- Биопотенциалдар. Тыныштық күй және әсер потенциалы

Содержание
- 2. Потенциалдар айырымы – бұл әр түрлі таңбадағы электр зарядтарының кеңістіктік бөлінуі болып табылады.
- 3. Тірі ағза жасушаларында, ұлпаларында пайда болатын потенциал айырмасын – биоэлектрлік потенциал д.а.
- 4. Тірі ағзада биоэлектрлік потенциалдардың пайда болуы жасуша мембранасында әр түрлі физика - химиялық градиенттердің болуына байланысты.
- 5. Медицинада электр өрісін зерттеуде ұлпалар мен мүшелердің биопотенциалдарын тіркеуге негізделінген диагностикалық әдістер: электрокардиография, электроэнцефалография, электромиография.
- 6. Ерітінділерде пайда болатын потенциалдар: электронды ионды
- 7. Электронды типтегі потенциалдар – еркін электрондардың, Ион типтегі потенциалдар – иондардың болуынан пайда болады.
- 9. Диффузия құбылысында пайда болатын, араласатын ерітінділерді аламыз. Концентрациясы көптен азға қарай жүреді
- 10. Фаза (күй, екі түрлі) аралық потенциалдар араласпайтын екі сұйықтың шекарасында пайда болады.
- 11. Ағзада тіркелетін биопотенциалдар – мембраналық потенциал.
- 12. Осыған байланысты мембрананың сыртқы және ішкі беттеріндегі потенциалдар айырымын - мембраналық потенциал д. а.
- 13. Мембраналық потенциал Тыныштық күй (қозғалыс жоқ кезде пайда болады) Әрекет болып екіге бөлінеді
- 14. Жасушаның беттік (сыртқы) мембранасының өткізгіштігі түрлі иондар үшін бірдей емес (таңдайды) Мембрананың екі жағындағы белгілі бір
- 15. Мембрананың сыртқы және ішкі жағы (моделдік жүйесі)
- 16. Иондар үшін тепе-теңдік күйге сәйкес келетін Нернст теңдеуі:
- 17. Мұндағы R – универсаль газ тұрақтысы, Т- абсолютті температура, F – Фарадей саны, [С2] және [С1]
- 18. Потенциалдың мембраналық теориясының негізін қалаушы 1902 жылы Бернштейн болды. Яғни калий иондарының өту диффузиясымен түсіндіріледі.
- 19. Тірі жасушаларда калий иондары жасуша аралық сұйықтарға қарағанда біраз есе көп. Нернст теңдеуімен есептелінген потенциалдар айырымы
- 20. ТКП-ның болуына және иондары себеп болады. Бұл иондар ағынының қосындылық тығыздығы:
- 21. Гольдман-Ходжкин-Катц теңдеуі
- 22. Иондар өтімділігі ағзаның күйіне байланысты болады. ТК-гі физиологиялық шарттарға байланысты түрлі иондардың өтімділік коэффициенттерінің қатынастары төмендегідей:
- 23. ТКП-на тек ғана K және Na иондары ғана үлестерін қосады. Мысалы, 30 град.С-ғы потенциал шамасы:
- 24. Гольдман-Ходжкин-Катц формуласымен есептелінген тыныштық күй потенциалы 60мВ болды. Гольдман теңдеуімен есептелінген мембраналық потенциал мәні Нернст теңдеуімен
- 25. Нернст және Гольдман теңдеулерінде иондардың мембрана арқылы активті тасмалдануы ескерілмеген. Мембраналық потенциалды есептеуде электрогендік иондық насостың
- 26. Мұндағы m – мембрана арқылы иондық насоспен тартылған натрий иондары мөлшерінің калий иондарының мөлшеріне қатынасын көрсетеді.
- 27. мембраналық потенциалды құруда калий концентрациясы градиентінің енгізілуін күшейтеді. Сондықтан Томас теңдеуімен есептелінген мембраналық потенциалдың мәні Гольдман
- 28. Жасушадағы биоэнергетикалық процесстердің және насосының жұмысының бұзылуы потенциалдың азаюуына ықпал етеді. Мұндай жағдайда мембраналық потенциал Гольдман
- 31. Мембраналық биопотенциалдарды зерттеу: 1.Микроэлектрод әдісімен жасушаішілік потенциалды өлшеу. 2. Биопотенциалды күшейткіш 3. Зерттеу объектісі ретінде ірі
- 32. Биопотенциалды өлшеу микроэлектрод әдісі
