- Водно-солевой обмен у водных организмов

Содержание
- 2. По степени солености водоемы условно подразделяются на пресные с соленостью менее 0,5 ‰, солоноватоводные – соленость
- 3. У большинства морских обитателей концентрация солей в организме близка к морской воде – такие живые организмы
- 5. У большинства беспозвоночных наблюдается некоторое повышение осмотического давления внутренней среды организма (гипертоничность), это обеспечивает постоянный приток
- 6. Механизмы осморегуляции: Многие водные организмы, живущие в гипертрофированной среде (морская вода), теряют воду путем осмоса и
- 8. Среди пресноводных нет изотоничных форм, концентрация жидкости в их клетках и тканях выше, чем в окружающей
- 9. Механизмом поддержания постоянства осмотического давления является активное выделение избытка воды через почки. Вода поступает в организм
- 10. Потери солей с мочой и эскрементами компенсируются активным переносом ионов из окружающей среды против градиента концентрации.
- 11. 1 Костные рыбы Задачи осморегуляции в море обратны пресноводному типу: в море концентрация солей несколько выше,
- 12. 1 Костные рыбы Реабсорбция ионов в почечных канальцах морских костных рыб резко снижена, но зато здесь
- 13. Осморегуляция в море 2 Хрящевые рыбы Концентрация солей в крови хрящевых рыб, также как и у
- 14. Осморегуляция в море 2 Хрящевые рыбы Во-вторых, в крови хрящевых рыб накапливается триметиламиноксид (ТМАО), обладающий высокой
- 15. Осморегуляция и жизненный цикл Чаще всего, даже если система осморегуляции животного развита хорошо, это не относится
- 16. Как уже можно догадаться, практически у всех животных осморегуляция и выделение продуктов собственного обмена веществ не
- 17. Это не помешало им приспособиться как к морским, так и к пресным водам, но эвригалинных форм
- 18. Губки и кишечнополостные. Не имеют специальных систем выделения, кроме тех, которые создают ток воды через их
- 19. Плоские и круглые черви. Снабжены протонефридиями – многоклеточными аналогами сократительных вакуолей, накапливающими межклеточную жидкость и выпрыскивающими
- 20. Кольчатые черви. Имеют метанефридии, в которые поступает жидкость из целомической полости, и имеющие систему дополнительной закачки
- 21. Моллюски. Имеют специальный орган (почку) для накопления полостной жидкости и вторичной ее трансформации (закачки или откачки
- 22. Иглокожие. Не имеют никаких систем осморегуляции и не способны адаптироваться к изменениям солености. Более того –
- 23. Ракообразные. Обычно имеют свой аналог почек – так называемые антеннальные, или зеленые железы, открывающеся на голове.
- 24. Насекомые. Поскольку возникли на суше, изначально имеют мощные системы экономии влаги – плотные наружние покровы (почти
- 25. Рыбы. Также имеют хорошо развитые почки, осуществляющие активный транспорт ионов в нужном направлении и обеспечивающие эффективную
- 33. Скачать презентацию

По степени солености водоемы условно подразделяются на пресные с соленостью менее
По степени солености водоемы условно подразделяются на пресные с соленостью менее

