Системы, которые создают корневое давление и ответственны за поднятие корневого водного раствора вверх по сосудам ксилемы презентация
- Системы, которые создают корневое давление и ответственны за поднятие корневого водного раствора вверх по сосудам ксилемы

Содержание
- 2. Системы, которые создают корневое давление и ответственны за поднятие корневого водного раствора (пасоки) вверх по сосудам
- 3. Верхний концевой двигатель – системы, обеспечивающие присасывающую силу листьев вследствие испарения воды через устьица и снижения
- 4. Воздух Ψ = −100.0 Mpa Лист (межклетник) Ψ = −7.0 Mpa Ксилемный сок (пасока) Клетки мезофилла
- 5. Минеральное питание растений Классификация элементов минерального питания и их функции в растении
- 6. Насколько важно минеральное питания? Морфология и урожайность растения напрямую зависит от доступности элементов минерального питания.
- 7. Концентрация минеральных веществ в самих растениях в большинстве случаев мало связана с их концентрацией в среде.
- 8. Коэффициенты накопления минеральных элементов (включая trace-элементы) некоторыми растениями
- 9. Концентрация питательных элементов в растительном материале при достаточном уровне обеспеченности
- 10. Макро и микроэлементы минерального питания.
- 11. CO2 O2 O2 CO2 H2O H2O Минеральные элементы
- 12. более 0.1% (содержание 1-20 г кг-1 сухого веса) Неминеральные элементы питания: H, O, & C Они
- 13. более 0.1% (содержание 1-20 г кг-1 сухого веса) Первичные и вторичные минеральные макроэлементы: Первичные – N,
- 14. более 0.1% (содержание 1-20 г кг-1 сухого веса) Первичные и вторичные минеральные макроэлементы: Вторичные – Ca,
- 15. более 0.1% (содержание 1-20 г кг-1 сухого веса) Органогены: H, O, C & N 95-98% веса
- 16. Важность of P, K, Ca, Mg & S K+ - основной осмотик, ответственен за генерацию и
- 17. Важность of P, K, Ca, Mg & S Ca2+ - главный сигнальный агент растительной клетки, ответственен
- 18. Важность of P, K, Ca, Mg & S Mg2+ - важнейший активатор ферментов и кофактор в
- 19. Здоровый лист кукурузы Дефицит фосфора Дефицит калия Дефицит азота
- 20. Микроэлементы: менее 0.1% на сухую массу ( Микроэлементы критически важны для жизни растений, но необходимы в
- 21. Микроэлементы: менее 0.1% на сухую массу ( Cu – имеет схожую функцию с железом. Входит в
- 22. Zn – нужен для стабильности клеточной стенки, мембран и ДНК, некоторых фотосинтетических реакциях, компонент важнейшего антиоксиданта
- 23. K Ca Fe Cu Zn Первичные роли минеральных элементов: - структурные компоненты - метаболические компоненты -
- 24. Катионы и состояние геля/золя. Катионы влияют на гидратацию (оводненность) цитоплазмы, причем щелочные металлы (например, K+) повышают
- 25. Хелаты металлов играют важную роль в жизни клеток. Наиболее важная функция большинства металлов – это участие
- 26. Хелаты металлов играют важную роль в жизни клеток. 2. Металлы стабилизируют определенную конфармационную структуру белковой молекулы,
- 27. Доступность минеральных элементов Зависит о характеристик почвы: pH, размера и состава почвенных частиц, других факторов. Большинство
- 29. Тройное правило Арнона (1939 г.) Элемент признается необходимым в случае, когда: 1. Растение без него не
- 30. Физиологические функции минеральных элементов
- 31. Физиологические функции минеральных элементов
- 33. Скачать презентацию