- 33. Әрекет потенциал Қозу жағдайында жасуша мен қоршаған орта арасындағы потенциал айырымы өзгереді. Осы кезде ӘП пайда
- 35. ӘП конденсатордың зарядталуы мен разрядталуы кезіндегі апериодтық (периодтық емес) үрдістер тәріздес болады.
- 36. 1 сурет . Әрекет потенциалды зерттеу
- 37. Нерв талшықтарында және қаңқа бұлшық еттерінде әрекет потенциалдың ұзақтығы 1 мс шамасында болады. (жүрек бұлшық еттерінде
- 38. Әрекет потенциалдың негізгі қасиеттері: Деполяризация потенциалының табалдырық мәнінің болуы Егер деполяризация потенциалы табалдырық мәнінен үлкен болса,
- 39. 1 сурет . Әрекет потенциалды зерттеу
- 40. Рефрактерлік период, әрекет потенциалдың пайда болу уақытындағы мембрананың қозбаған кезеңі және қозудан кейінгі қалдық құбылыстар. Қозу
- 41. Қозу кезінде натрий иондары үшін мембрананың өтімділігі күрт артады. Қозу кезеңінде Тыныштық потенциал кезінде әр түрлі
- 42. ӘП бірнеше фазадан тұрады: потенциал оң бағытқа қарай тез артады. Арту барысында жасушалық мембрана өзінің қалыпты
- 43. Деполяризация қисығы нолдік сызықтан өтіп, мембраналық потенциал оң болады. Осы оң фазаны ӘП –ң инверсиясы деп
- 44. Нерв талшықтарында әрекет потенциалдың реполяризация кезеңінде «іздік» потенциалдары байқалады. Реполяризация кезеңінің соңында потенциалдың күшеюін гиперполяризация деп
- 45. Қозу үрдісі ӘП және оның фазалары: 1 — деполяризация, 2 — инверсия, 3 —реполяризация, 4 —
- 46. Жергілікті жауап 2. Деполяризация. 3. Реполяризация. 4. Теріс потенциал. 5. Оң потенциал. Әрекет потенциалының фазалары
- 48. Мембрана қозуы Ходжкин -Хаксли теңдеуімен сипатталады. - мембрана арқылы өтетін ток, См - мембрананың сыйымдылығы, -
- 49. ӘП-дың маңызды қасиетінің бірі олардың нерв талшықтары бойымен таралуы.
- 50. Нерв жасушасы – нейрон – жасуша денесінен (сома), бірнеше өсінділер – дендриттерден және шығару өсіндісі –
- 51. Нейронның құрылымы Нейрондар — нерв жүйесінің негізігі элементі. Жасушаның ұзын өсінділері нерв импульстарын таратады.
- 52. Нейронның құрылымы
- 53. Нейрон денесінің диаметрі 4-5 –ден 135 мк-ге дейін болады. Нерв жасушасы денесінің формасы да әр түрлі
- 54. Нерв жасушасы денесінен нерв импульсы ұзын өсіндісі аксон бойынша таралады. Аксонның негізгі функциясы –нейронды пайда болған
- 56. Синапс Аксон
- 57. Тірі ағзаларда эл.нерв импульстары нәтижесінде (ӘП)- ақпарат рецепторлардан ми нейрондарына және одан бұлшық еттерге беріледі Нерв
- 58. Аксондар миелинді қабықшамен қапталады. Миелинді қабықша омдық кедергісі жоғары 80% липидтерден және 20% ақуыздан тұрады. Миелинді
- 59. Нерв талшықтарымен импульстың таралуы нерв талшықтарының құрылымымен анықталады. Құрылымы өткізгіш ретінде аксоплазма, электрлік изолятор ретінде миелинді
- 61. Олай болса, нерв талшықтары миелинді (майлы қабықты) (1) және миелинсіз (майлы қабықсыз) (2) болып бөлінеді Олардың
- 62. Миелинді нерв талшықтарында қозу үрдісі тек Ранвье үзілісінде ғана пайда болады. Қозу бір Ранвье үзілісінен екіншісіне
- 63. Миелинді нерв талшықтарымен нерв импульсының таралу механизмі
- 64. Миелинсіз нерв талшықтарында тығыз майлы қабықтар болмайды, оларда қозу үздіксіз және өте жай өтеді.
- 65. Миелинсіз нерв талшықтарымен импульстың таралу механизмі
- 66. Нерв талшығы бойымен қозудың таралуы механизмі I. Миелинсіз нерв талшығы; - қозу үздіксіз түрде таралады. II.
- 67. Әдебиеттер: 1. Арызханов Б.,Биологиялық физика,1990 ж. 2. Кошенов Б.К. Медициналық биофизика, ,2011г. 3. Тиманюк В.А., Животова
- 69. Скачать презентацию

