- Структура и организация генома, репликация ДНК

Содержание
- 2. Наследственный материал должен удовлетворять следующим требованиям: 1. Информативность: должен содержать информацию о строении и функции всего
- 3. Идентификация молекулы ДНК в качестве генетического материала Данные многих генетиков, включая Г.Менделя, согласуются с четырьмя перечисленными
- 4. Эксперименты Ф.Гриффитца с бактерией Streptococcus pneumoniae Гриффитц (1928) изучал бактерию (пневмококка) сейчас известную как Streptococcus pneumoniae
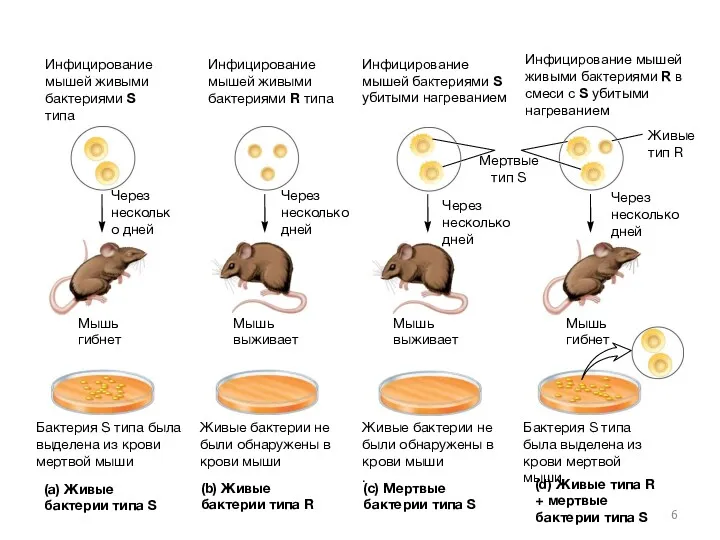
- 5. В 1928, Гриффитц провел эксперименты с использованием двух штаммов S. pneumoniae: тип IIIS и тип IIR
- 6. Инфицирование мышей живыми бактериями S типа Мышь гибнет Мертвые тип S Живые тип R Мышь гибнет
- 7. Гриффитц пришел к выводу, что что-то из мертвой бактерии S типа трансформировало бактерии R типа в
- 8. Голова Хвостовые нити Опорная плита Тело ДНК внутри капсидной оболочки Эксперимент Херши и Чейза с бактериофагом
- 9. Жизненный цикл бактериофага T2 Головка Бактериальная хромосома Клеточная стенка бактерии Генетический материал Оболочка фага (белок) связывается
- 10. Краткое описание эксперимента Херши и Чейза: Использование радиоизотопов для дифференциации ДНК и белка 32P специфично меченая
- 11. В 1956, A. Gierer и G. Schramm выделили РНК из вируса табачной мозаики (ВТМ) Очищенная РНК
- 12. Структура нуклеиновой кислоты ДНК и РНК – огромные молекулы с несколькими уровнями сложности 1. Нуклеотиды –
- 13. Фосфатная группа Сахар D-Дезоксирибоза (в ДНК) Пурины (двойное кольцо) Пиримидины (однокольцевые) Азотистые основания O O O
- 14. H H H O CH2 Основание (a) Основная структурная единица дезоксирибонуклеиновой кислоты (ДНК) (b) Основная структурная
- 15. Основание + сахар ? нуклеозид Пример Аденин + рибоза = Аденозин Аденин + дезоксирибоза = Дезоксиаденозин
- 16. Основание всегда присоединено здесь Аденозин Аденозин монофосфат Аденозин дифосфат Аденин Фосфатная группа Рибоза H O P
- 17. NH2 N O N O N O N Аденин (A) Гуанин (Г) Тимин (T) Основание Остов
- 18. В 1953 Джеймс Ватсон и Френсис Крик открыли двуспиральную структуру ДНК Открытие структуры ДНК (a) Watson
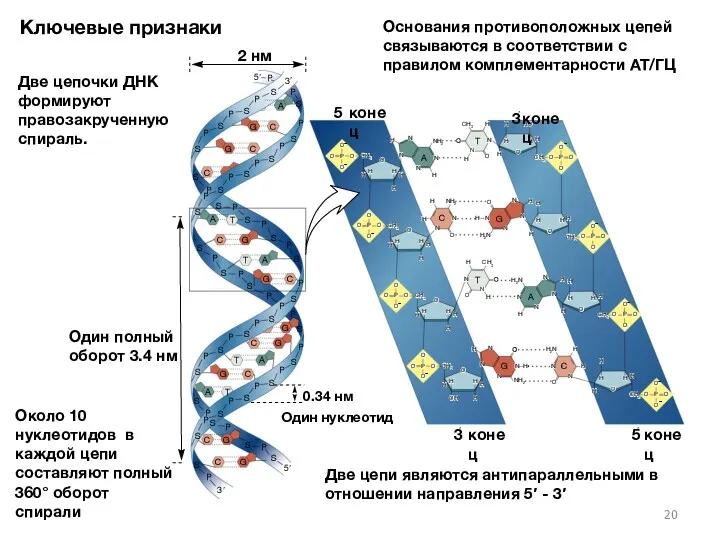
- 19. Основные структурные особенности Двуспиральная модель ДНК Две цепочки закручиваются вокруг общей оси 10 оснований и 3.4
- 20. H N H N N N G NH 2 H P S P S P 5
- 21. (b) Модель ДНК с заполненным пространством (a) Шарико-палочковая модель ДНК Малая бороздка Большая бороздка Малая бороздка
- 22. ДНК обвивается вокруг гистоновых белков Радиальные петли (300 нм в диаметре) Метафазная хромосома ДНК (диаметр 2
- 23. 3 метра ДНК/клетка 1013 клеток/человек 3 x 1013 метров ДНК/человек 3.8 x 108 метров от Земли
- 24. РЕПЛИКАЦИЯ ДНК Когда Уотсон и Крик предложили модель двойной спирали ДНК, то эта модель должна была
- 25. а) Механизм репликации ДНК b) Результат репликации Репликация ДНК Репликация ДНК – это процесс удвоения генетического
- 27. Уотсон и Крик предложили следующее: Поскольку ДНК содержит две нити, две цепи связанные между собой более
- 28. ДНК геликаза разрушает водородные связи между комплементарными цепями; Топоизомераза снимает положительную сверхспирализацию ДНК; белки SSB связываются
- 29. ДНК-полимераза III осуществляет на одиночных цепях синтез комплементарных цепей ДНК; для начала синтеза ДНК-полимеразе III необходимо
- 30. ДНК-полимераза III продолжает синтез цепи, постепенно заменяя нуклеотиды РНК между фрагментами; ДНК-лигаза сшивает фрагменты отстающей цепи
- 31. Аденин (А) Гуанин (Г) Урацил (У) Основания Остов Цитозинe (Ц) O H H H H O
- 32. Copyright ©The McGraw-Hill Companies, Inc. Permission required for reproduction or display Обычно РНК молекулы являются одноцепочечными,
- 33. A U A U U A G C C G C G A U U A
- 34. Many factors contribute to the tertiary structure of RNA For example Base-pairing and base stacking within
- 35. Транскрипция гена
- 36. ВВЕДЕНИЕ ДНК хранит информацию Для построения механизма (организма) обеспечивающего Не является самодостаточной молекулой Для построения и
- 37. Центральная догма генетики
- 38. Функция промотера – место определение транскрипционных факторов Транскрипционные факторы обеспечивают связь РНК полимеразы с промотером, формирующих
- 39. Ген может быть определен как дискретный участок ДНК транскрибируемый в РНК Ген так же называют транскрипционной
- 40. ДНК Регуляторная последовательность: место прикрепления регуляторного белка, определяющего уровень транскрипции Промотер: место прикрепления РНК полимеразы, определяет
- 41. Синтез РНК транскрипта РНК полимераза скользит вдоль ДНК, создавая открытый комплекс. Матричная цепочка ДНК используется в
- 42. Полипептидный синтез Процесс синтеза белка на матричной мРНК называется трансляцией, его основные этапы: активизация определенной аминокислоты
- 43. Модель бактериальной рибосомы 30S субъединица 50S субъединица Рибосомная РНК (рРНК): рибосома содержит 64% рРНК и 36%
- 44. Трансляция Субъединицы рибосомы Большая Малая UAC антикодон мРНК AUG Старт кодон UAG Стоп кодон Инициация AUG
- 45. Структура тРНК Транспортная РНК (тРНК): Осуществляет транспортировку аминокислот к рибосоме По трехмерной структуре тРНК напоминает форму
- 46. Структурные уровни формируемые белками В зависимости от аминокислотной последовательности Белковая субъединица
- 48. Скачать презентацию

