- Жизненный цикл клетки. Апоптоз. Старение клетки

Содержание
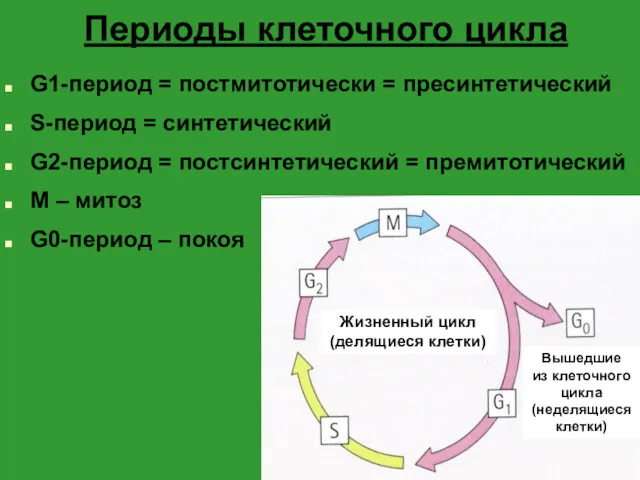
- 2. Периоды клеточного цикла G1-период = постмитотически = пресинтетический S-период = синтетический G2-период = постсинтетический = премитотический
- 3. G1-период начинается сразу после образования клетки в результате митоза содержание ДНК в ядре постоянно – 2n
- 4. S-период репликация молекулы ДНК (кроме центромерных участков) – 4n удвоения количества хромосомных белков дупликация центриолей
- 5. G2-период синтез ряда веществ, необходимых для прохождения митоза в т.ч. тубулина – белка микротрубочек веретена деления
- 6. Типы клеток по способности к делению 1) митотические (делящиеся) клетки: клетки базального слоя эпителия гемопоэтические клетки
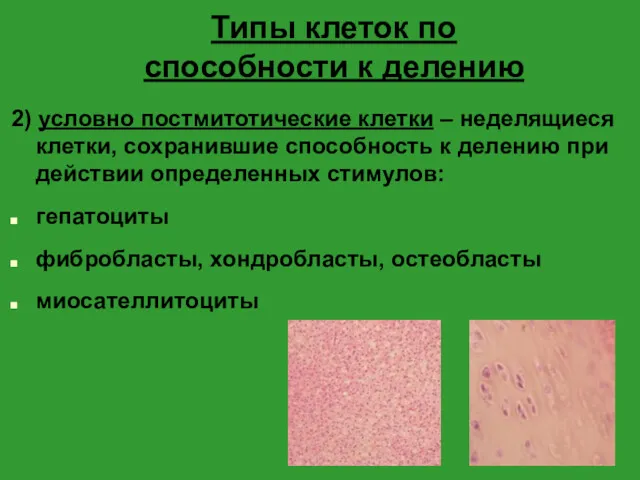
- 7. Типы клеток по способности к делению 2) условно постмитотические клетки – неделящиеся клетки, сохранившие способность к
- 8. Типы клеток по способности к делению 3) постмитотические клетки – неделящиеся клетки, окончательно потерявшие способность делиться:
- 9. Популяции клеток стационарные популяции: клетки мозга симпласты скелетной мышцы обновляющиеся популяции: клетки эпителия гемопоэтические клетки растущие
- 10. Выход клетки из митотического цикла M → G1 ↔ G0 ↔ G0 (D1) → G0 (D2)
- 11. Регуляция клеточного цикла циклинзависимые киназы (Cdks) – ключевая роль в смене фаз клеточного цикла Cdks –
- 12. Регуляция клеточного цикла ранняя стадия G1 – циклин D-Cdk4(6) поздняя стадия G1 – циклин Е-Cdk2 ранняя
- 13. Выход клетки из митоза комплекс MPF: максимальная активность – в метафазу митоза фосфорилирует фактор АРС –
- 14. Внеклеточные регуляторы пролиферации действие митогенов – факторы роста, некоторые гормоны, интерлейкины действие антимитогенов – фактор некроза
- 15. КЛЕТОЧНЫЙ ЦИКЛ КЛЕТОЧНЫЙ ЦИКЛ
- 16. Митоз от греч. Mítos – нить = кариокинез = непрямое деление клетки наиболее распространённый способ воспроизведения
- 18. Профаза в ядре: конденсация хромосом каждая хромосома состоит из двух хроматид, связанных во многих местах белками-когезинами
- 19. Профаза в цитоплазме: ЭПС и комплекс Гольджи распадаются на везикулы 2 диплосомы постепенно расходятся к полюсам
- 20. Метафаза хромосомы достигают максимальной степени конденсации хромосомы выстраиваются в экваториальной плоскости клетки – метафазная пластинка или
- 21. Метафаза микротрубочки веретена деления: 1) кинетохорные – связывают каждую хроматиду с одной из диплосом 2) полярные
- 22. Анафаза самая короткая стадия митоза хроматиды расходятся к полюсам клетки хроматиды ориентированы центромерными участками к полюсу,
- 23. Анафаза движение хромосом (2 причины): 1) изменение длины МТ веретена деления укорочение кинетохорных МТ удлинение полярных
- 24. Телофаза с хромосомами ассоциируются пузырьки кариолеммы в их стенки встраиваются комплексы ядерных пор через них в
- 25. Цитокинез происходит в поздней телофазе по экватору клетки формируется актиномиозиновое кольцо постепенно сжимаясь, кольцо стягивает плазмолемму,
- 26. Результаты цитокинеза: 1) Органеллы равномерно распределены между соседними клетками. 2) Сразу после деления дочерние клетки входят
- 28. Продолжительность митоза Продолжительность митоза зависит от: размеров клеток их плоидности числа ядер условий окружающей среды (температура)
- 29. Пролиферация клеток и апоптоз Количество клеток в ткани регулируется двумя процессами – пролиферацией клеток и «программированной,
- 30. Апоптоз Апоптоз – генетически запрограммированная гибель клеток, которая приводит к "аккуратной" разборке и удалению клеток Морфологическими
- 31. Апоптоз от греч. – опадание листьев
- 32. Причины апоптоза две группы: «неудовлетворительное» состояние клетки («апоптоз изнутри») «негативная» сигнализация снаружи, передающаяся через специальные рецепторы
- 33. «Апоптоз изнутри» повреждение хромосом: многочисленные разрывы ДНК нарушение конформации ДНК сшивки между цепями ДНК неправильная сегрегация
- 34. Причины повреждений разные виды облучений изменения температуры химические соединения нарушение питания клетки деление клетки повреждение клеточных
- 35. «Апоптоз по команде» клетка вполне жизнеспособна, но, с позиций целостного организма, она является ненужной или даже
- 36. «Апоптоз по команде» 2) при формировании и функционировании иммунной системы гибель аутореактивных клонов лимфоцитов гибель стимулированных
- 37. «Апоптоз по команде» 4) при функционировании женской половой системы гибель клеток атрезирующих фолликулов гибель клеток редуцирующего
- 38. Регуляция апоптоза одним из важнейших инструментов апоптоза является семейство цитоплазматических протеаз – каспаз каспазы способны в
- 39. Регуляция апоптоза мишени каспаз: некоторые белки цитоскелета ( → изменение формы клетки – появление инвагинаций и
- 40. Митохондриальные факторы апоптоза протеаза AIF и цитохром с – активаторы каспазного каскада в митохондриях – специальные
- 41. Белок р53 = транскрипционный фактор активируется в ответ на разнообразные повреждения клеточной структуры: нерепарированные разрывы и
- 42. Белок р53 активирует гены: ряда рецепторов, воспринимающих команду об апоптозе отвечающие за остановку клеточного деления белка
- 43. Итоги действия белка р53 клетка: либо задерживается на той или иной стадии митотического цикла и исправляет
- 44. Морфология апоптоза 1) конденсация хроматина и некоторое сжатие клетки (из-за конденсации цитоплазмы) хроматин приобретает вид плотных
- 45. Морфология апоптоза 2) фрагментация ядра ядро распадается на отдельные фрагменты, окруженные ядерной оболочкой и содержащие очень
- 46. Морфология апоптоза 3) изменение формы клетки на поверхности клетки появляются инвагинации цитоплазма приобретает вид лопастей с
- 47. Морфология апоптоза 4) фагоцитоз апоптозных телец окружающими клетками к фагоцитозу апоптозных телец способны не только фагоциты,
- 49. Сравнительная характеристика апоптоза и некроза
- 50. Старение клетки с возрастом прогрессивно страдает ряд функций клеток: снижаются окислительное фосфорилирование в митохондриях снижается синтез
- 51. Теломерная теория старения сформулирована А.М. Оловниковым в 1971 г во всех соматических клетках организма механизм репликации
- 52. Факты, подтверждающие теорию при каждом делении клетки длина теломерных участков сокращается на 50-100 н.п. в половых
- 53. Дополнительные предположения Как укорочение длины теломер влияет на клетку? достаточно длинные теломеры вызывают репрессию активности прилежащих
- 54. Морфологические изменения клетки при старении неправильные и дольчатые ядра полиморфные вакуолизированные митохондрий уменьшение эндоплазматического ретикулума деформация
- 56. Скачать презентацию
Периоды клеточного цикла
G1-период = постмитотически = пресинтетический
S-период = синтетический
G2-период = постсинтетический
Периоды клеточного цикла
G1-период = постмитотически = пресинтетический
S-период = синтетический
G2-период = постсинтетический