Потенциалдар айырымы – бұл әр түрлі таңбадағы электр зарядтарының кеңістіктік бөлінуі

Тірі ағза жасушаларында, ұлпаларында пайда болатын потенциал айырмасын – биоэлектрлік потенциал
Тірі ағза жасушаларында, ұлпаларында пайда болатын потенциал айырмасын – биоэлектрлік потенциал

Тірі ағзада биоэлектрлік потенциалдардың пайда болуы жасуша мембранасында әр түрлі физика
Тірі ағзада биоэлектрлік потенциалдардың пайда болуы жасуша мембранасында әр түрлі физика

Медицинада электр өрісін зерттеуде ұлпалар мен мүшелердің биопотенциалдарын тіркеуге негізделінген диагностикалық
Медицинада электр өрісін зерттеуде ұлпалар мен мүшелердің биопотенциалдарын тіркеуге негізделінген диагностикалық

Ерітінділерде пайда болатын потенциалдар:
электронды
ионды
Ерітінділерде пайда болатын потенциалдар:
электронды
ионды

Электронды типтегі потенциалдар – еркін электрондардың,
Ион типтегі потенциалдар – иондардың
Электронды типтегі потенциалдар – еркін электрондардың,
Ион типтегі потенциалдар – иондардың


Диффузия құбылысында пайда болатын, араласатын ерітінділерді аламыз.
Концентрациясы көптен азға қарай
Диффузия құбылысында пайда болатын, араласатын ерітінділерді аламыз. Концентрациясы көптен азға қарай

Фаза (күй, екі түрлі) аралық потенциалдар
араласпайтын екі сұйықтың шекарасында
Фаза (күй, екі түрлі) аралық потенциалдар араласпайтын екі сұйықтың шекарасында

Ағзада тіркелетін биопотенциалдар – мембраналық потенциал.
Ағзада тіркелетін биопотенциалдар – мембраналық потенциал.

Осыған байланысты мембрананың сыртқы және ішкі беттеріндегі потенциалдар айырымын -
Осыған байланысты мембрананың сыртқы және ішкі беттеріндегі потенциалдар айырымын -

Мембраналық потенциал
Тыныштық күй (қозғалыс жоқ кезде пайда болады)
Әрекет болып екіге бөлінеді
Мембраналық потенциал
Тыныштық күй (қозғалыс жоқ кезде пайда болады)
Әрекет болып екіге бөлінеді

Жасушаның беттік (сыртқы) мембранасының өткізгіштігі түрлі иондар үшін бірдей емес (таңдайды)
Мембрананың
Жасушаның беттік (сыртқы) мембранасының өткізгіштігі түрлі иондар үшін бірдей емес (таңдайды)
Мембрананың

Мембрананың сыртқы және ішкі жағы (моделдік жүйесі)
Мембрананың сыртқы және ішкі жағы (моделдік жүйесі)