Наследственный материал должен удовлетворять следующим требованиям:
1. Информативность: должен содержать информацию о
Наследственный материал должен удовлетворять следующим требованиям:
1. Информативность: должен содержать информацию о

Идентификация молекулы ДНК в качестве генетического материала
Данные многих генетиков, включая Г.Менделя,
Идентификация молекулы ДНК в качестве генетического материала
Данные многих генетиков, включая Г.Менделя,

Эксперименты Ф.Гриффитца с бактерией Streptococcus pneumoniae
Гриффитц (1928) изучал бактерию (пневмококка) сейчас
Эксперименты Ф.Гриффитца с бактерией Streptococcus pneumoniae
Гриффитц (1928) изучал бактерию (пневмококка) сейчас

В 1928, Гриффитц провел эксперименты с использованием двух штаммов S. pneumoniae:
В 1928, Гриффитц провел эксперименты с использованием двух штаммов S. pneumoniae:
Инфицирование мышей живыми бактериями S типа
Мышь гибнет
Мертвые
тип S
Живые
тип R
Мышь гибнет
Мышь выживает
Мышь
Инфицирование мышей живыми бактериями S типа
Мышь гибнет
Мертвые
тип S
Живые
тип R
Мышь гибнет
Мышь выживает
Мышь

Гриффитц пришел к выводу, что что-то из мертвой бактерии S типа
Гриффитц пришел к выводу, что что-то из мертвой бактерии S типа
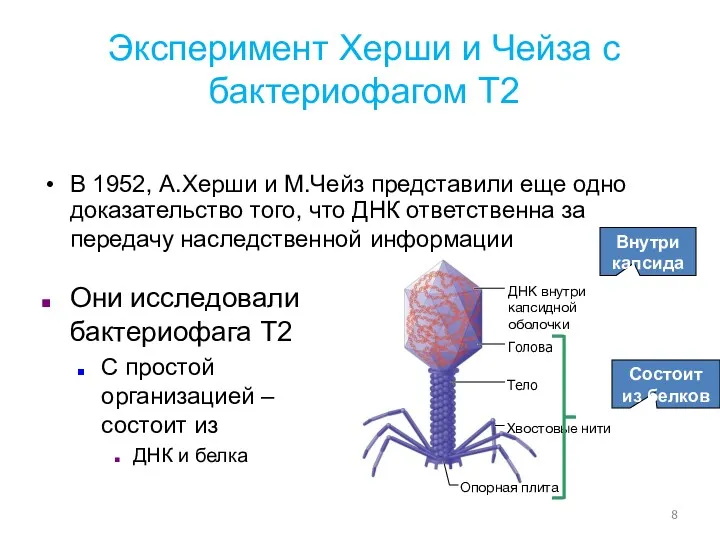
Голова
Хвостовые нити
Опорная плита
Тело
ДНК внутри капсидной оболочки
Эксперимент Херши и Чейза
Голова
Хвостовые нити
Опорная плита
Тело
ДНК внутри капсидной оболочки
Эксперимент Херши и Чейза
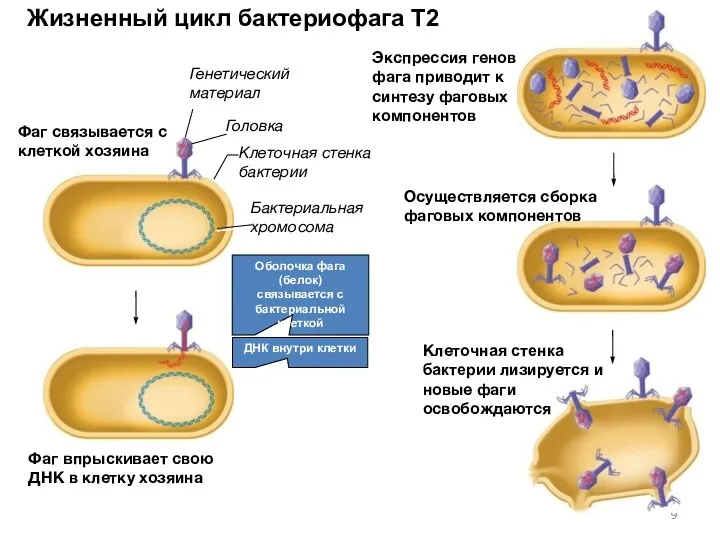
Жизненный цикл бактериофага T2
Головка
Бактериальная хромосома
Клеточная стенка бактерии
Генетический материал
Оболочка фага (белок)
Жизненный цикл бактериофага T2
Головка
Бактериальная хромосома
Клеточная стенка бактерии
Генетический материал
Оболочка фага (белок)

Краткое описание эксперимента Херши и Чейза:
Использование радиоизотопов для дифференциации ДНК и
Краткое описание эксперимента Херши и Чейза:
Использование радиоизотопов для дифференциации ДНК и

