- Закон Дриша: проспективные потенции области зародыша

Содержание
- 2. ОБРАЗОВАНИЕ И ДИФФЕРЕНЦИРОВКА ПЛАЗМОДИЯ У Dyctiostelium Жизненный состоит из чередования одноклеточной (одиночные амебы) и многоклеточной (плазмодий,
- 3. Классический опыт Дриша – изоляция бластомеров зародышей морского ежа после первого деления дробления с полной регуляцией
- 4. Из каждой сагиттальной половины бластулы Xenopus развивается нормальный пропорционально уменьшенный головастик с более высокой внутри-индивидуальной изменчивостью,
- 5. ОПЫТ ВИЛЬГЕЛЬМА РУ Обычно считается, что из живого бластомера развивается половина зародыша. Это неверно: живая половина
- 6. Считается, что закон Дриша относится к явлениям эмбриональной регуляции. В действительности же между регуляцией и изменчивостью
- 7. У гидроида Dynamena дробление не упорядочено, нет бластулы, и AP полярность планулы возникает независимо от AV
- 8. В НОРМАЛЬНОЙ ГАСТРУЛЕ XENOPUS ОБЛАСТИ С ЕДИНЫМ ПРОСПЕКТИВНЫМ ЗНАЧЕНИЕМ «СМЕШИВАЮТСЯ» ИЗ КЛЕТОК РАЗНЫХ ЗОН БЛАСТУЛЫ К
- 9. Обычные карты презумптивных закладок достоверны только при условии, что клетки, окружающие меченый участок, движутся точно так
- 10. Биллиард Гальтона (Galton’s board): модель, показывающая как формируется нормальное распределение: «детерминированный хаос» Для возникновения изменчивости достаточно
- 11. Судьба индивидуальных клеток зародышевого щитка у zebrafish: пунктир – границы щитка (A – передний, P –
- 12. НОРМАЛЬНОЕ РАСПРЕДЕЛЕНИЕ
- 13. МОДА ЧАСТОТНОГО РАСПРЕДЕЛЕНИЯ – МАКРОСОСТОЯНИЕ С НАИБОЛЬШИМ СТАТИСТИЧЕСКИМ ВЕСОМ Нормальное распределение макроскопического признака (размер особи) не
- 14. ОТКЛОНЕНИЕ ОТ НОРМАЛЬНОСТИ ОЗНАЧАЕТ КАУЗАЛЬНУЮ СВЯЗЬ МАКРО И МИКРОСОСТОЯНИЙ (+) эксцесс означает положительную (стабилизирующую), (-) эксцесс
- 15. ФЛЮКТУИРУЮЩАЯ АСИММЕТРИЯ БИЛАТЕРАЛЬНО СИММЕТРИЧНЫХ СТРУКТУР Ротовой диск головастиков состоит из билатерально симметричных рядов роговых зубчиков (например,
- 16. Закон Дриша в нормальном развитии – это нормальная изменчивость, вытекающая из относительной независимости судьбы от происхождения
- 17. Закон Дриша выполняется и при наличии, и при отсутствии клонирования клеток. Если клонирования нет, то каждая
- 18. Клонирование – это когда границы первичных эмбриональных закладок совпадают с границами бластомеров. У разных зародышей клон
- 19. Анимальные и вегетативные бластомеры приходят в контакт из-за редукции бластоцеля: изменчивость резко возрастает из-за конкуренции макромеров
- 20. При гетероквадрантном спиральном дроблении мезодермальным становится самый крупный макромер, так как он, из-за своих размеров, смещен
- 21. Эмбриональное развитие дрозофилы (ядра помечены флуоресцентной меткой
- 22. У высших насекомых (дрозофила), развитие которых считается детерминированным, случайный выбор смещается из эмбриогенеза в оогенез –
- 23. Два подхода к описанию развития: координатный (позициональная информация) и геометрический (геометрия движения клеток)
- 24. Позициональная информация дает объяснение дифференцировки осевых структур с помощью градиентов экспрессии генов в заранее заданной системе
- 25. Хотя термин «позициональная информация» продолжают использовать, «градиенты» концентрации «морфогенов» оказались иллюзией BMP – белки, поддерживающие «вентральное»
- 26. «Вентральное» состояние эктодермы (эпидермис + мезенхима) не просто существует, а активно поддерживается за счет замкнутой петли
- 27. Вместо «градиентов морфогенов» - ФИЗИОЛОГИЧЕСКИЕ ГРАДИЕНТЫ Чайлда, НЕ являющиеся градиентами каких-либо полей. Это параметрические градиенты, делающими
- 28. В нормальном развитии один и тот элемент организации может подстроиться к окружению, или подстроить его под
- 29. Меченый участок, занимающий латеральное положение, может сохранить его (А), или же оказаться в медианной зоне (Б)
- 30. Морфометрия, топология и морфогенез дорсальной губы бластопора гаструлы (ДГБ или DBL), образующей арку над желточной энтодермой
- 31. При количественной оценке изменчивость формы и размеров арки ДГБ (значения коэффициентов вариации количественных морфологических признаков) всегда
- 33. Масштабы изменчивости развития в генетически однородной выборке зародышей, развивающихся в оптимальных условиях, показывают широту его нормы
- 34. Компоненты коллективного движения наружного слоя клеток в гаструляции лягушки, доступные для измерения (I -III): Компоненты I-III
- 35. Индивидуальные траектории гаструляции лягушки (траектории изменения признака у отдельных зародышей) образуют «детерминированный хаос»: анализировать можно только
- 36. Графы корреляций между последовательными измерениями признаков ДГБ у индивидуальных зародышей Индивидуальные траектории развития неустойчивы (отсутствуют) в
- 37. Основная закономерность: изменчивость признаков максимальна в начале образования структуры и уменьшается по мере ее формирования (по
- 38. Второе правило: скорость направленного изменения признаков положительно связана с величиной их изменчивости (положительная связь удельной скорости
- 39. Такие же правила соблюдаются и в морфогенезе сериально гомологичных (повторяющихся) структур A – ротовой диск головастиков
- 40. Связь между удельной скоростью роста и изменчивостью для последовательных рядов зубчиков в ранней (красный цвет) и
- 41. Гистологический анализ показывает, что дополнительный ряд, онтогенетически самый поздний, развивается так, как должны были развиваться эволюционно
- 42. Беспорядочное распределение фаллоидина (А) упорядочивается по мере формирования борозды гастрального впячивания (Б) А Б То же
- 43. Аналогичным образом формируются зоны экспрессии генов, например, гена brachyury А Б В Стрелка – дорсальный полюс
- 44. Изменчивость связана с гетерохрониями между эпиболией и изменением формы дорсальной губы бластопора (ДГБ): при эпиболии ДГБ
- 45. Пунктирные стрелки – вход и выход клеток из переходной зоны ДГБ, наконечник – движение дорсального края
- 46. Осцилляции средних значений и дисперсии кривизны арки ДГБ в гаструляции лягушки достоверны в начальной фазе образования
- 47. Динамика и причины региональных различий в масштабах изменчивости Изменчивость радиуса более «молодого» латерального края ДГБ максимальна,
- 48. Латеральный край ДГБ и ВГБ не формируют главную ось тела только из-за того, что клетки краевой
- 49. СХОДСТВО ДИНАМИКИ ИЗМЕНЧИВОСТИ С ОТБОРОМ: при нормальном ходе гаструляции ДГБ состоит из фрагментов разной кривизны и
- 50. Кривизна губы бластопора (утолщенная стрелка) вытесняет исходную кривизну окружности краевой зоны (тонкие стрелки) подобно тому, как
- 51. ФУНДАМЕНТАЛЬНАЯ ТЕОРЕМА ЕСТЕСТВЕННОГО ОТБОРА (ТЕОРЕМА ФИШЕРА): простейшая (гаплоидная) модель Отбор максимизирует среднюю приспособленность W* со скоростью,
- 52. Центры двух окружностей с радиусами RL и RD сближаются со скоростью, равной величине их дисперсии (RL
- 53. Нормальная изменчивость хода эпиболии вьюна в синхронной фиксации зародышей одной кладки икры с сильной десинхронизацией развития:
- 54. В раннем морфогенезе вьюна зоны экспрессии генов (размер, форма расположение) изменчивы не меньше, чем морфология Изменчивость
- 55. Изменчивость зональной экспрессии генов подчиняется основному правилу – она максимальна при появлении зоны экспрессии данного гена
- 56. Последовательные стадии гаструляции вьюна: осцилляции, основанные на чередовании входа и выхода клеток из ЗК в области
- 57. Зона формирования ЗЩ может соответствовать утолщенной или более плотной части ЗК. В первом случае ЗЩ формируется
- 58. НОРМАЛЬНАЯ ИЗМЕНЧИВОСТЬ НАЧАЛА ГАСТРУЛЯЦИИ (ЗАРОДЫШИ РАЗНЫХ КЛАДОК) Гетерохронии между входом и выходом клеток из краевого валика
- 60. При факторном анализе нормальной изменчивости практически вся дисперсия количественных признаков распределяется между небольшим числом (не более
- 61. Корреляционные графы, связывающие значения количественных признаков на индивидуальных траекториях развития (в сериях последовательных измерений живых зародышей),
- 62. Морфогенетические корреляции (МК) – это прямая и обратная связь формы с организацией ее изменчивости на данном
- 63. Развитие – детерминированный процесс, если (и только если) новые МК возникают на последовательных стадиях развития в
- 64. У 10% зародышей вьюна ЗЩ возникает в точке замыкания ЗК: это тоже возмущение конечной амплитуды (триггер
- 65. ОНТОГЕНЕЗ ОТЛИЧАЕТСЯ ОТ МОРФОГЕНЕЗА ПОЯВЛЕНИЕМ ПРОСТРАНСТВЕННО-ВРЕМЕННЫХ КООРДИНАТ РАЗВИТИЯ Светлая заливка – бластодерма, темная заливка (более плотное
- 66. Сходная динамика изменчивости описана для эмбрионального взаимодействия генов в раннем развитии дрозофилы Нормальное развитие: AP –
- 67. На рисунке – зоны экспрессии (интенсивность флюоресценции мРНК) гена eve, маркирующего переднюю половину каждого сегмента, на
- 69. Скачать презентацию
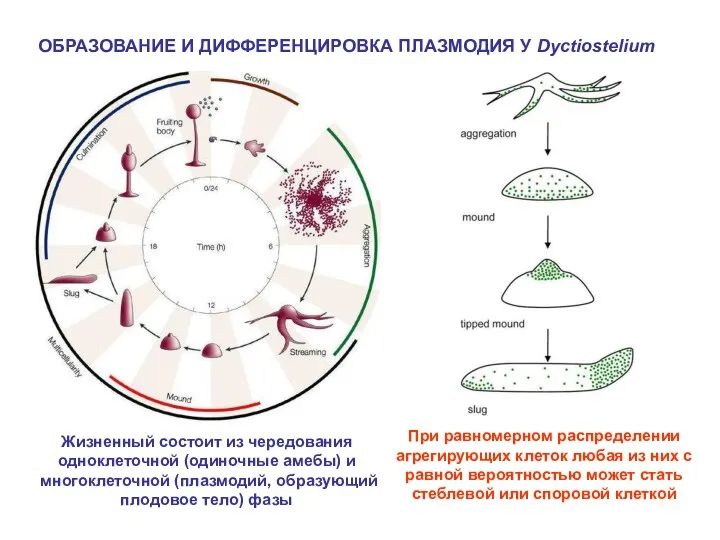
ОБРАЗОВАНИЕ И ДИФФЕРЕНЦИРОВКА ПЛАЗМОДИЯ У Dyctiostelium
Жизненный состоит из чередования одноклеточной (одиночные
ОБРАЗОВАНИЕ И ДИФФЕРЕНЦИРОВКА ПЛАЗМОДИЯ У Dyctiostelium
Жизненный состоит из чередования одноклеточной (одиночные
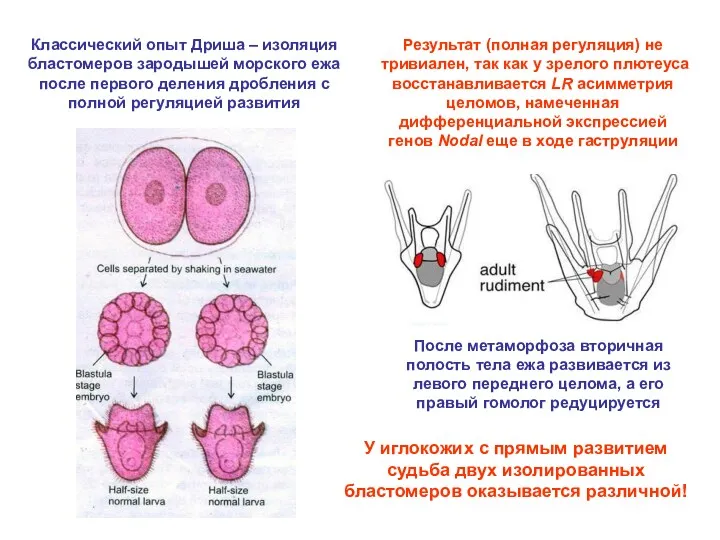
Классический опыт Дриша – изоляция бластомеров зародышей морского ежа после первого
Классический опыт Дриша – изоляция бластомеров зародышей морского ежа после первого

Из каждой сагиттальной половины бластулы Xenopus развивается нормальный пропорционально уменьшенный головастик
Из каждой сагиттальной половины бластулы Xenopus развивается нормальный пропорционально уменьшенный головастик
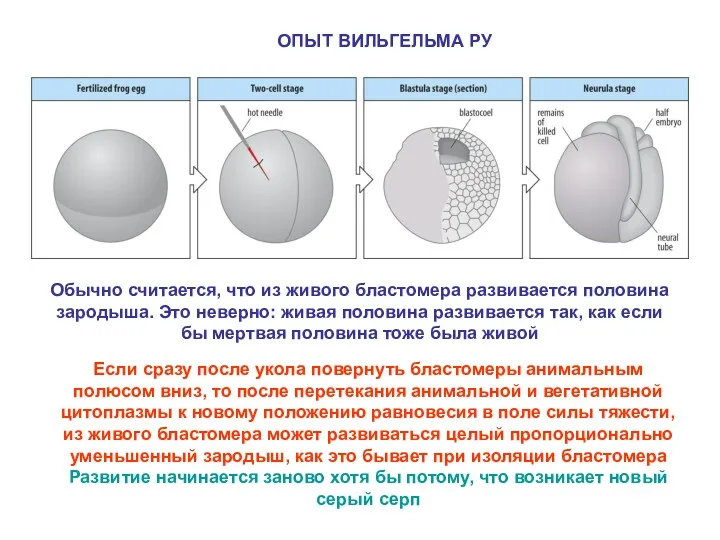
ОПЫТ ВИЛЬГЕЛЬМА РУ
Обычно считается, что из живого бластомера развивается половина зародыша.
