- Антиген. Врожденный и адаптивный иммунитет

Содержание
- 2. Врожденный и адаптивный иммунитет Врожденный Имеется в организме с рождения Быстрая реакция на появление патогенов Низкая
- 3. Антиген В-клеточный рецептор — антитело Антиген — молекула, способная вызвать иммунный ответ. Антиген является, как правило,
- 4. Связывание антигена с Т-клеточным рецептором Т-клеточный рецептор (TCR) связывается с антигеном не напрямую, а в комплексе
- 5. Антиген-специфичные рецепторы В- и Т-лимфоцитов Рецепторы В-лимфоцитов — иммуноглобулины, состоят из двух легких и двух тяжелых
- 6. Вариабельные участки антител
- 7. Вариативность антител V(D)J-рекомбинация
- 8. Вариативность антител V(D)J-рекомбинация Комбинация разных цепей при образовании антитела (κ/λ) Комбинация генных сегментов V(D)J-рекомбинации Соматическая гипермутация
- 9. Вариативность Т-клеточного рецептора V(D)J-рекомбинация Вариативность полученных в ходе рекомбинации генов, кодирующих Т-клеточный рецептор, сравнима с вариативностью
- 10. Главный комплекс гистосовместимости MHC (Major histocompatibility complex), он же HLA (human leukocyte antigen) у человека, —
- 11. MHC первого и второго класса MHC I класса связываются с TCR (Т-клеточным рецептором) «Т-киллеров», MHC II
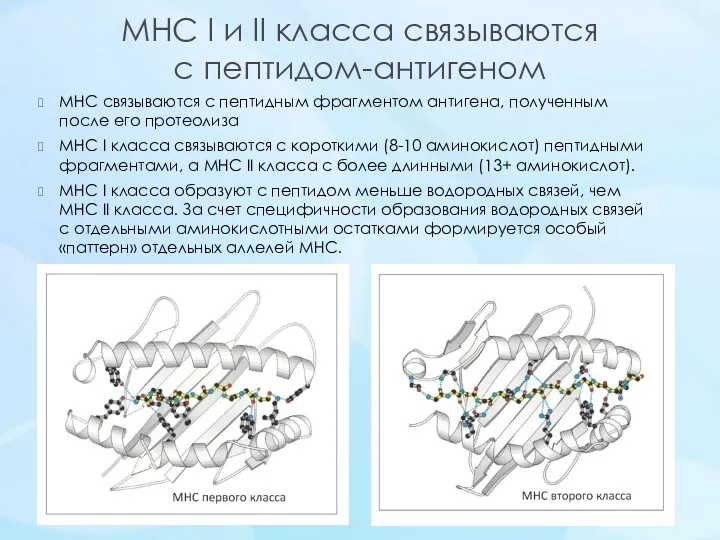
- 12. MHC I и II класса связываются с пептидом-антигеном МНС связываются с пептидным фрагментом антигена, полученным после
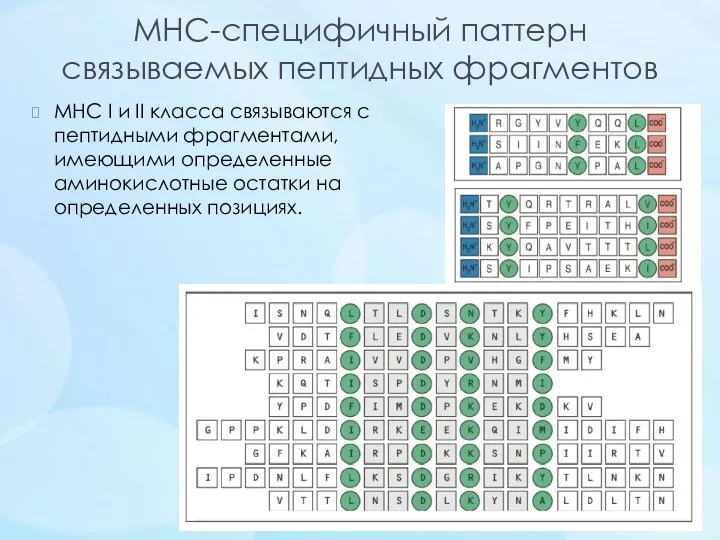
- 13. MHC-специфичный паттерн связываемых пептидных фрагментов МНС I и II класса связываются с пептидными фрагментами, имеющими определенные
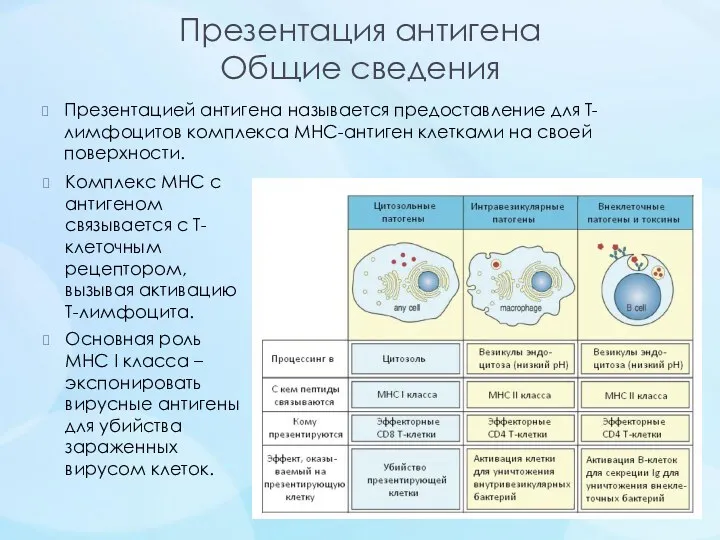
- 14. Презентация антигена Общие сведения Презентацией антигена называется предоставление для Т-лимфоцитов комплекса MHC-антиген клетками на своей поверхности.