В 1956, A. Gierer и G. Schramm выделили РНК из вируса
В 1956, A. Gierer и G. Schramm выделили РНК из вируса
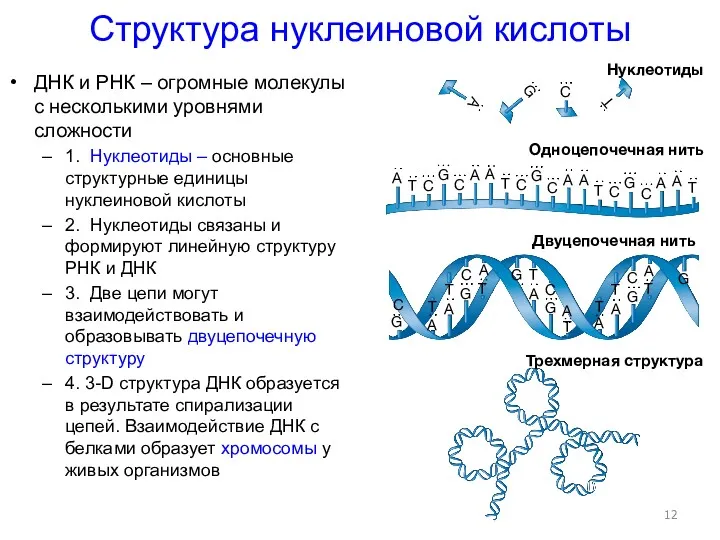
Структура нуклеиновой кислоты
ДНК и РНК – огромные молекулы с несколькими уровнями
Структура нуклеиновой кислоты
ДНК и РНК – огромные молекулы с несколькими уровнями
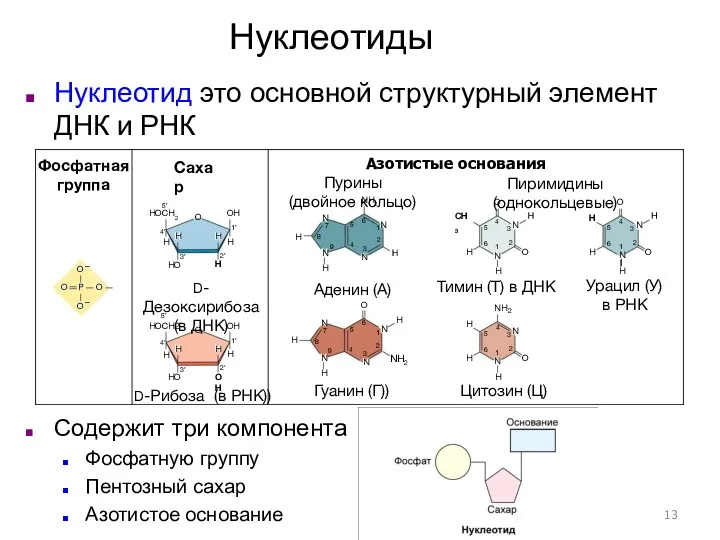
Фосфатная группа
Сахар
D-Дезоксирибоза (в ДНК)
Пурины
(двойное кольцо)
Пиримидины (однокольцевые)
Азотистые основания
O
O
O
–
O
–
P
H
H
H
HO
OH
O
HOCH2
H
H
D-Рибоза (в РНК))
H
OH
H
HO
OH
O
HOCH2
H
H
Урацил (У) в
Фосфатная группа
Сахар
D-Дезоксирибоза (в ДНК)
Пурины
(двойное кольцо)
Пиримидины (однокольцевые)
Азотистые основания
O
O
O
–
O
–
P
H
H
H
HO
OH
O
HOCH2
H
H
D-Рибоза (в РНК))
H
OH
H
HO
OH
O
HOCH2
H
H
Урацил (У) в
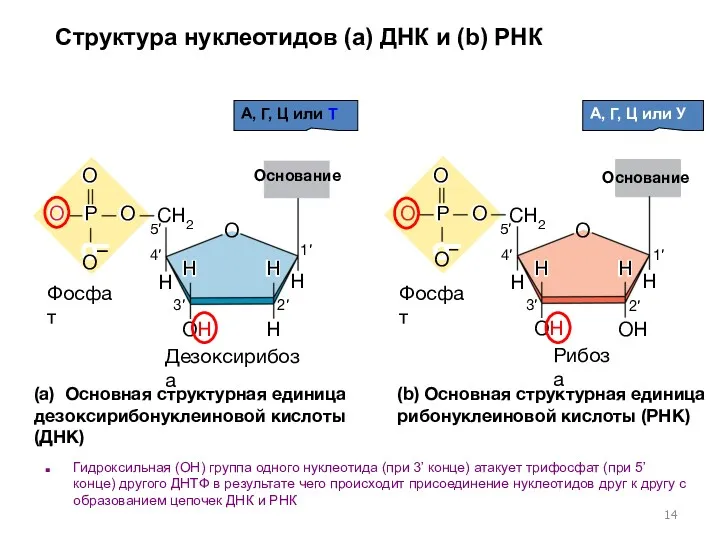
H
H
H
O
CH2
Основание
(a) Основная структурная единица дезоксирибонуклеиновой кислоты (ДНК)
(b) Основная структурная единица рибонуклеиновой
H
H
H
O
CH2
Основание
(a) Основная структурная единица дезоксирибонуклеиновой кислоты (ДНК)
(b) Основная структурная единица рибонуклеиновой

Основание + сахар ? нуклеозид
Пример
Аденин + рибоза = Аденозин
Аденин
Основание + сахар ? нуклеозид
Пример
Аденин + рибоза = Аденозин
Аденин
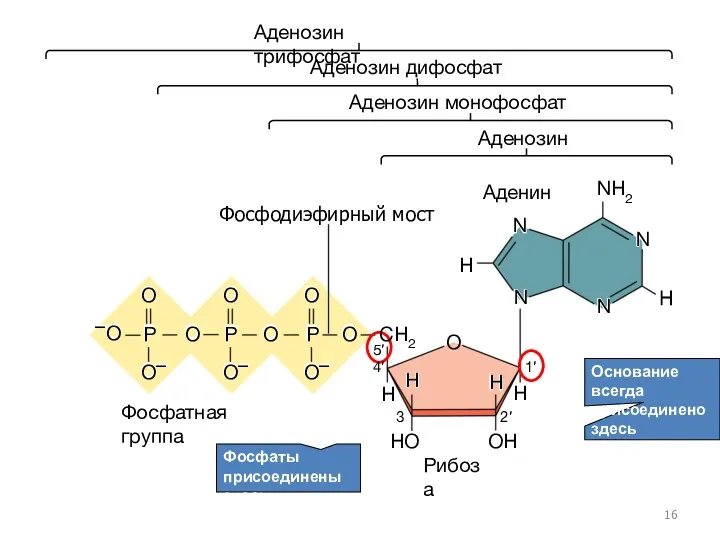
Основание всегда присоединено здесь
Аденозин
Аденозин монофосфат
Аденозин дифосфат
Аденин
Фосфатная группа
Рибоза
H
O
P
CH2
O
–
O
O
P
O
–
O
O
O
–
O
P
O
–
H
OH
HO
O
2′
3
1′
4′
5′
Аденозин трифосфат
NH2
H
Фосфодиэфирный мост
Основание всегда присоединено здесь
Аденозин
Аденозин монофосфат
Аденозин дифосфат
Аденин
Фосфатная группа
Рибоза
H
O
P
CH2
O
–
O
O
P
O
–
O
O
O
–
O
P
O
–
H
OH
HO
O
2′
3
1′
4′
5′
Аденозин трифосфат
NH2
H
Фосфодиэфирный мост
NH2
N
O
N
O
N
O
N
Аденин (A)
Гуанин (Г)
Тимин (T)
Основание
Остов
Цитозин (Ц)
O
H
H
H
H
H
H
O
O
O
O
–
P
CH2
O
–
H
H
H
H
H
H
H
O
O
O
O
P
CH2
O
–
NH2
N
N
H
N
N
H
H
H
H
H
O
O
O
O
P
CH2
O
–
NH2
H
N
N
N
H
N
H
H
H
OH
H
H
O
O
O
O
P
CH2
O
–
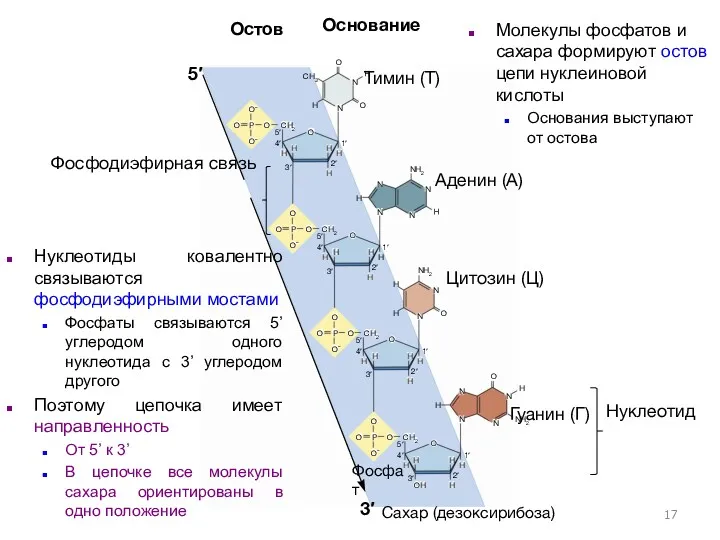
Нуклеотид
Фосфодиэфирная связь
Сахар (дезоксирибоза)
Фосфат
3′
5′
5′
4′
1′
2′
3′
5′
4′
1′
2′
3′
5′
4′
1′
2′
3′
5′
4′
1′
2′
3′
CH3
Нуклеотиды ковалентно связываются фосфодиэфирными мостами
Фосфаты
NH2
N
O
N
O
N
O
N
Аденин (A)
Гуанин (Г)
Тимин (T)
Основание
Остов
Цитозин (Ц)
O
H
H
H
H
H
H
O
O
O
O
–
P
CH2
O
–
H
H
H
H
H
H
H
O
O
O
O
P
CH2
O
–
NH2
N
N
H
N
N
H
H
H
H
H
O
O
O
O
P
CH2
O
–
NH2
H
N
N
N
H
N
H
H
H
OH
H
H
O
O
O
O
P
CH2
O
–
Нуклеотид
Фосфодиэфирная связь
Сахар (дезоксирибоза)
Фосфат
3′
5′
5′
4′
1′
2′
3′
5′
4′
1′
2′
3′
5′
4′
1′
2′
3′
5′
4′
1′
2′
3′
CH3
Нуклеотиды ковалентно связываются фосфодиэфирными мостами
Фосфаты