ОПЫТ ВИЛЬГЕЛЬМА РУ
Обычно считается, что из живого бластомера развивается половина зародыша.
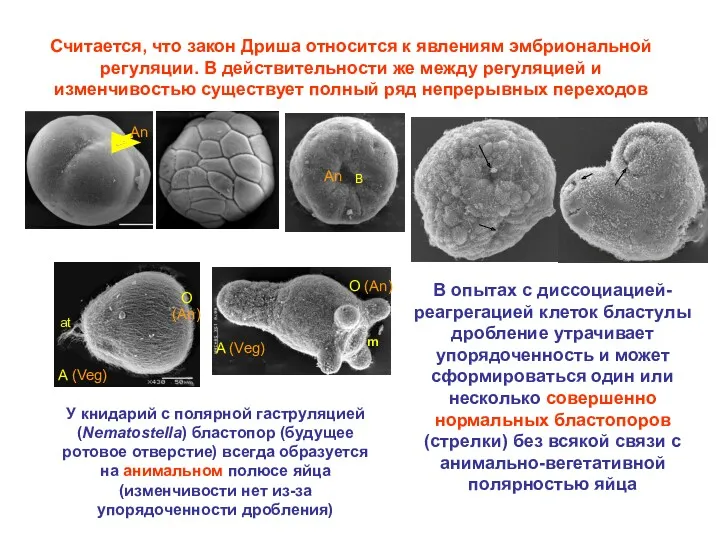
Считается, что закон Дриша относится к явлениям эмбриональной регуляции. В действительности
Считается, что закон Дриша относится к явлениям эмбриональной регуляции. В действительности
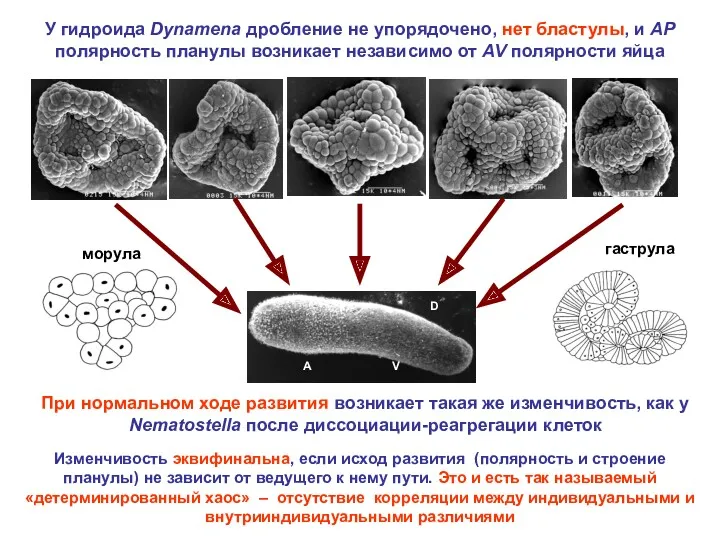
У гидроида Dynamena дробление не упорядочено, нет бластулы, и AP полярность
У гидроида Dynamena дробление не упорядочено, нет бластулы, и AP полярность
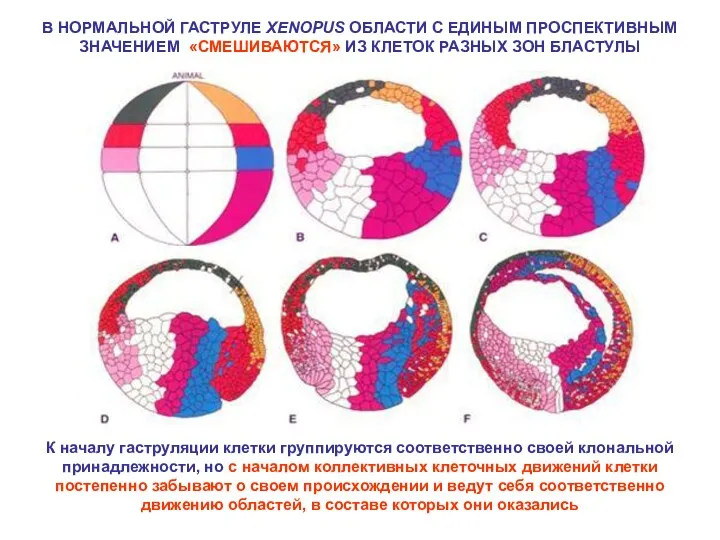
В НОРМАЛЬНОЙ ГАСТРУЛЕ XENOPUS ОБЛАСТИ С ЕДИНЫМ ПРОСПЕКТИВНЫМ ЗНАЧЕНИЕМ «СМЕШИВАЮТСЯ» ИЗ
В НОРМАЛЬНОЙ ГАСТРУЛЕ XENOPUS ОБЛАСТИ С ЕДИНЫМ ПРОСПЕКТИВНЫМ ЗНАЧЕНИЕМ «СМЕШИВАЮТСЯ» ИЗ
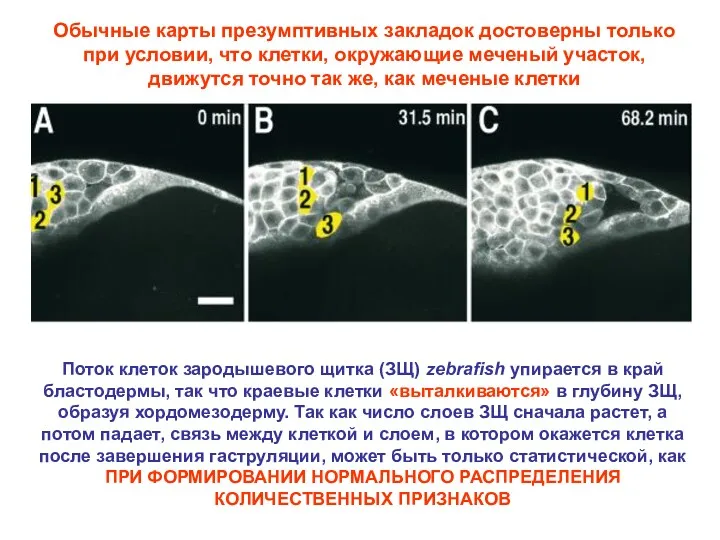
Обычные карты презумптивных закладок достоверны только при условии, что клетки, окружающие
Обычные карты презумптивных закладок достоверны только при условии, что клетки, окружающие
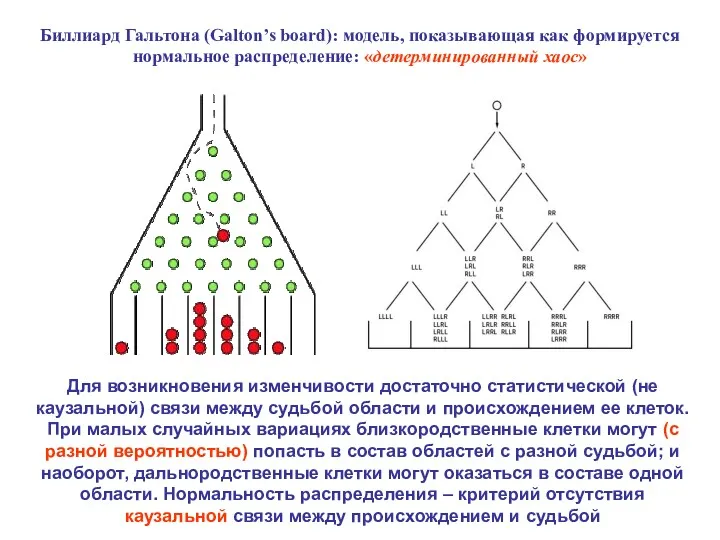
Биллиард Гальтона (Galton’s board): модель, показывающая как формируется нормальное распределение: «детерминированный
Биллиард Гальтона (Galton’s board): модель, показывающая как формируется нормальное распределение: «детерминированный
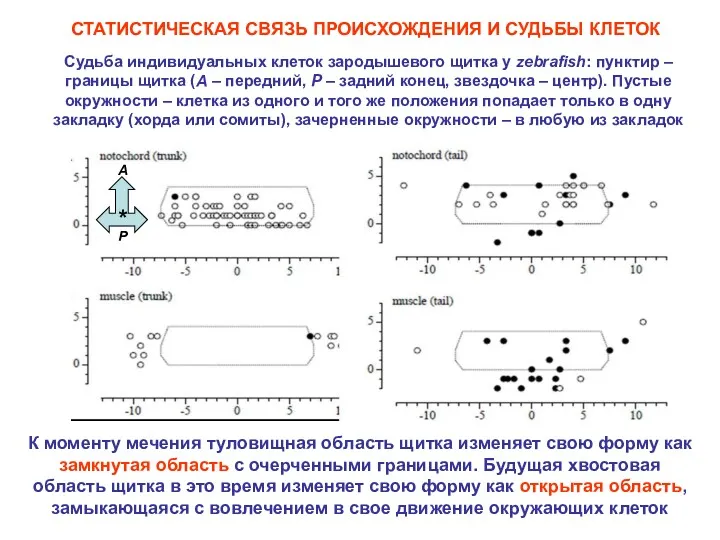
Судьба индивидуальных клеток зародышевого щитка у zebrafish: пунктир – границы щитка
Судьба индивидуальных клеток зародышевого щитка у zebrafish: пунктир – границы щитка
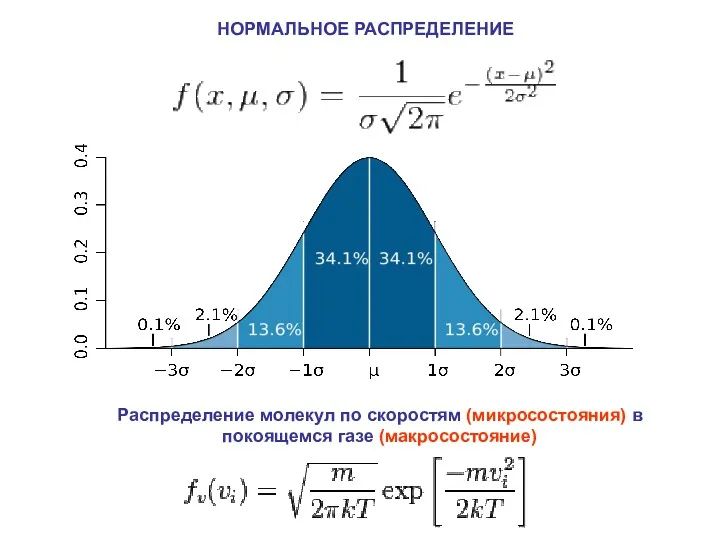
НОРМАЛЬНОЕ РАСПРЕДЕЛЕНИЕ
НОРМАЛЬНОЕ РАСПРЕДЕЛЕНИЕ
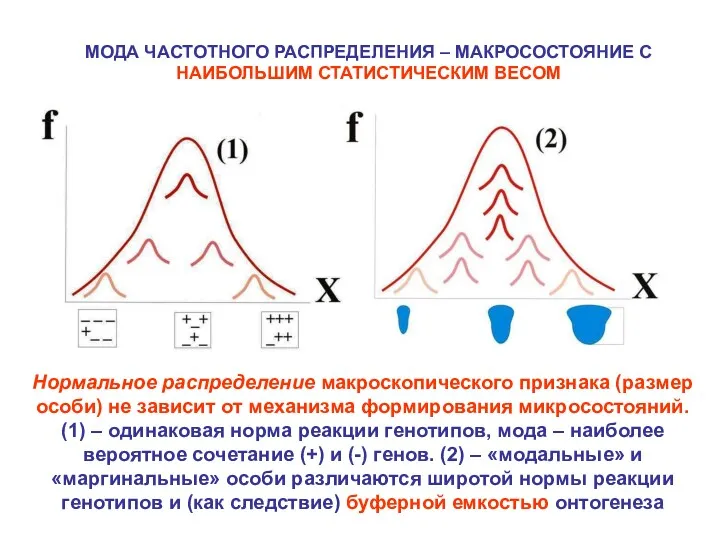
МОДА ЧАСТОТНОГО РАСПРЕДЕЛЕНИЯ – МАКРОСОСТОЯНИЕ С НАИБОЛЬШИМ СТАТИСТИЧЕСКИМ ВЕСОМ
Нормальное распределение
МОДА ЧАСТОТНОГО РАСПРЕДЕЛЕНИЯ – МАКРОСОСТОЯНИЕ С НАИБОЛЬШИМ СТАТИСТИЧЕСКИМ ВЕСОМ
Нормальное распределение
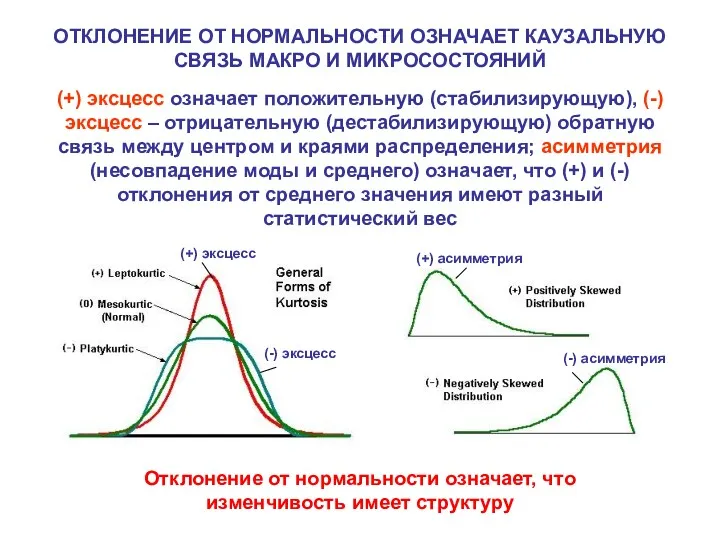
ОТКЛОНЕНИЕ ОТ НОРМАЛЬНОСТИ ОЗНАЧАЕТ КАУЗАЛЬНУЮ СВЯЗЬ МАКРО И МИКРОСОСТОЯНИЙ
(+) эксцесс
