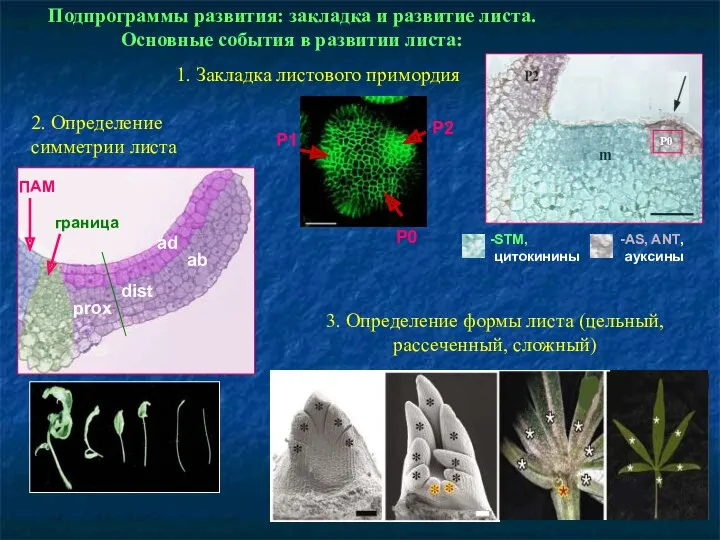
Подпрограммы развития: закладка и развитие листа.
Основные события в развитии листа:
1.
Подпрограммы развития: закладка и развитие листа.
Основные события в развитии листа:
1.
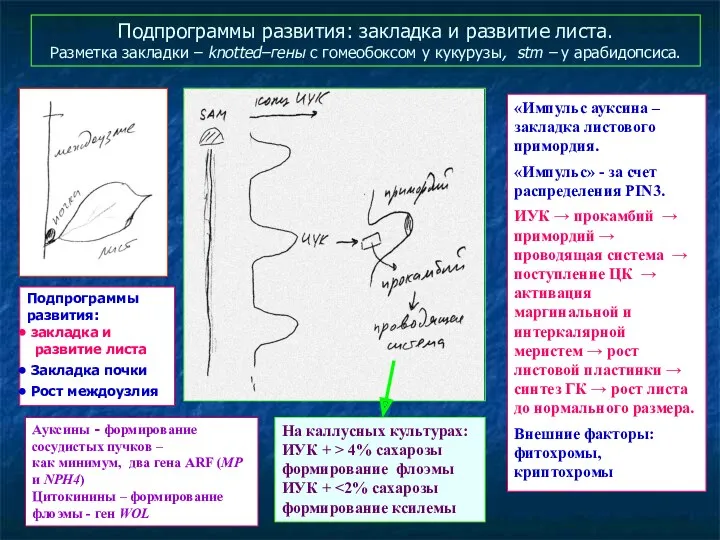
Подпрограммы развития: закладка и развитие листа.
Разметка закладки – knotted–гены с
Подпрограммы развития: закладка и развитие листа. Разметка закладки – knotted–гены с
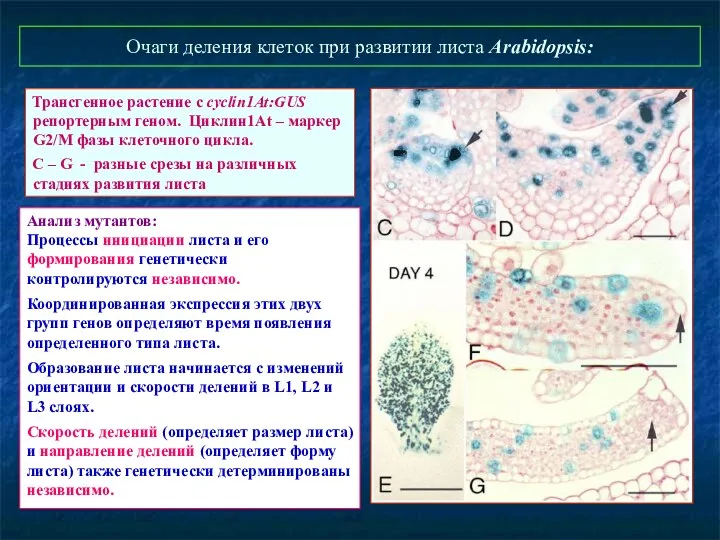
Очаги деления клеток при развитии листа Arabidopsis:
Трансгенное растение с cyclin1At:GUS репортерным
Очаги деления клеток при развитии листа Arabidopsis:
Трансгенное растение с cyclin1At:GUS репортерным
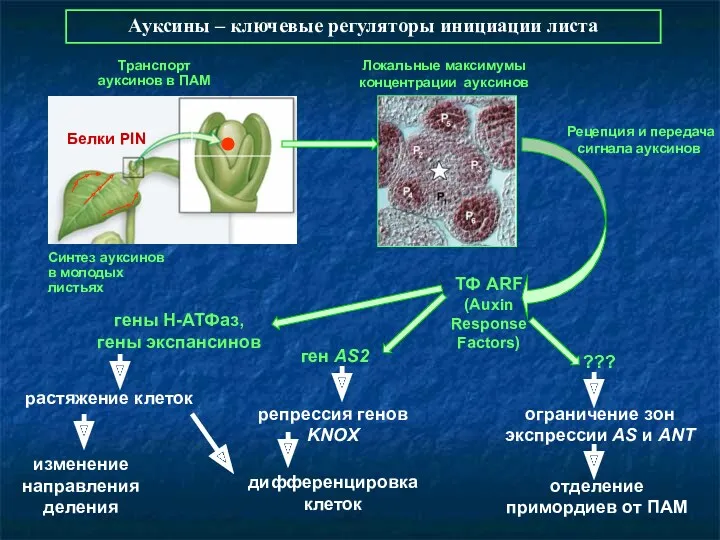
Ауксины – ключевые регуляторы инициации листа
Белки PIN
Синтез ауксинов в молодых листьях
Транспорт
Ауксины – ключевые регуляторы инициации листа
Белки PIN
Синтез ауксинов в молодых листьях
Транспорт
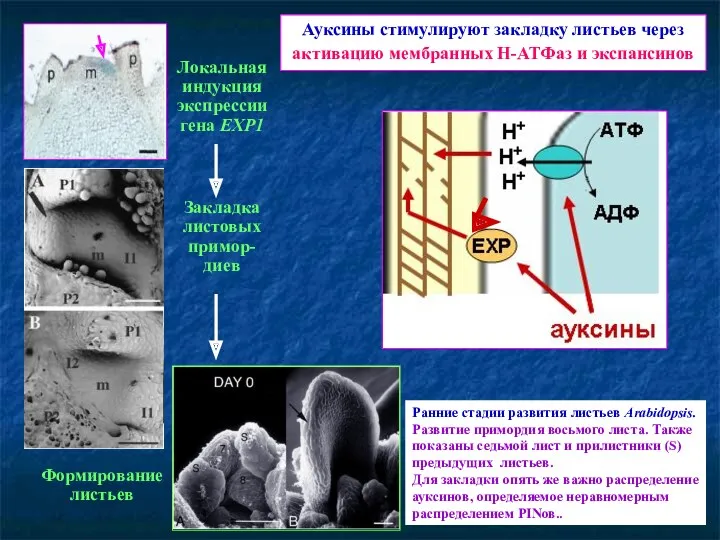
Ауксины стимулируют закладку листьев через активацию мембранных Н-АТФаз и экспансинов