В 1953 Джеймс Ватсон и Френсис Крик открыли двуспиральную структуру ДНК
Открытие
В 1953 Джеймс Ватсон и Френсис Крик открыли двуспиральную структуру ДНК
Открытие

Основные структурные особенности
Двуспиральная модель ДНК
Две цепочки закручиваются вокруг общей оси
10 оснований
Основные структурные особенности
Двуспиральная модель ДНК
Две цепочки закручиваются вокруг общей оси
10 оснований
H
N
H
N
N
N
G
NH
2
H
P
S
P
S
P
5
′
конец
3
′
конец
H
NH
2
N
N
H
H
H
H
H
O
O
O
O
P
CH
2
O
H
H
H
OH
H
H
O
O
O
O
P
CH
2
O
H
H
H
H
H
O
O
O
O
P
CH
2
O
CH
2
H
H
H
H
H
O
O
O
O
P
O
T
Ключевые признаки
Две цепочки ДНК формируют правозакрученную спираль.
Основания противоположных цепей связываются в
H
N
H
N
N
N
G
NH
2
H
P
S
P
S
P
5
′
конец
3
′
конец
H
NH
2
N
N
H
H
H
H
H
O
O
O
O
P
CH
2
O
H
H
H
OH
H
H
O
O
O
O
P
CH
2
O
H
H
H
H
H
O
O
O
O
P
CH
2
O
CH
2
H
H
H
H
H
O
O
O
O
P
O
T
Ключевые признаки
Две цепочки ДНК формируют правозакрученную спираль.
Основания противоположных цепей связываются в
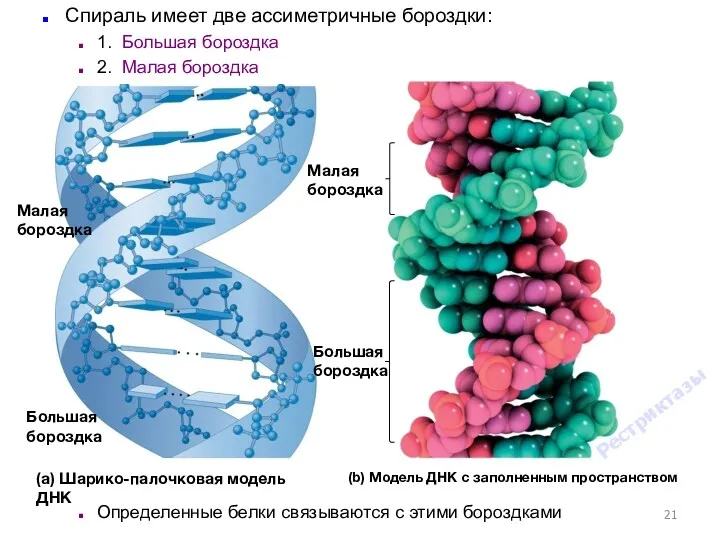
(b) Модель ДНК с заполненным пространством
(a) Шарико-палочковая модель ДНК
Малая бороздка
Большая бороздка
Малая
(b) Модель ДНК с заполненным пространством
(a) Шарико-палочковая модель ДНК
Малая бороздка
Большая бороздка
Малая
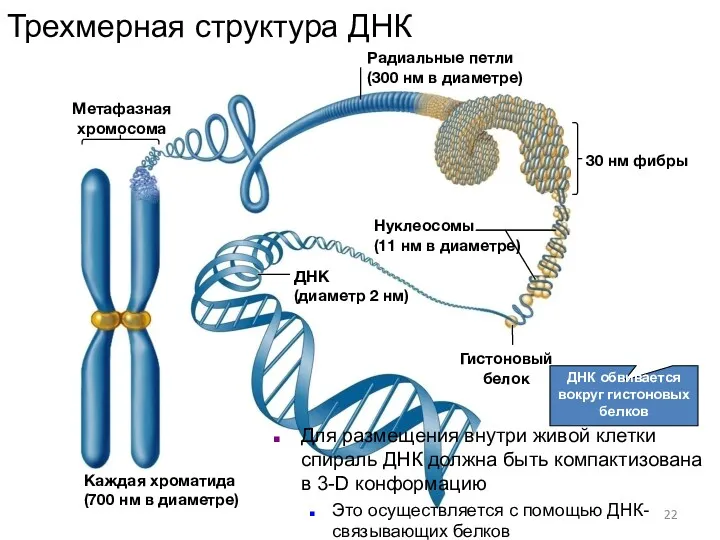
ДНК обвивается вокруг гистоновых белков
Радиальные петли
(300 нм в диаметре)
Метафазная хромосома
ДНК
(диаметр 2
ДНК обвивается вокруг гистоновых белков
Радиальные петли
(300 нм в диаметре)
Метафазная хромосома
ДНК
(диаметр 2

3 метра ДНК/клетка
1013 клеток/человек
3 x 1013 метров ДНК/человек
3.8 x 108 метров
3 метра ДНК/клетка
1013 клеток/человек
3 x 1013 метров ДНК/человек
3.8 x 108 метров