ОТКЛОНЕНИЕ ОТ НОРМАЛЬНОСТИ ОЗНАЧАЕТ КАУЗАЛЬНУЮ СВЯЗЬ МАКРО И МИКРОСОСТОЯНИЙ
(+) эксцесс

ФЛЮКТУИРУЮЩАЯ АСИММЕТРИЯ БИЛАТЕРАЛЬНО СИММЕТРИЧНЫХ СТРУКТУР
Ротовой диск головастиков состоит из билатерально симметричных
ФЛЮКТУИРУЮЩАЯ АСИММЕТРИЯ БИЛАТЕРАЛЬНО СИММЕТРИЧНЫХ СТРУКТУР
Ротовой диск головастиков состоит из билатерально симметричных
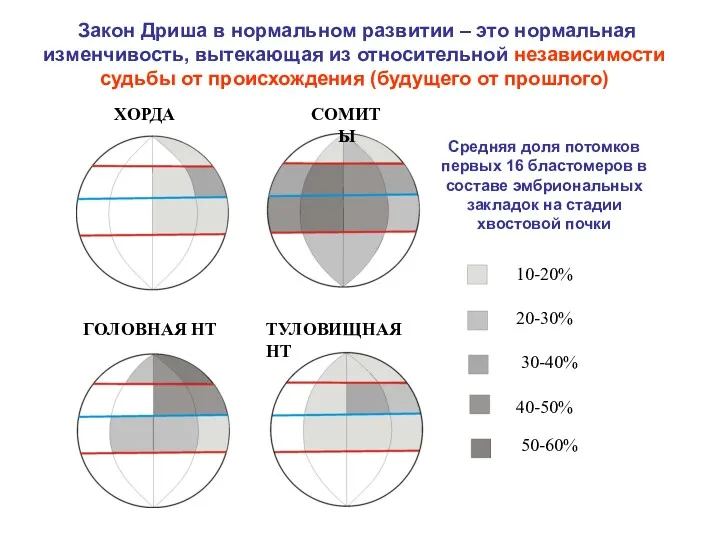
Закон Дриша в нормальном развитии – это нормальная изменчивость, вытекающая
Закон Дриша в нормальном развитии – это нормальная изменчивость, вытекающая
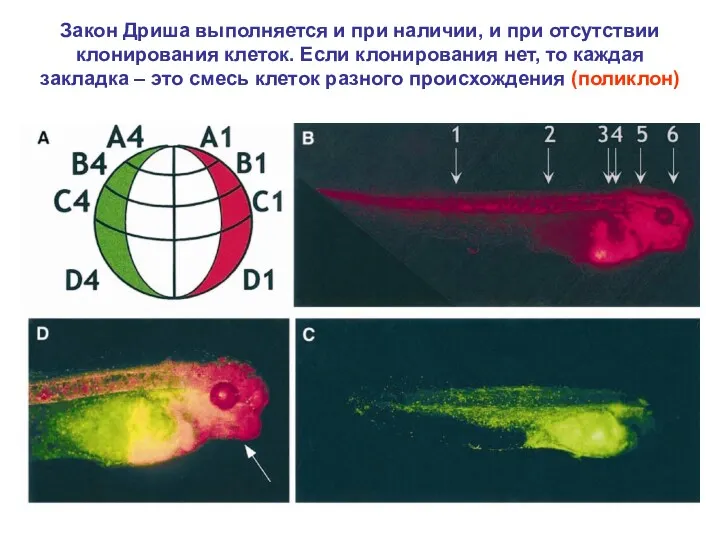
Закон Дриша выполняется и при наличии, и при отсутствии клонирования клеток.
Закон Дриша выполняется и при наличии, и при отсутствии клонирования клеток.
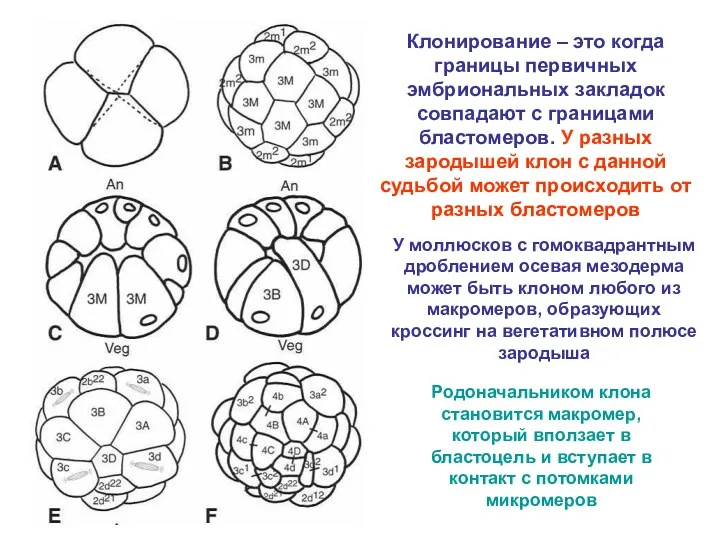
Клонирование – это когда границы первичных эмбриональных закладок совпадают с границами
Клонирование – это когда границы первичных эмбриональных закладок совпадают с границами
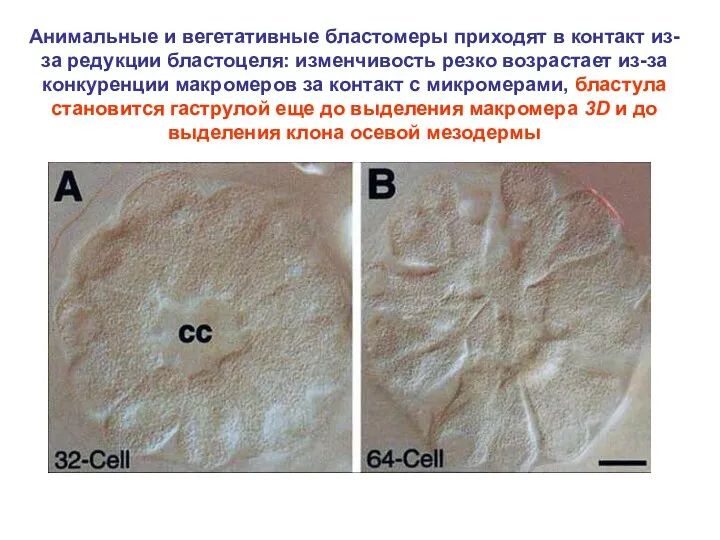
Анимальные и вегетативные бластомеры приходят в контакт из-за редукции бластоцеля: изменчивость
Анимальные и вегетативные бластомеры приходят в контакт из-за редукции бластоцеля: изменчивость
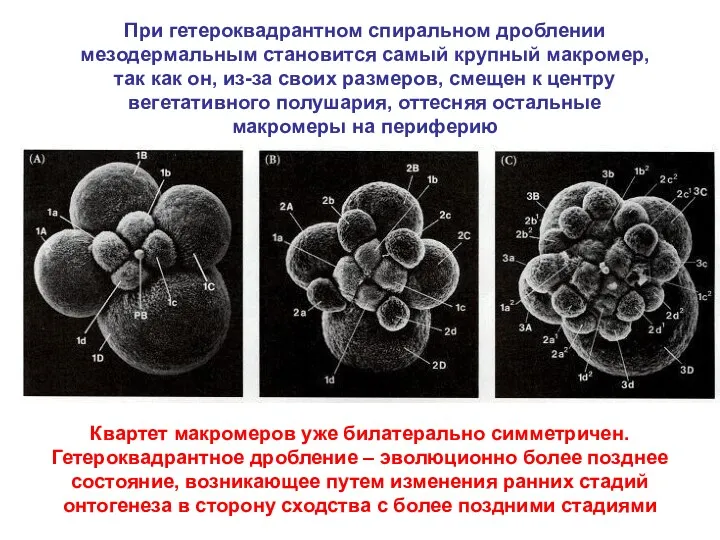
При гетероквадрантном спиральном дроблении мезодермальным становится самый крупный макромер, так как
При гетероквадрантном спиральном дроблении мезодермальным становится самый крупный макромер, так как
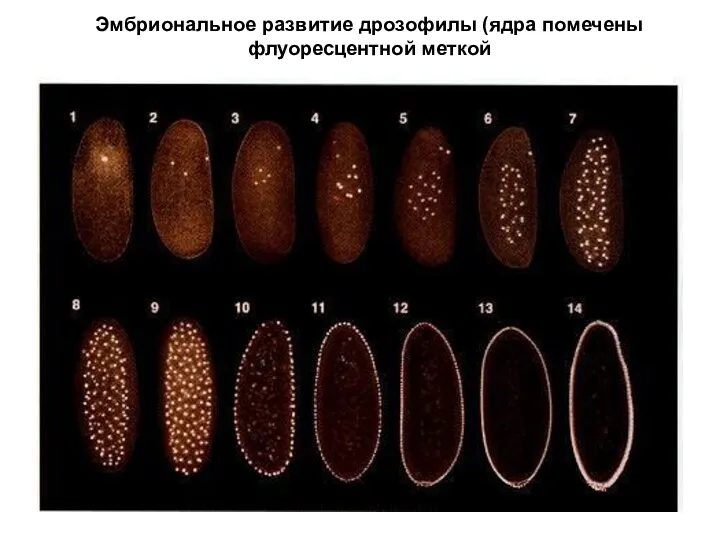
Эмбриональное развитие дрозофилы (ядра помечены флуоресцентной меткой
Эмбриональное развитие дрозофилы (ядра помечены флуоресцентной меткой
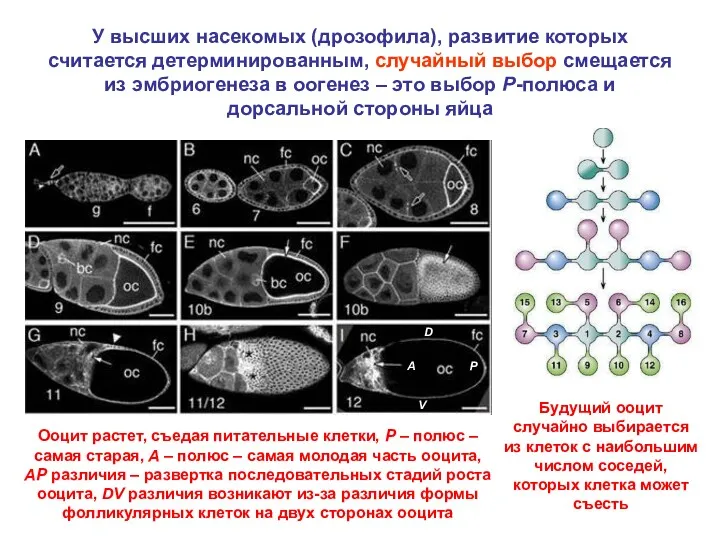
У высших насекомых (дрозофила), развитие которых считается детерминированным, случайный выбор смещается
У высших насекомых (дрозофила), развитие которых считается детерминированным, случайный выбор смещается
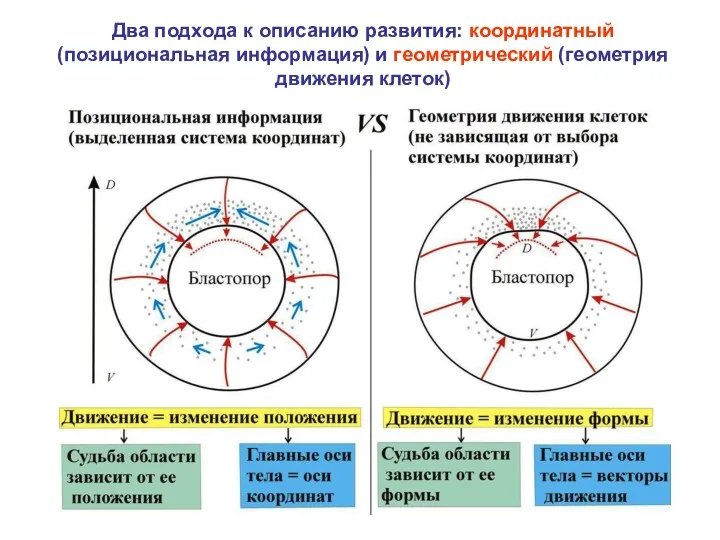