У большинства морских обитателей концентрация солей в организме близка к морской
У большинства морских обитателей концентрация солей в организме близка к морской
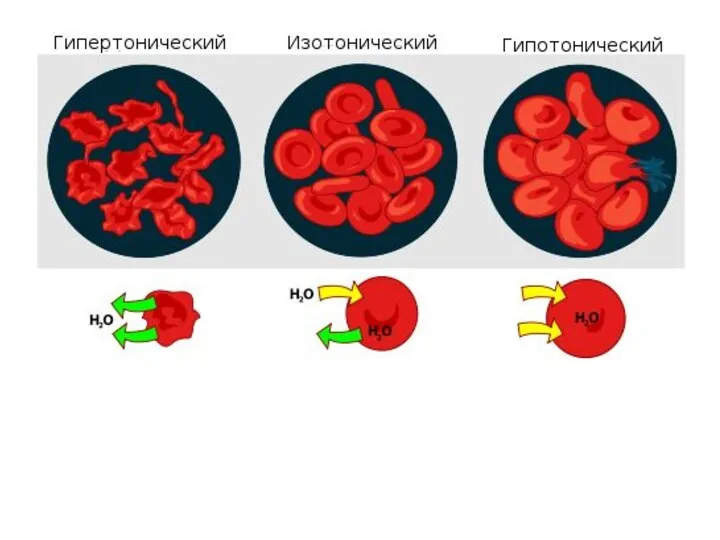
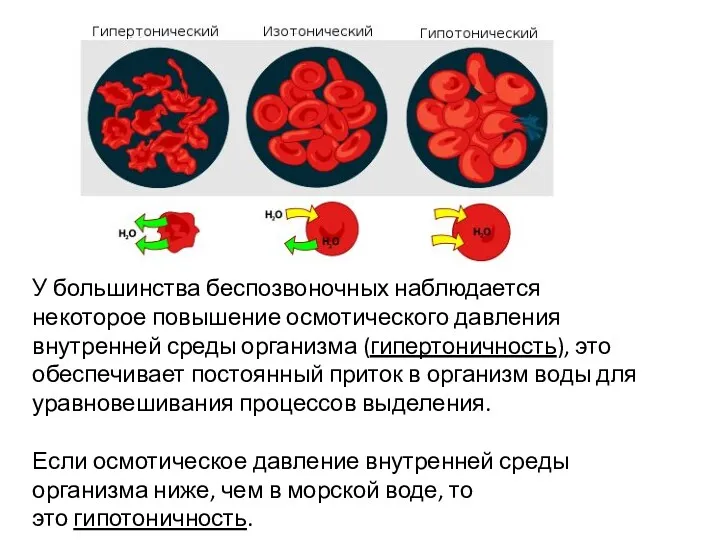
У большинства беспозвоночных наблюдается некоторое повышение осмотического давления внутренней среды организма
У большинства беспозвоночных наблюдается некоторое повышение осмотического давления внутренней среды организма