РЕПЛИКАЦИЯ ДНК
Когда Уотсон и Крик предложили модель двойной спирали ДНК,
РЕПЛИКАЦИЯ ДНК
Когда Уотсон и Крик предложили модель двойной спирали ДНК,
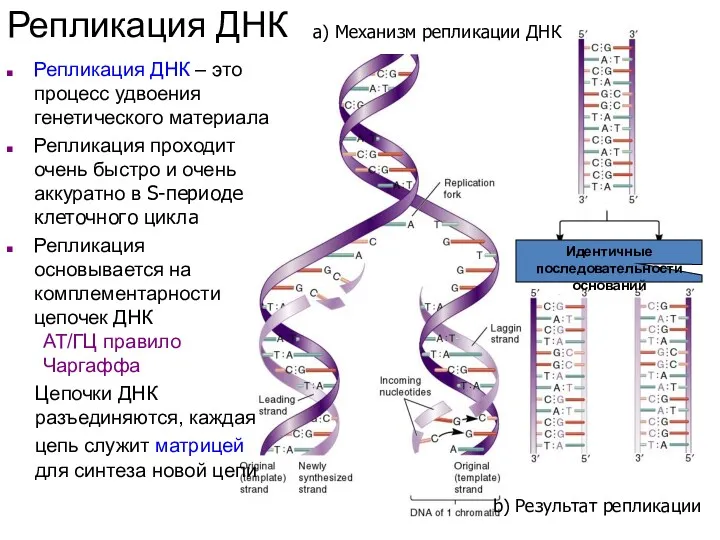
а) Механизм репликации ДНК
b) Результат репликации
Репликация ДНК
Репликация ДНК – это процесс
а) Механизм репликации ДНК
b) Результат репликации
Репликация ДНК
Репликация ДНК – это процесс


Уотсон и Крик предложили следующее:
Поскольку ДНК содержит две нити, две цепи
Уотсон и Крик предложили следующее:
Поскольку ДНК содержит две нити, две цепи
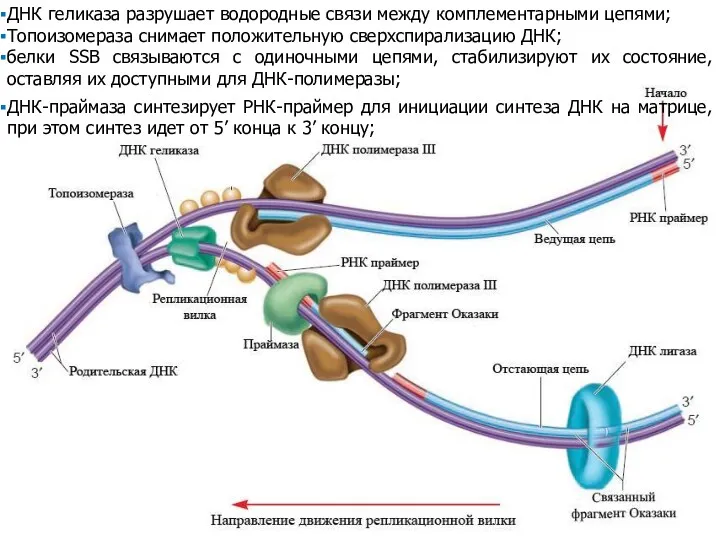
ДНК геликаза разрушает водородные связи между комплементарными цепями;
Топоизомераза снимает положительную сверхспирализацию
ДНК геликаза разрушает водородные связи между комплементарными цепями;
Топоизомераза снимает положительную сверхспирализацию
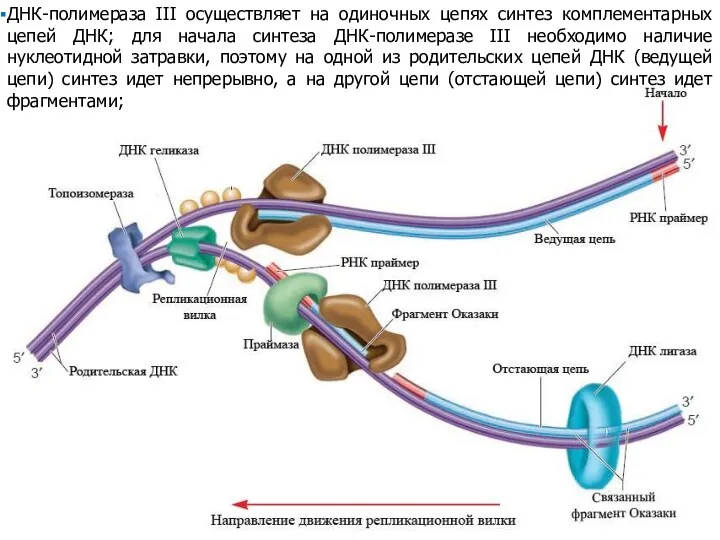
ДНК-полимераза III осуществляет на одиночных цепях синтез комплементарных цепей ДНК; для
ДНК-полимераза III осуществляет на одиночных цепях синтез комплементарных цепей ДНК; для
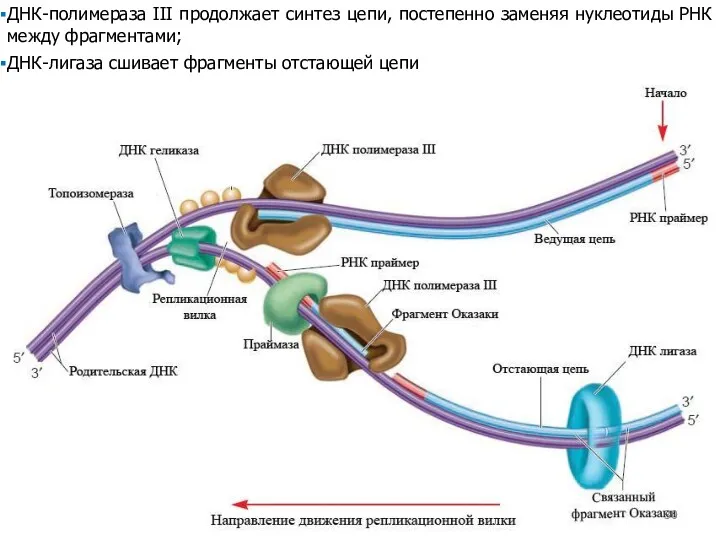
ДНК-полимераза III продолжает синтез цепи, постепенно заменяя нуклеотиды РНК между фрагментами;
ДНК-лигаза
ДНК-полимераза III продолжает синтез цепи, постепенно заменяя нуклеотиды РНК между фрагментами;
ДНК-лигаза
Аденин (А)
Гуанин (Г)
Урацил (У)
Основания
Остов
Цитозинe (Ц)
O
H
H
H
H
O
O
O
O
–
P
CH2
O
–
H
H
H
H
O
O
O
P
CH2
O
–
NH2
H
H
H
H
H
O
O
O
O
P
CH2
O
–
H
H
H
H
OH
H
H
O
O
O
O
P
CH2
O
–
Сахар (рибоза)
Фосфат
5′
4′
1′
2′
3′
5′
4′
1′
2′
3′
5′
4′
1′
2′
3′
5′
4′
1′
2′
3′
OH
OH
OH
OH
Нуклеотид РНК
Фосфодиэфирная связь
3′
5′
NH2
O
H
H
H
O
N
H
O
N
NH2
H
H
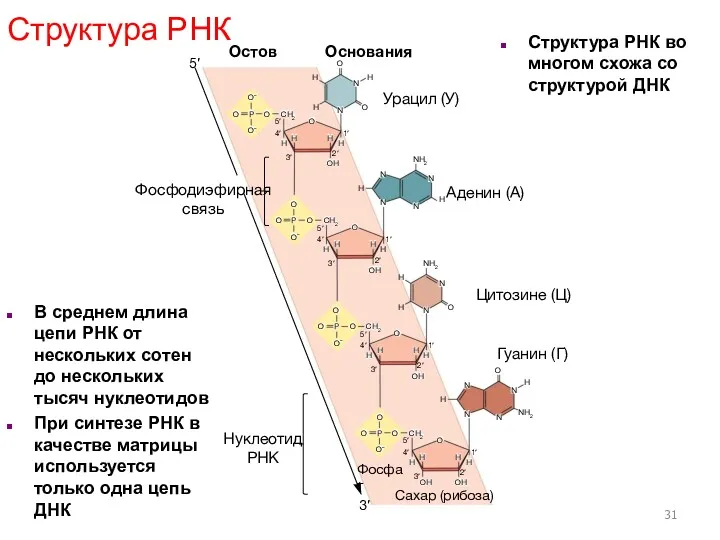
В среднем длина цепи
Аденин (А)
Гуанин (Г)
Урацил (У)
Основания
Остов
Цитозинe (Ц)
O
H
H
H
H
O
O
O
O
–
P
CH2
O
–
H
H
H
H
O
O
O
P
CH2
O
–
NH2
H
H
H
H
H
O
O
O
O
P
CH2
O
–
H
H
H
H
OH
H
H
O
O
O
O
P
CH2
O
–
Сахар (рибоза)
Фосфат
5′
4′
1′
2′
3′
5′
4′
1′
2′
3′
5′
4′
1′
2′
3′
5′
4′
1′
2′
3′
OH
OH
OH
OH
Нуклеотид РНК
Фосфодиэфирная связь
3′
5′
NH2
O
H
H
H
O
N
H
O
N
NH2
H
H
В среднем длина цепи