G1-период
начинается сразу после образования клетки в результате митоза
содержание ДНК в ядре
G1-период
начинается сразу после образования клетки в результате митоза
содержание ДНК в ядре
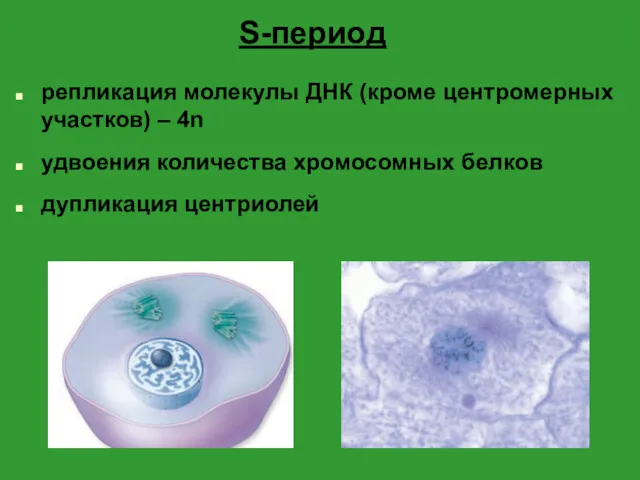
S-период
репликация молекулы ДНК (кроме центромерных участков) – 4n
удвоения количества хромосомных белков
дупликация
S-период
репликация молекулы ДНК (кроме центромерных участков) – 4n
удвоения количества хромосомных белков
дупликация
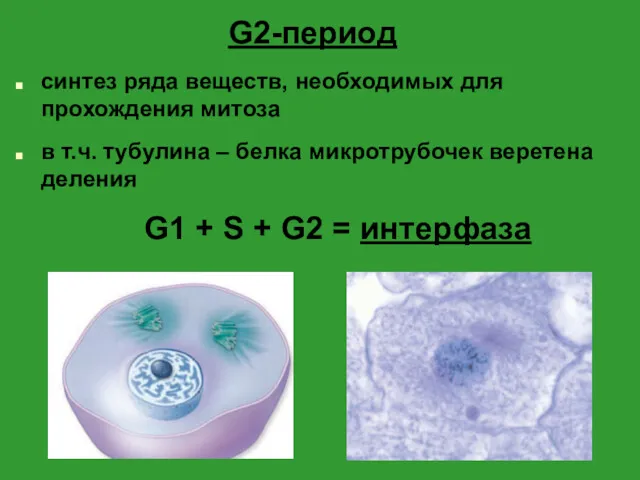
G2-период
синтез ряда веществ, необходимых для прохождения митоза
в т.ч. тубулина – белка
G2-период
синтез ряда веществ, необходимых для прохождения митоза
в т.ч. тубулина – белка
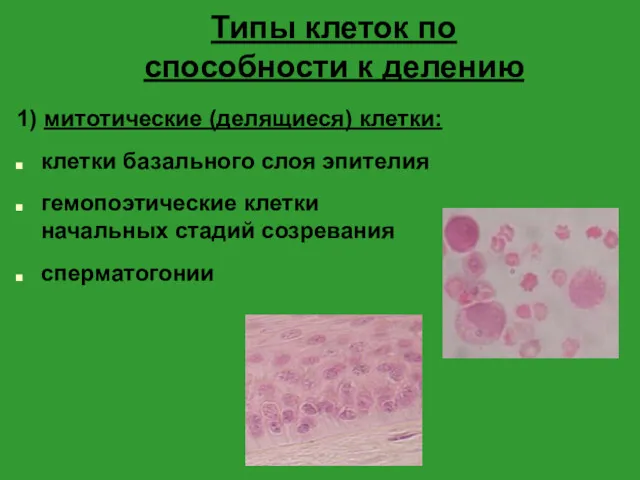
Типы клеток по способности к делению
1) митотические (делящиеся) клетки:
клетки базального слоя
Типы клеток по способности к делению
1) митотические (делящиеся) клетки:
клетки базального слоя
Типы клеток по способности к делению
2) условно постмитотические клетки – неделящиеся
Типы клеток по способности к делению
2) условно постмитотические клетки – неделящиеся