Локальная индукция
Ауксины стимулируют закладку листьев через активацию мембранных Н-АТФаз и экспансинов
Локальная индукция

Гомеозисные гены
Гомеозисные гены — гены, определяющие процессы роста и дифференцировки в организме.
Гомеозисные гены
Гомеозисные гены — гены, определяющие процессы роста и дифференцировки в организме.
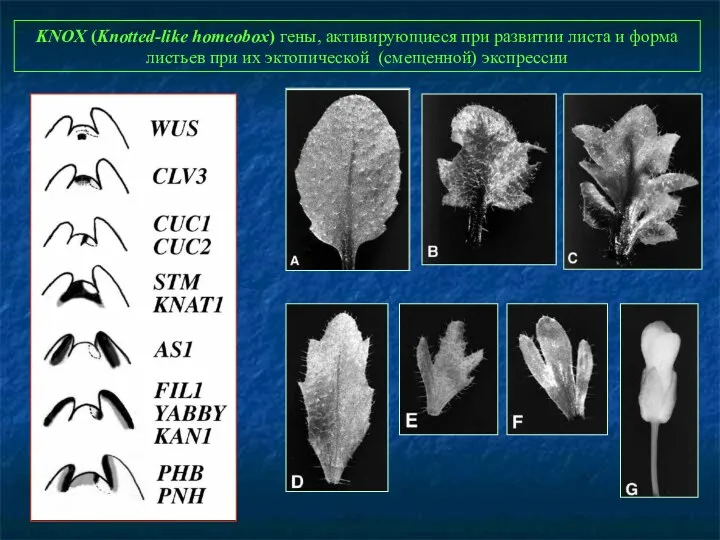
KNOX (Knotted-like homeobox) гены, активирующиеся при развитии листа и форма листьев
KNOX (Knotted-like homeobox) гены, активирующиеся при развитии листа и форма листьев

Форма розеточных листьев у мутантов по KNOX - генам
Форма розеточных листьев у мутантов по KNOX - генам
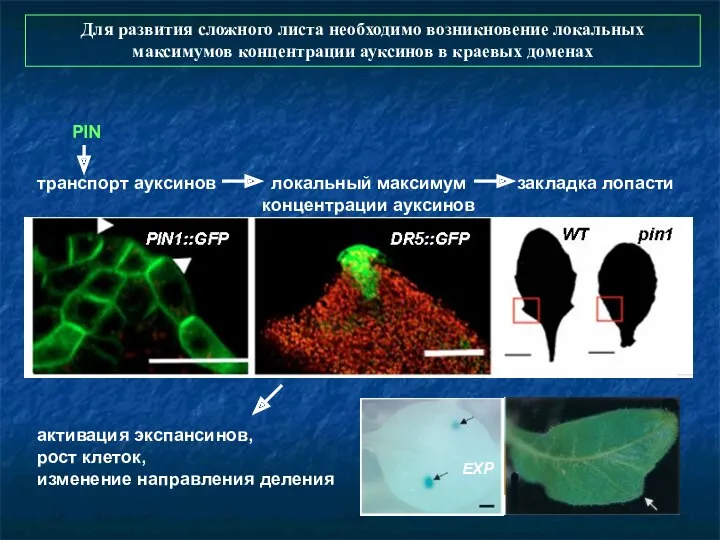
PIN
транспорт ауксинов
локальный максимум концентрации ауксинов
закладка лопасти
активация экспансинов,
рост клеток,
изменение
PIN
транспорт ауксинов
локальный максимум концентрации ауксинов
закладка лопасти
активация экспансинов,
рост клеток,
изменение

Подпрограммы развития: закладка пазушных почек и ветвление