Системы, которые создают корневое давление и ответственны за поднятие корневого водного
Системы, которые создают корневое давление и ответственны за поднятие корневого водного
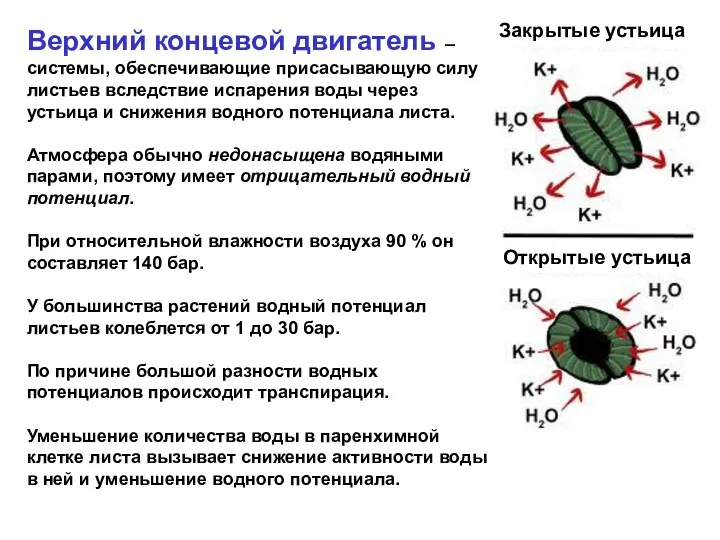
Верхний концевой двигатель – системы, обеспечивающие присасывающую силу листьев вследствие испарения
Верхний концевой двигатель – системы, обеспечивающие присасывающую силу листьев вследствие испарения
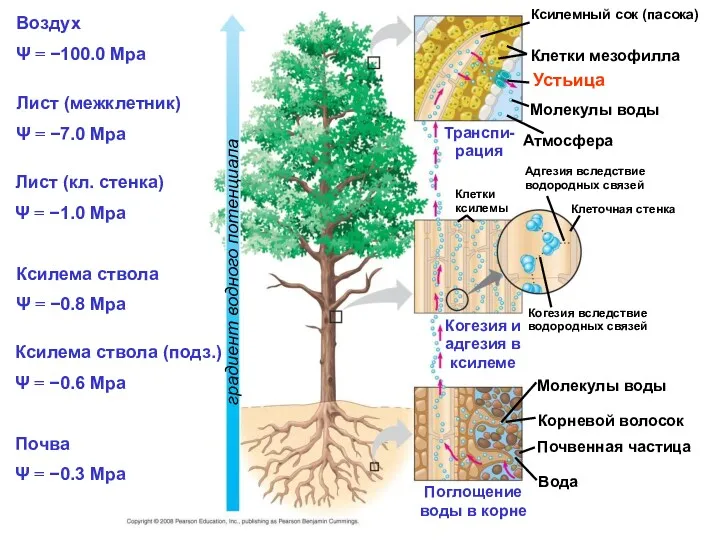
Воздух
Ψ = −100.0 Mpa
Лист (межклетник)
Ψ = −7.0 Mpa
Ксилемный сок (пасока)
Клетки
Воздух
Ψ = −100.0 Mpa
Лист (межклетник)
Ψ = −7.0 Mpa
Ксилемный сок (пасока)
Клетки

Минеральное
питание растений
Классификация элементов минерального питания и их функции в
Минеральное
питание растений
Классификация элементов минерального питания и их функции в
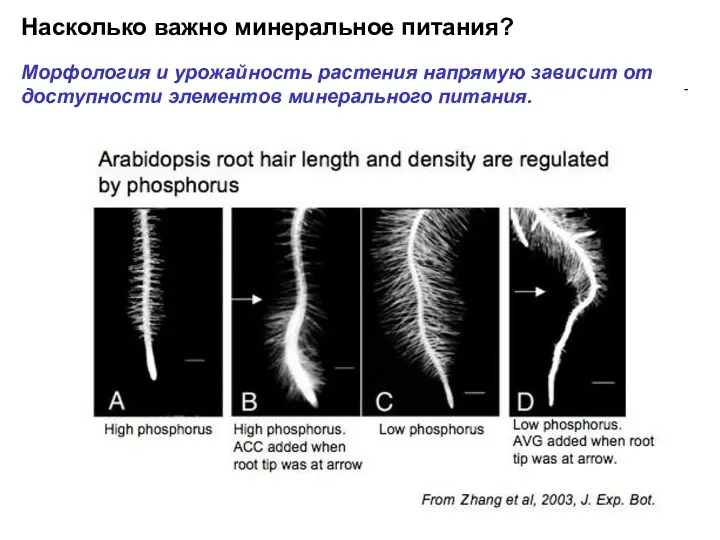
Насколько важно минеральное питания?
Морфология и урожайность растения напрямую зависит от
Насколько важно минеральное питания?
Морфология и урожайность растения напрямую зависит от