Типы клеток по способности к делению
3) постмитотические клетки – неделящиеся клетки,
Типы клеток по способности к делению
3) постмитотические клетки – неделящиеся клетки,

Популяции клеток
стационарные популяции:
клетки мозга
симпласты скелетной мышцы
обновляющиеся популяции:
клетки эпителия
гемопоэтические клетки
растущие популяции:
клетки желез
клетки
Популяции клеток
стационарные популяции:
клетки мозга
симпласты скелетной мышцы
обновляющиеся популяции:
клетки эпителия
гемопоэтические клетки
растущие популяции:
клетки желез
клетки

Выход клетки из митотического цикла
M → G1 ↔ G0 ↔
Выход клетки из митотического цикла
M → G1 ↔ G0 ↔

Регуляция клеточного цикла
циклинзависимые киназы (Cdks) – ключевая роль в смене фаз
Регуляция клеточного цикла
циклинзависимые киназы (Cdks) – ключевая роль в смене фаз

Регуляция клеточного цикла
ранняя стадия G1 – циклин D-Cdk4(6)
поздняя стадия G1 –
Регуляция клеточного цикла
ранняя стадия G1 – циклин D-Cdk4(6)
поздняя стадия G1 –

Выход клетки из митоза
комплекс MPF:
максимальная активность – в метафазу митоза
фосфорилирует
Выход клетки из митоза
комплекс MPF:
максимальная активность – в метафазу митоза
фосфорилирует