Иондар үшін тепе-теңдік күйге сәйкес келетін
Нернст теңдеуі:
Иондар үшін тепе-теңдік күйге сәйкес келетін
Нернст теңдеуі:

Мұндағы R – универсаль газ тұрақтысы,
Т- абсолютті температура,
F –
Мұндағы R – универсаль газ тұрақтысы,
Т- абсолютті температура,
F –

Потенциалдың мембраналық теориясының негізін қалаушы 1902 жылы Бернштейн болды. Яғни калий

Тірі жасушаларда калий иондары жасуша аралық сұйықтарға қарағанда біраз есе көп.
Тірі жасушаларда калий иондары жасуша аралық сұйықтарға қарағанда біраз есе көп.

ТКП-ның болуына
және
иондары себеп болады. Бұл иондар ағынының қосындылық тығыздығы:
ТКП-ның болуына
және
иондары себеп болады. Бұл иондар ағынының қосындылық тығыздығы:

Гольдман-Ходжкин-Катц теңдеуі
Гольдман-Ходжкин-Катц теңдеуі

Иондар өтімділігі ағзаның күйіне байланысты болады. ТК-гі физиологиялық шарттарға байланысты түрлі
Иондар өтімділігі ағзаның күйіне байланысты болады. ТК-гі физиологиялық шарттарға байланысты түрлі

ТКП-на тек ғана K және Na иондары ғана үлестерін қосады. Мысалы,
ТКП-на тек ғана K және Na иондары ғана үлестерін қосады. Мысалы,

Гольдман-Ходжкин-Катц формуласымен есептелінген тыныштық күй потенциалы 60мВ болды.
Гольдман теңдеуімен
Гольдман-Ходжкин-Катц формуласымен есептелінген тыныштық күй потенциалы 60мВ болды.
Гольдман теңдеуімен

Нернст және Гольдман теңдеулерінде иондардың мембрана арқылы активті тасмалдануы ескерілмеген. Мембраналық
Нернст және Гольдман теңдеулерінде иондардың мембрана арқылы активті тасмалдануы ескерілмеген. Мембраналық

Мұндағы m – мембрана арқылы иондық насоспен тартылған натрий иондары мөлшерінің
Мұндағы m – мембрана арқылы иондық насоспен тартылған натрий иондары мөлшерінің

мембраналық потенциалды құруда калий
концентрациясы градиентінің енгізілуін
күшейтеді.
Сондықтан Томас теңдеуімен есептелінген
мембраналық потенциалды құруда калий
концентрациясы градиентінің енгізілуін
күшейтеді.
Сондықтан Томас теңдеуімен есептелінген

Жасушадағы биоэнергетикалық процесстердің және насосының жұмысының бұзылуы потенциалдың азаюуына ықпал етеді. Мұндай
Жасушадағы биоэнергетикалық процесстердің және насосының жұмысының бұзылуы потенциалдың азаюуына ықпал етеді. Мұндай



Мембраналық биопотенциалдарды зерттеу:
1.Микроэлектрод әдісімен жасушаішілік потенциалды өлшеу.
2. Биопотенциалды күшейткіш
3.
Мембраналық биопотенциалдарды зерттеу:
1.Микроэлектрод әдісімен жасушаішілік потенциалды өлшеу.
2. Биопотенциалды күшейткіш
3.
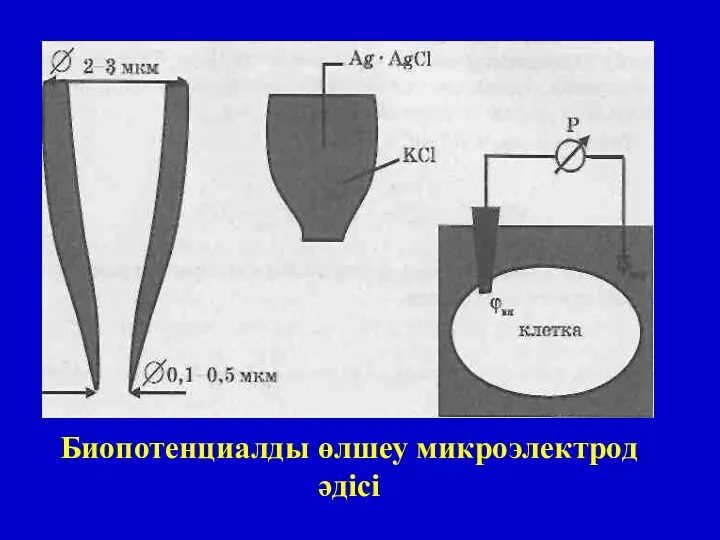
Биопотенциалды өлшеу микроэлектрод әдісі
Биопотенциалды өлшеу микроэлектрод әдісі