После образования листа (но никогда
Подпрограммы развития: закладка пазушных почек и ветвление
После образования листа (но никогда

Подпрограммы развития: рост междоузлия
Формирование листа → синтез ГК → транспорт в
Подпрограммы развития: рост междоузлия
Формирование листа → синтез ГК → транспорт в
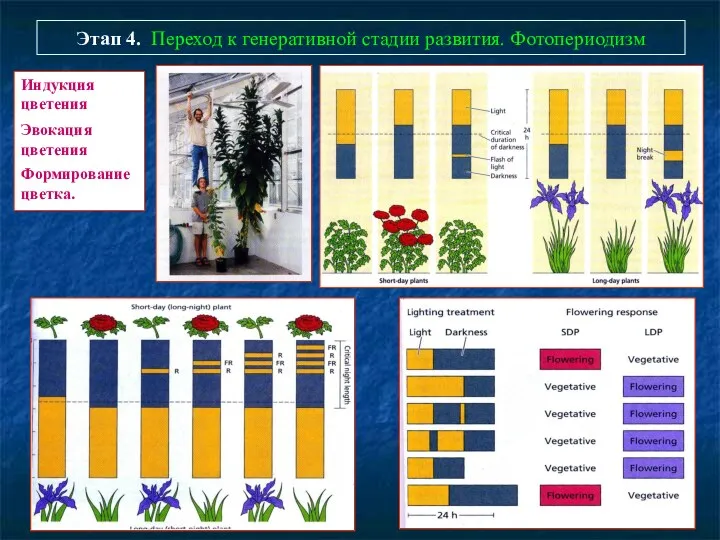
Этап 4. Переход к генеративной стадии развития. Фотопериодизм
Индукция цветения
Эвокация цветения
Формирование
цветка.
Этап 4. Переход к генеративной стадии развития. Фотопериодизм
Индукция цветения
Эвокация цветения
Формирование
цветка.
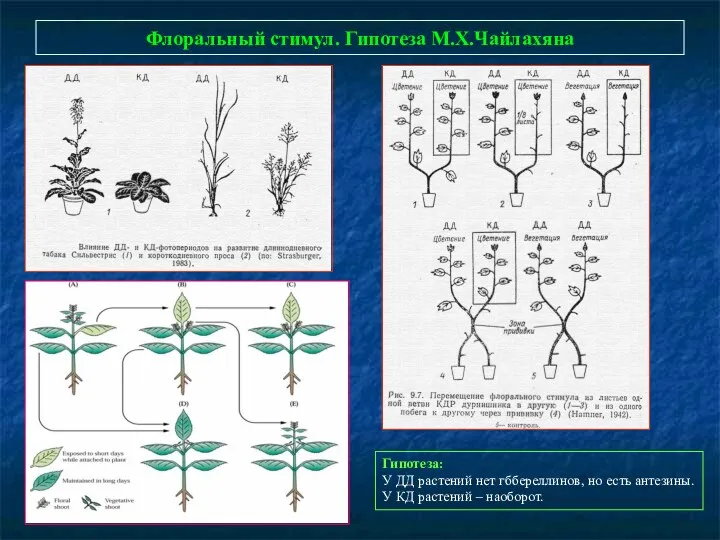
Флоральный стимул. Гипотеза М.Х.Чайлахяна
Гипотеза:
У ДД растений нет гббереллинов, но есть антезины.
У
Флоральный стимул. Гипотеза М.Х.Чайлахяна
Гипотеза:
У ДД растений нет гббереллинов, но есть антезины.
У
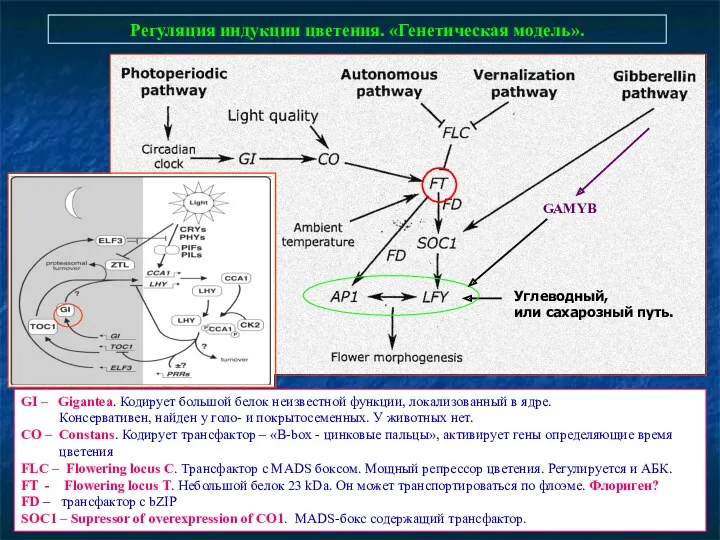
Регуляция индукции цветения. «Генетическая модель».
GI – Gigantea. Кодирует большой белок неизвестной
Регуляция индукции цветения. «Генетическая модель».