Внеклеточные регуляторы пролиферации
действие митогенов – факторы роста, некоторые гормоны, интерлейкины
действие антимитогенов
Внеклеточные регуляторы пролиферации
действие митогенов – факторы роста, некоторые гормоны, интерлейкины
действие антимитогенов
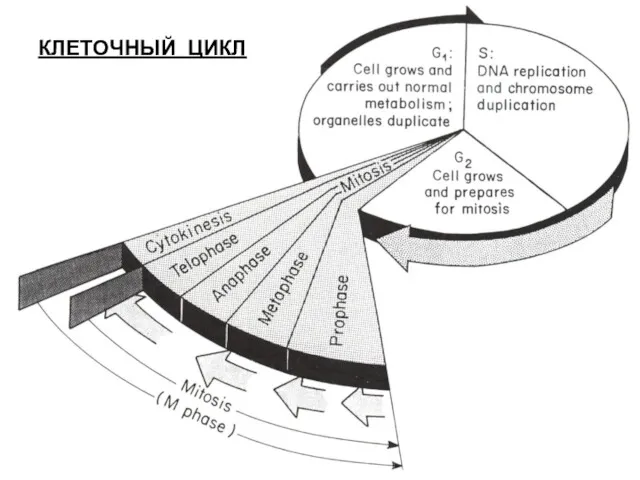
КЛЕТОЧНЫЙ ЦИКЛ
КЛЕТОЧНЫЙ ЦИКЛ
КЛЕТОЧНЫЙ ЦИКЛ
КЛЕТОЧНЫЙ ЦИКЛ

Митоз
от греч. Mítos – нить
= кариокинез = непрямое деление клетки
наиболее распространённый
Митоз
от греч. Mítos – нить
= кариокинез = непрямое деление клетки
наиболее распространённый

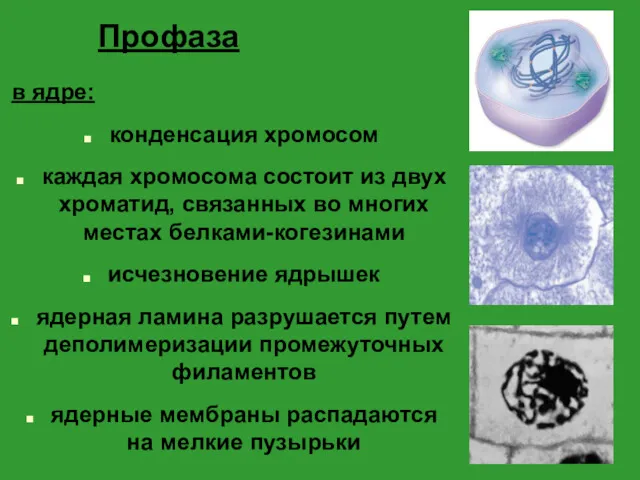
Профаза
в ядре:
конденсация хромосом
каждая хромосома состоит из двух хроматид, связанных во многих
Профаза
в ядре:
конденсация хромосом
каждая хромосома состоит из двух хроматид, связанных во многих
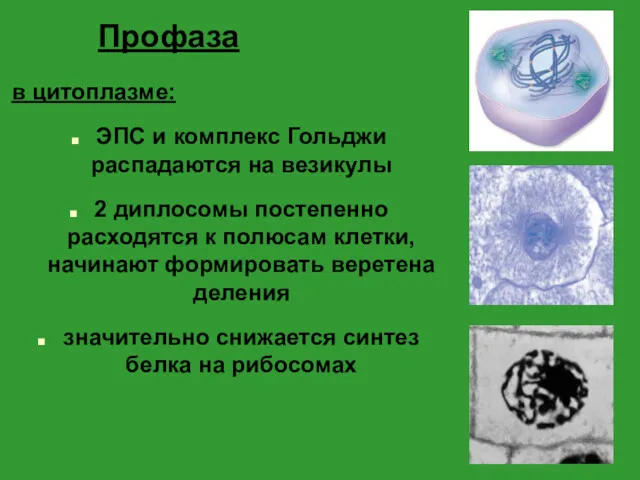
Профаза
в цитоплазме:
ЭПС и комплекс Гольджи распадаются на везикулы
2 диплосомы постепенно расходятся
Профаза
в цитоплазме:
ЭПС и комплекс Гольджи распадаются на везикулы
2 диплосомы постепенно расходятся
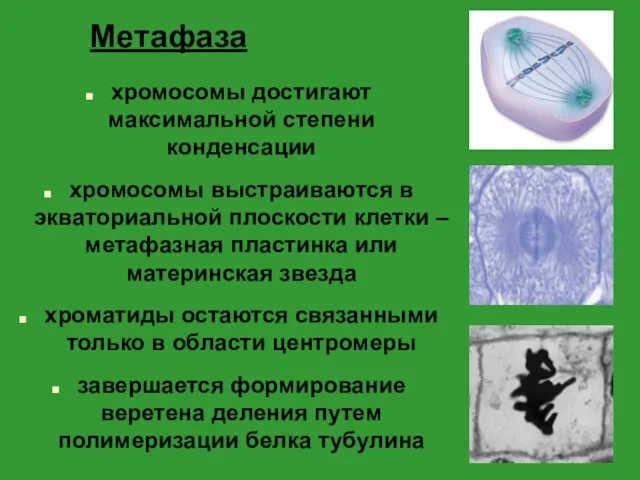
Метафаза
хромосомы достигают максимальной степени конденсации
хромосомы выстраиваются в экваториальной плоскости клетки –
Метафаза
хромосомы достигают максимальной степени конденсации
хромосомы выстраиваются в экваториальной плоскости клетки –