Два подхода к описанию развития: координатный (позициональная информация) и геометрический (геометрия
Два подхода к описанию развития: координатный (позициональная информация) и геометрический (геометрия
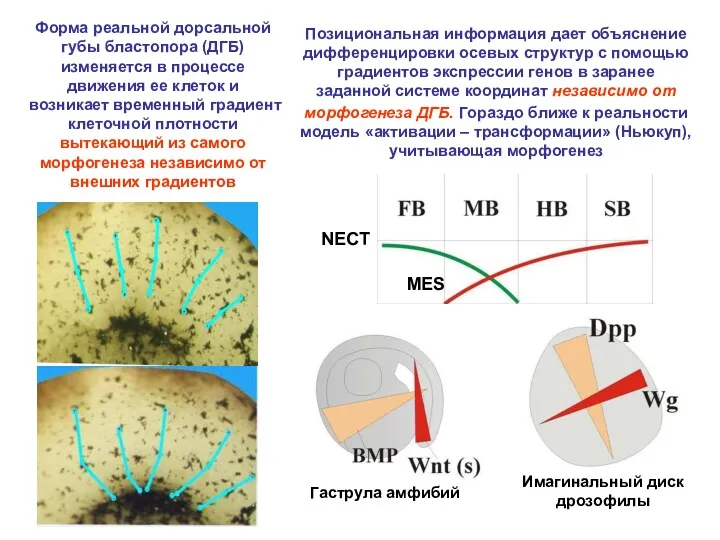
Позициональная информация дает объяснение дифференцировки осевых структур с помощью градиентов экспрессии
Позициональная информация дает объяснение дифференцировки осевых структур с помощью градиентов экспрессии
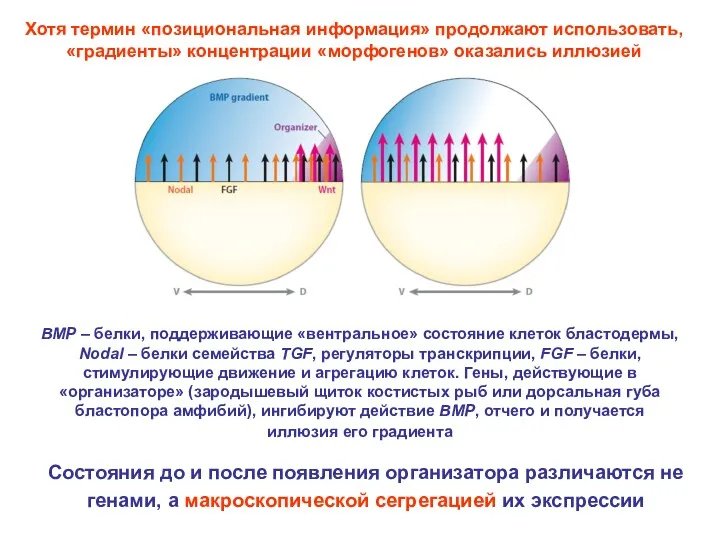
Хотя термин «позициональная информация» продолжают использовать, «градиенты» концентрации «морфогенов» оказались иллюзией
BMP
Хотя термин «позициональная информация» продолжают использовать, «градиенты» концентрации «морфогенов» оказались иллюзией
BMP
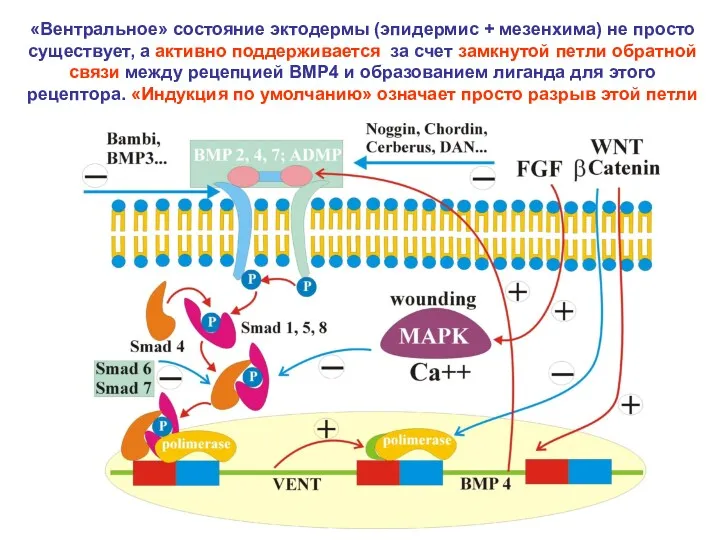
«Вентральное» состояние эктодермы (эпидермис + мезенхима) не просто существует, а активно
«Вентральное» состояние эктодермы (эпидермис + мезенхима) не просто существует, а активно
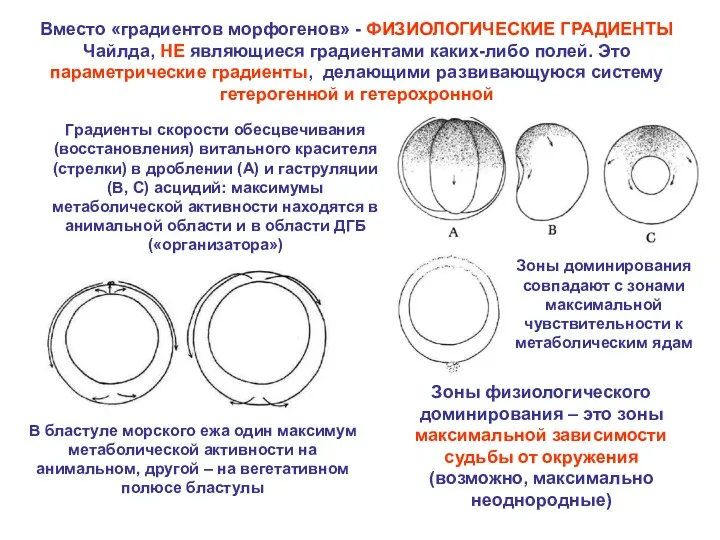
Вместо «градиентов морфогенов» - ФИЗИОЛОГИЧЕСКИЕ ГРАДИЕНТЫ Чайлда, НЕ являющиеся градиентами каких-либо
Вместо «градиентов морфогенов» - ФИЗИОЛОГИЧЕСКИЕ ГРАДИЕНТЫ Чайлда, НЕ являющиеся градиентами каких-либо
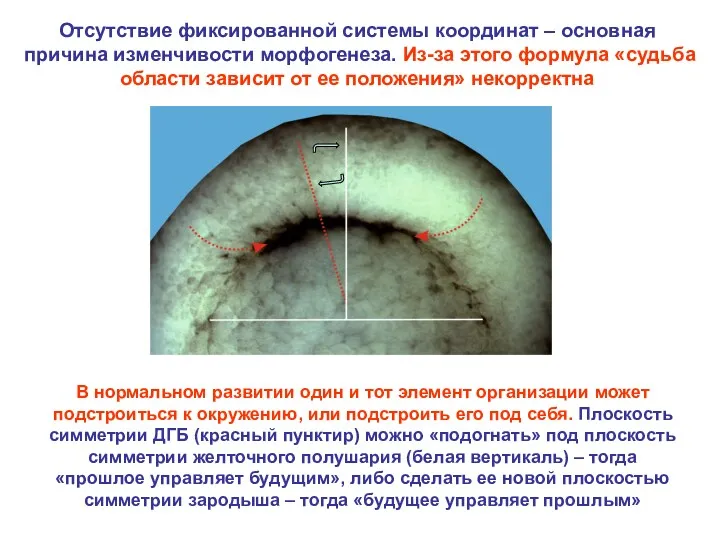
В нормальном развитии один и тот элемент организации может подстроиться к
В нормальном развитии один и тот элемент организации может подстроиться к
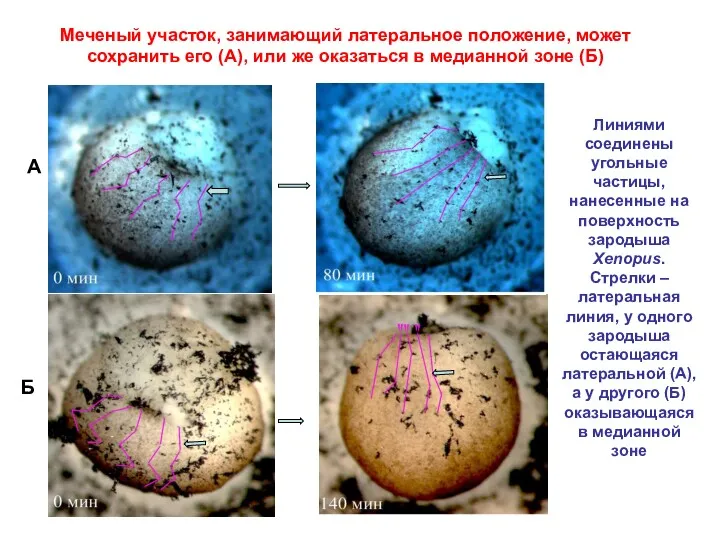
Меченый участок, занимающий латеральное положение, может сохранить его (А), или же
Меченый участок, занимающий латеральное положение, может сохранить его (А), или же
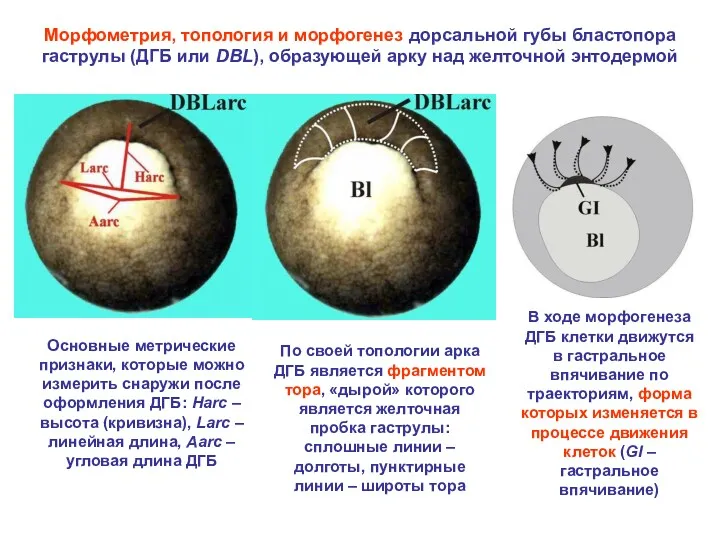
Морфометрия, топология и морфогенез дорсальной губы бластопора гаструлы (ДГБ или DBL),
Морфометрия, топология и морфогенез дорсальной губы бластопора гаструлы (ДГБ или DBL),
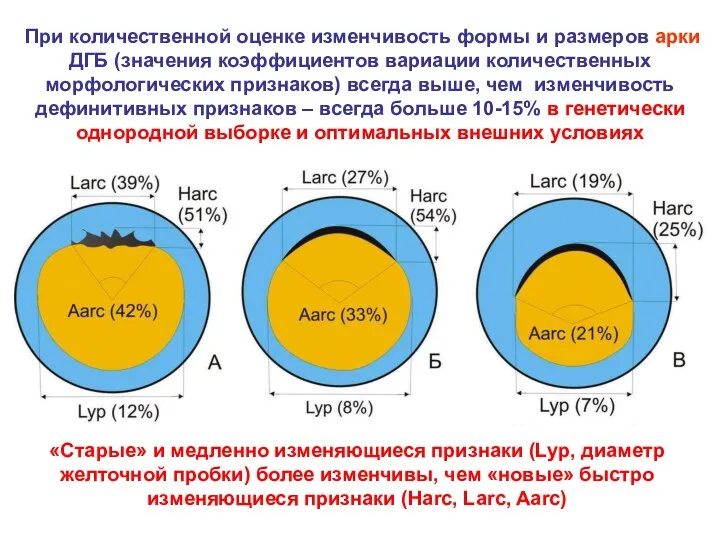
При количественной оценке изменчивость формы и размеров арки ДГБ (значения коэффициентов
