- Жасушааралық сигнализацияның және интеграцияның молекулярлық механизмдері

Содержание
- 2. Жасушааралық сигнализация және интеграцияның молекулалық механизмдері. Жасуша ерекше сигнал жауабына молекулалық каскадтық механизмді жібереді, ол биохимиялық
- 3. Жасушадан тыс сигналдар химиялық және физикалық болып бөлінеді. Химиялық сигналдар: аминокишқылдар, ақуыздар, гормондар, нуклеотидтер, май қышқылдары
- 4. Барлық сигналдық молекулалар 3 классқа бөлінеді: Гормондар. Ішкі секреция бездерінде синтезделеді, содан кейін қан ағымы арқылы
- 5. Жасушалық жауаптың ерекшелігі рецептор түрімен анықталынады, өйткені жасушаларда рецепторлар жиынтығы бар, олар орналасады: 1- жасушада (беткейлік,
- 7. I – жасушалық мембрана арқылы ерігіш майлардың өтуі; II – сигналдық малекулалардың рецепторлармен байланысы және оның
- 8. Жасушаның беткейлік рецепторларымен байланысты өтетін сигналдардың берілу механизмдері. Рецепторлардың бұл жиынтығы мембранадағы интегралдық ақуыздар жатады. E=extracellular
- 9. Сигналды берудің жалпы кестесінің 5 негізгі этаптары бар. Сигналды берудің бұзылуы әрбір этапта болуы мүмкін, ол
- 10. 4 этап- эффектор активациясы бұл G-ақуыздың - альфа суббірлігі. 5 этап – сигналдың әлсіреуі. Немесе эффектордың
- 11. G-ақуыздар – бұлар 3 суббірліктен тұратын тримерлер. Бұл ақуыздар мембрана құрамына кірмейді, бір жағынан мембрананың цитоплазмаға
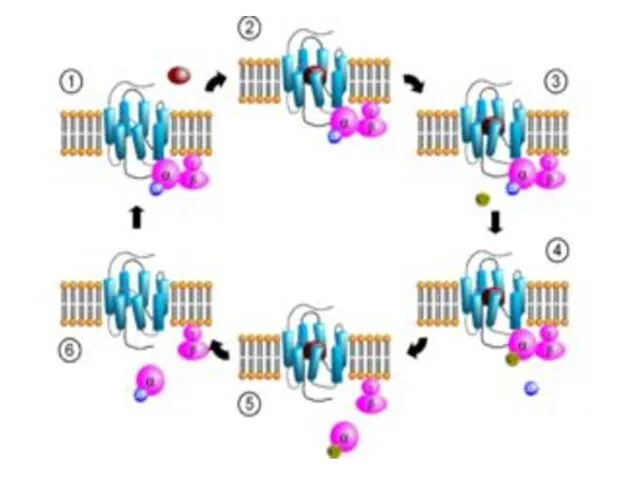
- 12. Лиганданың рецептормен байланысуы рецептордың конформациясын өзгертуге әкеледі және оның G-ақуызға байланысушылығы жоғарлайды (2).
- 13. Түзілген гормон-рецептор- G-ақуыз –ГДФ комплексі, G-ақуыздағы α-протомердің ГДФ-ке байланысушылық қабілетін төмендетіп ГТФ-ке байланысушылығын жоғарлатады. ГДФ ГТФ-ке
- 14. Бұл жағдай комплекстің α - суббірлік пен ГТФ және βγ - димер түзілуіне әкеледі. α -
- 15. α-суббірлік – ГТФ-тің нысанамен байланысуы α-суббірліктің ГТФазалық белсенділігін стимулдейді. α-протомердің белсенді орталығындағы ГТФ-тің дефосфорленуі оның нысана-молекулаға
- 16. Нысана ретінде, α-суббірлік – ГТФ комплексі үшін, мембранамен байланысқан ферменттер (аденилатциклаза, фосфолипаза С) және иондық каналдар
- 18. Мембранамен байланысқан рецепторлардың үш типі бар: Иондық каналдар G-ақуызбен өтетін Ферменттермен өтетін
- 19. Иондық каналдар рецепторлар ретінде. Лиганданың каналдық рецепторлық ақуыздарымен байланысуы қажетті иондардың енуі мен шығуына әкелетін, каналдың
- 20. Иондық каналдардың төрт типі белгілі. Кальций каналдары лигандамен басқарылатын каналға жатады. Иондық канал лигандамен байланысып, энергияның
- 21. Каналдардың ашылуы жоғары энергетикалық фосфаттың қосылуының арқасында жүреді. Б. Фосфорленумен басқарылады
- 22. Электрлік басқарылатын каналдар (В), мембраналық электрлік потенциалдың өзгеруі кезінде ашылады. Механикалық- каналдар жасуша мембранасына және цитоқаңқаға
- 23. Алғашқы мессенджерлер ақуыз белсенділігіне, сонымен қатар жасушаның белсенділігіне қалай әсер етеді? Бұл үшін сырттан келген сигнал
- 24. Екіншілік (қосымша) мессенджерлер (тасымалдаушылар) – бұл кішкентай молекулалар, рецептор активациясына жауап беретін кезде, жасушада жылдам және
- 25. Бес қосымша екінші мессенджерлер бар, жасушаішілік сигнализация процесінде үлкен маңызды роль атқарады. 1 – циклдік аденозинмонофосфат