GI – Gigantea. Кодирует большой белок неизвестной
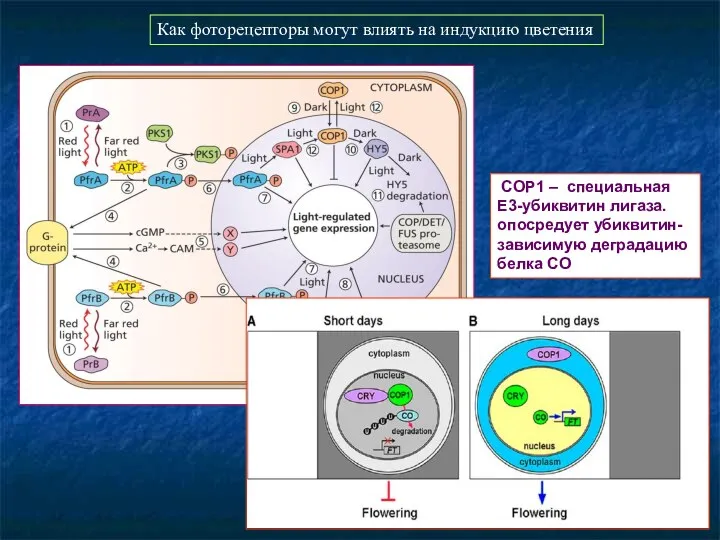
COP1 – специальная Е3-убиквитин лигаза. опосредует убиквитин-зависимую деградацию белка CO
Как
COP1 – специальная Е3-убиквитин лигаза. опосредует убиквитин-зависимую деградацию белка CO
Как
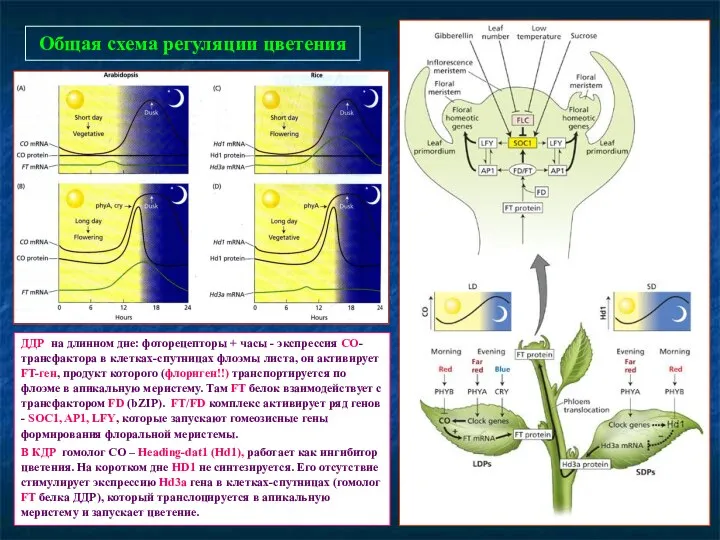
Общая схема регуляции цветения
ДДР на длинном дне: фоторецепторы + часы -
Общая схема регуляции цветения
ДДР на длинном дне: фоторецепторы + часы -
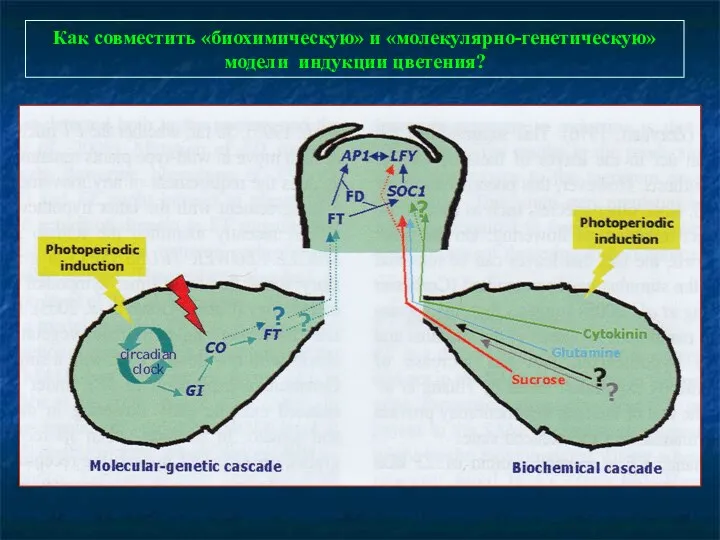
Как совместить «биохимическую» и «молекулярно-генетическую» модели индукции цветения?
Как совместить «биохимическую» и «молекулярно-генетическую» модели индукции цветения?