Концентрация минеральных веществ в самих растениях в большинстве случаев мало связана
Концентрация минеральных веществ в самих растениях в большинстве случаев мало связана
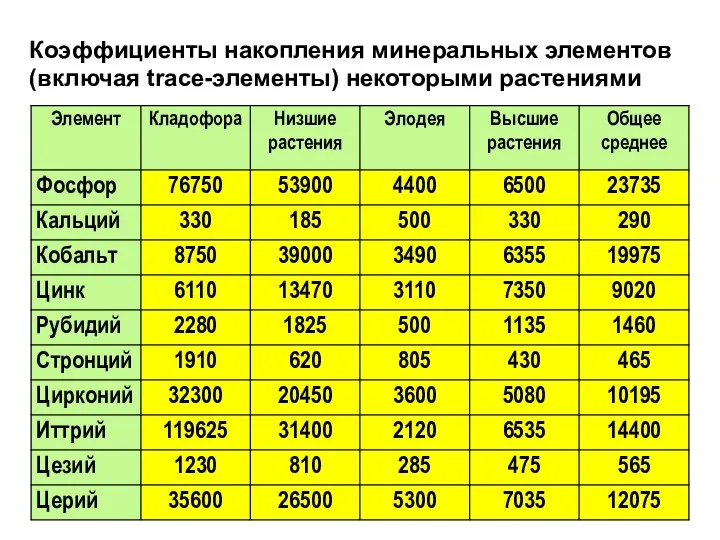
Коэффициенты накопления минеральных элементов (включая trace-элементы) некоторыми растениями
Коэффициенты накопления минеральных элементов (включая trace-элементы) некоторыми растениями
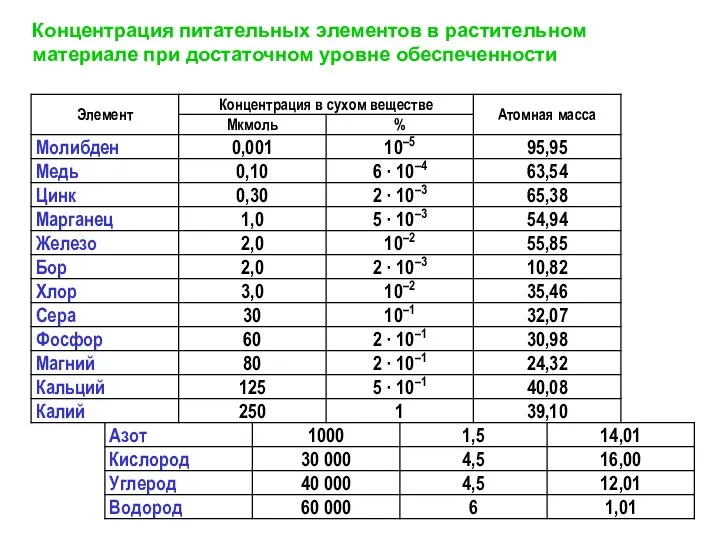
Концентрация питательных элементов в растительном материале при достаточном уровне обеспеченности
Концентрация питательных элементов в растительном материале при достаточном уровне обеспеченности

Макро и
микроэлементы
минерального
питания.
Макро и
микроэлементы
минерального
питания.
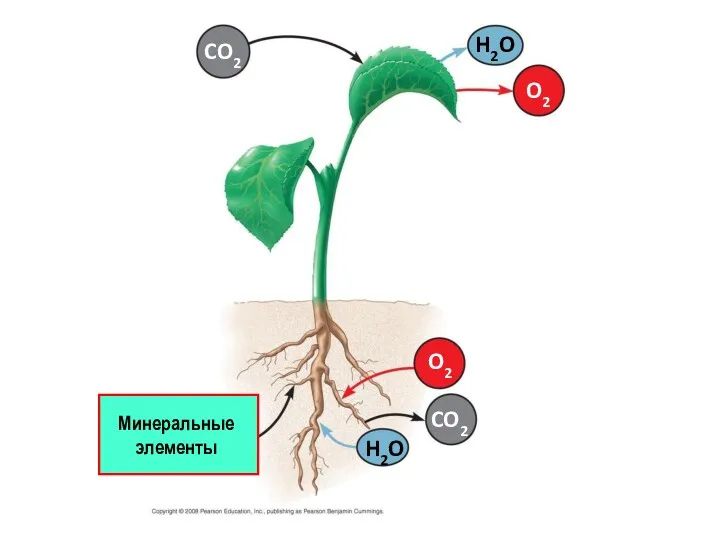
CO2
O2
O2
CO2
H2O
H2O
Минеральные элементы
CO2
O2
O2
CO2
H2O
H2O
Минеральные элементы

более 0.1%
(содержание 1-20 г кг-1 сухого веса)
Неминеральные элементы питания: H, O,
более 0.1%
(содержание 1-20 г кг-1 сухого веса)
Неминеральные элементы питания: H, O,

более 0.1%
(содержание 1-20 г кг-1 сухого веса)
Первичные и вторичные минеральные макроэлементы:
Первичные
более 0.1%
(содержание 1-20 г кг-1 сухого веса)
Первичные и вторичные минеральные макроэлементы:
Первичные