- 26. цАМФ – бауырдағы көмірсулардың және май ұлпаларындағы үшглицеридтердің катаболизміне, бүйректегі сулардың реабсорбциясына, кальцийдің қалыпты алмасуына, стероидтық
- 27. Азот оксиді. Бұрыннан бері стенокардия кезінде нитроглицеріннің әсері белгілі, бірақ оның әсер ету механизмі 1990 жылы
- 28. Көптеген жасушаішілік сигналдық молекулалар молекулалық «ашып-жабатын-переключатель» сияқты жұмыс істейді. Олар белсенсіз ақуыздарды белсендендіреді немесе белсенділерді белсенсіздендіруі
- 30. Мысал ретінде цАМФ гликогеннің пайда болуына қалай әсер ететінін қарастырамыз. Сигналдың аденилатциклазалық жүйесі. Қауыпті немесе стрессті
- 31. Бауыр жасушасына адреналиннің әсері. Адреналин ағзада «fight or flight» (соғысу немесе қашу) деп аталатын эффектіні шақырады
- 32. Адреналиннің байланысуы рецептордың кеңістіктегі құрылымының өзгеруіне алып келеді, ол өз кезегінде жасушалық мембрананың ішкі беткейінде орналасқан
- 33. цАМФ-мен белсенген протеинкиназа тағы келесі ферментті – киназаны фосфорлайды. Киназа, гликогенфосфорилазаны фосфорлайды, нәтижесінде гликогеннен глюкоза молекуласын
- 34. Путь аденилатциклазы.Путь аденилатциклазы.exe
- 35. Оба ауруы мысалы : Оба вибрионының токсині ГТФ-тің деңгейіне және G-ақуыздың белсенділігіне әсер етеді. Нәтижесінде, Обамен
- 36. Қалыпты жағдайда фосфодиэстераза ферментінің әсерінен цАМФ жасушада жылдам инактивирленеді, циклдік емес АМФ айналады. Bordetella pertussis бактериясымен
- 37. Екінші жағдай. Бұл жағдайда молекулалық нысана қызметін фосфолипаза С атқарады (инозитолфосфаттық жүйе). Фосфолипаза С-ны (инозитолфосфаттық жүйені)
- 38. Кальцийдің жасушаішілік концентрациясына жүйке импульстарынан басқа да сигналдар әсер етеді. Мысалы, бұл цАМФ болуы мүмкін. Қандағы
- 39. Арнайы жасушадан тыс сигналдардың әсерінен және G-ақуыздардың бақылауымен екі молекула пайда болады – диацилглицерол және инозитолтрифосфат.
- 40. Сыртқы сигналға жауап беру үшін босап шыққан кальций ионы кальмодулинмен байланысады, содан соң киназа ферментімен байланысады
- 41. ФИФ -фосфотидил инозитолбисфосфат, ПКС- протеинкиназа С, ИФ-3- инозитол 3-фосфат, ДАГ- диацилглицирид, ФС-фосфотидилсерин
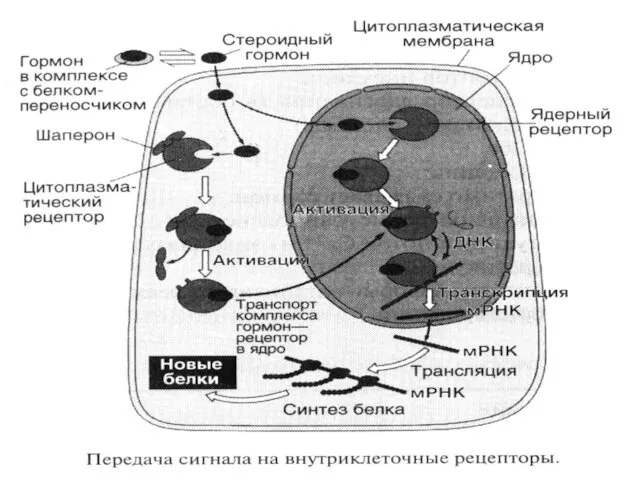
- 42. Мембранамен байланыспаған рецепторлармен сигналды беру механизмі. Жасушаішілік рецепторлар цитозолда басқа ақуыздармен кешене түзеді, немесе ядро мембраналарында
- 43. Цитоплазмалық рецептордың құрылысы Цитозолдық рецепторлар ірі ақуыздар шаперондармен комплекс түрінде болады. Бұл рецепторларда да үш қызметтік
- 44. Жасуша ішілік рецептор арқылы сигнал берілуінің этаптары (стероидтық гормондар мысалында). Гормонның рецептормен байланысуы шаперондардың босауына әкеледі.
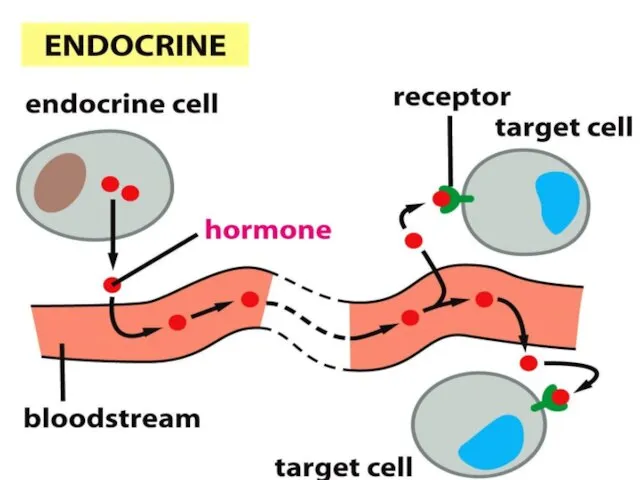
- 46. Жасушаға сигналдық молекулаларды жеткізу жолдары. Эндокриндік механизм – жасушааралық сигнализацияның көбірек зерттелінген тәсілі (гормон – эндокриндік
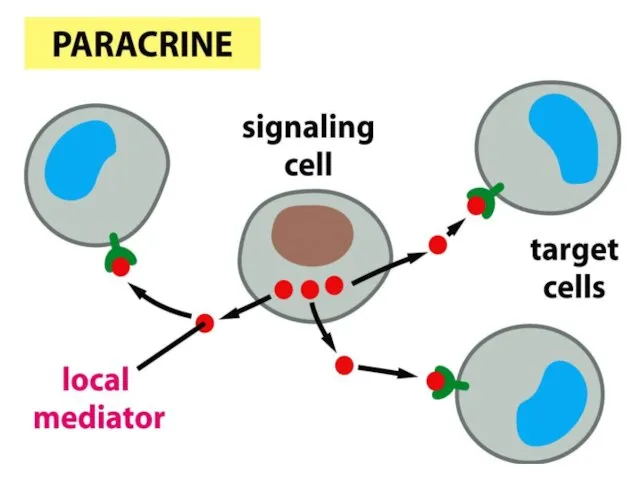
- 48. Паракриндік механизм – жасушалар сигналды молекулаларды жасап шығарады, олар сол ұлпалардағы жақын көрші жасушаларды белсендіреді. Сигналды
- 50. Аутокриндік механизм – бұл жергілікті (локалді) қатынас –шектелген сигналдарға жауап беру механизмі. Жасуша өзіне меншікті сигналға
- 51. Юкстакриндік сигналды жүйе – жасушаның жапсырылу процесіне қатысады (қан жасушалары бір-бірімен, немесе қабыну кезінде тамыр жүйесінің
- 54. Скачать презентацию