Әрекет потенциал
Қозу жағдайында жасуша мен қоршаған орта арасындағы потенциал айырымы өзгереді.
Әрекет потенциал
Қозу жағдайында жасуша мен қоршаған орта арасындағы потенциал айырымы өзгереді.


ӘП конденсатордың зарядталуы мен разрядталуы кезіндегі апериодтық (периодтық емес)
ӘП конденсатордың зарядталуы мен разрядталуы кезіндегі апериодтық (периодтық емес)
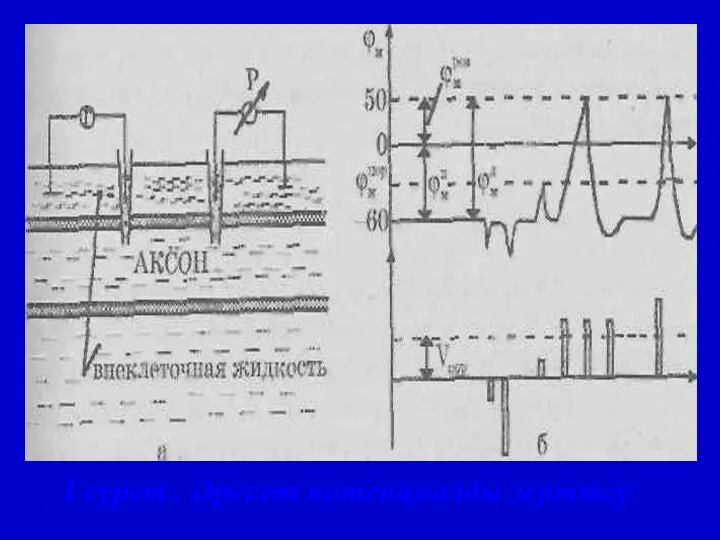
1 сурет . Әрекет потенциалды зерттеу
1 сурет . Әрекет потенциалды зерттеу

Нерв талшықтарында және қаңқа бұлшық еттерінде әрекет потенциалдың ұзақтығы 1
Нерв талшықтарында және қаңқа бұлшық еттерінде әрекет потенциалдың ұзақтығы 1

Әрекет потенциалдың негізгі қасиеттері:
Деполяризация потенциалының табалдырық мәнінің болуы
Егер деполяризация потенциалы табалдырық
Әрекет потенциалдың негізгі қасиеттері:
Деполяризация потенциалының табалдырық мәнінің болуы
Егер деполяризация потенциалы табалдырық

1 сурет . Әрекет потенциалды зерттеу
1 сурет . Әрекет потенциалды зерттеу

Рефрактерлік период, әрекет потенциалдың пайда болу уақытындағы мембрананың қозбаған кезеңі және
Рефрактерлік период, әрекет потенциалдың пайда болу уақытындағы мембрананың қозбаған кезеңі және

Қозу кезінде натрий иондары үшін мембрананың өтімділігі күрт артады.
Қозу
Қозу кезінде натрий иондары үшін мембрананың өтімділігі күрт артады.
Қозу