Механизмы осморегуляции:
Многие водные организмы, живущие в гипертрофированной среде (морская вода), теряют
Механизмы осморегуляции:
Многие водные организмы, живущие в гипертрофированной среде (морская вода), теряют
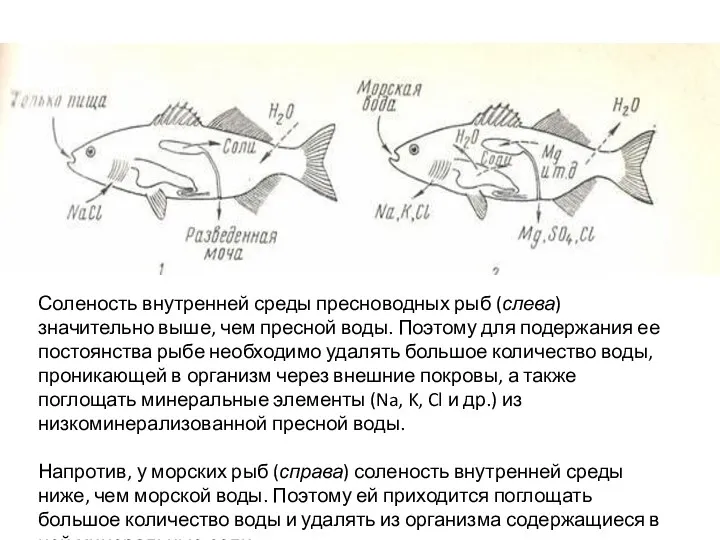
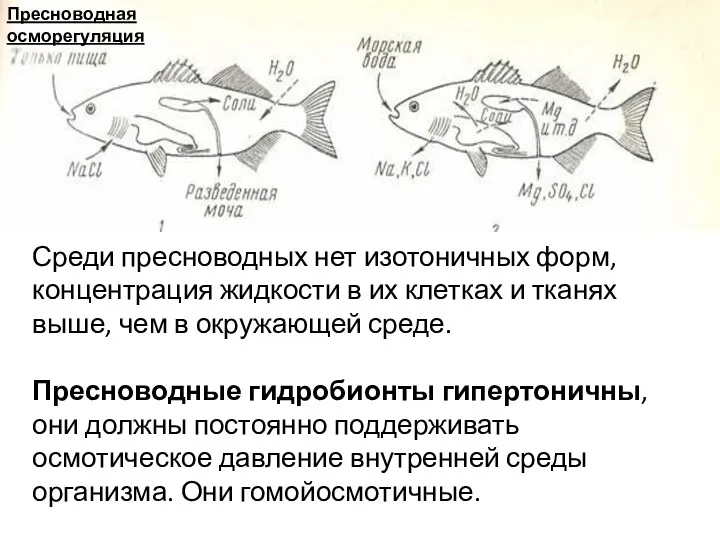
Среди пресноводных нет изотоничных форм, концентрация жидкости в их клетках и
Среди пресноводных нет изотоничных форм, концентрация жидкости в их клетках и
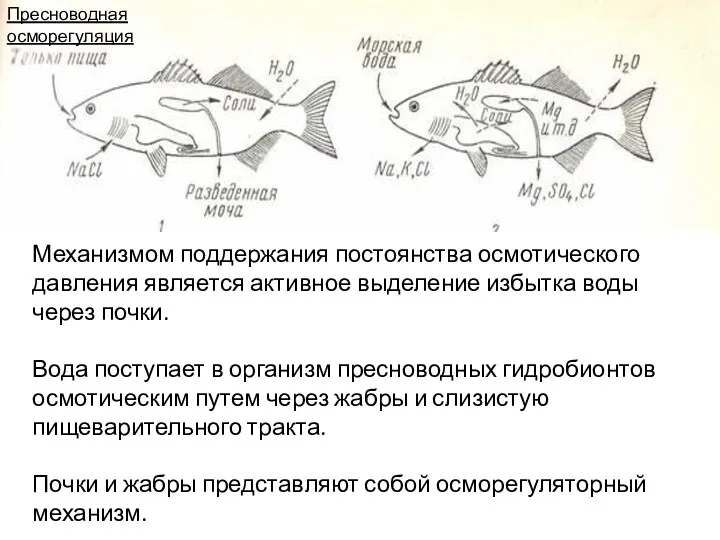
Механизмом поддержания постоянства осмотического давления является активное выделение избытка воды через
Механизмом поддержания постоянства осмотического давления является активное выделение избытка воды через
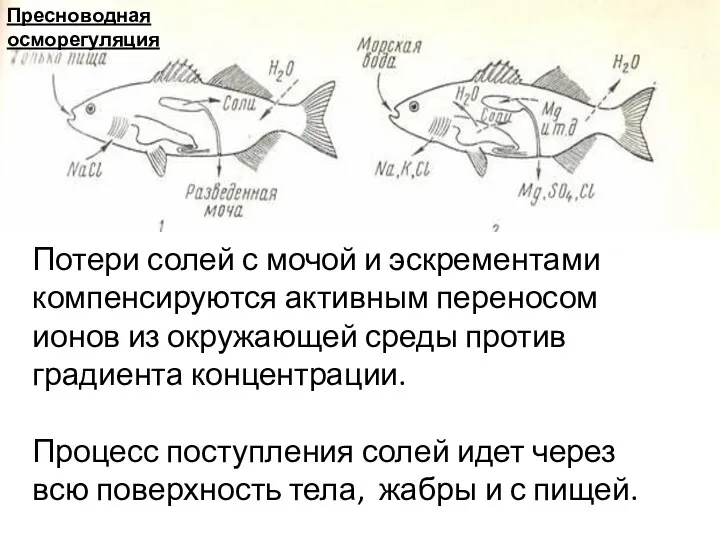
Потери солей с мочой и эскрементами компенсируются активным переносом ионов из
Потери солей с мочой и эскрементами компенсируются активным переносом ионов из