Copyright ©The McGraw-Hill Companies, Inc. Permission required for reproduction or display
Обычно
Copyright ©The McGraw-Hill Companies, Inc. Permission required for reproduction or display
Обычно
A
U
A
U
U
A
G
C
C
G
C
G
A
U
U
A
U
A
C
G
C
G
C
G
C
G
C
G
A
A
U
U
G
G
C
C
C
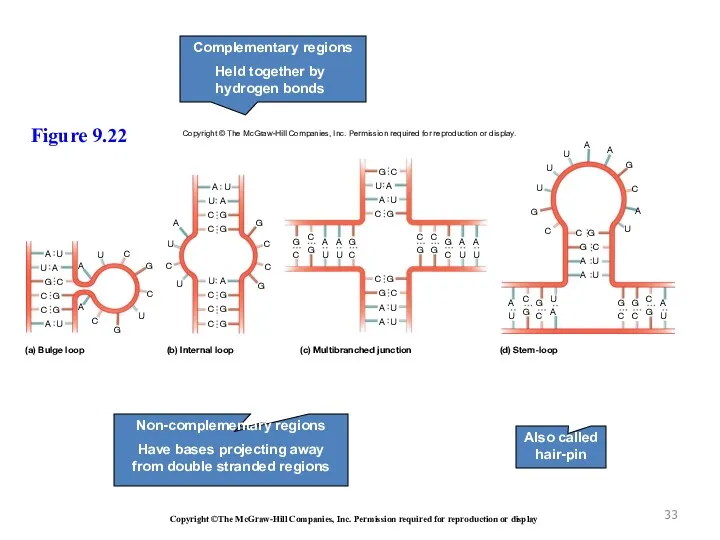
(a) Bulge loop
(b) Internal loop
(c) Multibranched junction
(d) Stem-loop
G
C
C
G
U
A
A
U
G
C
G
C
C
G
A
U
A
U
A
U
G
C
A
U
U
A
G
C
C
G
C
G
G
C
G
C
C
G
A
U
A
U
G
C
C
G
C
G
A
U
A
U
A
U
U
G
G
C
C
C
A
U
A
U
G
C
C
G
A
A
A
U
U
U
U
G
G
C
C
Copyright © The McGraw-Hill
A
U
A
U
U
A
G
C
C
G
C
G
A
U
U
A
U
A
C
G
C
G
C
G
C
G
C
G
A
A
U
U
G
G
C
C
C
(a) Bulge loop
(b) Internal loop
(c) Multibranched junction
(d) Stem-loop
G
C
C
G
U
A
A
U
G
C
G
C
C
G
A
U
A
U
A
U
G
C
A
U
U
A
G
C
C
G
C
G
G
C
G
C
C
G
A
U
A
U
G
C
C
G
C
G
A
U
A
U
A
U
U
G
G
C
C
C
A
U
A
U
G
C
C
G
A
A
A
U
U
U
U
G
G
C
C
Copyright © The McGraw-Hill
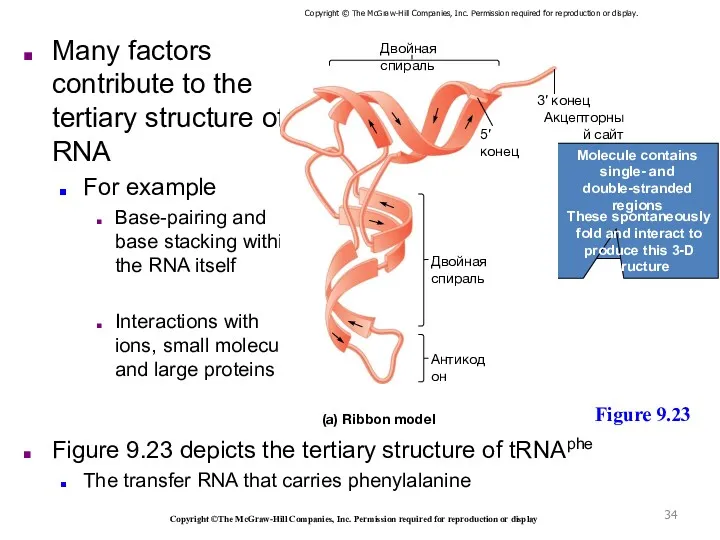
Many factors contribute to the tertiary structure of RNA
For example
Base-pairing and
Many factors contribute to the tertiary structure of RNA
For example
Base-pairing and