Жасушааралық сигнализация және интеграцияның молекулалық механизмдері.
Жасуша ерекше сигнал
жауабына молекулалық каскадтық
Жасушааралық сигнализация және интеграцияның молекулалық механизмдері.
Жасуша ерекше сигнал
жауабына молекулалық каскадтық

Жасушадан тыс сигналдар химиялық және физикалық болып бөлінеді.
Химиялық сигналдар: аминокишқылдар, ақуыздар,
Жасушадан тыс сигналдар химиялық және физикалық болып бөлінеді.
Химиялық сигналдар: аминокишқылдар, ақуыздар,

Барлық сигналдық молекулалар 3 классқа бөлінеді:
Гормондар. Ішкі секреция бездерінде синтезделеді,
Барлық сигналдық молекулалар 3 классқа бөлінеді:
Гормондар. Ішкі секреция бездерінде синтезделеді,

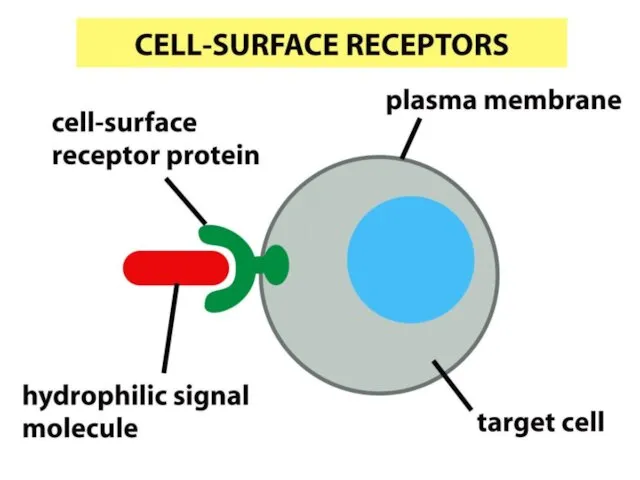
Жасушалық жауаптың ерекшелігі рецептор түрімен анықталынады, өйткені жасушаларда рецепторлар жиынтығы бар,
Жасушалық жауаптың ерекшелігі рецептор түрімен анықталынады, өйткені жасушаларда рецепторлар жиынтығы бар,
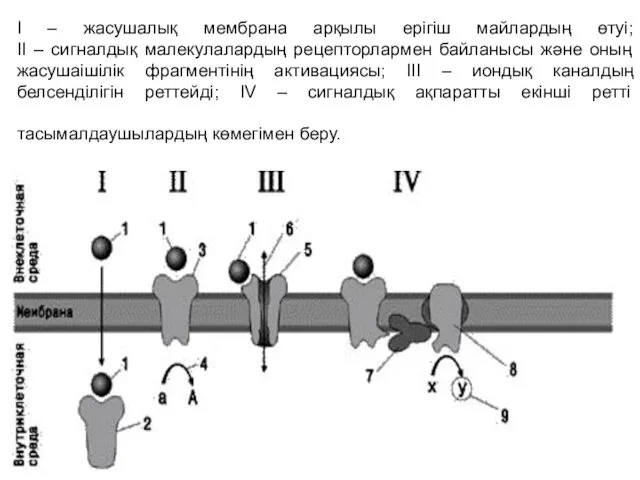
I – жасушалық мембрана арқылы ерігіш майлардың өтуі;
II – сигналдық малекулалардың
I – жасушалық мембрана арқылы ерігіш майлардың өтуі; II – сигналдық малекулалардың
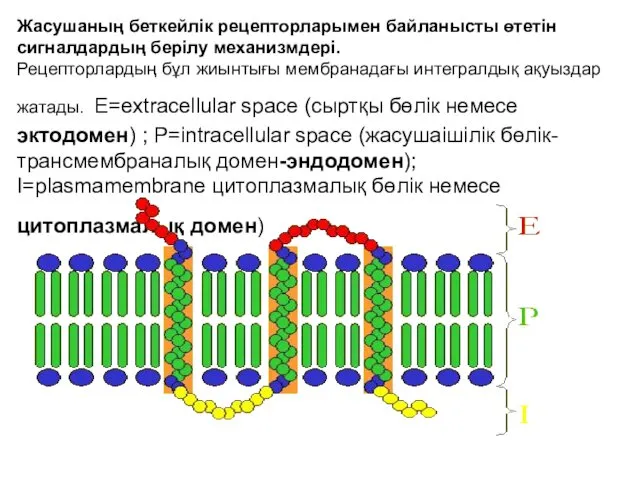
Жасушаның беткейлік рецепторларымен байланысты өтетін сигналдардың берілу механизмдері.
Рецепторлардың бұл жиынтығы
Жасушаның беткейлік рецепторларымен байланысты өтетін сигналдардың берілу механизмдері. Рецепторлардың бұл жиынтығы