Четыре пути регулирования цветения…
Фотопериодический путь. Начинается в листьях. Участвуют фитохромы и
Четыре пути регулирования цветения…
Фотопериодический путь. Начинается в листьях. Участвуют фитохромы и
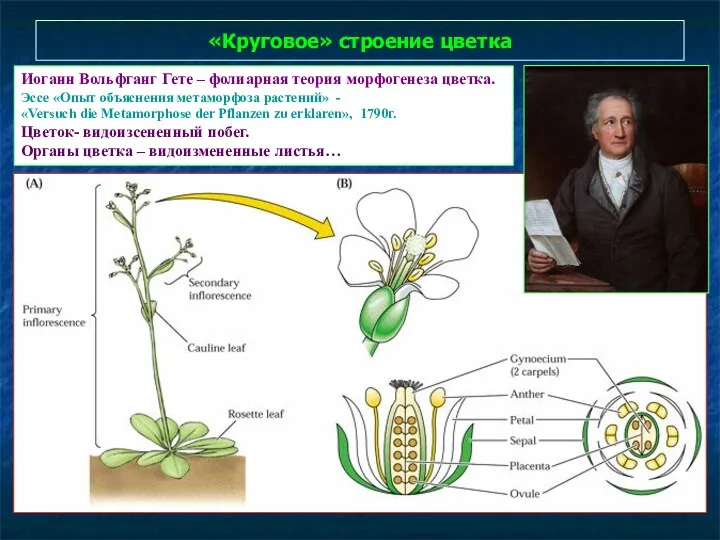
«Круговое» строение цветка
Иоганн Вольфганг Гете – фолиарная теория морфогенеза цветка. Эссе
«Круговое» строение цветка
Иоганн Вольфганг Гете – фолиарная теория морфогенеза цветка. Эссе
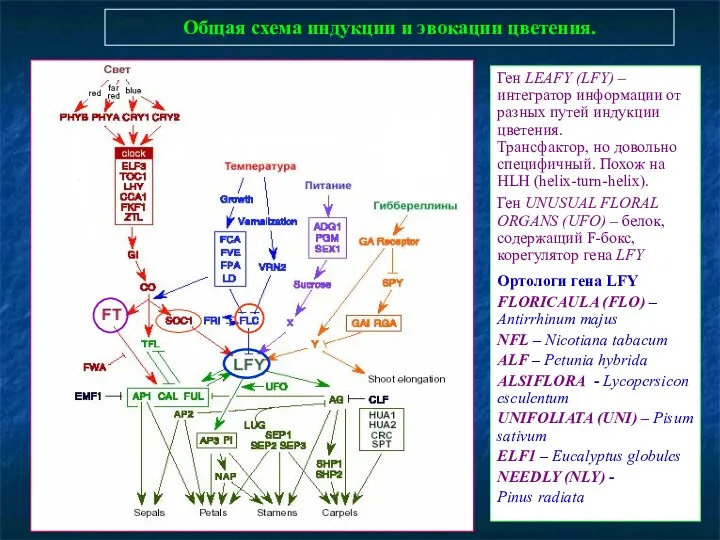
Общая схема индукции и эвокации цветения.
Ген LEAFY (LFY) – интегратор информации
Общая схема индукции и эвокации цветения.
Ген LEAFY (LFY) – интегратор информации
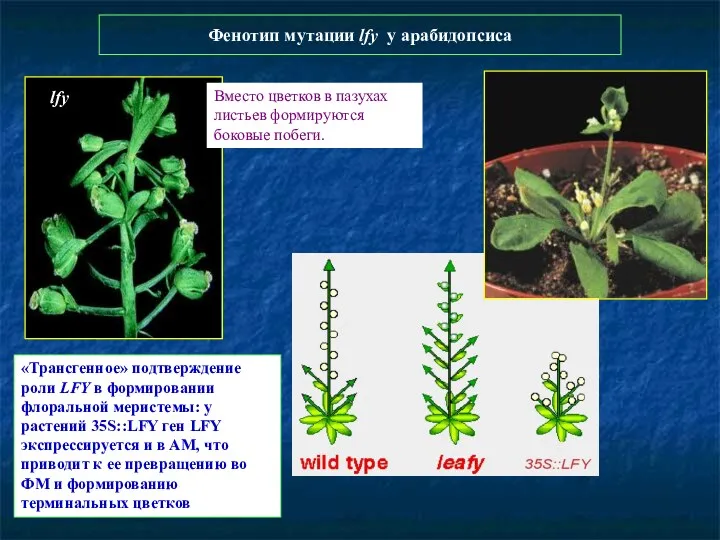
Фенотип мутации lfy у арабидопсиса
lfy
Вместо цветков в пазухах листьев формируются
Фенотип мутации lfy у арабидопсиса
lfy
Вместо цветков в пазухах листьев формируются
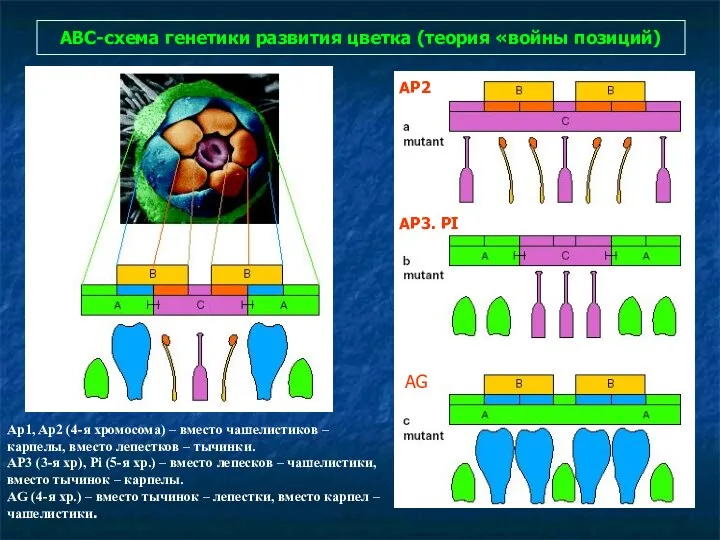
АВС-схема генетики развития цветка (теория «войны позиций)
Ap1, Ap2 (4-я хромосома) –
АВС-схема генетики развития цветка (теория «войны позиций)
Ap1, Ap2 (4-я хромосома) –

Мутанты арабидопсиса с точки зрения АВС-системы и гены.