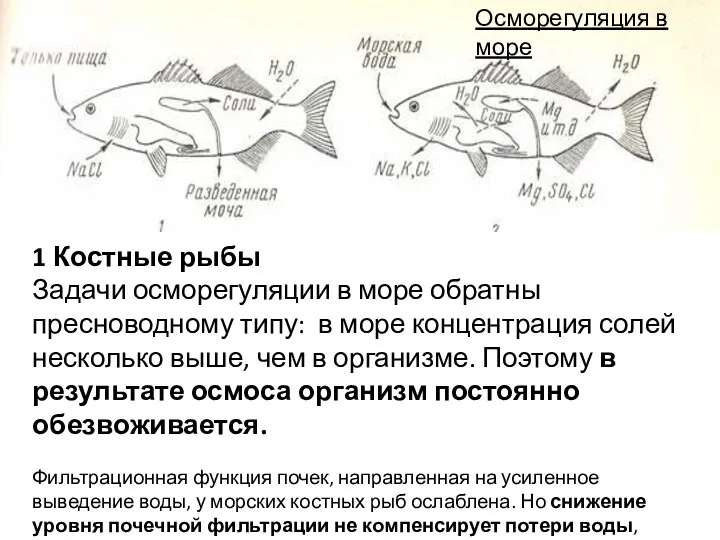
1 Костные рыбы
Задачи осморегуляции в море обратны пресноводному типу: в море
1 Костные рыбы
Задачи осморегуляции в море обратны пресноводному типу: в море
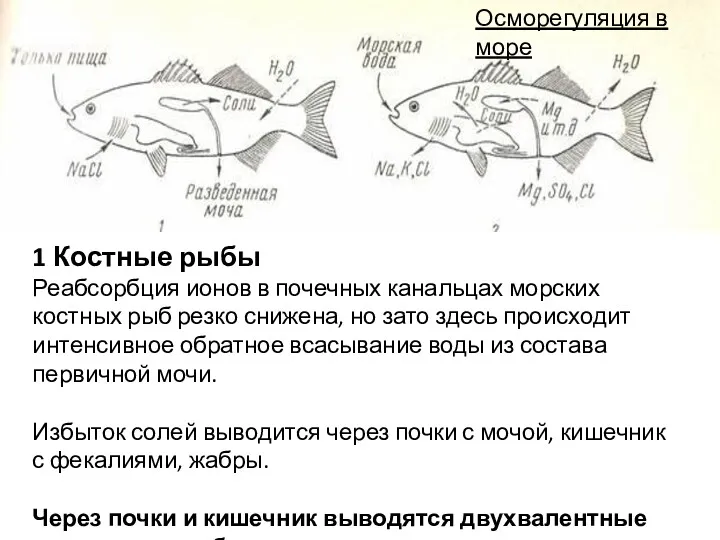
1 Костные рыбы
Реабсорбция ионов в почечных канальцах морских костных рыб резко
1 Костные рыбы
Реабсорбция ионов в почечных канальцах морских костных рыб резко
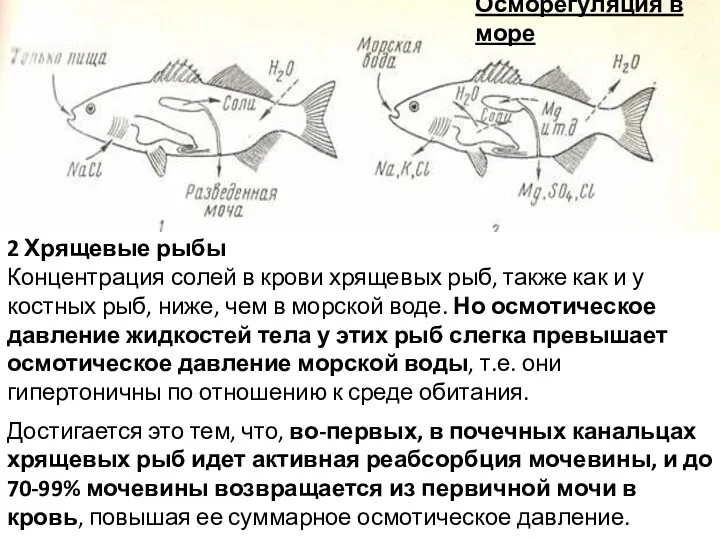
Осморегуляция в море
2 Хрящевые рыбы
Концентрация солей в крови хрящевых рыб, также
Осморегуляция в море
2 Хрящевые рыбы
Концентрация солей в крови хрящевых рыб, также
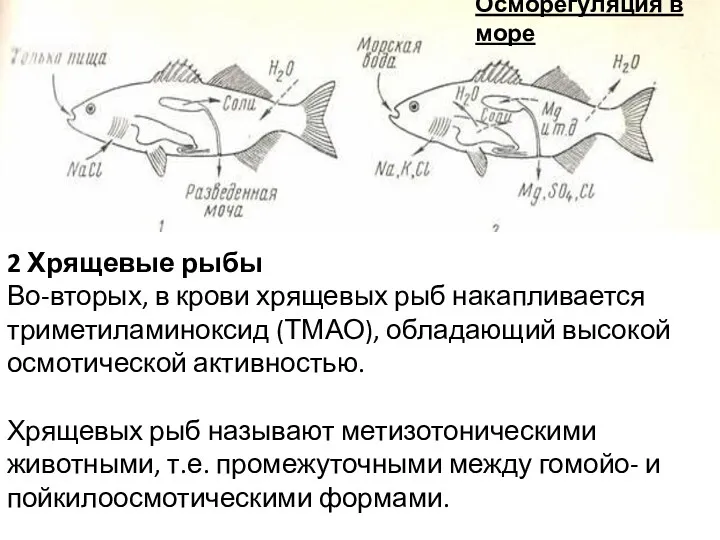
Осморегуляция в море
2 Хрящевые рыбы
Во-вторых, в крови хрящевых рыб накапливается триметиламиноксид
Осморегуляция в море
2 Хрящевые рыбы
Во-вторых, в крови хрящевых рыб накапливается триметиламиноксид