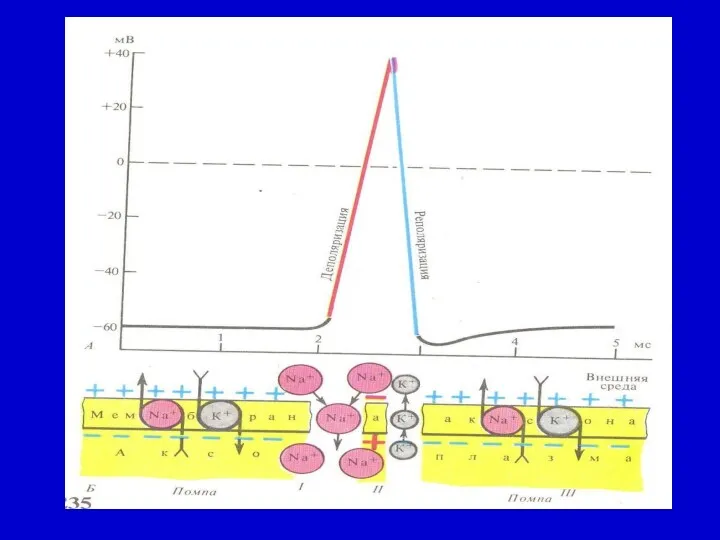
ӘП бірнеше фазадан тұрады: потенциал оң бағытқа қарай тез артады. Арту
ӘП бірнеше фазадан тұрады: потенциал оң бағытқа қарай тез артады. Арту

Деполяризация қисығы нолдік сызықтан өтіп, мембраналық потенциал оң болады.
Осы оң
Деполяризация қисығы нолдік сызықтан өтіп, мембраналық потенциал оң болады.
Осы оң

Нерв талшықтарында әрекет потенциалдың реполяризация кезеңінде «іздік» потенциалдары байқалады.
Реполяризация кезеңінің соңында
Нерв талшықтарында әрекет потенциалдың реполяризация кезеңінде «іздік» потенциалдары байқалады.
Реполяризация кезеңінің соңында

Қозу үрдісі
ӘП және оның фазалары:
1 — деполяризация,
2 —
Қозу үрдісі
ӘП және оның фазалары:
1 — деполяризация,
2 —

Жергілікті жауап
2. Деполяризация. 3. Реполяризация.
4. Теріс потенциал. 5. Оң
Жергілікті жауап
2. Деполяризация. 3. Реполяризация.
4. Теріс потенциал. 5. Оң

Мембрана қозуы Ходжкин -Хаксли теңдеуімен сипатталады.
- мембрана арқылы өтетін
Мембрана қозуы Ходжкин -Хаксли теңдеуімен сипатталады.
- мембрана арқылы өтетін

ӘП-дың маңызды қасиетінің бірі олардың нерв талшықтары бойымен таралуы.

Нерв жасушасы – нейрон – жасуша денесінен (сома), бірнеше өсінділер –
Нерв жасушасы – нейрон – жасуша денесінен (сома), бірнеше өсінділер –

Нейронның құрылымы
Нейрондар — нерв жүйесінің негізігі элементі.
Жасушаның ұзын өсінділері нерв
Нейрондар — нерв жүйесінің негізігі элементі.
Жасушаның ұзын өсінділері нерв

Нейронның құрылымы
Нейронның құрылымы

Нейрон денесінің диаметрі 4-5 –ден 135 мк-ге дейін болады. Нерв жасушасы

Нерв жасушасы денесінен нерв импульсы
ұзын өсіндісі аксон бойынша таралады.
Нерв жасушасы денесінен нерв импульсы
ұзын өсіндісі аксон бойынша таралады.
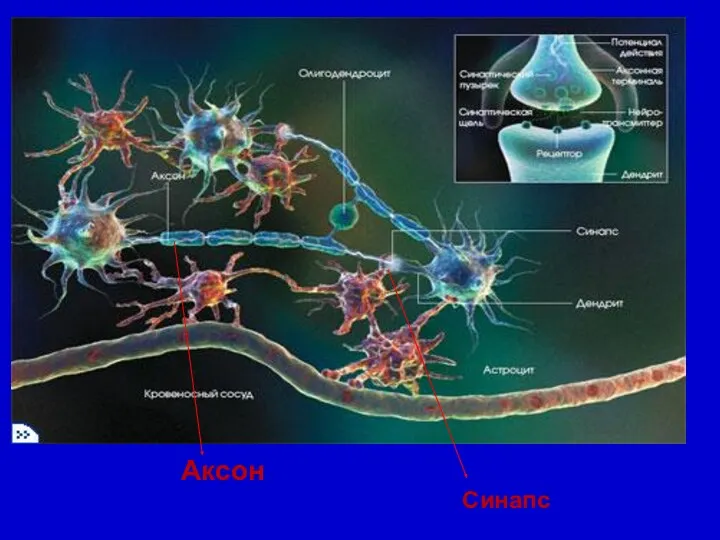
Синапс
Аксон
Синапс
Аксон