AP1, AP2 – функция
Мутанты арабидопсиса с точки зрения АВС-системы и гены.
AP1, AP2 – функция
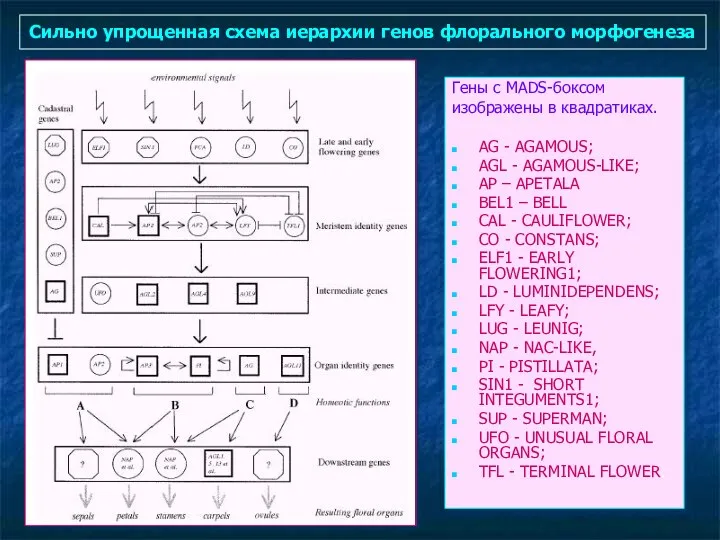
Сильно упрощенная схема иерархии генов флорального морфогенеза
Гены с MADS-боксом
изображены в
Сильно упрощенная схема иерархии генов флорального морфогенеза
Гены с MADS-боксом
изображены в
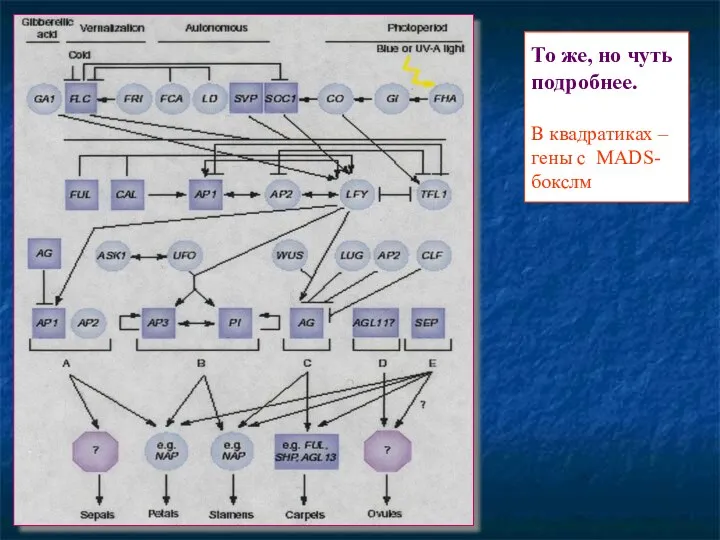
То же, но чуть подробнее.
В квадратиках – гены с MADS-бокслм
То же, но чуть подробнее.
В квадратиках – гены с MADS-бокслм
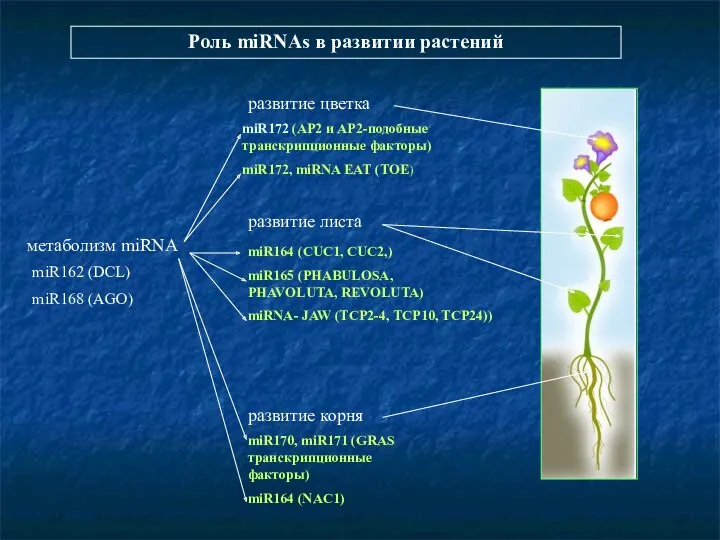
Роль miRNAs в развитии растений
развитие цветка
развитие листа
развитие корня
miR170, miR171 (GRAS транскрипционные
Роль miRNAs в развитии растений
развитие цветка
развитие листа
развитие корня
miR170, miR171 (GRAS транскрипционные

A. wt B. ap2-9 C. 35S::MIR172a-1
Фенотип трансгенных растений 35S::MIR172a-1 повторяет
A. wt B. ap2-9 C. 35S::MIR172a-1
Фенотип трансгенных растений 35S::MIR172a-1 повторяет
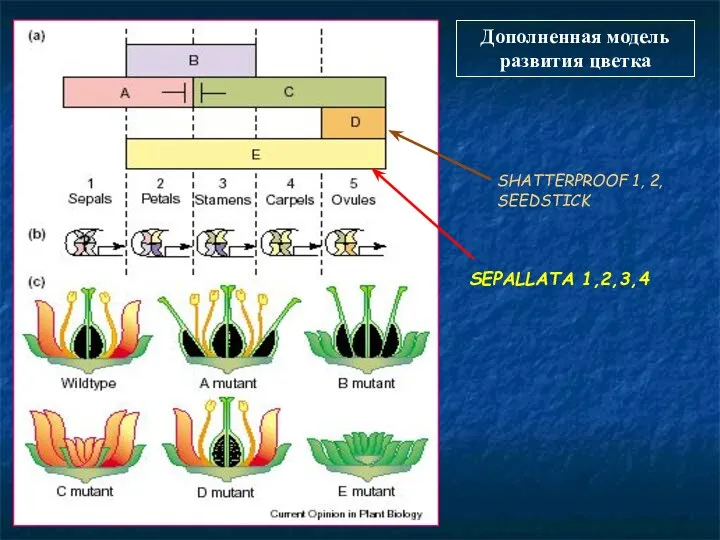
Дополненная модель развития цветка
SHATTERPROOF 1, 2,
SEEDSTICK
SEPALLATA 1,2,3,4
Дополненная модель развития цветка
SHATTERPROOF 1, 2,
SEEDSTICK
SEPALLATA 1,2,3,4
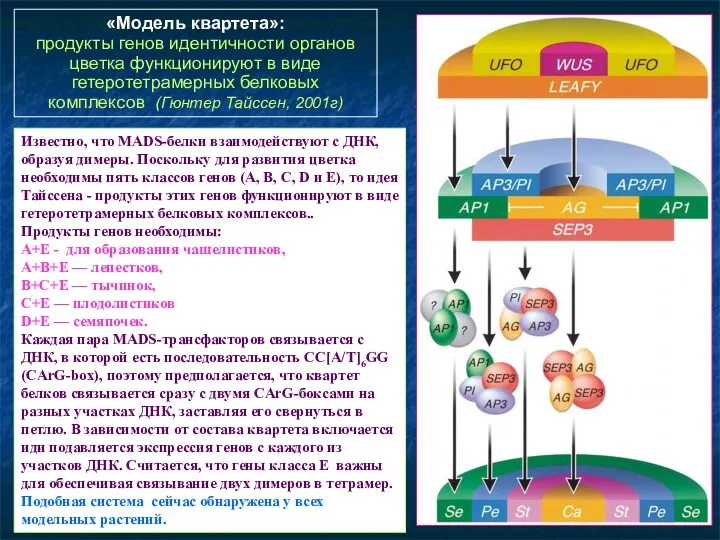
«Модель квартета»:
продукты генов идентичности органов цветка функционируют в виде гетеротетрамерных
«Модель квартета»:
продукты генов идентичности органов цветка функционируют в виде гетеротетрамерных
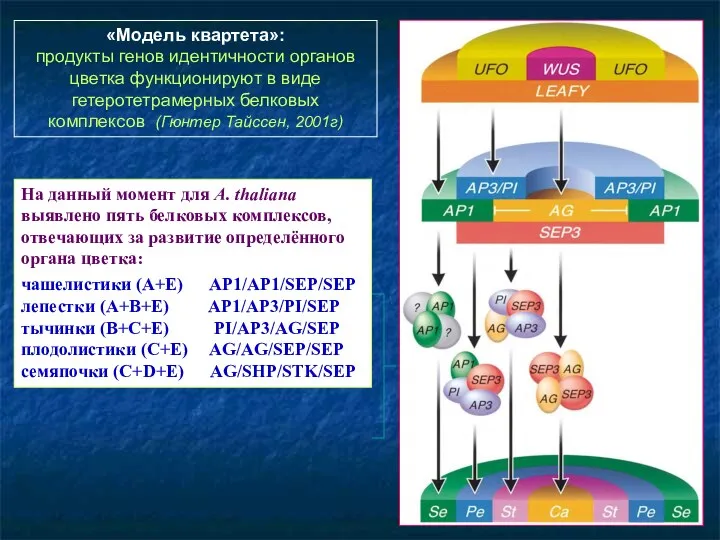
«Модель квартета»:
продукты генов идентичности органов цветка функционируют в виде гетеротетрамерных
