- Частная физиология ЦНС

Содержание
- 2. Физиология спинного мозга Спинной мозг является наиболее простым, примитивным по строению и физиологическим функциям отделом центральной
- 3. С П И Н Н О Й М О З Г РЕФЛЕКТОРНАЯ ФУНКЦИЯ (РЕФЛЕКСЫ СПИННОГО МОЗГА)
- 4. Рефлекторная деятельность спинного мозга обеспечивает местные сегментарные рефлектор-ные акты организма. Как совокупность нервных центров спинной мозг
- 5. В спинном мозге расположены центры вегетативной нервной системы: 1. симпатического отдела, иннервирующие сердце, сосуды, органы пищеварения
- 6. СПИНАЛЬНАЯ ДВИГАТЕЛЬНАЯ СИСТЕМА В регуляции мышечной активности принимают участие: кора, подкорковые ядра, мозжечок, ствол мозга, спинной
- 7. СПИНАЛЬНЫЙ ШОК Спинальный шок – это обратимое угнетение двигательных и вегетатив-ных рефлексов спинного мозга вследствие выключения
- 8. Спинальный шок выражен у животных, головной мозг которых в большей степени преобладает над спинным мозгом. По
- 9. МЕХАНИЗМ СПИНАЛЬНОГО ШОКА Главным в механизме спинального шока является нарушение связей спинного мозга с остальными отделами
- 10. ПРИНЦИП ОБРАТНОЙ СВЯЗИ ИЛИ ВТОРИЧНОЙ АФФЕРЕНТАЦИИ В мышце имеются как экстрафу-зальные волокна, представляющие основную массу рабочих
- 11. В спинном мозге различают 2 механизма активации альфа-мотонейронов: 1. прямое влияние пирамид-ных клеток коры мозга на
- 12. СТРОЕНИЕ И ФУНКЦИЯ МЫШЕЧНЫХ ПРОПРИОРЕЦЕПТОРОВ Ядерносумчатое интрафузальное мышечное волокно одним концом крепится к ядерной сумке, другим
- 13. Рецепторы ядерной сумки возбуждаются тремя способами: 1. при расслаблении мышцы интрафузальное мышечное волокно растягивается, что ведет
- 15. ВЗАИМООТНОШЕНИЯ МЕЖДУ ЭКСТРАФУЗАЛЬНЫМИ И ИНТРАФУЗАЛЬНЫМИ МЫШЕЧНЫМИ ВОЛОКНАМИ: При сокращении экстрафузальных волокон происходит расслабление интрафузальных волокон, и,
- 16. Расслабление экстрафузальных волокон ведет к натяжению интрафу-зальных волокон, приводящее к раздражению рецепторов ядерной сумки. Импульсы от
- 18. МЕХАНИЗМ ШАГАТЕЛЬНОГО РЕФЛЕКСА: Когда необходимо повышение тонуса мышц сгибателей от гамма-мотонейронов спинного мозга следуют импульсы, ведущие
- 19. ВЛИЯНИЕ ДВИГАТЕЛЬНЫХ ЦЕНТРОВ ГОЛОВНОГО МОЗГА НА СПИННОЙ МОЗГ В нормальных условиях деятельность спинного мозга контролируется и
- 20. Нисходящие пути от супраспинальных отделов можно разделить на 2 группы, оказывающие противоположные влияния на нейроны спинного
- 21. ФИЗИОЛОГИЯ ПРОДОЛГОВАТОГО МОЗГА ПРОДОЛГОВАТЫЙ МОЗГ ТАКЖЕ ВЫПОЛНЯЕТ 2 ОСНОВНЫЕ ФУНКЦИИ: РЕФЛЕКТОРНУЮ И ПРОВОДНИКОВУЮ. Основная биологическая роль
- 22. ФИЗИОЛОГИЯ СРЕДНЕГО МОЗГА Главными образованиями среднего мозга являются: красное ядро, черная субстанция, ядра 3 и 4
- 23. Они обеспечивают ориентировочные рефлексы на свет: поворот глаз, головы, туловища в сторону источника света, а также
- 24. Черная субстанция выполняет следующие функции: участвует в сложных координацион-ных движениях акта еды (последо-вательность жевания и глотания);
- 25. Красное ядро играет важную роль при осуществлении движений. Получая информацию от двигательных нейронов коры головного мозга,
- 26. Функция красного ядра была установлена в опытах с перерезкой ствола мозга на уровне переднего края задних
- 28. Механизм децеребрационной ригидности Доказано, что нейроны красного ядра вызывают активацию альфа-мотонейронов преимущественно мышц-сгибателей и тормозят активность
- 29. В норме красное ядро тормозит активность вестибулярных ядер. При перерезке ствола мозга ниже красного ядра вестибуляр-ные
- 30. Децеребрационная ригидность не возникает, если перерезка мозга проведена ниже латерального вестибулярного ядра или нарушена целостность вестибуло-спинального
- 31. Децеребрационная ригидность не наступает при перерезке ствола мозга выше красного ядра. В этом случае красное ядро
- 32. ФИЗИОЛОГИЯ МОЗЖЕЧКА Мозжечок как надсегментарный орган, входящий в систему регуляции движений, выполняет следующие функции: 1. Координация
- 33. Механизм работы мозжечка: В мозжечок поступает информация от вестибулярного аппарата, мышеч-ных проприорецепторов, кожных рецепторов, коры мозга.