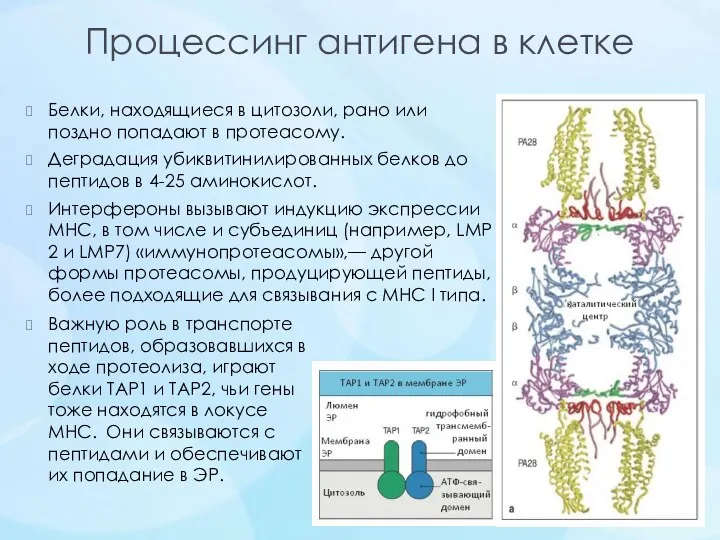
- 15. Процессинг антигена в клетке Белки, находящиеся в цитозоли, рано или поздно попадают в протеасому. Деградация убиквитинилированных
- 16. Транспорт MHC I класса в эндоплазматический ретикулум Для сборки MHC I класса необходимо связаться с антигеном
- 17. Нарушения презентацию антигена Вирусы Многие вирусы с помощью различных белков-эвазинов пытаются нарушить презентацию антигена MHC I
- 18. Нарушения презентации антигена Опухолевые клетки Опухолевые клетки часто характеризуются наличие так называемых «неоантигенов» — антигенов, не
- 19. Презентация антигена MHC II класса Антиген попадает в клетку не из цитозоли. Антиген захватывается путем эндоцитоза
- 20. Инвариантная цепь и MHC II класса В эндоплазматическом ретикулуме МНС II связывается с инвариантной цепью (CD74).
- 21. Нарушения презентации антигена Паразиты Toxoplasma gondii стимулирует экспрессию CD74 (инвариантной цепи) и подавляет экспрессию МНС I
- 22. Кросс-презентация Презентация антигена MHC I класса антигенами, которые попали в клетку извне, а не из цитозоля.
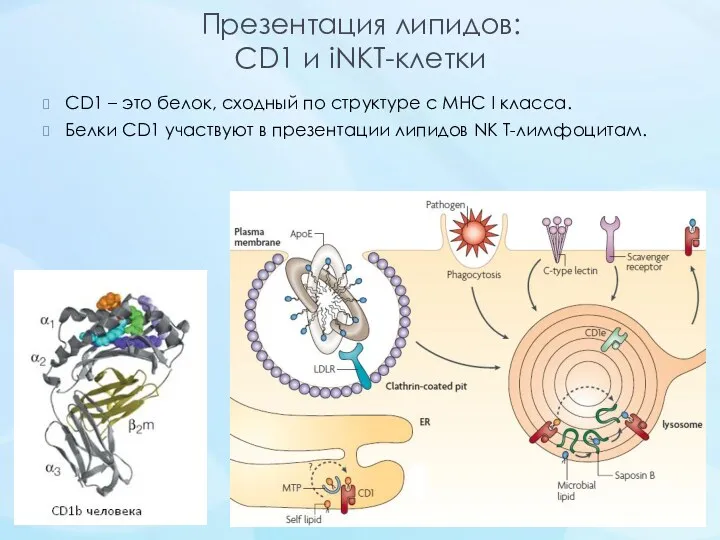
- 23. Презентация липидов: CD1 и iNKT-клетки CD1 – это белок, сходный по структуре с MHC I класса.
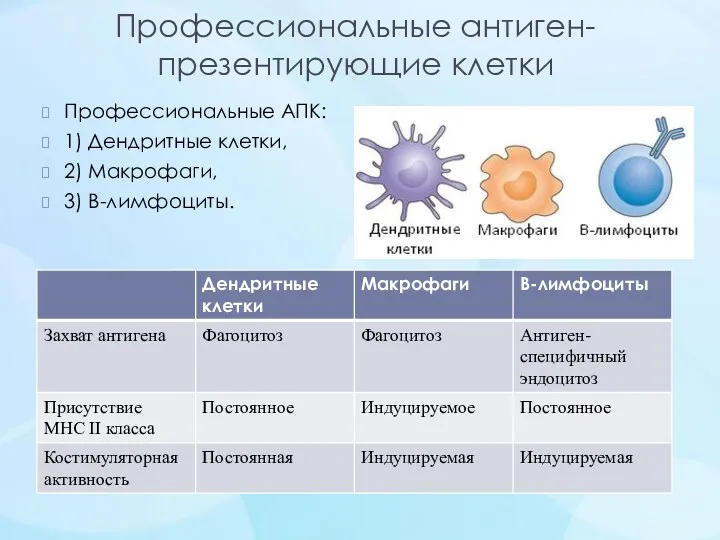
- 24. Профессиональные антиген-презентирующие клетки Профессиональные АПК: 1) Дендритные клетки, 2) Макрофаги, 3) В-лимфоциты.