Сигналды берудің жалпы кестесінің 5 негізгі этаптары бар.
Сигналды берудің бұзылуы әрбір
Сигналды берудің жалпы кестесінің 5 негізгі этаптары бар.
Сигналды берудің бұзылуы әрбір

4 этап- эффектор активациясы бұл G-ақуыздың - альфа суббірлігі.
5 этап –
4 этап- эффектор активациясы бұл G-ақуыздың - альфа суббірлігі.
5 этап –

G-ақуыздар – бұлар 3 суббірліктен
тұратын тримерлер. Бұл ақуыздар
мембрана құрамына
G-ақуыздар – бұлар 3 суббірліктен
тұратын тримерлер. Бұл ақуыздар
мембрана құрамына
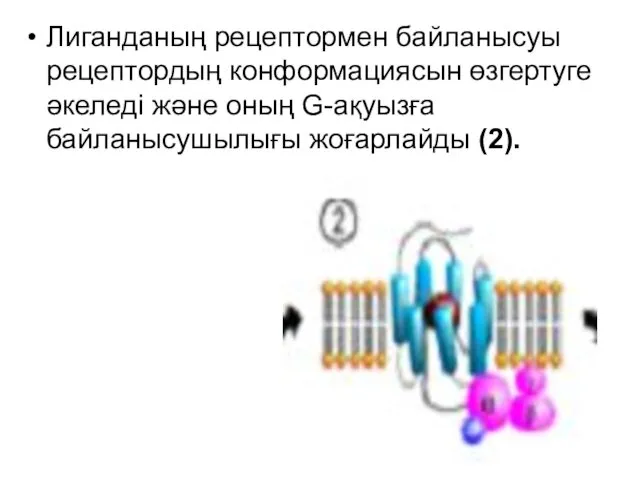
Лиганданың рецептормен байланысуы рецептордың конформациясын өзгертуге әкеледі және оның G-ақуызға байланысушылығы
Лиганданың рецептормен байланысуы рецептордың конформациясын өзгертуге әкеледі және оның G-ақуызға байланысушылығы
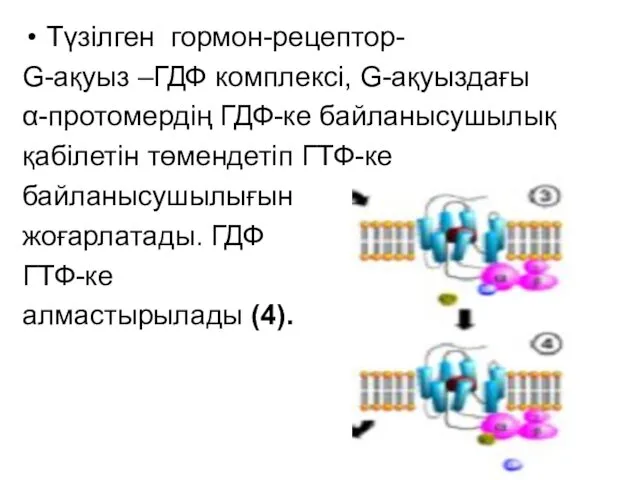
Түзілген гормон-рецептор-
G-ақуыз –ГДФ комплексі, G-ақуыздағы
α-протомердің ГДФ-ке байланысушылық
қабілетін төмендетіп
Түзілген гормон-рецептор-
G-ақуыз –ГДФ комплексі, G-ақуыздағы
α-протомердің ГДФ-ке байланысушылық
қабілетін төмендетіп
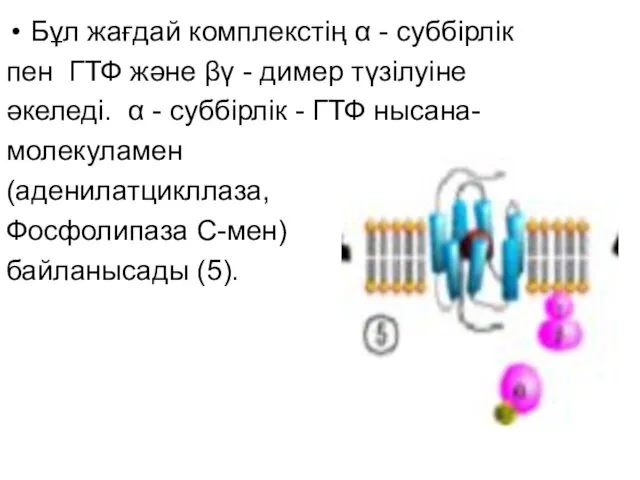
Бұл жағдай комплекстің α - суббірлік
пен ГТФ және βγ -
Бұл жағдай комплекстің α - суббірлік
пен ГТФ және βγ -