При количественной оценке изменчивость формы и размеров арки ДГБ (значения коэффициентов
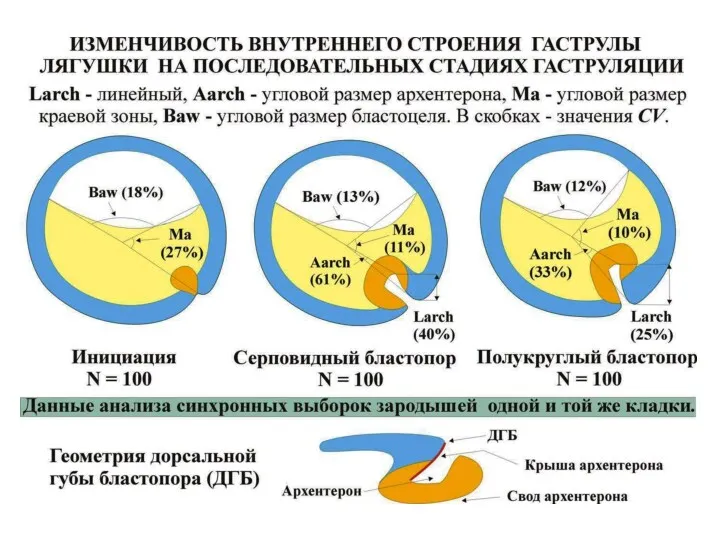

Масштабы изменчивости развития в генетически однородной выборке зародышей, развивающихся в оптимальных
Масштабы изменчивости развития в генетически однородной выборке зародышей, развивающихся в оптимальных
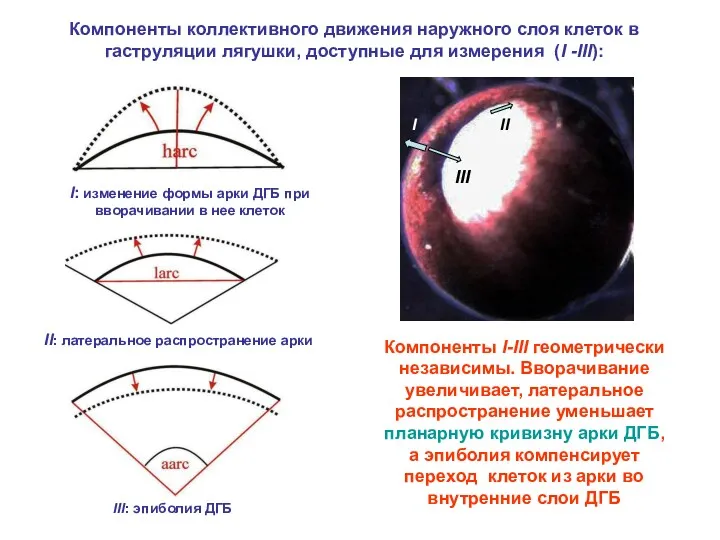
Компоненты коллективного движения наружного слоя клеток в гаструляции лягушки, доступные для
Компоненты коллективного движения наружного слоя клеток в гаструляции лягушки, доступные для
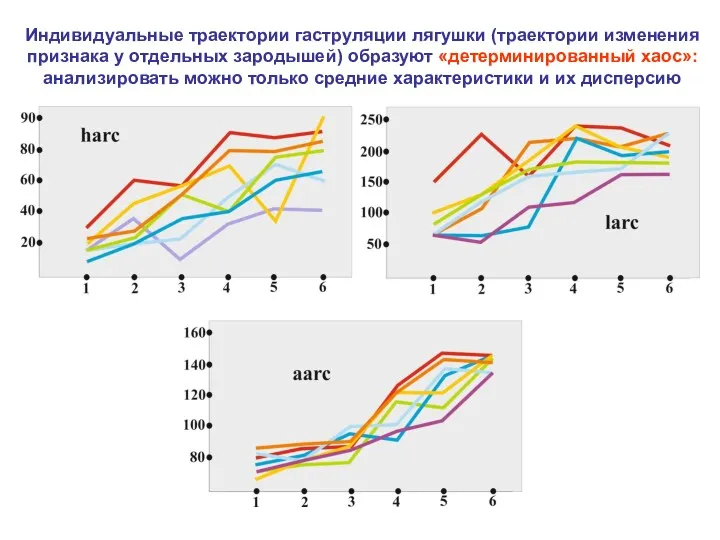
Индивидуальные траектории гаструляции лягушки (траектории изменения признака у отдельных зародышей) образуют
Индивидуальные траектории гаструляции лягушки (траектории изменения признака у отдельных зародышей) образуют
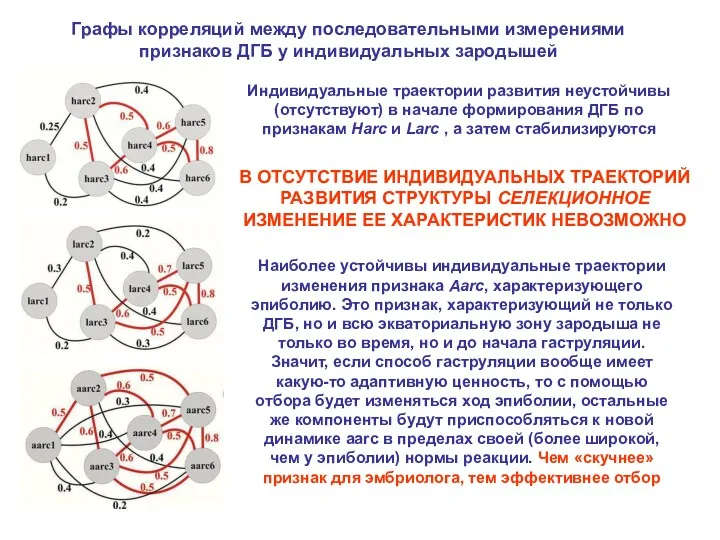
Графы корреляций между последовательными измерениями признаков ДГБ у индивидуальных зародышей
Индивидуальные
Графы корреляций между последовательными измерениями признаков ДГБ у индивидуальных зародышей
Индивидуальные
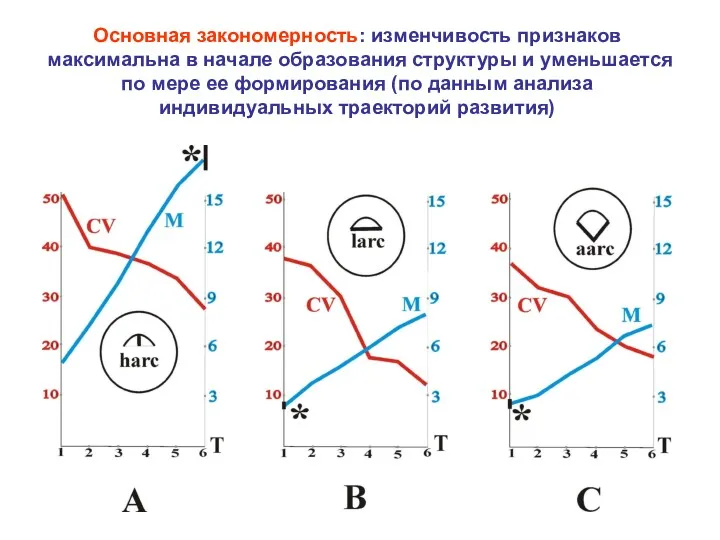
Основная закономерность: изменчивость признаков максимальна в начале образования структуры и уменьшается
Основная закономерность: изменчивость признаков максимальна в начале образования структуры и уменьшается
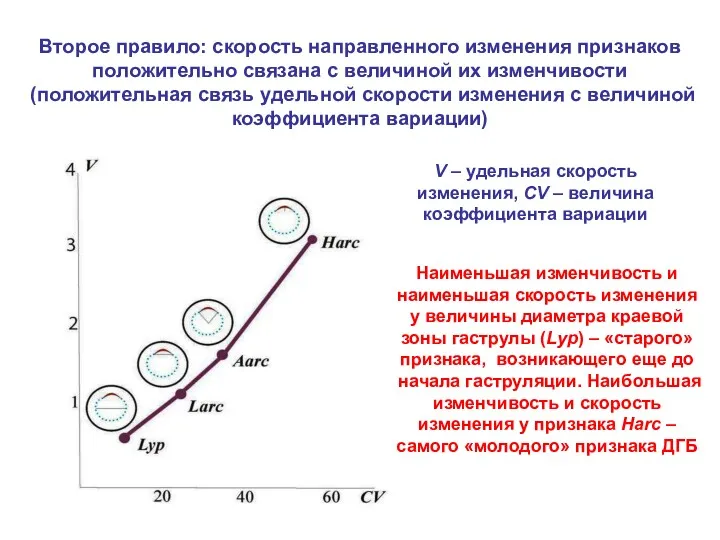
Второе правило: скорость направленного изменения признаков положительно связана с величиной их
Второе правило: скорость направленного изменения признаков положительно связана с величиной их
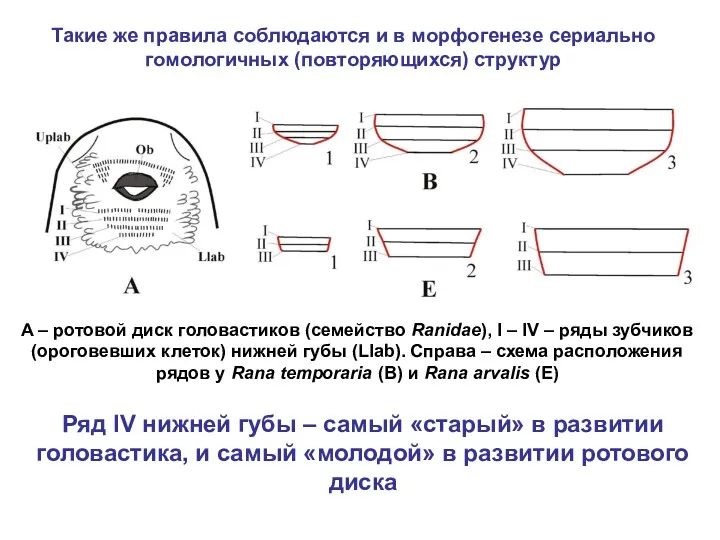
Такие же правила соблюдаются и в морфогенезе сериально гомологичных (повторяющихся) структур
Такие же правила соблюдаются и в морфогенезе сериально гомологичных (повторяющихся) структур
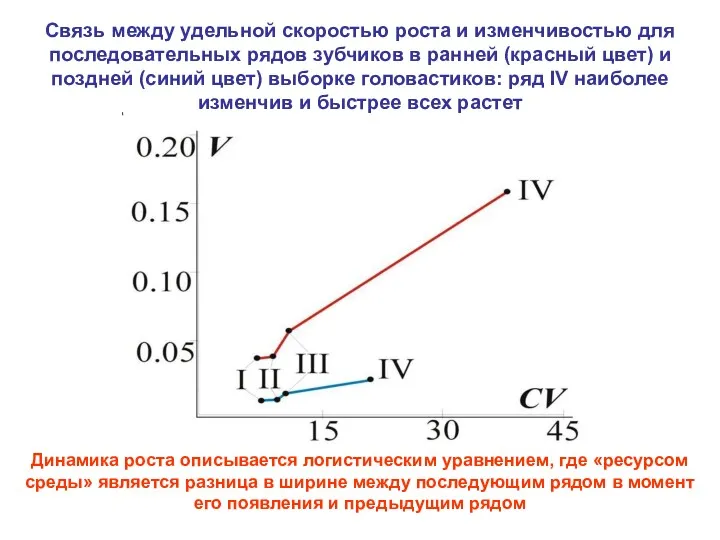