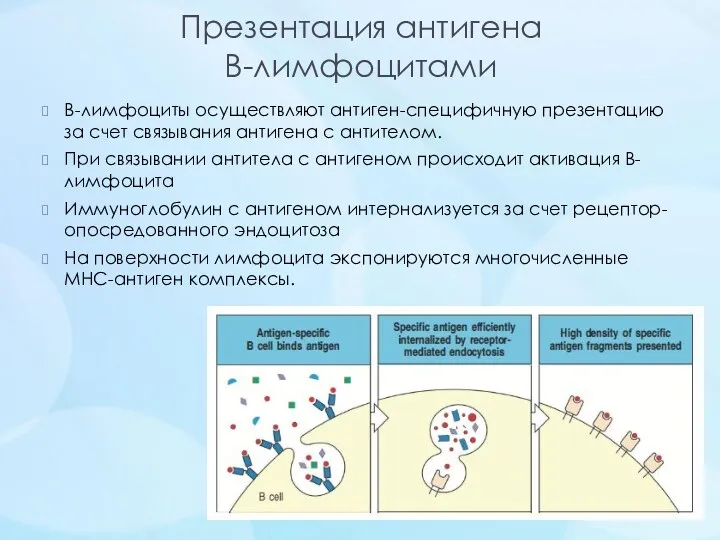
- 25. Презентация антигена В-лимфоцитами В-лимфоциты осуществляют антиген-специфичную презентацию за счет связывания антигена с антителом. При связывании антитела
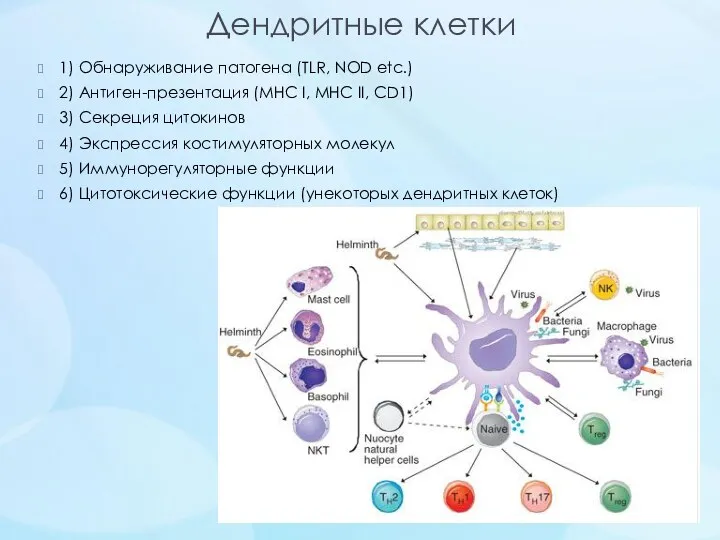
- 26. Дендритные клетки 1) Обнаруживание патогена (TLR, NOD etc.) 2) Антиген-презентация (MHC I, MHC II, CD1) 3)
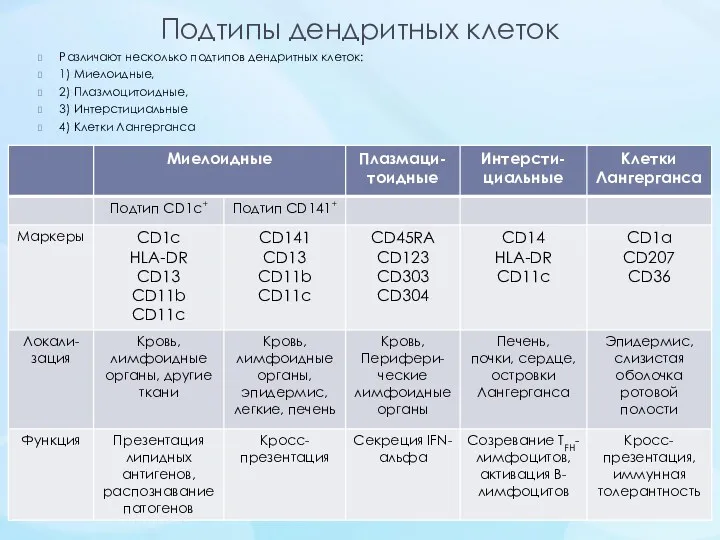
- 27. Подтипы дендритных клеток Различают несколько подтипов дендритных клеток: 1) Миелоидные, 2) Плазмоцитоидные, 3) Интерстициальные 4) Клетки
- 28. Спасибо за внимание!