- 34. Функции мозжечка были изучены в опытах с экстирпацией мозжечка. Лючиани в 1893 г. описал симпто-мокомплекс моторных
- 35. Для поражения мозжечка характерна атаксия, т.е. нарушение координа-ции движений. Атаксия возникает вследствие прекращения поступле-ния в мозжечок
- 36. Мозжечок является подкорковым центром координации движений, посредством которого кора головного мозга осуществляет высшую координацию быстрых целенаправленных
- 37. Асинергия означает нарушение содружественной деятельности мышц, что ведет к расстройству движений, требующих одновременного сокращения нескольких групп
- 38. Механизм адиадохокинеза: при выполнении любого произ-вольного движения пирамидные клетки коры мозга посылают к мотонейронам спинного мозга
- 39. Влияние мозжечка облегчает спин-ному мозгу легкую смену движе-ния на противоположное, т.е. подгатавливает спинной мозг к выполнению
- 40. Мозжечок контролирует висцераль-ные функции организма. При раздражении мозжечка возникают расстройства, характерные для возбуждения симпатической нервной системы,
- 41. Нарушение функций мозжечка влияет на высшую нервную деятельность: ослабляется сила процессов возбуждения и тормо-жения, развивается инертность
- 42. Р Е Т И К У Л Я Р Н А Я Ф О Р М
- 43. Нейронная организация РФ Ретикулярная формация берет начало в шейной части спинного мозга между боковыми и задними
- 44. Морфологические особенности РФ: 1. Нейроны РФ имеют слабоветвящиеся дендриты. Аксоны делятся на восходя-щую и нисходящую ветви,
- 45. Связи РФ с другими отделами ЦНС РФ является структурой, расположен-ной на путях всех входящих в мозг
- 46. Нисходящие ретикуло-спинальные пути. Они берут начало от ядер заднего мозга и спускаются в спинной мозг по
- 47. Нисходящие влияния РФ По нисходящим путям РФ оказывает на спинной мозг как активирующее , так и
- 48. Тормозное влияние РФ на спинномозговые центры осуществляется двояким путем: 1. за счет ослабления сенсорного входа в
- 49. Активирующее влияние РФ на рефлекторную деятельность спинного мозга было обнаружено при раздраже-нии латеральных областей РФ вароли-ева
- 50. Активирующее влияние на спинной мозг осуществляется двумя путями: 1. путем снижения порога возбуди-мости нейронов спинного мозга.
- 51. РФ регулирует возбудимость альфа-мотонейронов частично по моносинап-тическим путям, частично через систему вставочных нейронов и через гамма-мотонейронов
- 52. Регуляция мышечного тонуса происходит, в основном, при участии покрышки среднего мозга по двум ретикулоспинальным путям: быстропроводящему
- 53. Восходящие ретикулярные влияния Восходящие ретикуло-кортикальные влияния были открыты Моруцци и Мегуном в конце 40-х годов прошлого
- 54. Регистрируется α-ритм в положении лежа или сидя с закрытыми глазами и с расслабленной мускулатурой. β-ритм, частота
- 55. Методы исследования. РИТМЫ БОДРСТВОВАНИЯ : Бета-ритм: 14 – 30 в сек, 25 мкВ. Альфа-ритм: 8 –
- 56. Анализ частотного спектра ЭЭГ позволяет судить о функциональном состоянии коры головного мозга. Моруцци и Мегун, раздражая
- 57. Реакцию активации в ЭЭГ можно вызвать при раздражении различных отделов продолговатого и среднего мозга, таламуса, суб-
- 58. Активирующее восходящее влияние РФ на кору головного мозга является непременным условием поддержания бодрствующего состояния мозга. Если
- 59. Активация коры мозга может возникать при любом афферентном воздействии. На уровне ствола мозга афферентное возбуждение, возникающее
- 60. Пример: классический лемнисковый путь, передающий в мозг сигналы о прикосновении к коже, давления на нее ,
- 61. От ретикулярной формации возбуж-дение в виде мощных восходящих потоков направляется к коре головного мозга, генерализованно активируя
- 63. Если раздражение ретикулярных ядер ствола мозга вызывает генерализован-ное активирующее действие, то раздражение ретикулярных ядер таламуса проявляет
- 64. Существует корковый контроль над деятельностью РФ. Воздействуя на нейроны РФ, кора головного мозга способна изменять их
- 65. Таким образом, ретикулярная формация является структурой, отвечающей за состояние бодрствования, структурой, формирующей восходящую активирующую ретикулярную систему,
- 76. Скачать презентацию