Транскрипция гена
Транскрипция гена

ВВЕДЕНИЕ
ДНК хранит информацию
Для построения механизма (организма) обеспечивающего
Не является самодостаточной молекулой
Для
ВВЕДЕНИЕ
ДНК хранит информацию
Для построения механизма (организма) обеспечивающего
Не является самодостаточной молекулой
Для
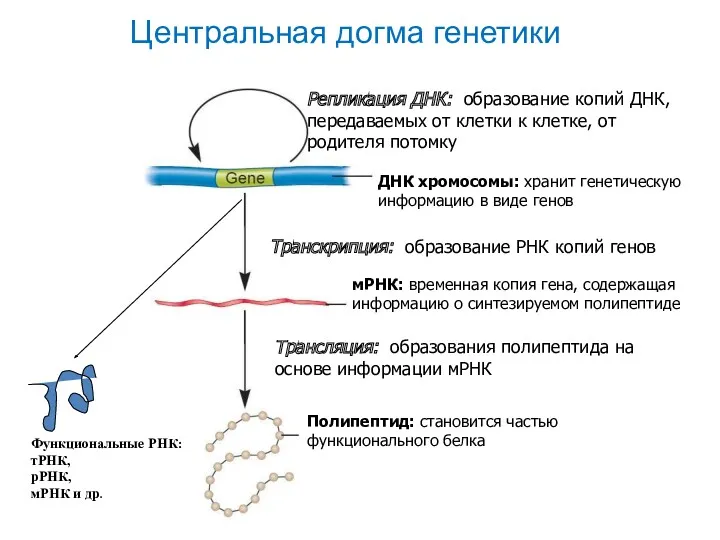
Центральная догма генетики
Центральная догма генетики
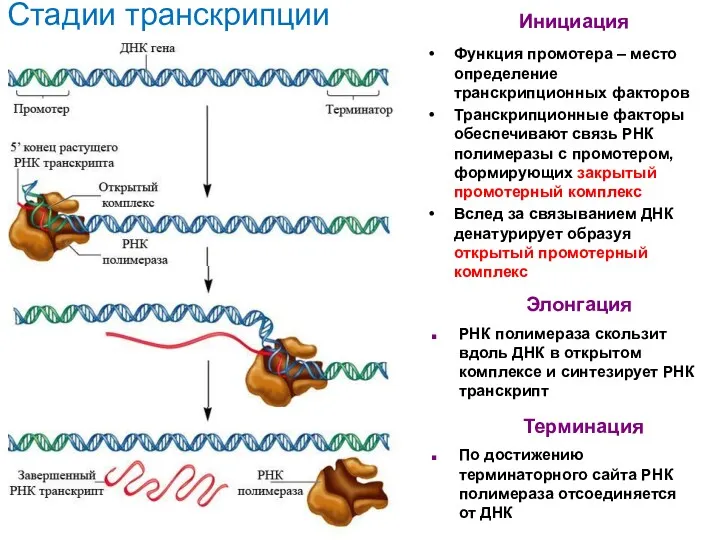
Функция промотера – место определение транскрипционных факторов
Транскрипционные факторы обеспечивают связь
Функция промотера – место определение транскрипционных факторов
Транскрипционные факторы обеспечивают связь

Ген может быть определен как дискретный участок ДНК транскрибируемый в РНК
Ген
Ген может быть определен как дискретный участок ДНК транскрибируемый в РНК
Ген
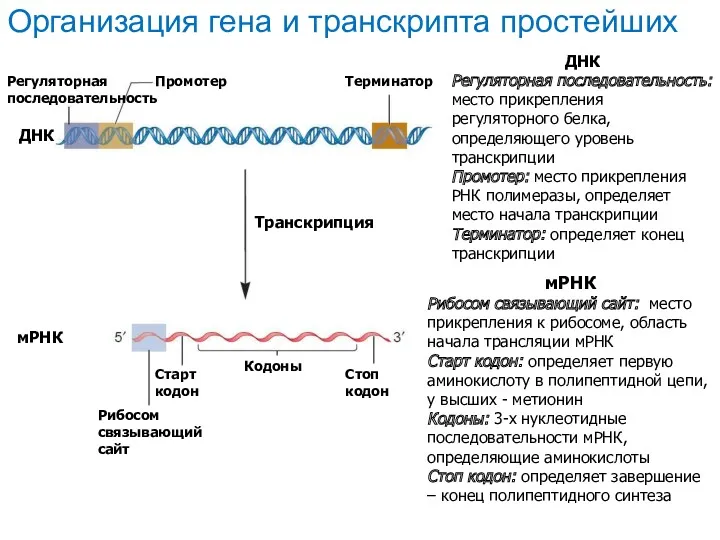
ДНК
Регуляторная последовательность: место прикрепления регуляторного белка, определяющего уровень транскрипции
Промотер: место
ДНК
Регуляторная последовательность: место прикрепления регуляторного белка, определяющего уровень транскрипции
Промотер: место
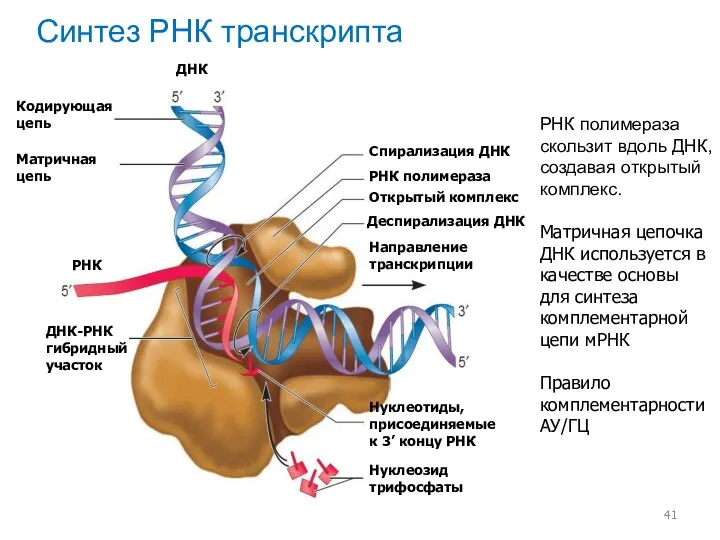
Синтез РНК транскрипта
РНК полимераза скользит вдоль ДНК, создавая открытый комплекс.
Матричная цепочка
Синтез РНК транскрипта
РНК полимераза скользит вдоль ДНК, создавая открытый комплекс.
Матричная цепочка