более 0.1%
(содержание 1-20 г кг-1 сухого веса)
Первичные и вторичные минеральные макроэлементы:
Вторичные
более 0.1%
(содержание 1-20 г кг-1 сухого веса)
Первичные и вторичные минеральные макроэлементы:
Вторичные

более 0.1%
(содержание 1-20 г кг-1 сухого веса)
Органогены: H, O, C &
более 0.1%
(содержание 1-20 г кг-1 сухого веса)
Органогены: H, O, C &

Важность of P, K, Ca, Mg & S
K+ - основной
Важность of P, K, Ca, Mg & S
K+ - основной

Важность of P, K, Ca, Mg & S
Ca2+ - главный
Важность of P, K, Ca, Mg & S
Ca2+ - главный

Важность of P, K, Ca, Mg & S
Mg2+ - важнейший
Важность of P, K, Ca, Mg & S
Mg2+ - важнейший
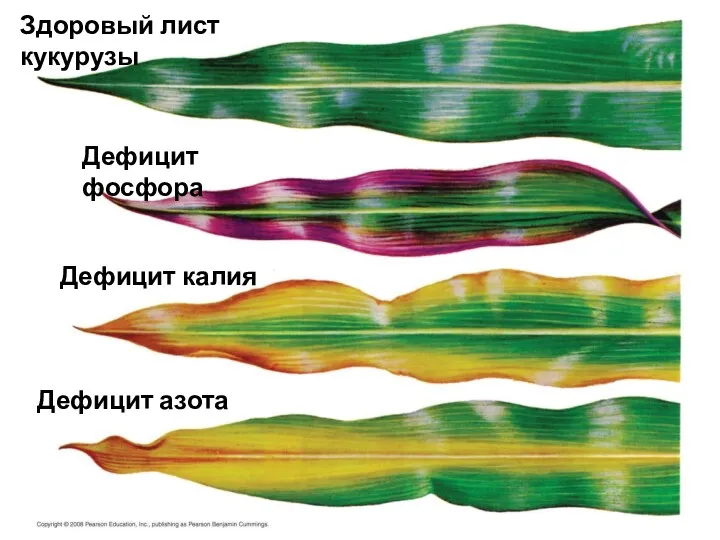
Здоровый лист кукурузы
Дефицит фосфора
Дефицит калия
Дефицит азота
Здоровый лист кукурузы
Дефицит фосфора
Дефицит калия
Дефицит азота

Микроэлементы:
менее 0.1% на сухую массу
(<1 г кг-1 сухого веса)
Микроэлементы критически важны
Микроэлементы:
менее 0.1% на сухую массу
(<1 г кг-1 сухого веса)
Микроэлементы критически важны

Микроэлементы:
менее 0.1% на сухую массу
(<1 г кг-1 сухого веса)
Cu – имеет
Микроэлементы:
менее 0.1% на сухую массу
(<1 г кг-1 сухого веса)
Cu – имеет

Zn – нужен для стабильности клеточной стенки, мембран и ДНК, некоторых
Zn – нужен для стабильности клеточной стенки, мембран и ДНК, некоторых

K
Ca
Fe
Cu
Zn
Первичные роли минеральных элементов:
- структурные компоненты
- метаболические компоненты
- активаторы и/или регуляторы
K
Ca
Fe
Cu
Zn
Первичные роли минеральных элементов:
- структурные компоненты
- метаболические компоненты
- активаторы и/или регуляторы

Катионы и состояние геля/золя.
Катионы влияют на гидратацию (оводненность) цитоплазмы, причем щелочные
Катионы и состояние геля/золя.
Катионы влияют на гидратацию (оводненность) цитоплазмы, причем щелочные

Хелаты металлов играют важную роль в жизни клеток.
Наиболее важная функция
Хелаты металлов играют важную роль в жизни клеток.
Наиболее важная функция

Хелаты металлов играют важную роль в жизни клеток.
2. Металлы стабилизируют
Хелаты металлов играют важную роль в жизни клеток.
2. Металлы стабилизируют

Доступность минеральных элементов
Зависит о характеристик почвы: pH, размера и состава почвенных
Доступность минеральных элементов
Зависит о характеристик почвы: pH, размера и состава почвенных