Осморегуляция и жизненный цикл
Чаще всего, даже если система осморегуляции животного развита
Осморегуляция и жизненный цикл
Чаще всего, даже если система осморегуляции животного развита

Как уже можно догадаться, практически у всех животных осморегуляция и выделение
Как уже можно догадаться, практически у всех животных осморегуляция и выделение
Это не помешало им приспособиться как к морским, так и к пресным водам,
Это не помешало им приспособиться как к морским, так и к пресным водам,
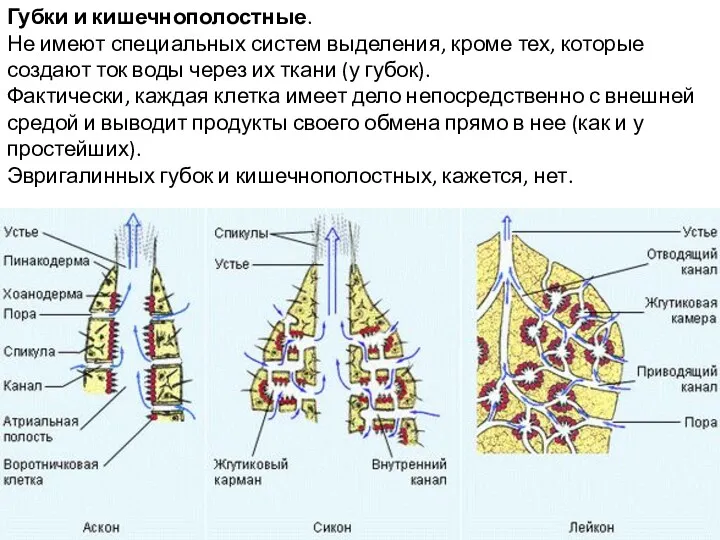
Губки и кишечнополостные.
Не имеют специальных систем выделения, кроме тех, которые
Губки и кишечнополостные.
Не имеют специальных систем выделения, кроме тех, которые
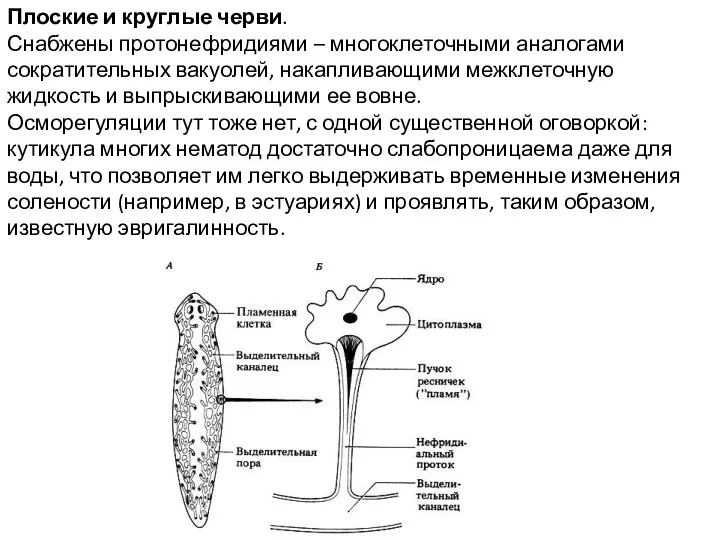
Плоские и круглые черви.
Снабжены протонефридиями – многоклеточными аналогами сократительных вакуолей,
Плоские и круглые черви.
Снабжены протонефридиями – многоклеточными аналогами сократительных вакуолей,