α-суббірлік – ГТФ-тің нысанамен
байланысуы α-суббірліктің ГТФазалық
белсенділігін стимулдейді. α-протомердің
α-суббірлік – ГТФ-тің нысанамен
байланысуы α-суббірліктің ГТФазалық
белсенділігін стимулдейді. α-протомердің

Нысана ретінде, α-суббірлік – ГТФ комплексі үшін, мембранамен байланысқан ферменттер (аденилатциклаза,
Нысана ретінде, α-суббірлік – ГТФ комплексі үшін, мембранамен байланысқан ферменттер (аденилатциклаза,

Мембранамен байланысқан рецепторлардың үш типі бар:
Иондық каналдар
G-ақуызбен өтетін
Ферменттермен өтетін
Мембранамен байланысқан рецепторлардың үш типі бар:
Иондық каналдар
G-ақуызбен өтетін
Ферменттермен өтетін

Иондық каналдар рецепторлар ретінде.
Лиганданың каналдық рецепторлық ақуыздарымен байланысуы қажетті иондардың
Иондық каналдар рецепторлар ретінде.
Лиганданың каналдық рецепторлық ақуыздарымен байланысуы қажетті иондардың
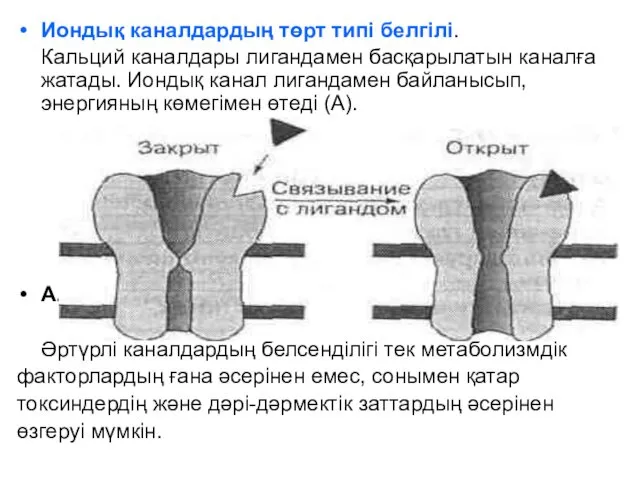
Иондық каналдардың төрт типі белгілі.
Кальций каналдары лигандамен басқарылатын каналға жатады.
Иондық каналдардың төрт типі белгілі.
Кальций каналдары лигандамен басқарылатын каналға жатады.
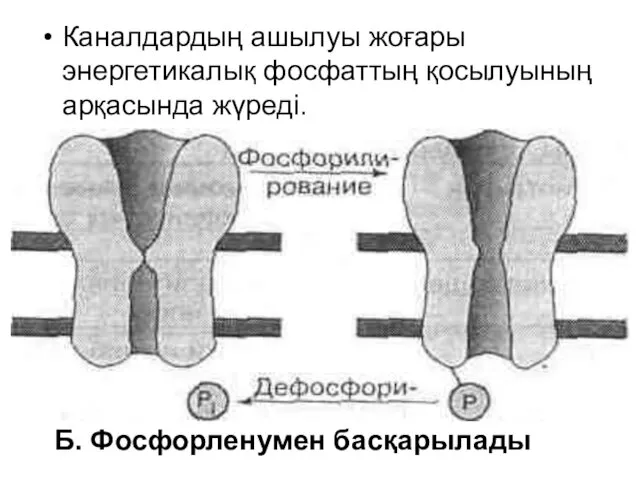
Каналдардың ашылуы жоғары энергетикалық фосфаттың қосылуының арқасында жүреді.
Б. Фосфорленумен басқарылады
Каналдардың ашылуы жоғары энергетикалық фосфаттың қосылуының арқасында жүреді.
Б. Фосфорленумен басқарылады
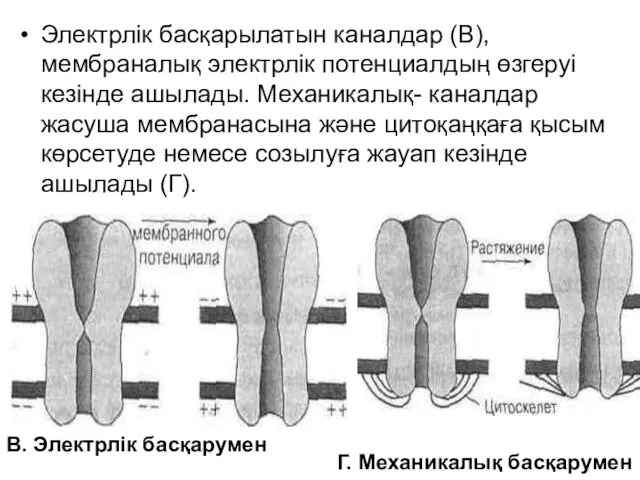
Электрлік басқарылатын каналдар (В), мембраналық электрлік потенциалдың өзгеруі кезінде ашылады. Механикалық-
Электрлік басқарылатын каналдар (В), мембраналық электрлік потенциалдың өзгеруі кезінде ашылады. Механикалық-