- 30. Скачать презентацию
Врожденный и адаптивный иммунитет
Врожденный
Имеется в организме с рождения
Быстрая реакция на появление патогенов
Низкая специфичность
Низкая
Врожденный и адаптивный иммунитет
Врожденный
Имеется в организме с рождения
Быстрая реакция на появление патогенов
Низкая специфичность
Низкая

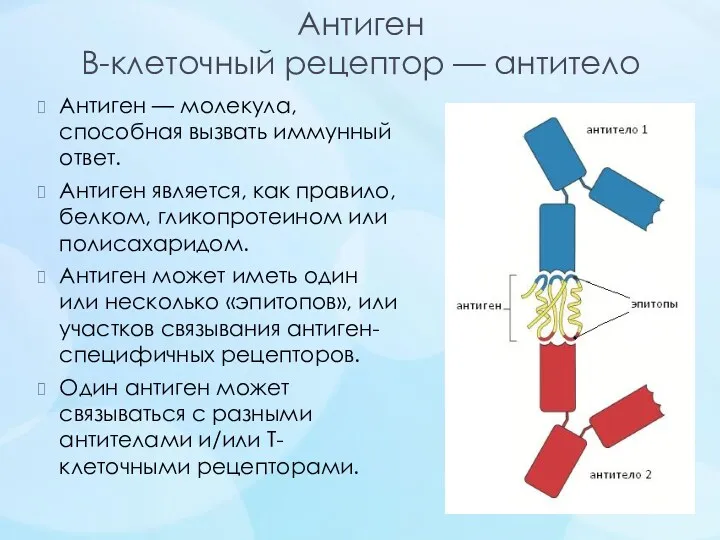
Антиген
В-клеточный рецептор — антитело
Антиген — молекула, способная вызвать иммунный ответ.
Антиген является, как правило,
Антиген
В-клеточный рецептор — антитело
Антиген — молекула, способная вызвать иммунный ответ.
Антиген является, как правило,
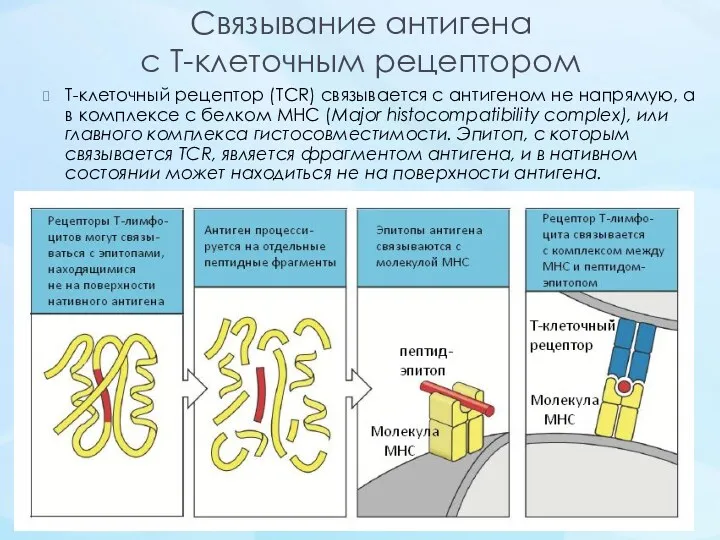
Связывание антигена
с Т-клеточным рецептором
Т-клеточный рецептор (TCR) связывается с антигеном не напрямую, а
Связывание антигена
с Т-клеточным рецептором
Т-клеточный рецептор (TCR) связывается с антигеном не напрямую, а
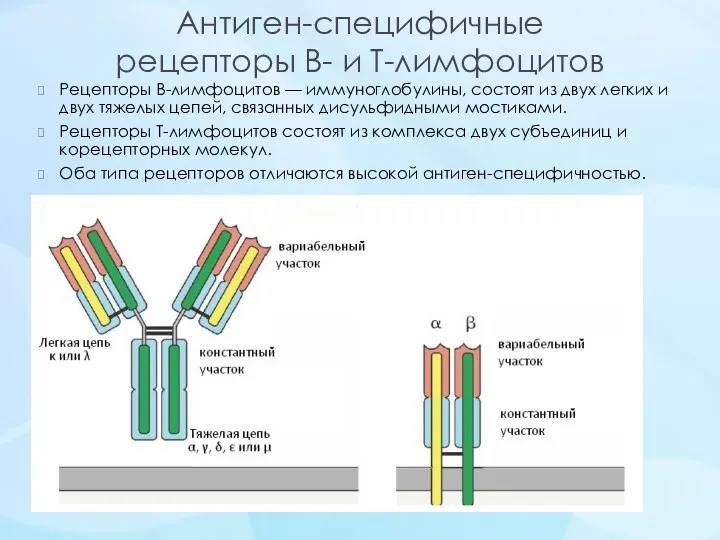
Антиген-специфичные
рецепторы В- и Т-лимфоцитов
Рецепторы В-лимфоцитов — иммуноглобулины, состоят из двух легких и
Антиген-специфичные
рецепторы В- и Т-лимфоцитов
Рецепторы В-лимфоцитов — иммуноглобулины, состоят из двух легких и
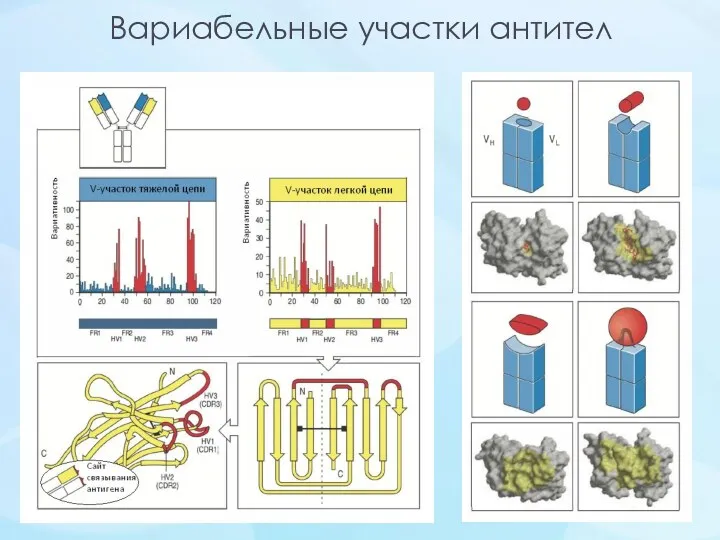
Вариабельные участки антител
Вариабельные участки антител
Вариативность антител
V(D)J-рекомбинация
Вариативность антител
V(D)J-рекомбинация
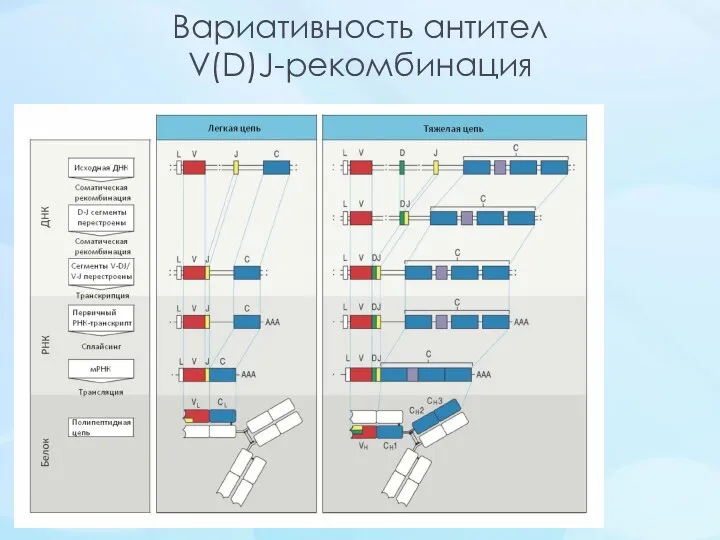
Вариативность антител
V(D)J-рекомбинация
Комбинация разных цепей при образовании антитела (κ/λ)