Кольчатые черви.
Имеют метанефридии, в которые поступает жидкость из целомической полости,
Кольчатые черви.
Имеют метанефридии, в которые поступает жидкость из целомической полости,

Моллюски.
Имеют специальный орган (почку) для накопления полостной жидкости и вторичной
Моллюски.
Имеют специальный орган (почку) для накопления полостной жидкости и вторичной

Иглокожие.
Не имеют никаких систем осморегуляции и не способны адаптироваться к
Иглокожие.
Не имеют никаких систем осморегуляции и не способны адаптироваться к

Ракообразные.
Обычно имеют свой аналог почек – так называемые антеннальные, или
Ракообразные.
Обычно имеют свой аналог почек – так называемые антеннальные, или

Насекомые.
Поскольку возникли на суше, изначально имеют мощные системы экономии влаги
Насекомые.
Поскольку возникли на суше, изначально имеют мощные системы экономии влаги

Рыбы.
Также имеют хорошо развитые почки, осуществляющие активный транспорт ионов в
Рыбы.
Также имеют хорошо развитые почки, осуществляющие активный транспорт ионов в





 Development
Development Фізіологія червоної крові. Еритрон. Дихальні пігменти
Фізіологія червоної крові. Еритрон. Дихальні пігменти Качественное и количественное определение витамина С в продуктах и оценка обеспеченности рациона школьника и студента витамином С
Качественное и количественное определение витамина С в продуктах и оценка обеспеченности рациона школьника и студента витамином С презентация по теме: Правда о пиве
презентация по теме: Правда о пиве Микрогаметофит. Созревание, покой и прорастание
Микрогаметофит. Созревание, покой и прорастание Внеклассное мероприятие День Зиновия Синичника
Внеклассное мероприятие День Зиновия Синичника Клетки – маленькие лаборатории
Клетки – маленькие лаборатории Разнообразие пород гусей
Разнообразие пород гусей Обмен белков: Индивидуальные пути обмена аминокислот
Обмен белков: Индивидуальные пути обмена аминокислот ткани человека
ткани человека Микрофлора организма человека
Микрофлора организма человека Сравнительный анализ проращивания семян горчицы в разных образцах талой воды и дистиллированной воды
Сравнительный анализ проращивания семян горчицы в разных образцах талой воды и дистиллированной воды Сущность жизни и свойства живого
Сущность жизни и свойства живого Подцарство одноклеточные
Подцарство одноклеточные Отряд Черепахи
Отряд Черепахи Фауна. Животные пойменных лугов
Фауна. Животные пойменных лугов Класс двудольные. Семейство розоцветные
Класс двудольные. Семейство розоцветные Безопасные ягоды и грибы
Безопасные ягоды и грибы Тестовыя работа по теме: Вирусы
Тестовыя работа по теме: Вирусы Доказательства эволюции органического мира
Доказательства эволюции органического мира Генная инженерия бактерий
Генная инженерия бактерий Цветы и растения, как символы стран мира
Цветы и растения, как символы стран мира Формирование исследовательских умений и навыков на уроках биологии и химии и во внеурочное время
Формирование исследовательских умений и навыков на уроках биологии и химии и во внеурочное время Физиология центральной нервной системы
Физиология центральной нервной системы Общая характеристика типа Моллюски
Общая характеристика типа Моллюски Белки как биополимеры, их состав, строение и функции в клетке
Белки как биополимеры, их состав, строение и функции в клетке Лишайники, мхи
Лишайники, мхи Методы антропогенетики
Методы антропогенетики