Алғашқы мессенджерлер ақуыз белсенділігіне, сонымен қатар жасушаның белсенділігіне қалай әсер етеді?
Бұл
Алғашқы мессенджерлер ақуыз белсенділігіне, сонымен қатар жасушаның белсенділігіне қалай әсер етеді?
Бұл

Екіншілік (қосымша) мессенджерлер
(тасымалдаушылар) – бұл кішкентай
молекулалар, рецептор активациясына
жауап
Екіншілік (қосымша) мессенджерлер
(тасымалдаушылар) – бұл кішкентай
молекулалар, рецептор активациясына
жауап

Бес қосымша екінші мессенджерлер бар, жасушаішілік сигнализация процесінде үлкен маңызды роль
Бес қосымша екінші мессенджерлер бар, жасушаішілік сигнализация процесінде үлкен маңызды роль

цАМФ – бауырдағы көмірсулардың және май ұлпаларындағы үшглицеридтердің катаболизміне, бүйректегі сулардың
цАМФ – бауырдағы көмірсулардың және май ұлпаларындағы үшглицеридтердің катаболизміне, бүйректегі сулардың

Азот оксиді. Бұрыннан бері стенокардия кезінде нитроглицеріннің әсері белгілі, бірақ оның
Азот оксиді. Бұрыннан бері стенокардия кезінде нитроглицеріннің әсері белгілі, бірақ оның

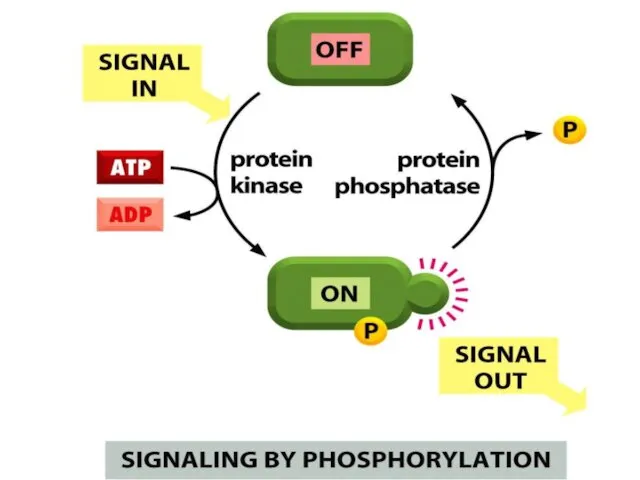
Көптеген жасушаішілік сигналдық молекулалар молекулалық «ашып-жабатын-переключатель» сияқты жұмыс істейді. Олар белсенсіз
Көптеген жасушаішілік сигналдық молекулалар молекулалық «ашып-жабатын-переключатель» сияқты жұмыс істейді. Олар белсенсіз

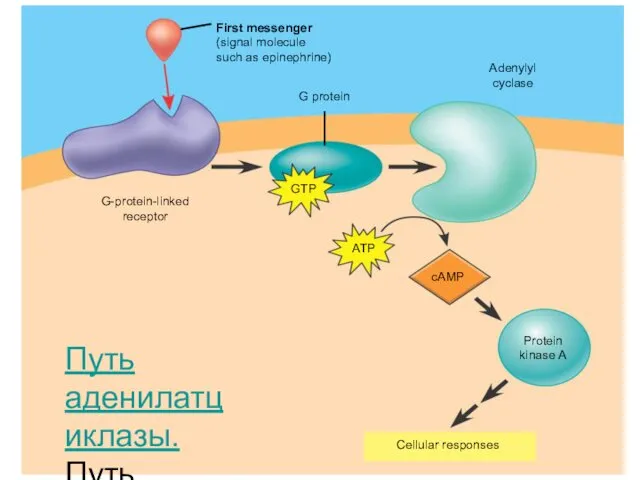
Мысал ретінде цАМФ гликогеннің пайда болуына қалай әсер ететінін қарастырамыз.
Сигналдың аденилатциклазалық
Мысал ретінде цАМФ гликогеннің пайда болуына қалай әсер ететінін қарастырамыз.
Сигналдың аденилатциклазалық

Бауыр жасушасына адреналиннің әсері. Адреналин ағзада «fight or flight» (соғысу немесе
Бауыр жасушасына адреналиннің әсері. Адреналин ағзада «fight or flight» (соғысу немесе