Метафаза
микротрубочки веретена деления:
1) кинетохорные – связывают каждую хроматиду с одной из
Метафаза
микротрубочки веретена деления:
1) кинетохорные – связывают каждую хроматиду с одной из
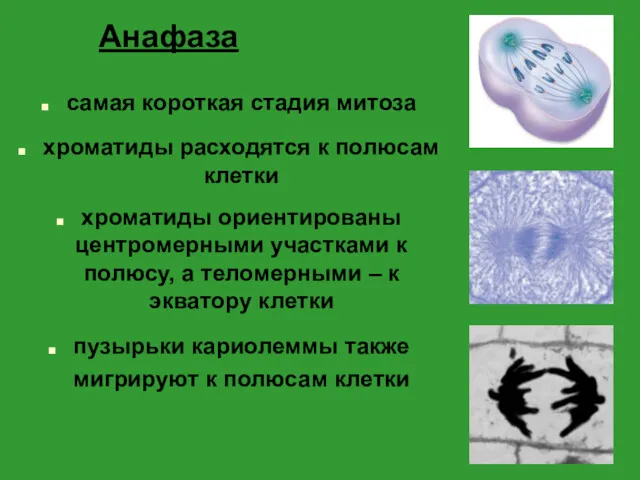
Анафаза
самая короткая стадия митоза
хроматиды расходятся к полюсам клетки
хроматиды ориентированы центромерными участками
Анафаза
самая короткая стадия митоза
хроматиды расходятся к полюсам клетки
хроматиды ориентированы центромерными участками
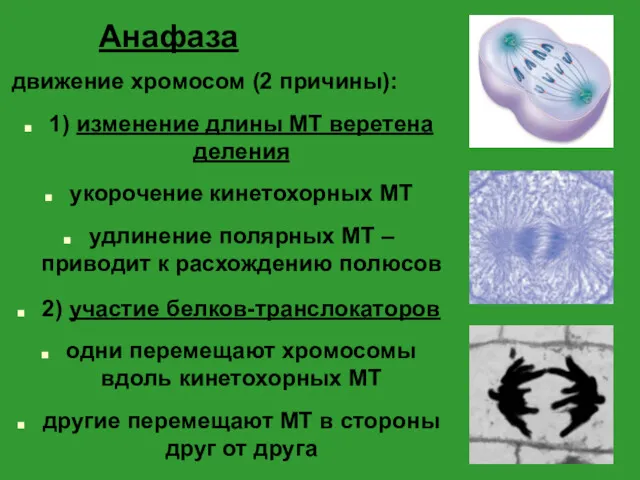
Анафаза
движение хромосом (2 причины):
1) изменение длины МТ веретена деления
укорочение кинетохорных МТ
удлинение
Анафаза
движение хромосом (2 причины):
1) изменение длины МТ веретена деления
укорочение кинетохорных МТ
удлинение

Телофаза
с хромосомами ассоциируются пузырьки кариолеммы
в их стенки встраиваются комплексы ядерных пор
через
Телофаза
с хромосомами ассоциируются пузырьки кариолеммы
в их стенки встраиваются комплексы ядерных пор
через
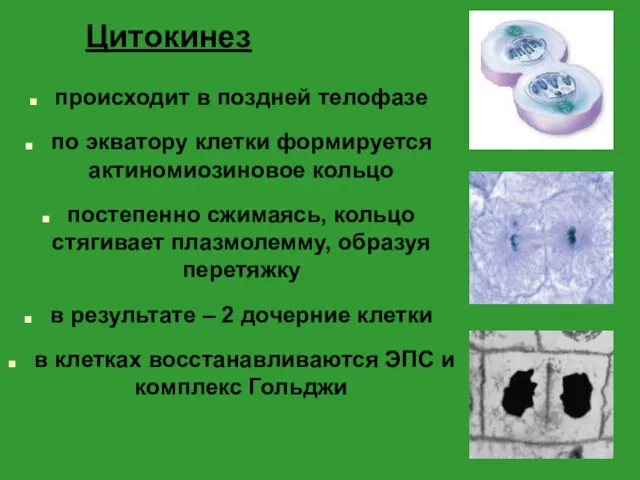
Цитокинез
происходит в поздней телофазе
по экватору клетки формируется актиномиозиновое кольцо
постепенно сжимаясь, кольцо
Цитокинез
происходит в поздней телофазе
по экватору клетки формируется актиномиозиновое кольцо
постепенно сжимаясь, кольцо