Тірі ағзаларда эл.нерв импульстары нәтижесінде (ӘП)- ақпарат рецепторлардан ми нейрондарына және
Тірі ағзаларда эл.нерв импульстары нәтижесінде (ӘП)- ақпарат рецепторлардан ми нейрондарына және

Аксондар миелинді қабықшамен қапталады. Миелинді қабықша омдық кедергісі жоғары 80%
Аксондар миелинді қабықшамен қапталады. Миелинді қабықша омдық кедергісі жоғары 80%

Нерв талшықтарымен импульстың таралуы нерв талшықтарының құрылымымен анықталады.
Құрылымы өткізгіш ретінде аксоплазма,
Нерв талшықтарымен импульстың таралуы нерв талшықтарының құрылымымен анықталады.
Құрылымы өткізгіш ретінде аксоплазма,


Олай болса, нерв талшықтары миелинді (майлы қабықты) (1) және миелинсіз (майлы
Олай болса, нерв талшықтары миелинді (майлы қабықты) (1) және миелинсіз (майлы

Миелинді нерв талшықтарында қозу үрдісі тек Ранвье үзілісінде ғана пайда болады.
Миелинді нерв талшықтарында қозу үрдісі тек Ранвье үзілісінде ғана пайда болады.
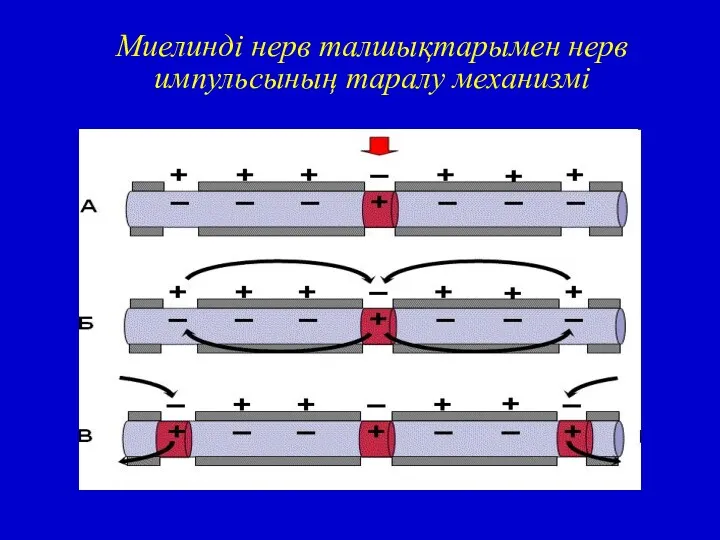
Миелинді нерв талшықтарымен нерв импульсының таралу механизмі
Миелинді нерв талшықтарымен нерв импульсының таралу механизмі