Адреналиннің байланысуы рецептордың кеңістіктегі құрылымының өзгеруіне алып келеді, ол өз кезегінде
Адреналиннің байланысуы рецептордың кеңістіктегі құрылымының өзгеруіне алып келеді, ол өз кезегінде

цАМФ-мен белсенген протеинкиназа тағы келесі ферментті – киназаны фосфорлайды.
Киназа, гликогенфосфорилазаны
фосфорлайды,
цАМФ-мен белсенген протеинкиназа тағы келесі ферментті – киназаны фосфорлайды.
Киназа, гликогенфосфорилазаны
фосфорлайды,
Путь аденилатциклазы.Путь аденилатциклазы.exe
Путь аденилатциклазы.Путь аденилатциклазы.exe

Оба ауруы мысалы :
Оба вибрионының токсині ГТФ-тің деңгейіне және G-ақуыздың
Оба ауруы мысалы :
Оба вибрионының токсині ГТФ-тің деңгейіне және G-ақуыздың

Қалыпты жағдайда фосфодиэстераза ферментінің әсерінен цАМФ жасушада жылдам инактивирленеді, циклдік емес
Қалыпты жағдайда фосфодиэстераза ферментінің әсерінен цАМФ жасушада жылдам инактивирленеді, циклдік емес

Екінші жағдай. Бұл жағдайда молекулалық нысана қызметін фосфолипаза С атқарады (инозитолфосфаттық
Екінші жағдай. Бұл жағдайда молекулалық нысана қызметін фосфолипаза С атқарады (инозитолфосфаттық

Кальцийдің жасушаішілік концентрациясына жүйке импульстарынан басқа да сигналдар әсер етеді. Мысалы,
Кальцийдің жасушаішілік концентрациясына жүйке импульстарынан басқа да сигналдар әсер етеді. Мысалы,

Арнайы жасушадан тыс сигналдардың әсерінен және G-ақуыздардың бақылауымен екі молекула пайда
Арнайы жасушадан тыс сигналдардың әсерінен және G-ақуыздардың бақылауымен екі молекула пайда

Сыртқы сигналға жауап беру үшін босап шыққан кальций ионы кальмодулинмен байланысады,
Сыртқы сигналға жауап беру үшін босап шыққан кальций ионы кальмодулинмен байланысады,
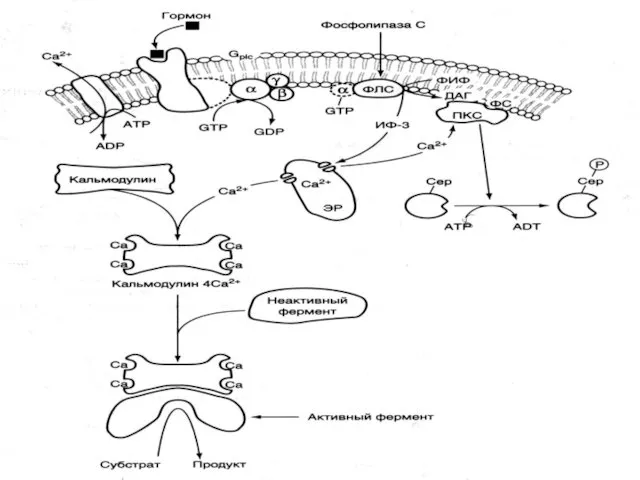
ФИФ -фосфотидил инозитолбисфосфат, ПКС- протеинкиназа С, ИФ-3- инозитол 3-фосфат, ДАГ- диацилглицирид,
ФИФ -фосфотидил инозитолбисфосфат, ПКС- протеинкиназа С, ИФ-3- инозитол 3-фосфат, ДАГ- диацилглицирид,

Мембранамен байланыспаған рецепторлармен сигналды беру механизмі.
Жасушаішілік рецепторлар цитозолда басқа ақуыздармен кешене
Мембранамен байланыспаған рецепторлармен сигналды беру механизмі.
Жасушаішілік рецепторлар цитозолда басқа ақуыздармен кешене

Цитоплазмалық рецептордың құрылысы
Цитозолдық рецепторлар ірі ақуыздар шаперондармен комплекс түрінде болады.
Цитоплазмалық рецептордың құрылысы Цитозолдық рецепторлар ірі ақуыздар шаперондармен комплекс түрінде болады.

Жасуша ішілік рецептор арқылы сигнал берілуінің этаптары (стероидтық гормондар мысалында).
Гормонның рецептормен
Жасуша ішілік рецептор арқылы сигнал берілуінің этаптары (стероидтық гормондар мысалында). Гормонның рецептормен
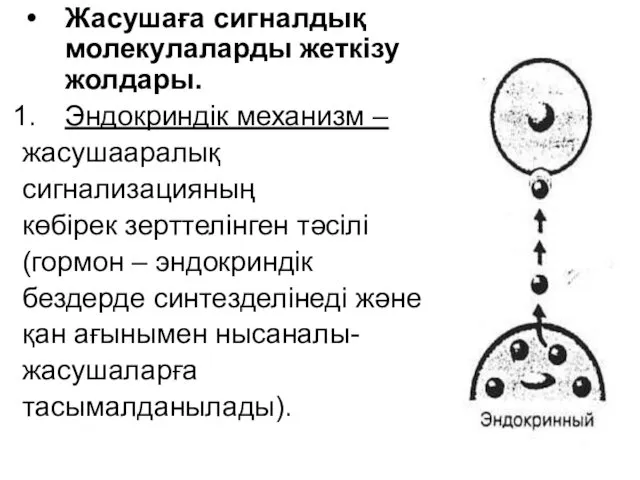
Жасушаға сигналдық молекулаларды жеткізу жолдары.
Эндокриндік механизм –
жасушааралық
сигнализацияның
көбірек зерттелінген
Жасушаға сигналдық молекулаларды жеткізу жолдары.
Эндокриндік механизм –
жасушааралық
сигнализацияның
көбірек зерттелінген