Комбинация генных сегментов V(D)J-рекомбинации
Соматическая гипермутация
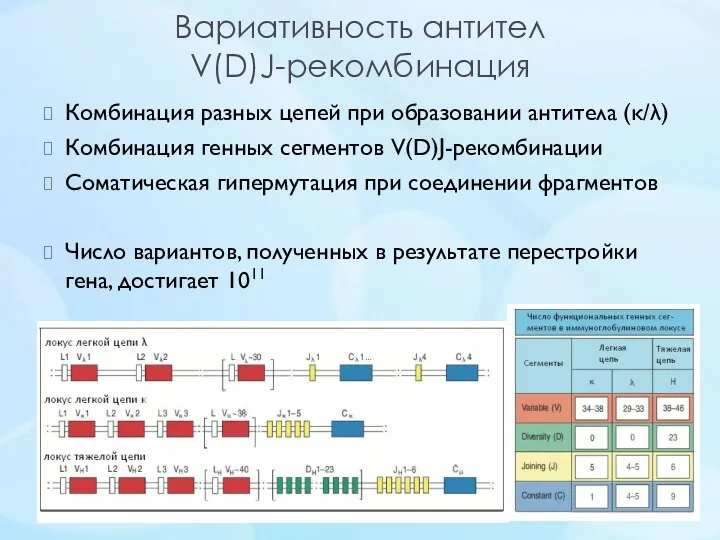
Вариативность антител
V(D)J-рекомбинация
Комбинация разных цепей при образовании антитела (κ/λ)
Комбинация генных сегментов V(D)J-рекомбинации
Соматическая гипермутация
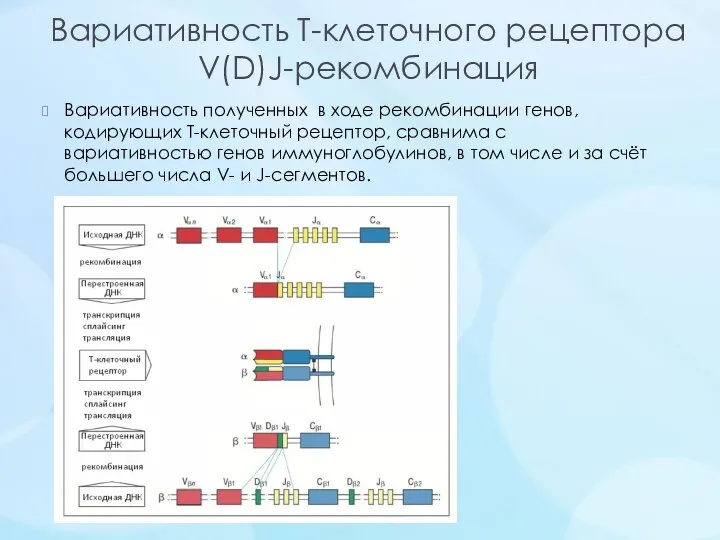
Вариативность Т-клеточного рецептора
V(D)J-рекомбинация
Вариативность полученных в ходе рекомбинации генов, кодирующих Т-клеточный рецептор, сравнима с
Вариативность Т-клеточного рецептора
V(D)J-рекомбинация
Вариативность полученных в ходе рекомбинации генов, кодирующих Т-клеточный рецептор, сравнима с
Главный комплекс
гистосовместимости
MHC (Major histocompatibility complex), он же HLA (human leukocyte antigen) у
Главный комплекс
гистосовместимости
MHC (Major histocompatibility complex), он же HLA (human leukocyte antigen) у

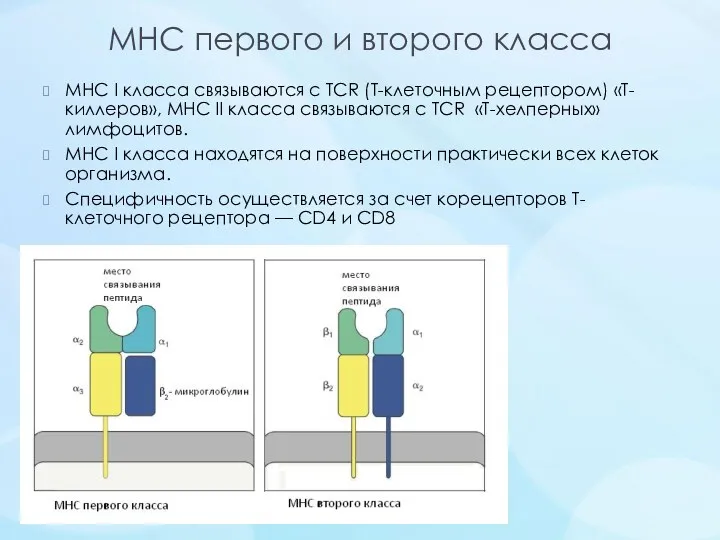
MHC первого и второго класса
MHC I класса связываются с TCR (Т-клеточным рецептором) «Т-киллеров»,
MHC первого и второго класса
MHC I класса связываются с TCR (Т-клеточным рецептором) «Т-киллеров»,
MHC I и II класса связываются
с пептидом-антигеном
МНС связываются с пептидным фрагментом антигена,
MHC I и II класса связываются
с пептидом-антигеном
МНС связываются с пептидным фрагментом антигена,
MHC-специфичный паттерн
связываемых пептидных фрагментов
МНС I и II класса связываются с пептидными фрагментами,
MHC-специфичный паттерн
связываемых пептидных фрагментов