Миелинсіз нерв талшықтарында тығыз майлы қабықтар болмайды, оларда қозу үздіксіз және
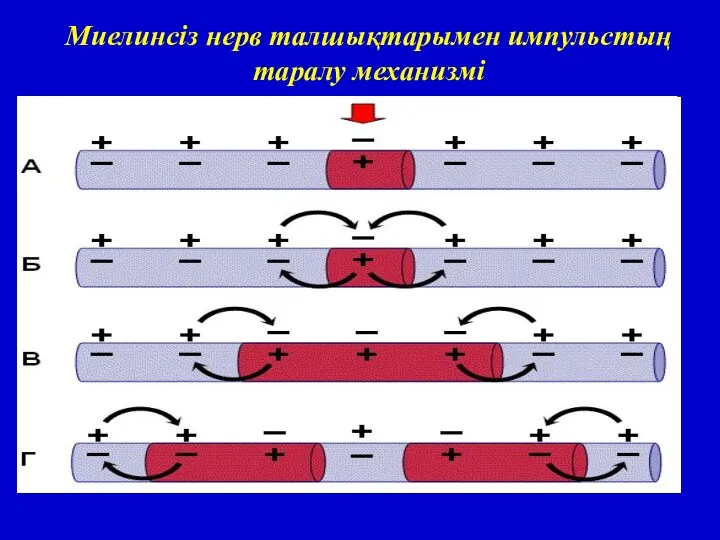
Миелинсіз нерв талшықтарымен импульстың таралу механизмі
Миелинсіз нерв талшықтарымен импульстың таралу механизмі

Нерв талшығы бойымен қозудың таралуы
механизмі
I. Миелинсіз нерв талшығы;
- қозу
Нерв талшығы бойымен қозудың таралуы
механизмі
I. Миелинсіз нерв талшығы;
- қозу

Әдебиеттер:
1. Арызханов Б.,Биологиялық физика,1990 ж.
2. Кошенов Б.К. Медициналық биофизика, ,2011г.
3.
Әдебиеттер:
1. Арызханов Б.,Биологиялық физика,1990 ж.
2. Кошенов Б.К. Медициналық биофизика, ,2011г.
3.
 Парогенераторы АЭС. Первичные теплоносители. (Тема 3)
Парогенераторы АЭС. Первичные теплоносители. (Тема 3) Сила трения
Сила трения Синтез наноматериалов золь-гель методом. (Лекция 5)
Синтез наноматериалов золь-гель методом. (Лекция 5) Nanophotonics class 4. Density of states
Nanophotonics class 4. Density of states Изопроцессы
Изопроцессы Сложение скоростей
Сложение скоростей ТРИЗ Электричество
ТРИЗ Электричество Все о ракетах
Все о ракетах Физика в профессии Слесарь по ремонту автомобилей
Физика в профессии Слесарь по ремонту автомобилей Электрооборудование автомобилей. Автоматическая коробка переключения передач с электронным управлением
Электрооборудование автомобилей. Автоматическая коробка переключения передач с электронным управлением Методы оптической молекулярной спектроскопии
Методы оптической молекулярной спектроскопии Types of Forces
Types of Forces Магнітне поле постійного струму
Магнітне поле постійного струму Внутренняя энергия. Первый закон термодинамики
Внутренняя энергия. Первый закон термодинамики Заттың агрегаттық күйлерінің өзгеруі. 8 сынып
Заттың агрегаттық күйлерінің өзгеруі. 8 сынып Тонкостенные оболочки. Основные допущения. Уравнение Лапласа. Лекция 8
Тонкостенные оболочки. Основные допущения. Уравнение Лапласа. Лекция 8 Тяговый расчет автомобиля
Тяговый расчет автомобиля Сообщающие сосуды
Сообщающие сосуды Лампа накаливания
Лампа накаливания Interest in the synthesis of metal nanoparticles by explosion
Interest in the synthesis of metal nanoparticles by explosion Электромагнитные колебания
Электромагнитные колебания Взаимодействие электромагнитных световых волн с веществом. Поляризация света. Виды поляризации
Взаимодействие электромагнитных световых волн с веществом. Поляризация света. Виды поляризации Полевые транзисторы FET (field-effect transistor). Устройство, принципы работы полевых транзисторов различных типов
Полевые транзисторы FET (field-effect transistor). Устройство, принципы работы полевых транзисторов различных типов Физика. Физические основы механики
Физика. Физические основы механики Критерії роботоздатності деталей машин. (Лекція 2)
Критерії роботоздатності деталей машин. (Лекція 2) Дыбыс, кең мағынасында
Дыбыс, кең мағынасында Magnit orici
Magnit orici Законы постоянного тока
Законы постоянного тока