Паракриндік механизм –
жасушалар сигналды
молекулаларды жасап
шығарады, олар сол
ұлпалардағы жақын
Паракриндік механизм –
жасушалар сигналды
молекулаларды жасап
шығарады, олар сол
ұлпалардағы жақын

Аутокриндік механизм –
бұл жергілікті (локалді)
қатынас –шектелген
сигналдарға жауап
беру
Аутокриндік механизм –
бұл жергілікті (локалді)
қатынас –шектелген
сигналдарға жауап
беру
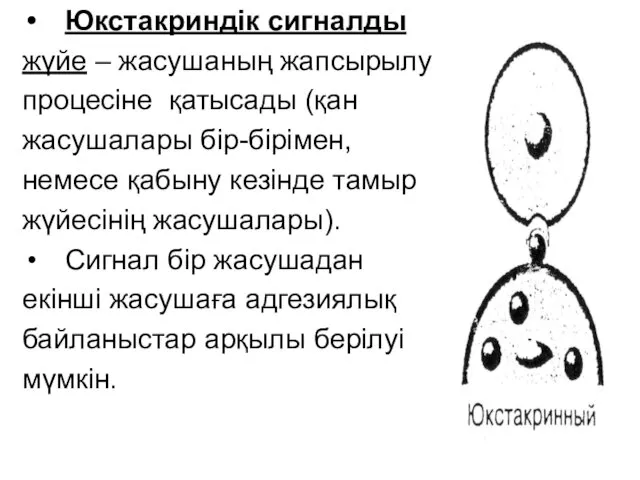
Юкстакриндік сигналды
жүйе – жасушаның жапсырылу
процесіне қатысады (қан
жасушалары бір-бірімен,
Юкстакриндік сигналды
жүйе – жасушаның жапсырылу
процесіне қатысады (қан
жасушалары бір-бірімен,
 Постэмбриональное развитие насекомых
Постэмбриональное развитие насекомых Семейство Бобовые
Семейство Бобовые Общий путь катаболизма. (Лекция 9)
Общий путь катаболизма. (Лекция 9) Клеточная оболочка
Клеточная оболочка Ткани растений
Ткани растений Бактерии. Их роль в природе и жизни человека
Бактерии. Их роль в природе и жизни человека Как прекрасен мир бабочек
Как прекрасен мир бабочек Жасыл балдырлар бөлімі – Chlorophyta
Жасыл балдырлар бөлімі – Chlorophyta Заповедник Нургуш - жемчужина Вятского края
Заповедник Нургуш - жемчужина Вятского края Знакомство с лисой
Знакомство с лисой Презентация к уроку Жгутиконосцы. Инфузории. 7 класс
Презентация к уроку Жгутиконосцы. Инфузории. 7 класс Разнообразие пасленовых (акцент на цветки)
Разнообразие пасленовых (акцент на цветки) Самоедские собаки
Самоедские собаки Функциональная анатомия костей туловища. Соединения костей туловища
Функциональная анатомия костей туловища. Соединения костей туловища Трихинеллез. Устойчивость возбудителя. Лабораторная диагностика
Трихинеллез. Устойчивость возбудителя. Лабораторная диагностика Черника на подоконнике
Черника на подоконнике Биотехнология растений. Трансгенные растения (часть 2)
Биотехнология растений. Трансгенные растения (часть 2) Мутації проти варіацій в еволюції
Мутації проти варіацій в еволюції Физиология мышечного сокращения
Физиология мышечного сокращения Класс Пресмыкающиеся. Особенности внешнего и внутреннего строения. Многообразие. Происхождение
Класс Пресмыкающиеся. Особенности внешнего и внутреннего строения. Многообразие. Происхождение Презентация Физминутки на уроках биологии, химии
Презентация Физминутки на уроках биологии, химии Органические вещества. Углеводы моносахариды
Органические вещества. Углеводы моносахариды Конформация биомакромолекул. Виды конформационных перестроек
Конформация биомакромолекул. Виды конформационных перестроек Понятие о катаболизме и анаболизме. Основы питания. Незаменимые пищевые факторы. Биоэнергетика. Структурная организация ЦПЭ
Понятие о катаболизме и анаболизме. Основы питания. Незаменимые пищевые факторы. Биоэнергетика. Структурная организация ЦПЭ История возникновения и развития живого на Земле
История возникновения и развития живого на Земле Элементарные факторы эволюции
Элементарные факторы эволюции Палеонтологические свидетельства эволюции
Палеонтологические свидетельства эволюции Происхождение рыб
Происхождение рыб