МНС I и II класса связываются с пептидными фрагментами,
Презентация антигена
Общие сведения
Презентацией антигена называется предоставление для Т-лимфоцитов комплекса MHC-антиген клетками на своей
Презентация антигена
Общие сведения
Презентацией антигена называется предоставление для Т-лимфоцитов комплекса MHC-антиген клетками на своей
Процессинг антигена в клетке
Белки, находящиеся в цитозоли, рано или поздно попадают в протеасому.
Процессинг антигена в клетке
Белки, находящиеся в цитозоли, рано или поздно попадают в протеасому.
Транспорт MHC I класса в
эндоплазматический ретикулум
Для сборки MHC I класса необходимо связаться
Транспорт MHC I класса в
эндоплазматический ретикулум
Для сборки MHC I класса необходимо связаться

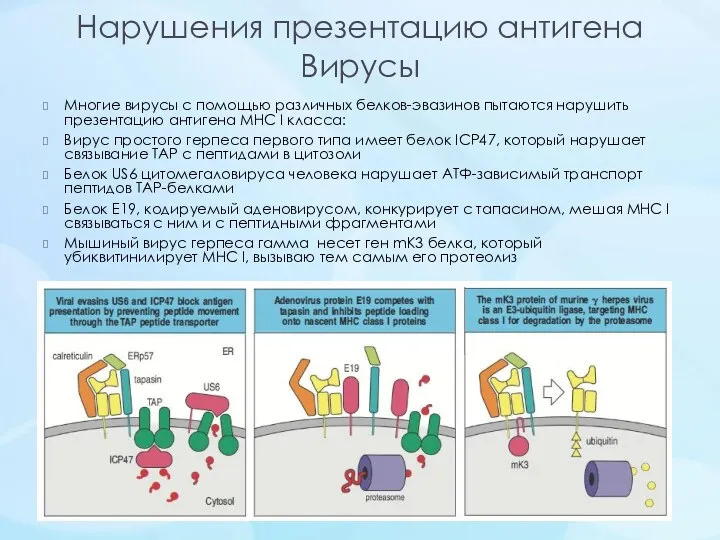
Нарушения презентацию антигена
Вирусы
Многие вирусы с помощью различных белков-эвазинов пытаются нарушить презентацию антигена MHC
Нарушения презентацию антигена
Вирусы
Многие вирусы с помощью различных белков-эвазинов пытаются нарушить презентацию антигена MHC
Нарушения презентации антигена
Опухолевые клетки
Опухолевые клетки часто характеризуются наличие так называемых «неоантигенов» — антигенов,
Нарушения презентации антигена
Опухолевые клетки
Опухолевые клетки часто характеризуются наличие так называемых «неоантигенов» — антигенов,

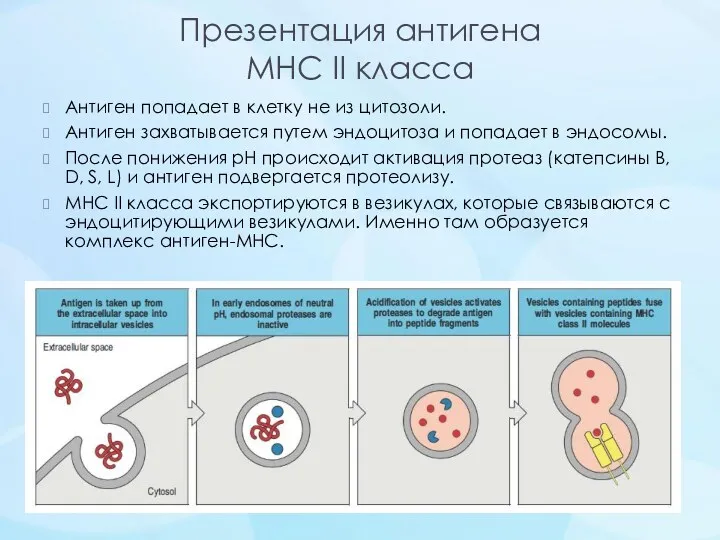
Презентация антигена
MHC II класса
Антиген попадает в клетку не из цитозоли.