Результаты цитокинеза:
1) Органеллы равномерно распределены между
соседними клетками.
2) Сразу после деления дочерние
Результаты цитокинеза:
1) Органеллы равномерно распределены между
соседними клетками.
2) Сразу после деления дочерние
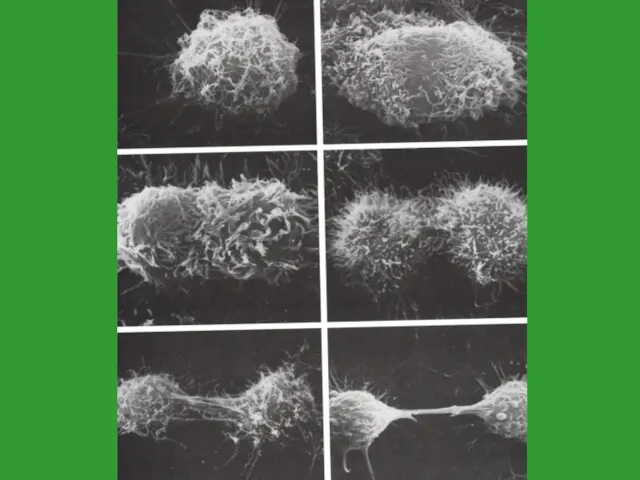

Продолжительность митоза
Продолжительность митоза зависит от:
размеров клеток
их плоидности
числа ядер
условий окружающей среды
Продолжительность митоза
Продолжительность митоза зависит от:
размеров клеток
их плоидности
числа ядер
условий окружающей среды

Пролиферация клеток и апоптоз
Количество клеток в ткани регулируется двумя процессами –
Пролиферация клеток и апоптоз
Количество клеток в ткани регулируется двумя процессами –

Апоптоз
Апоптоз – генетически запрограммированная гибель клеток, которая приводит к "аккуратной" разборке
Апоптоз
Апоптоз – генетически запрограммированная гибель клеток, которая приводит к "аккуратной" разборке

Апоптоз
от греч. – опадание листьев
Апоптоз
от греч. – опадание листьев

Причины апоптоза
две группы:
«неудовлетворительное» состояние клетки («апоптоз изнутри»)
«негативная» сигнализация снаружи, передающаяся через
Причины апоптоза
две группы:
«неудовлетворительное» состояние клетки («апоптоз изнутри»)
«негативная» сигнализация снаружи, передающаяся через

«Апоптоз изнутри»
повреждение хромосом:
многочисленные разрывы ДНК
нарушение конформации ДНК
сшивки между цепями ДНК
неправильная сегрегация
«Апоптоз изнутри»
повреждение хромосом:
многочисленные разрывы ДНК
нарушение конформации ДНК
сшивки между цепями ДНК
неправильная сегрегация

Причины повреждений
разные виды облучений
изменения температуры
химические соединения
нарушение питания клетки
деление клетки
повреждение клеточных структур,
Причины повреждений
разные виды облучений
изменения температуры
химические соединения
нарушение питания клетки
деление клетки
повреждение клеточных структур,

«Апоптоз по команде»
клетка вполне жизнеспособна, но, с позиций
целостного организма,
«Апоптоз по команде»
клетка вполне жизнеспособна, но, с позиций
целостного организма,

«Апоптоз по команде»
2) при формировании и функционировании иммунной системы
гибель аутореактивных
«Апоптоз по команде»
2) при формировании и функционировании иммунной системы
гибель аутореактивных

«Апоптоз по команде»
4) при функционировании женской половой системы
гибель клеток атрезирующих
«Апоптоз по команде»
4) при функционировании женской половой системы
гибель клеток атрезирующих

Регуляция апоптоза
одним из важнейших инструментов апоптоза является семейство цитоплазматических протеаз –
Регуляция апоптоза
одним из важнейших инструментов апоптоза является семейство цитоплазматических протеаз –

Регуляция апоптоза
мишени каспаз:
некоторые белки цитоскелета ( → изменение формы клетки –
Регуляция апоптоза
мишени каспаз:
некоторые белки цитоскелета ( → изменение формы клетки –

Митохондриальные факторы апоптоза
протеаза AIF и цитохром с – активаторы каспазного каскада
в
Митохондриальные факторы апоптоза
протеаза AIF и цитохром с – активаторы каспазного каскада
в

Белок р53
= транскрипционный фактор
активируется в ответ на разнообразные
повреждения клеточной
Белок р53
= транскрипционный фактор
активируется в ответ на разнообразные
повреждения клеточной

Белок р53
активирует гены:
ряда рецепторов, воспринимающих команду об апоптозе
отвечающие за
Белок р53
активирует гены:
ряда рецепторов, воспринимающих команду об апоптозе
отвечающие за