Связь между удельной скоростью роста и изменчивостью для последовательных рядов зубчиков
Связь между удельной скоростью роста и изменчивостью для последовательных рядов зубчиков
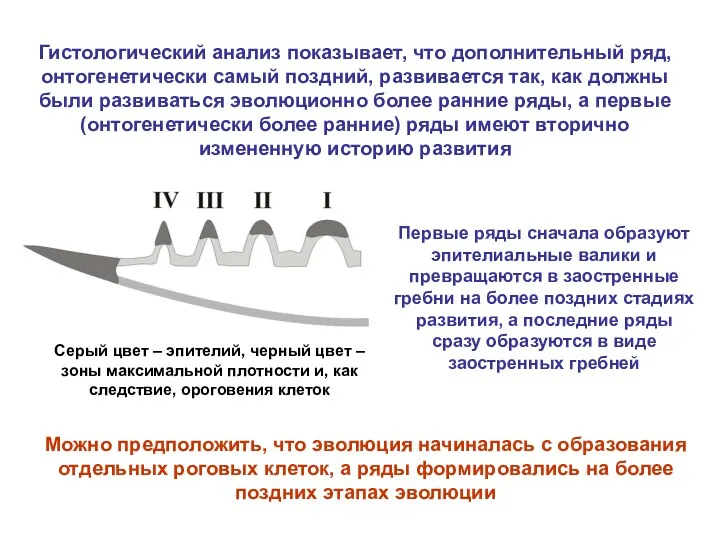
Гистологический анализ показывает, что дополнительный ряд, онтогенетически самый поздний, развивается так,
Гистологический анализ показывает, что дополнительный ряд, онтогенетически самый поздний, развивается так,
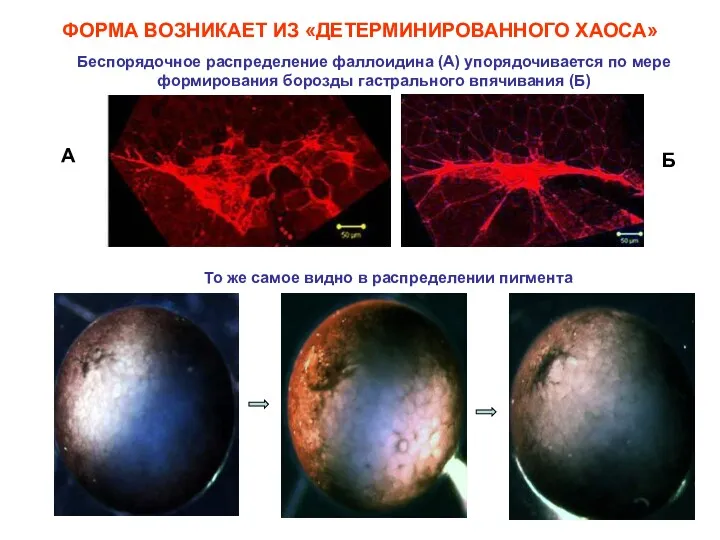
Беспорядочное распределение фаллоидина (А) упорядочивается по мере формирования борозды гастрального впячивания
Беспорядочное распределение фаллоидина (А) упорядочивается по мере формирования борозды гастрального впячивания
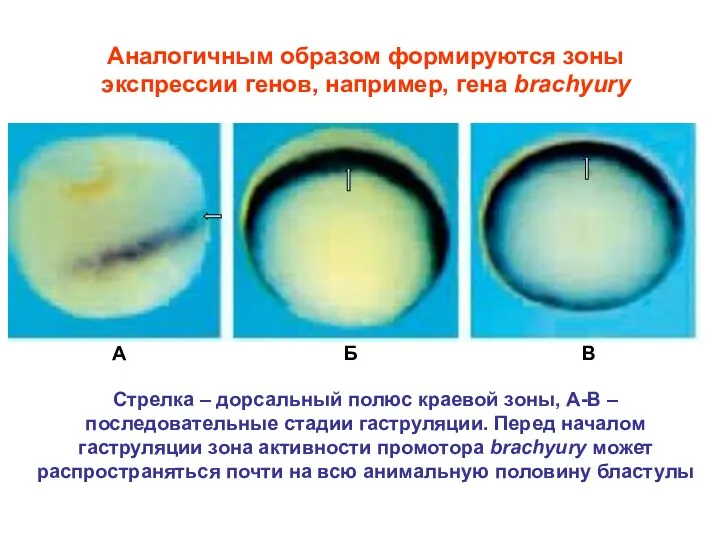
Аналогичным образом формируются зоны экспрессии генов, например, гена brachyury
А
Б
В
Стрелка – дорсальный
Аналогичным образом формируются зоны экспрессии генов, например, гена brachyury
А
Б
В
Стрелка – дорсальный

Изменчивость связана с гетерохрониями между эпиболией и изменением формы дорсальной губы
Изменчивость связана с гетерохрониями между эпиболией и изменением формы дорсальной губы

Пунктирные стрелки – вход и выход клеток из переходной зоны ДГБ,
Пунктирные стрелки – вход и выход клеток из переходной зоны ДГБ,
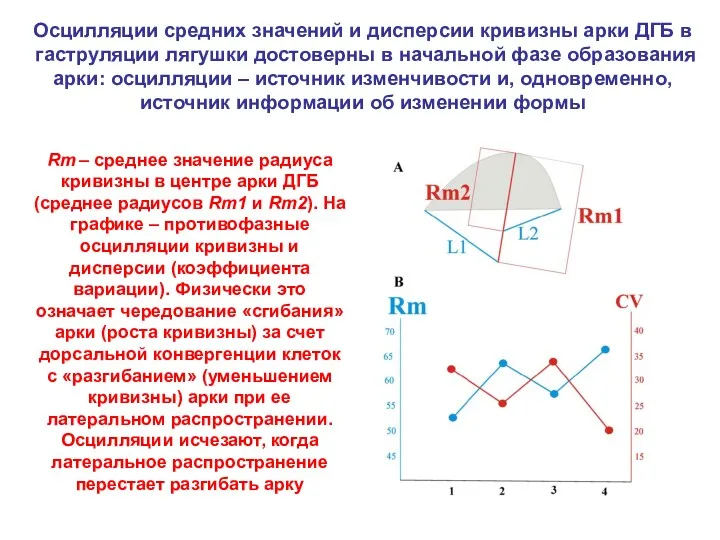
Осцилляции средних значений и дисперсии кривизны арки ДГБ в гаструляции лягушки
Осцилляции средних значений и дисперсии кривизны арки ДГБ в гаструляции лягушки
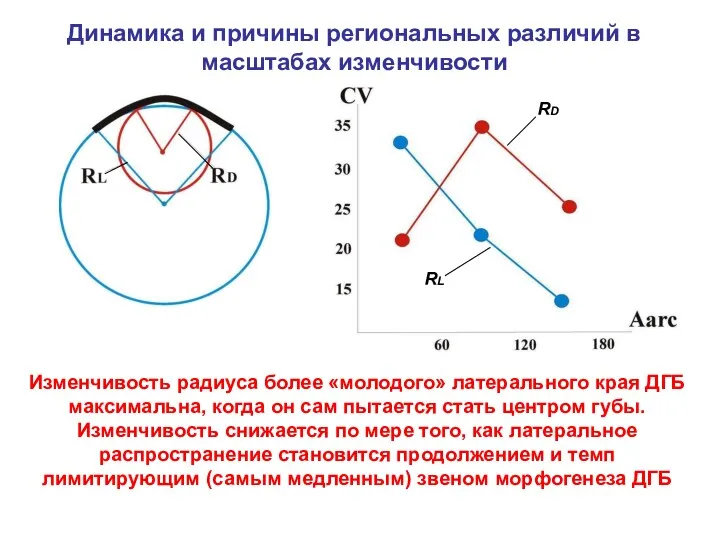
Динамика и причины региональных различий в масштабах изменчивости
Изменчивость радиуса более «молодого»
Динамика и причины региональных различий в масштабах изменчивости
Изменчивость радиуса более «молодого»
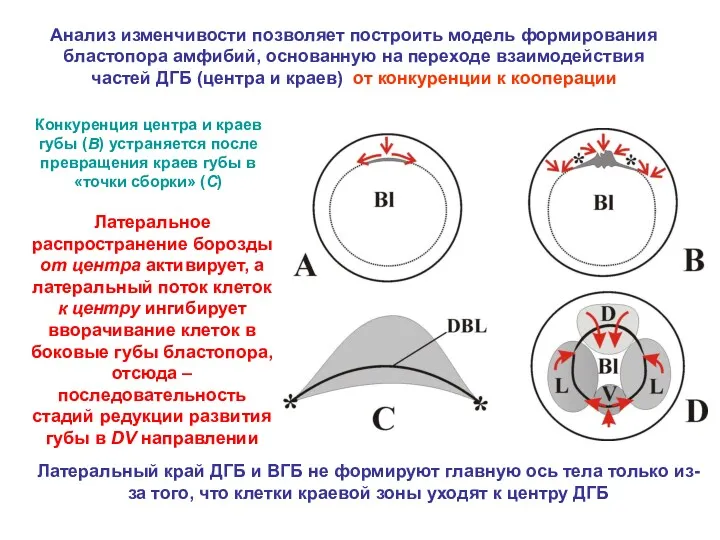
Латеральный край ДГБ и ВГБ не формируют главную ось тела только
Латеральный край ДГБ и ВГБ не формируют главную ось тела только
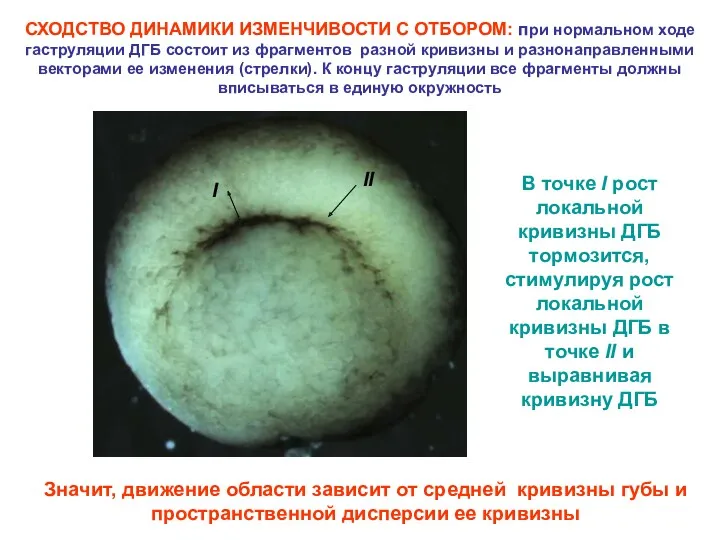