Антиген захватывается путем
Презентация антигена
MHC II класса
Антиген попадает в клетку не из цитозоли.
Антиген захватывается путем
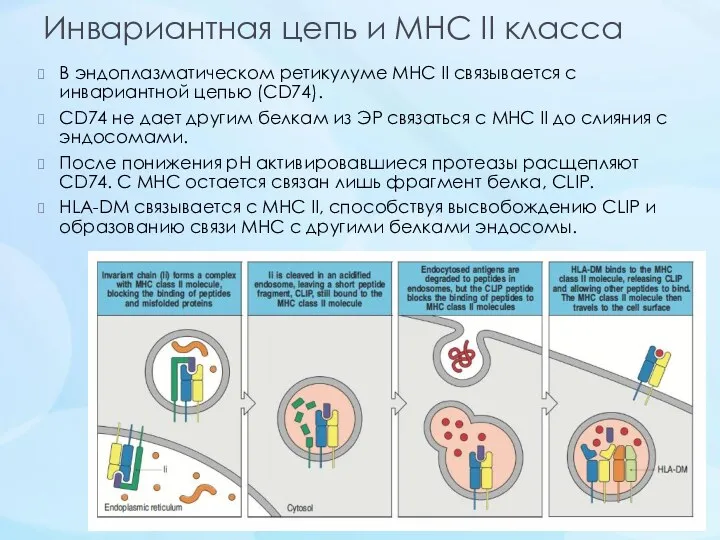
Инвариантная цепь и MHC II класса
В эндоплазматическом ретикулуме МНС II связывается с инвариантной
Инвариантная цепь и MHC II класса
В эндоплазматическом ретикулуме МНС II связывается с инвариантной
Нарушения презентации антигена
Паразиты
Toxoplasma gondii стимулирует экспрессию CD74 (инвариантной цепи) и подавляет экспрессию МНС
Нарушения презентации антигена
Паразиты
Toxoplasma gondii стимулирует экспрессию CD74 (инвариантной цепи) и подавляет экспрессию МНС

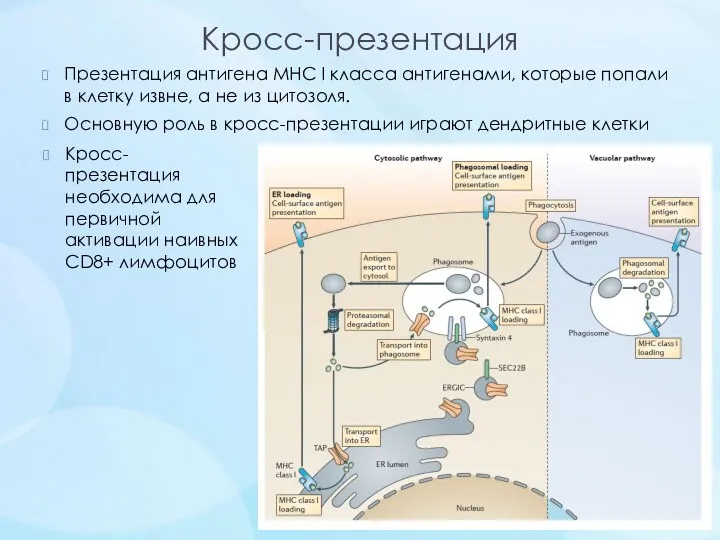
Кросс-презентация
Презентация антигена MHC I класса антигенами, которые попали в клетку извне, а
Кросс-презентация
Презентация антигена MHC I класса антигенами, которые попали в клетку извне, а
Презентация липидов:
CD1 и iNKT-клетки
CD1 – это белок, сходный по структуре с MHC I
Презентация липидов:
CD1 и iNKT-клетки
CD1 – это белок, сходный по структуре с MHC I
Профессиональные антиген-презентирующие клетки
Профессиональные АПК:
1) Дендритные клетки,
2) Макрофаги,
3) В-лимфоциты.
Профессиональные антиген-презентирующие клетки
Профессиональные АПК:
1) Дендритные клетки,
2) Макрофаги,
3) В-лимфоциты.
Презентация антигена
В-лимфоцитами
В-лимфоциты осуществляют антиген-специфичную презентацию за счет связывания антигена с антителом.
При связывании
Презентация антигена
В-лимфоцитами
В-лимфоциты осуществляют антиген-специфичную презентацию за счет связывания антигена с антителом.