«Модель квартета»:
продукты генов идентичности органов цветка функционируют в виде гетеротетрамерных

Мутанты по структуре цветка сходны у разных растений
Мутанты по структуре цветка сходны у разных растений
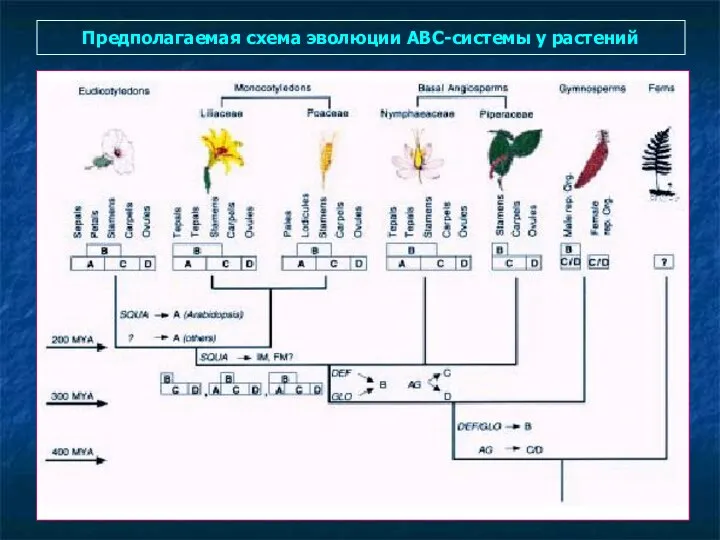
Предполагаемая схема эволюции АВС-системы у растений
Предполагаемая схема эволюции АВС-системы у растений

Детерминация пола у растений
Определение (детерминация) пола у растений - формирование признаков
Детерминация пола у растений
Определение (детерминация) пола у растений - формирование признаков

Аллосомы или sex-хромосомы - хромосомы отличающиеся по морфологии и количеству в
Аллосомы или sex-хромосомы - хромосомы отличающиеся по морфологии и количеству в

Хромосомное определение пола у растений
Большинство видов растений, у которых определение пола
Хромосомное определение пола у растений
Большинство видов растений, у которых определение пола
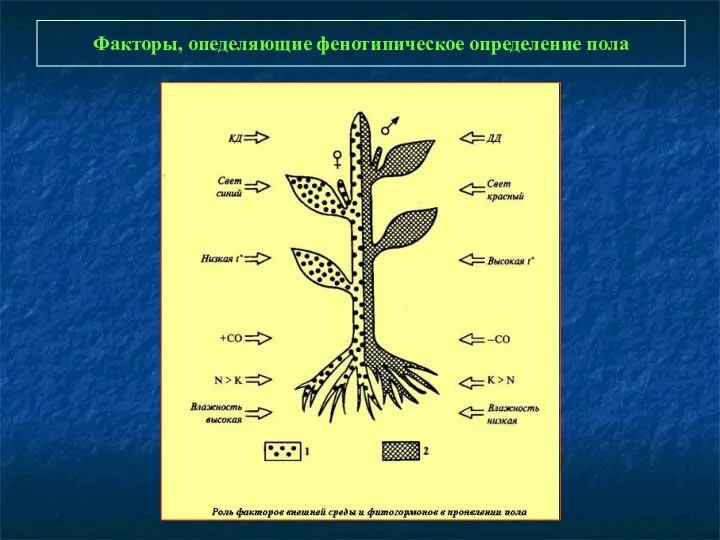
Факторы, опеделяющие фенотипическое определение пола
Факторы, опеделяющие фенотипическое определение пола


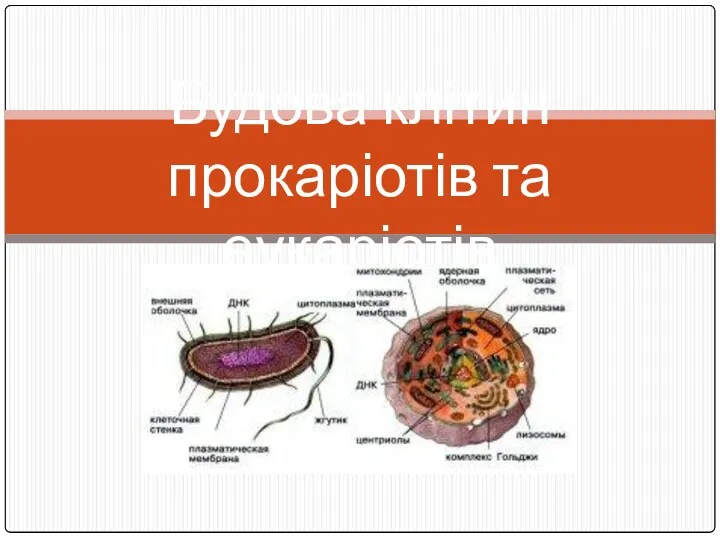 Будова клітин прокаріотів та еукаріотів
Будова клітин прокаріотів та еукаріотів Наследственность и патология
Наследственность и патология Развитие (мозговой пузырь)
Развитие (мозговой пузырь) Многолетние декоративные травянистые растения
Многолетние декоративные травянистые растения Организменная среда обитания. 5 класс
Организменная среда обитания. 5 класс Половое размножение животных
Половое размножение животных Витамины. Классификация витаминов