Итоги действия белка р53
клетка:
либо задерживается на той или иной стадии митотического
Итоги действия белка р53
клетка:
либо задерживается на той или иной стадии митотического

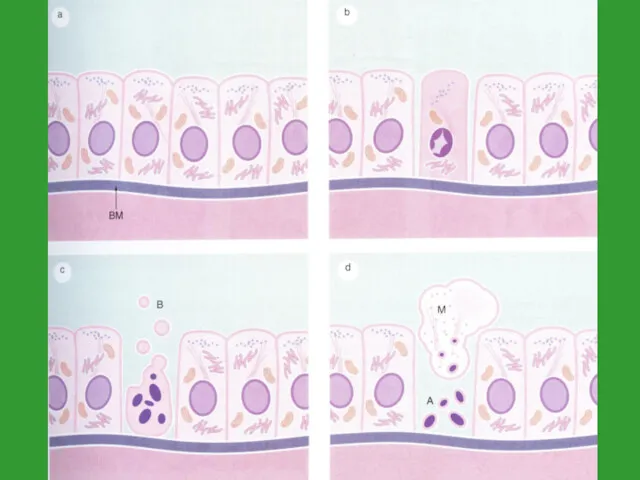
Морфология апоптоза
1) конденсация хроматина и некоторое сжатие клетки (из-за конденсации цитоплазмы)
хроматин
Морфология апоптоза
1) конденсация хроматина и некоторое сжатие клетки (из-за конденсации цитоплазмы)
хроматин
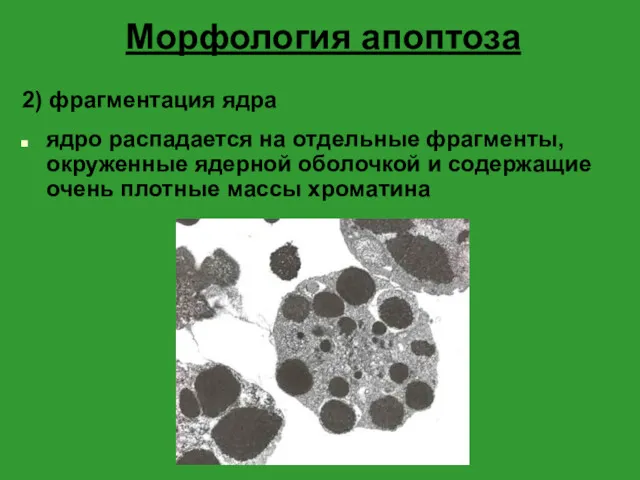
Морфология апоптоза
2) фрагментация ядра
ядро распадается на отдельные фрагменты, окруженные ядерной оболочкой
Морфология апоптоза
2) фрагментация ядра
ядро распадается на отдельные фрагменты, окруженные ядерной оболочкой
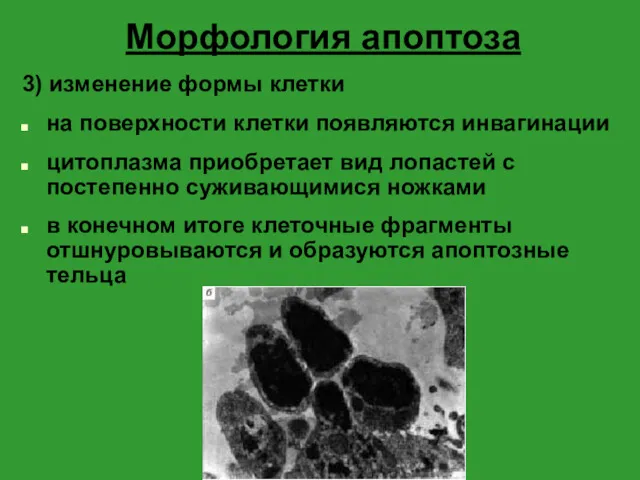
Морфология апоптоза
3) изменение формы клетки
на поверхности клетки появляются инвагинации
цитоплазма приобретает вид
Морфология апоптоза
3) изменение формы клетки
на поверхности клетки появляются инвагинации
цитоплазма приобретает вид
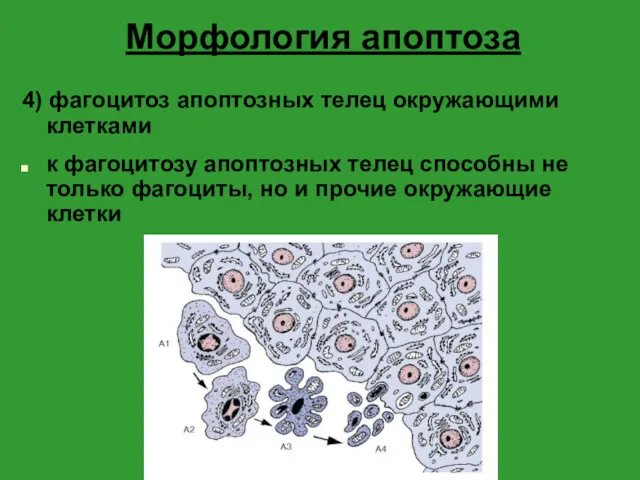
Морфология апоптоза
4) фагоцитоз апоптозных телец окружающими клетками
к фагоцитозу апоптозных телец способны
Морфология апоптоза
4) фагоцитоз апоптозных телец окружающими клетками
к фагоцитозу апоптозных телец способны
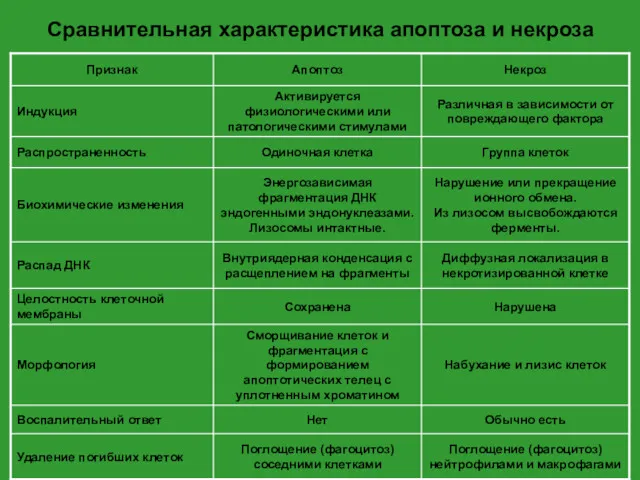
Сравнительная характеристика апоптоза и некроза
Сравнительная характеристика апоптоза и некроза