Тройное правило Арнона (1939 г.)
Элемент признается необходимым в случае, когда:
Тройное правило Арнона (1939 г.)
Элемент признается необходимым в случае, когда:
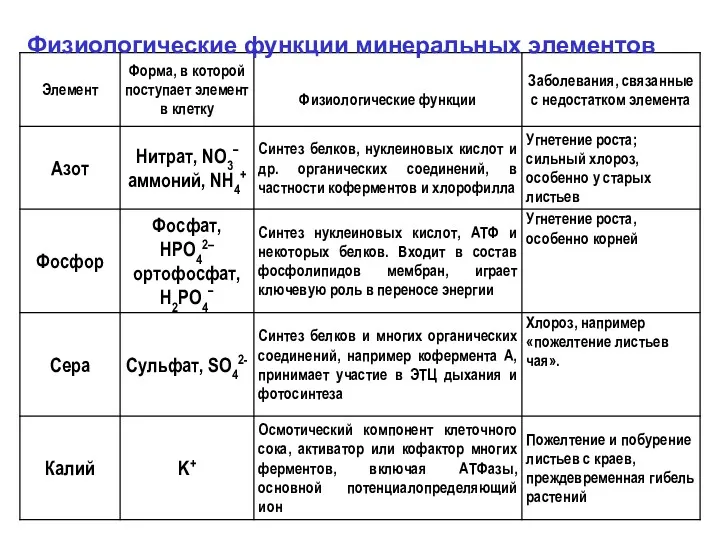
Физиологические функции минеральных элементов
Физиологические функции минеральных элементов
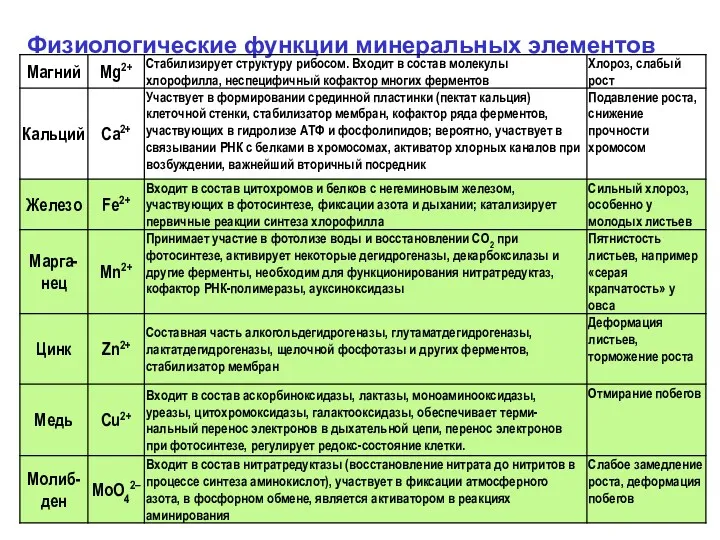
Физиологические функции минеральных элементов
Физиологические функции минеральных элементов
 Antigen-antibody reactions and selected tests
Antigen-antibody reactions and selected tests Строение эукариотических клеток
Строение эукариотических клеток Лекарственные растения рек Золотой Китат и Алчедат
Лекарственные растения рек Золотой Китат и Алчедат Огуречные именины. Правила игры
Огуречные именины. Правила игры Семейство Паслёновые
Семейство Паслёновые Поведінка риб
Поведінка риб Хищные растения
Хищные растения Майстерність маскування
Майстерність маскування Разнообразие животных
Разнообразие животных Применение БАДов
Применение БАДов Органические вещества
Органические вещества урок красная книга Мордовии
урок красная книга Мордовии Типы развития насекомых. Насекомые с неполным превращением
Типы развития насекомых. Насекомые с неполным превращением ВКР: Иммобилизованный биокатализатор на основе адгезированных амидазосодержащих клеток родококков для синтеза акриловой кислоты
ВКР: Иммобилизованный биокатализатор на основе адгезированных амидазосодержащих клеток родококков для синтеза акриловой кислоты Морфокинезиологический анализ пояса верхних конечностей
Морфокинезиологический анализ пояса верхних конечностей Грудная клетка. Лекция № 10
Грудная клетка. Лекция № 10 Nepryamoy_ontogenez
Nepryamoy_ontogenez Исследовательская работа Мой домашний питомец
Исследовательская работа Мой домашний питомец Живые ископаемые
Живые ископаемые Коммуникации у животных
Коммуникации у животных Расы человека
Расы человека Ферменты – 1
Ферменты – 1 Тип Nemathelminthes (Круглые Черви)
Тип Nemathelminthes (Круглые Черви) Будова і функції шкіри
Будова і функції шкіри Healthy eating
Healthy eating Покрытосеменные. Отличительные признаки покрытосеменных
Покрытосеменные. Отличительные признаки покрытосеменных Структура і функції білків. Ферменти. Вітаміни, гормони, фактори росту, їх роль у життєдіяльності організмів
Структура і функції білків. Ферменти. Вітаміни, гормони, фактори росту, їх роль у життєдіяльності організмів Питание бактерий
Питание бактерий