Полипептидный синтез
Процесс синтеза белка на матричной мРНК называется трансляцией, его основные
Полипептидный синтез
Процесс синтеза белка на матричной мРНК называется трансляцией, его основные
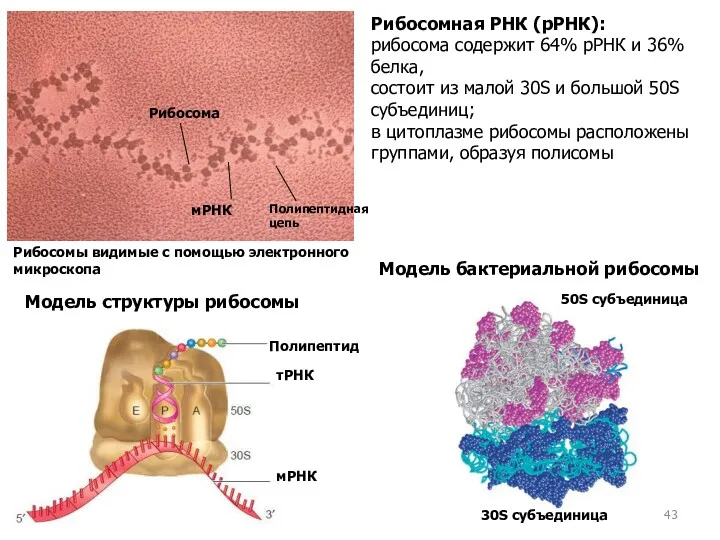
Модель бактериальной рибосомы
30S субъединица
50S субъединица
Рибосомная РНК (рРНК):
рибосома содержит 64% рРНК
Модель бактериальной рибосомы
30S субъединица
50S субъединица
Рибосомная РНК (рРНК):
рибосома содержит 64% рРНК
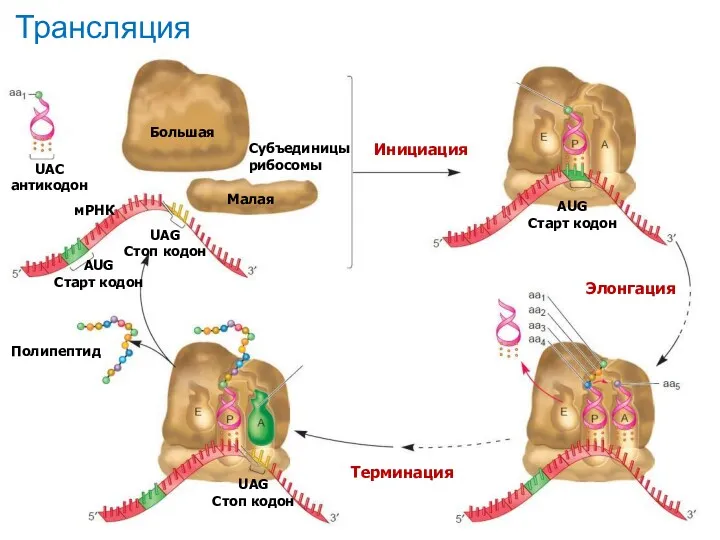
Трансляция
Субъединицы рибосомы
Большая
Малая
UAC
антикодон
мРНК
AUG
Старт кодон
UAG
Стоп кодон
Инициация
AUG
Старт кодон
Элонгация
Терминация
UAG
Стоп кодон
Полипептид
Трансляция
Субъединицы рибосомы
Большая
Малая
UAC
антикодон
мРНК
AUG
Старт кодон
UAG
Стоп кодон
Инициация
AUG
Старт кодон
Элонгация
Терминация
UAG
Стоп кодон
Полипептид
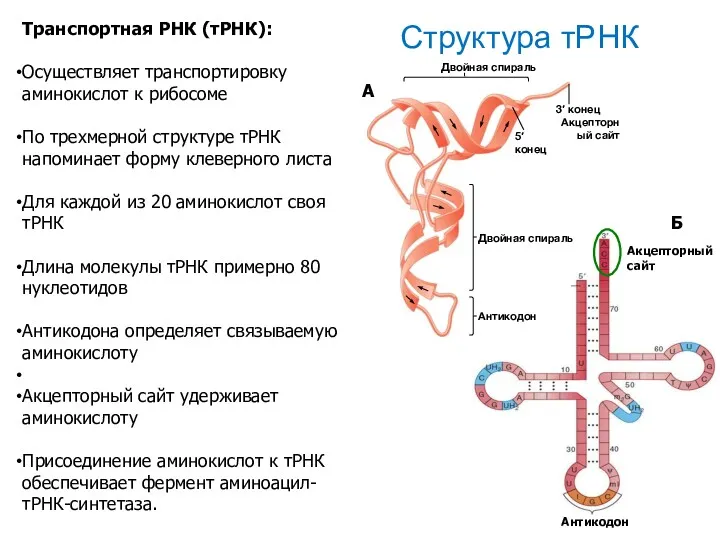
Структура тРНК
Транспортная РНК (тРНК):
Осуществляет транспортировку аминокислот к рибосоме
По трехмерной
Структура тРНК
Транспортная РНК (тРНК):
Осуществляет транспортировку аминокислот к рибосоме
По трехмерной
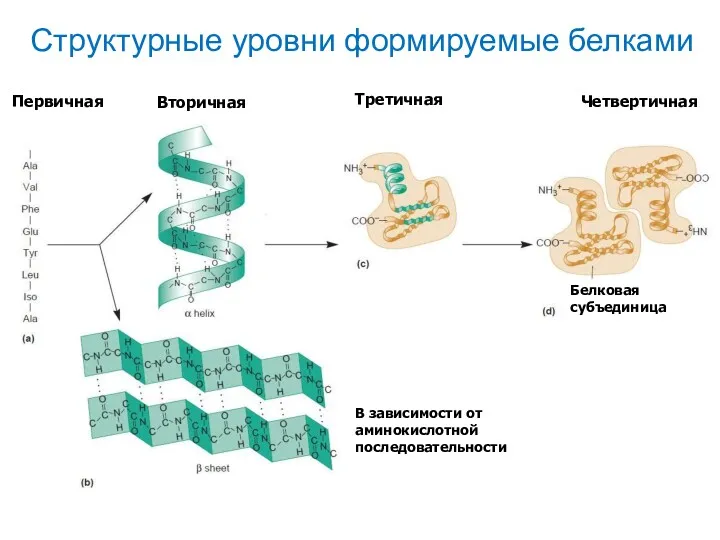
Структурные уровни формируемые белками
В зависимости от аминокислотной последовательности
Белковая субъединица
Структурные уровни формируемые белками
В зависимости от аминокислотной последовательности
Белковая субъединица
 Царства живой природы
Царства живой природы Работа и сила мышц. Строение сократительного аппарата поперечно-полосатой мышечной ткани
Работа и сила мышц. Строение сократительного аппарата поперечно-полосатой мышечной ткани Органы дыхания
Органы дыхания Гены и аллели. Аллельные взаимодействия
Гены и аллели. Аллельные взаимодействия Dinoflagellates
Dinoflagellates Торговий Дім АКТИВ-HARVEST. Добрива для позакореневого живлення
Торговий Дім АКТИВ-HARVEST. Добрива для позакореневого живлення Отряд Стрекозы
Отряд Стрекозы Рефлекторный принцип деятельности ЦНС. Возбуждение и торможение. Функции нейронов и нейроглии. Физиология рецепторов
Рефлекторный принцип деятельности ЦНС. Возбуждение и торможение. Функции нейронов и нейроглии. Физиология рецепторов Создание, исследование и испытание доступного биовегетария
Создание, исследование и испытание доступного биовегетария Асимметрия полушарий головного мозга человека
Асимметрия полушарий головного мозга человека Закон Менделя
Закон Менделя презентация к уроку Органические молекулы - углеводы
презентация к уроку Органические молекулы - углеводы Составление тренировочных программ для мужчин различных типов сложения
Составление тренировочных программ для мужчин различных типов сложения Репчатый лук
Репчатый лук Презентация Организация исследовательской работы
Презентация Организация исследовательской работы Презентация к уроку Эволюция строения и функций органов и их систем
Презентация к уроку Эволюция строения и функций органов и их систем Опорно-двигательная система человека
Опорно-двигательная система человека Репродукція клітин
Репродукція клітин Protein Structure and Function
Protein Structure and Function Инструменты для груминга
Инструменты для груминга klasss
klasss Подцарство одноклеточные
Подцарство одноклеточные Осьминоги. Головоногие моллюски
Осьминоги. Головоногие моллюски Особенности размножения рыб
Особенности размножения рыб Урок-игра Кровь и кровообращение
Урок-игра Кровь и кровообращение Для начинающих улитководов
Для начинающих улитководов Класс Земноводные, или Амфибии (Amphibia)
Класс Земноводные, или Амфибии (Amphibia) Вид. Критерии вида
Вид. Критерии вида