Старение клетки
с возрастом прогрессивно страдает ряд функций
клеток:
снижаются окислительное фосфорилирование в
Старение клетки
с возрастом прогрессивно страдает ряд функций
клеток:
снижаются окислительное фосфорилирование в

Теломерная теория старения
сформулирована А.М. Оловниковым в 1971 г
во всех соматических клетках
Теломерная теория старения
сформулирована А.М. Оловниковым в 1971 г
во всех соматических клетках

Факты, подтверждающие теорию
при каждом делении клетки длина теломерных участков сокращается
Факты, подтверждающие теорию
при каждом делении клетки длина теломерных участков сокращается

Дополнительные предположения
Как укорочение длины теломер влияет на клетку?
достаточно длинные теломеры вызывают
Дополнительные предположения
Как укорочение длины теломер влияет на клетку?
достаточно длинные теломеры вызывают

Морфологические изменения клетки при старении
неправильные и дольчатые ядра
полиморфные вакуолизированные митохондрий
уменьшение эндоплазматического
Морфологические изменения клетки при старении
неправильные и дольчатые ядра
полиморфные вакуолизированные митохондрий
уменьшение эндоплазматического
 Приспособленность организмов к среде обитания
Приспособленность организмов к среде обитания Кожа и ее строение
Кожа и ее строение Биохимия нервной, мышечной и соединительной ткани
Биохимия нервной, мышечной и соединительной ткани Грамотрицательные бактерии
Грамотрицательные бактерии Ветеринарно-санитарная экспертиза продуктов убоя животных и сырья при инвазионных болезнях
Ветеринарно-санитарная экспертиза продуктов убоя животных и сырья при инвазионных болезнях Презентация Экологическая игра Моя планета-человеческий дом
Презентация Экологическая игра Моя планета-человеческий дом Дыхание растений, животных, человека
Дыхание растений, животных, человека Систематика найпростіших. Тип апікомплексні
Систематика найпростіших. Тип апікомплексні Эволюция систем органов
Эволюция систем органов Рефлекс, рефлекторная дуга и обратная афферентация
Рефлекс, рефлекторная дуга и обратная афферентация Генетика – наука, объясняющая, почему ты похож на своего отца
Генетика – наука, объясняющая, почему ты похож на своего отца 20231022_tkani_rasteniy
20231022_tkani_rasteniy Растительноядные животные. Отличия в строении зубов
Растительноядные животные. Отличия в строении зубов Оплодотворение. Что такое оплодотворение
Оплодотворение. Что такое оплодотворение Процессы роста и развития организма человека
Процессы роста и развития организма человека Морфофункциональные особенности отдельных участков СОПР
Морфофункциональные особенности отдельных участков СОПР Строение и химический состав клетки
Строение и химический состав клетки Обмен простых белков. Образование биологически активных аминов
Обмен простых белков. Образование биологически активных аминов Углеводы. 9 класс
Углеводы. 9 класс Пресмыкающиеся или рептилии
Пресмыкающиеся или рептилии Половозрелая форма сосальщика
Половозрелая форма сосальщика Наука зоология
Наука зоология Питание клетки
Питание клетки Женские половые органы
Женские половые органы Половые железы. Функции
Половые железы. Функции Презентация к уроку биологии в 6 классе Половое размножение растений
Презентация к уроку биологии в 6 классе Половое размножение растений Исследования метагенома
Исследования метагенома Межвидовые взаимоотношения организмов. Виды биотических взаимоотношений (1)
Межвидовые взаимоотношения организмов. Виды биотических взаимоотношений (1)