СХОДСТВО ДИНАМИКИ ИЗМЕНЧИВОСТИ С ОТБОРОМ: при нормальном ходе гаструляции ДГБ состоит
СХОДСТВО ДИНАМИКИ ИЗМЕНЧИВОСТИ С ОТБОРОМ: при нормальном ходе гаструляции ДГБ состоит
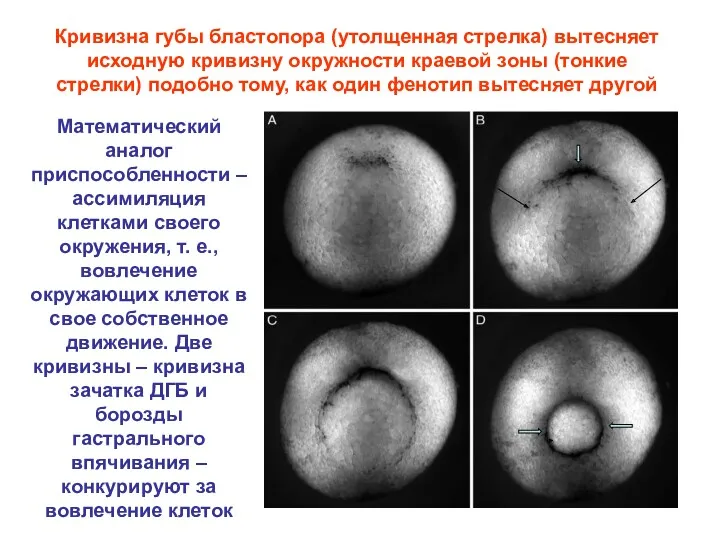
Кривизна губы бластопора (утолщенная стрелка) вытесняет исходную кривизну окружности краевой зоны
Кривизна губы бластопора (утолщенная стрелка) вытесняет исходную кривизну окружности краевой зоны
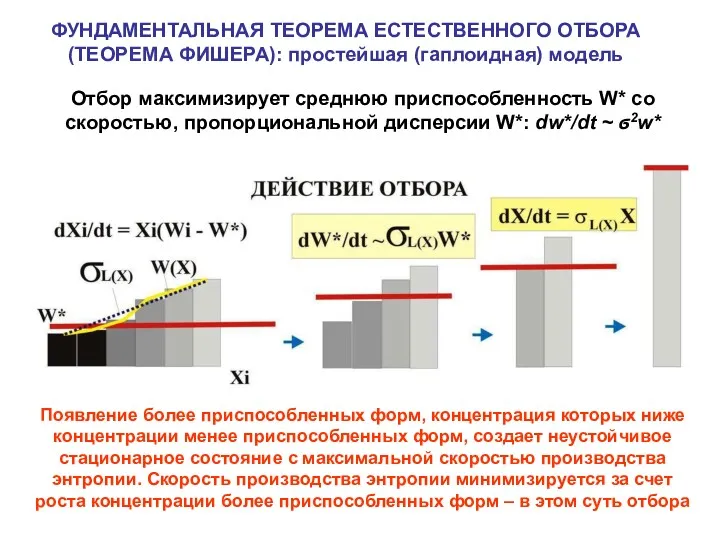
ФУНДАМЕНТАЛЬНАЯ ТЕОРЕМА ЕСТЕСТВЕННОГО ОТБОРА (ТЕОРЕМА ФИШЕРА): простейшая (гаплоидная) модель
Отбор максимизирует среднюю
ФУНДАМЕНТАЛЬНАЯ ТЕОРЕМА ЕСТЕСТВЕННОГО ОТБОРА (ТЕОРЕМА ФИШЕРА): простейшая (гаплоидная) модель
Отбор максимизирует среднюю
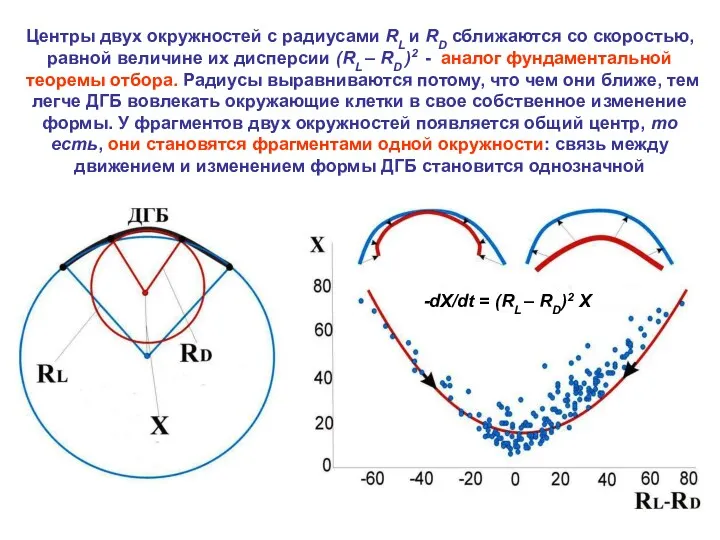
Центры двух окружностей с радиусами RL и RD сближаются со скоростью,
Центры двух окружностей с радиусами RL и RD сближаются со скоростью,
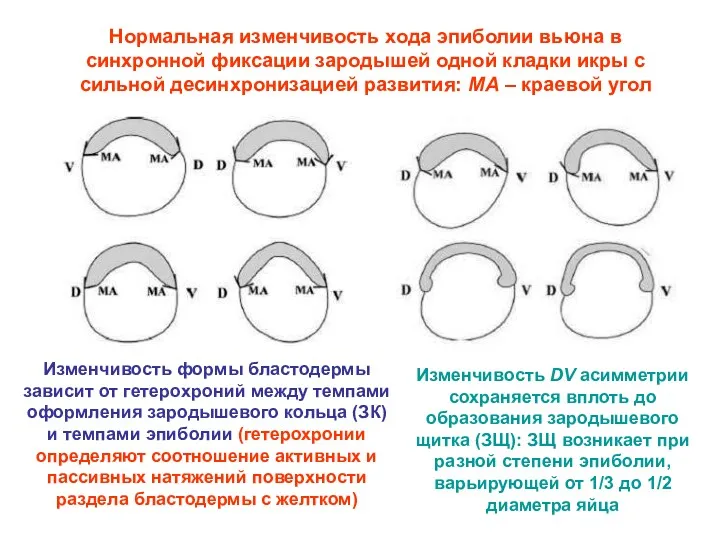
Нормальная изменчивость хода эпиболии вьюна в синхронной фиксации зародышей одной кладки
Нормальная изменчивость хода эпиболии вьюна в синхронной фиксации зародышей одной кладки
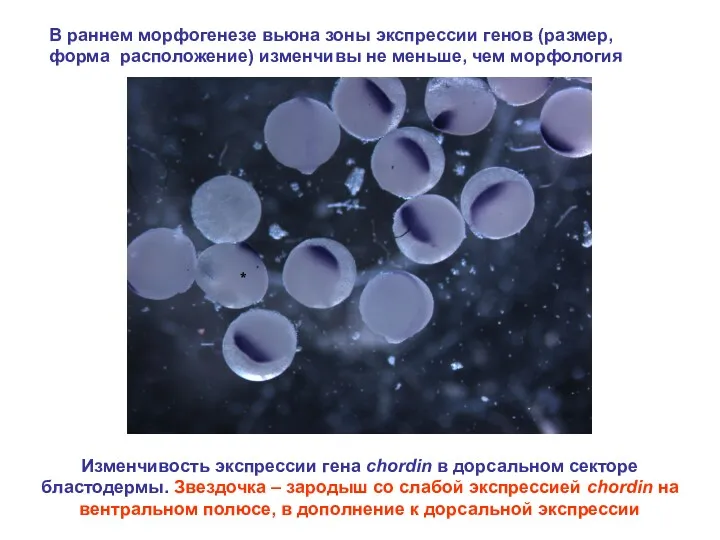
В раннем морфогенезе вьюна зоны экспрессии генов (размер, форма расположение) изменчивы
В раннем морфогенезе вьюна зоны экспрессии генов (размер, форма расположение) изменчивы

Изменчивость зональной экспрессии генов подчиняется основному правилу – она максимальна при
Изменчивость зональной экспрессии генов подчиняется основному правилу – она максимальна при
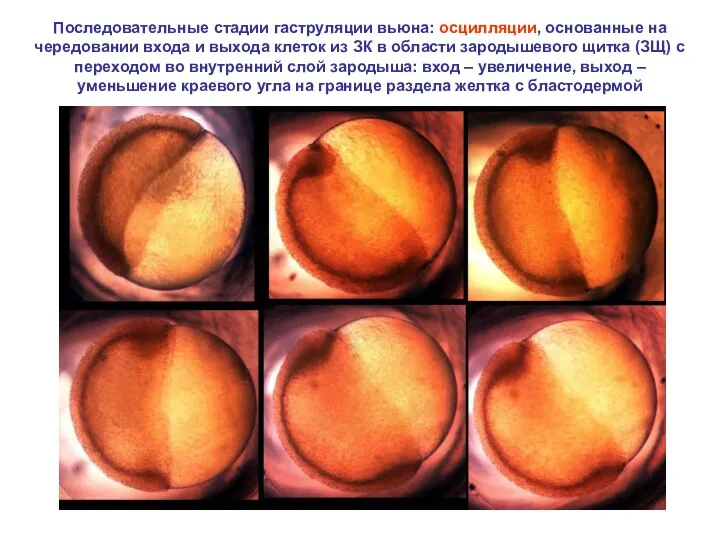
Последовательные стадии гаструляции вьюна: осцилляции, основанные на чередовании входа и выхода
Последовательные стадии гаструляции вьюна: осцилляции, основанные на чередовании входа и выхода
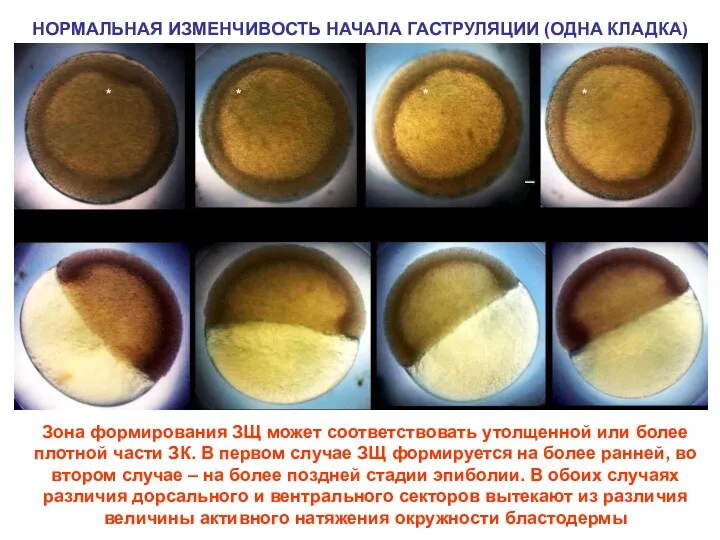
Зона формирования ЗЩ может соответствовать утолщенной или более плотной части ЗК.
Зона формирования ЗЩ может соответствовать утолщенной или более плотной части ЗК.