При связывании
Дендритные клетки
1) Обнаруживание патогена (TLR, NOD etc.)
2) Антиген-презентация (MHC I, MHC II, CD1)
3)
Дендритные клетки
1) Обнаруживание патогена (TLR, NOD etc.)
2) Антиген-презентация (MHC I, MHC II, CD1)
3)
Подтипы дендритных клеток
Различают несколько подтипов дендритных клеток:
1) Миелоидные,
2) Плазмоцитоидные,
3) Интерстициальные
4) Клетки Лангерганса
Подтипы дендритных клеток
Различают несколько подтипов дендритных клеток:
1) Миелоидные,
2) Плазмоцитоидные,
3) Интерстициальные
4) Клетки Лангерганса
Спасибо за внимание!
Спасибо за внимание!

 Аускультация тәсілінің даму тарихы
Аускультация тәсілінің даму тарихы Нестабильная стенокардия
Нестабильная стенокардия Современные методы лечения скрытого немелкоклеточного рака легкого (НМРЛ)
Современные методы лечения скрытого немелкоклеточного рака легкого (НМРЛ) Инфекциялық процес сипаттамасы. Бактериялардың патогенділігі мен токсигенділігі. Вирустардың жұқпалылығы
Инфекциялық процес сипаттамасы. Бактериялардың патогенділігі мен токсигенділігі. Вирустардың жұқпалылығы Мышцы туловища, груди и живота
Мышцы туловища, груди и живота Практические основы клинической фармакологии. Седативно-гипнотические и метаболические средства
Практические основы клинической фармакологии. Седативно-гипнотические и метаболические средства Дифференциальная диагностика синдрома Рея и миопатии Дюшена. Клинический случай
Дифференциальная диагностика синдрома Рея и миопатии Дюшена. Клинический случай Неотложные состояния в гинекологии
Неотложные состояния в гинекологии Қышқыл-сілтінің тепетеңдігінің биохимиясы және клиникалық физиологиясы
Қышқыл-сілтінің тепетеңдігінің биохимиясы және клиникалық физиологиясы Кардиотонические средства
Кардиотонические средства Режимы дня, труда и отдыха
Режимы дня, труда и отдыха Эпидемиология, общие понятия, задачи, исследовательские методики
Эпидемиология, общие понятия, задачи, исследовательские методики Гастроэзофагальды ауру
Гастроэзофагальды ауру Здоровый образ жизни — образ жизни отдельного человека с целью профилактики болезней и укрепления здоровья
Здоровый образ жизни — образ жизни отдельного человека с целью профилактики болезней и укрепления здоровья Анатомия лимфатической системы
Анатомия лимфатической системы Пиодермии: классификация, симптомы, лечение
Пиодермии: классификация, симптомы, лечение Кисель Александр Андреевич (1859 - 1938)
Кисель Александр Андреевич (1859 - 1938) Буллезный дерматоз. Пузырчатка
Буллезный дерматоз. Пузырчатка Выписка, хранение, применение лекарственных средств, парентеральные и энтеральные способы введения
Выписка, хранение, применение лекарственных средств, парентеральные и энтеральные способы введения Гастрит ауруы қазақша
Гастрит ауруы қазақша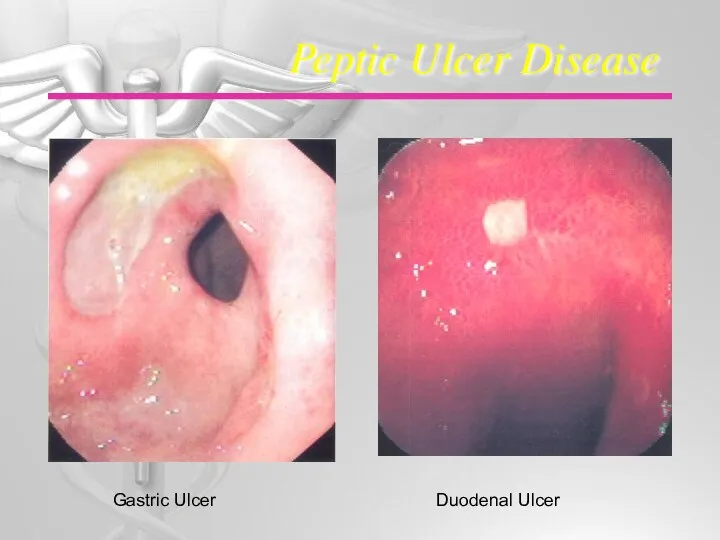 Peptic ulcer disease
Peptic ulcer disease Ұрықтың туа біткен патологиясының алдын алу және ерте диагностикасы. Жүкті әйелдің жеке картасын талдау
Ұрықтың туа біткен патологиясының алдын алу және ерте диагностикасы. Жүкті әйелдің жеке картасын талдау Современные алгоритмы лечения сахарного диабета 2 типа
Современные алгоритмы лечения сахарного диабета 2 типа Формирование воздушной струи в процессе преодоления нарушений звукопроизношения
Формирование воздушной струи в процессе преодоления нарушений звукопроизношения Гемолитические анемии
Гемолитические анемии Введение в фармакологию
Введение в фармакологию Лекарственные средства, влияющие на функцию органов дыхания
Лекарственные средства, влияющие на функцию органов дыхания Основные синдромы болезней нервной системы у животных
Основные синдромы болезней нервной системы у животных