Витамины. Классификация витаминов Транскрипция у эукариот
Транскрипция у эукариот Пищеварительная система. Ротовая полость
Пищеварительная система. Ротовая полость Tissues: group of cells that are similar in structure and function
Tissues: group of cells that are similar in structure and function Общая вирусология
Общая вирусология Биотехнология, ее направления
Биотехнология, ее направления Индустрия ферментов
Индустрия ферментов Значение растений в природе и жизни человека
Значение растений в природе и жизни человека Строение и работа сердца. Круги кровообращения
Строение и работа сердца. Круги кровообращения Мутации: закономерность или парадокс?
Мутации: закономерность или парадокс? Онтогенез и филогенез нервной системы
Онтогенез и филогенез нервной системы Внешнее строение рыб
Внешнее строение рыб Презентация к уроку Насекомые
Презентация к уроку Насекомые Высшая нервная деятельность человека. 8 класс
Высшая нервная деятельность человека. 8 класс Растительные сообщества
Растительные сообщества Синквейн Жизнь
Синквейн Жизнь Кожа. Анатомия кожи. Функции кожи
Кожа. Анатомия кожи. Функции кожи Семейства Розоцветные и Паслёновые
Семейства Розоцветные и Паслёновые Проект. Использование современных технологий в обучении биологии школьников
Проект. Использование современных технологий в обучении биологии школьников Экологические группы растений
Экологические группы растений Хижі птахи
Хижі птахи Физиология продолговатого мозга и моста
Физиология продолговатого мозга и моста