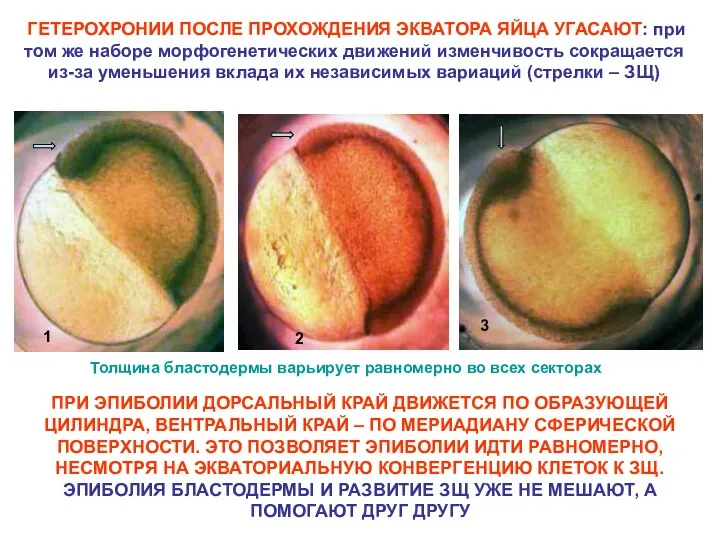
НОРМАЛЬНАЯ ИЗМЕНЧИВОСТЬ НАЧАЛА ГАСТРУЛЯЦИИ (ЗАРОДЫШИ РАЗНЫХ КЛАДОК)
Гетерохронии между входом и выходом
НОРМАЛЬНАЯ ИЗМЕНЧИВОСТЬ НАЧАЛА ГАСТРУЛЯЦИИ (ЗАРОДЫШИ РАЗНЫХ КЛАДОК)
Гетерохронии между входом и выходом
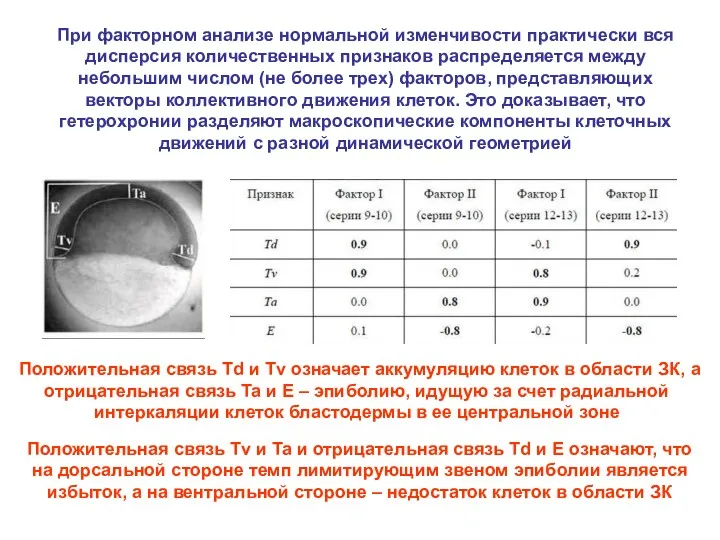
При факторном анализе нормальной изменчивости практически вся дисперсия количественных признаков распределяется
При факторном анализе нормальной изменчивости практически вся дисперсия количественных признаков распределяется
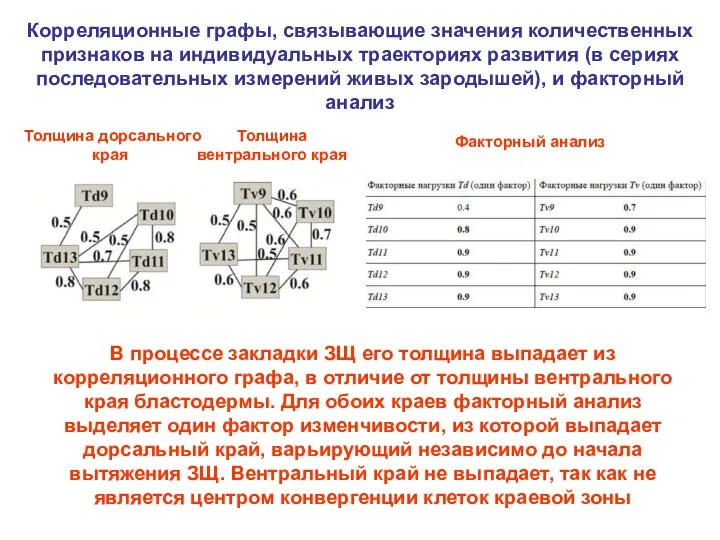
Корреляционные графы, связывающие значения количественных признаков на индивидуальных траекториях развития (в
Корреляционные графы, связывающие значения количественных признаков на индивидуальных траекториях развития (в
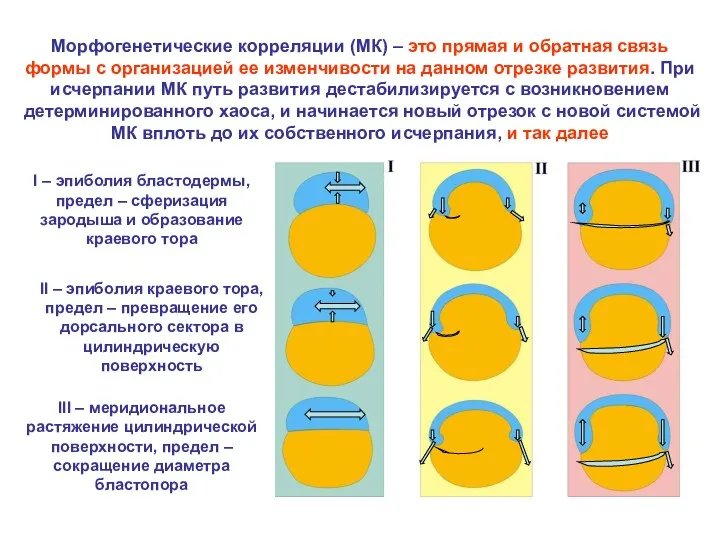
Морфогенетические корреляции (МК) – это прямая и обратная связь формы с
Морфогенетические корреляции (МК) – это прямая и обратная связь формы с
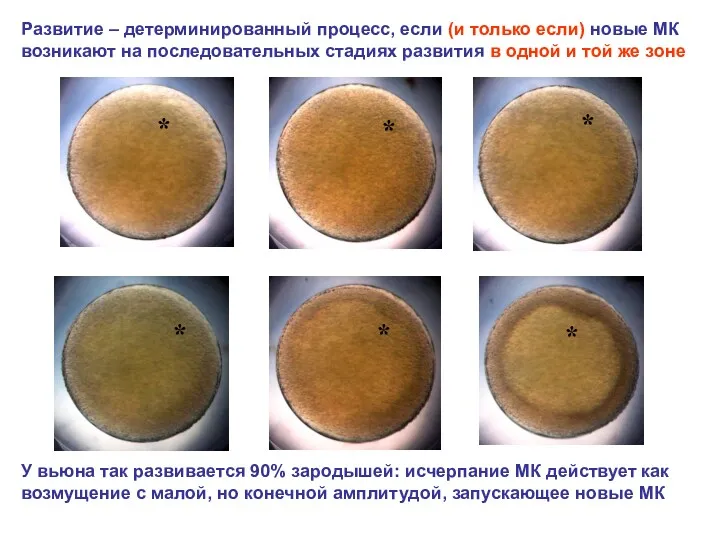
Развитие – детерминированный процесс, если (и только если) новые МК возникают
Развитие – детерминированный процесс, если (и только если) новые МК возникают
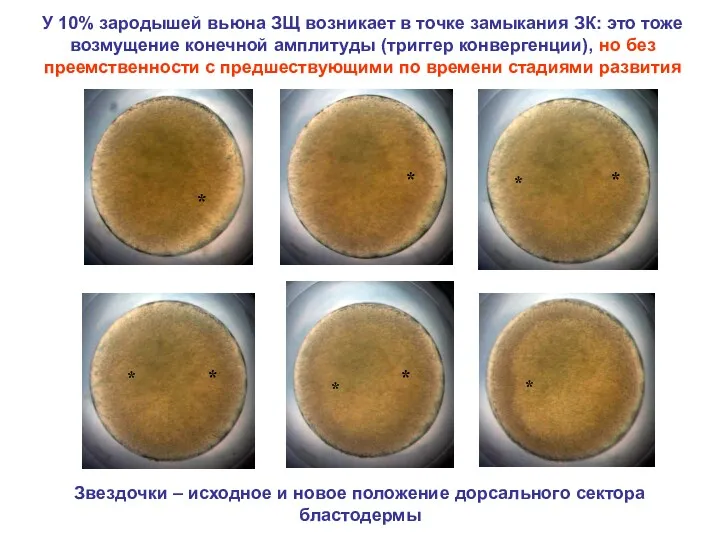
У 10% зародышей вьюна ЗЩ возникает в точке замыкания ЗК: это
У 10% зародышей вьюна ЗЩ возникает в точке замыкания ЗК: это
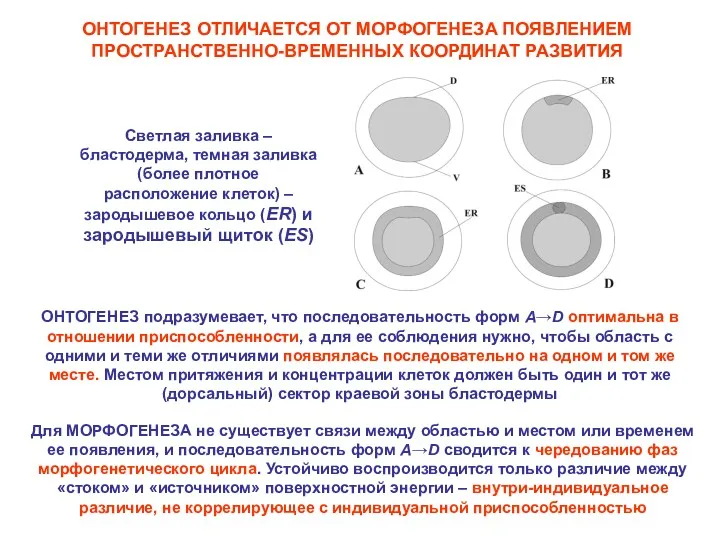
ОНТОГЕНЕЗ ОТЛИЧАЕТСЯ ОТ МОРФОГЕНЕЗА ПОЯВЛЕНИЕМ ПРОСТРАНСТВЕННО-ВРЕМЕННЫХ КООРДИНАТ РАЗВИТИЯ
Светлая заливка – бластодерма,
ОНТОГЕНЕЗ ОТЛИЧАЕТСЯ ОТ МОРФОГЕНЕЗА ПОЯВЛЕНИЕМ ПРОСТРАНСТВЕННО-ВРЕМЕННЫХ КООРДИНАТ РАЗВИТИЯ
Светлая заливка – бластодерма,
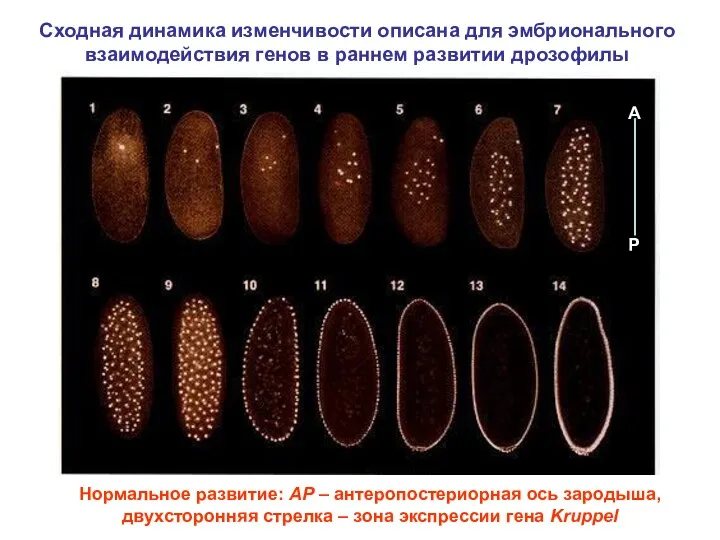
Сходная динамика изменчивости описана для эмбрионального взаимодействия генов в раннем развитии
Сходная динамика изменчивости описана для эмбрионального взаимодействия генов в раннем развитии
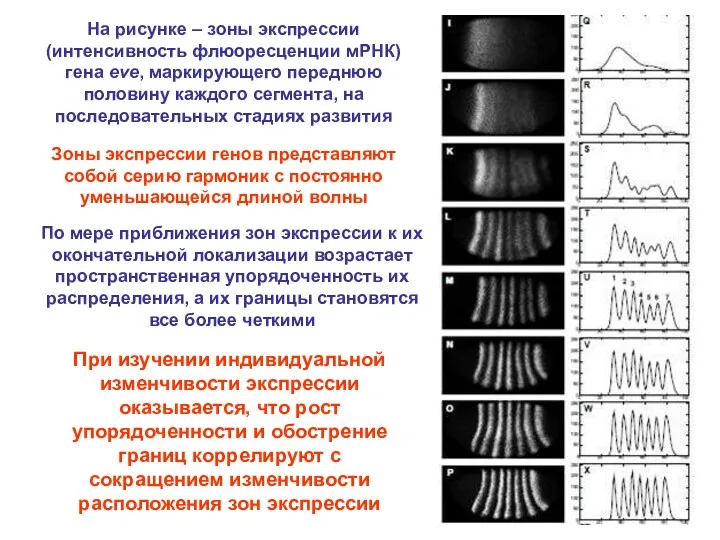
На рисунке – зоны экспрессии (интенсивность флюоресценции мРНК) гена eve, маркирующего
На рисунке – зоны экспрессии (интенсивность флюоресценции мРНК) гена eve, маркирующего
 Корень. Внешнее и внутреннее строение корня
Корень. Внешнее и внутреннее строение корня газообмен в легких
газообмен в легких ГМО в нашей жизни
ГМО в нашей жизни Сибирский кедр
Сибирский кедр Пищеварение в полости рта: состав и свойства слюны
Пищеварение в полости рта: состав и свойства слюны Биологическая обработка органических отходов , их характеристика и принципы обработки
Биологическая обработка органических отходов , их характеристика и принципы обработки Биологические ритмы. Сон и его значение
Биологические ритмы. Сон и его значение Основы гидро - и гемодинамики
Основы гидро - и гемодинамики Холинергиялық синапс құрылысы мен қызметінің байланысы
Холинергиялық синапс құрылысы мен қызметінің байланысы Моногибридное скрещивание.Задачи.
Моногибридное скрещивание.Задачи. Двигательные системы
Двигательные системы Вирусы. Строение и свойства вирусов
Вирусы. Строение и свойства вирусов Эволюция нервной, кровеносной и выделительной систем органов
Эволюция нервной, кровеносной и выделительной систем органов Общая вирусология
Общая вирусология Чому мігрують тварини
Чому мігрують тварини Определение резистентности бактерий к антибиотикам и наночастицам методами сканирующей зондовой микроскопии
Определение резистентности бактерий к антибиотикам и наночастицам методами сканирующей зондовой микроскопии Хвойные растения
Хвойные растения Тварини минулого
Тварини минулого Типы развития насекомых
Типы развития насекомых Декоративные качества цветков
Декоративные качества цветков Многообразие и значение земноводных
Многообразие и значение земноводных Конкурс Учитель года-2016 Республиканский этап.
Конкурс Учитель года-2016 Республиканский этап. Семейство пингвины
Семейство пингвины Центральная нервная система
Центральная нервная система Основы молекулярной биологии
Основы молекулярной биологии Костная система человека
Костная система человека Медицинская арахноэнтомология
Медицинская арахноэнтомология Малина звичайна
Малина звичайна