Физиология спинного мозга
Спинной мозг является наиболее простым, примитивным по строению
Физиология спинного мозга
Спинной мозг является наиболее простым, примитивным по строению
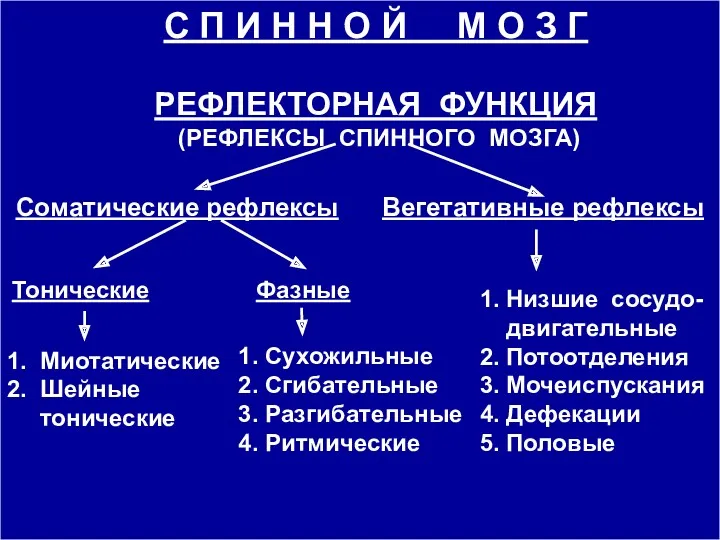
С П И Н Н О Й М О З Г
РЕФЛЕКТОРНАЯ
С П И Н Н О Й М О З Г
РЕФЛЕКТОРНАЯ

Рефлекторная деятельность спинного мозга обеспечивает местные сегментарные рефлектор-ные акты
Рефлекторная деятельность спинного мозга обеспечивает местные сегментарные рефлектор-ные акты

В спинном мозге расположены центры вегетативной нервной системы:
1. симпатического
В спинном мозге расположены центры вегетативной нервной системы:
1. симпатического

СПИНАЛЬНАЯ ДВИГАТЕЛЬНАЯ СИСТЕМА
В регуляции мышечной активности принимают участие: кора,
СПИНАЛЬНАЯ ДВИГАТЕЛЬНАЯ СИСТЕМА
В регуляции мышечной активности принимают участие: кора,

СПИНАЛЬНЫЙ ШОК
Спинальный шок – это обратимое угнетение двигательных
СПИНАЛЬНЫЙ ШОК
Спинальный шок – это обратимое угнетение двигательных

Спинальный шок выражен у животных, головной мозг которых в большей степени

МЕХАНИЗМ СПИНАЛЬНОГО ШОКА
Главным в механизме спинального шока является
МЕХАНИЗМ СПИНАЛЬНОГО ШОКА
Главным в механизме спинального шока является

ПРИНЦИП ОБРАТНОЙ СВЯЗИ ИЛИ ВТОРИЧНОЙ АФФЕРЕНТАЦИИ
В мышце имеются
ПРИНЦИП ОБРАТНОЙ СВЯЗИ ИЛИ ВТОРИЧНОЙ АФФЕРЕНТАЦИИ
В мышце имеются

В спинном мозге различают 2 механизма активации альфа-мотонейронов:
1. прямое влияние
В спинном мозге различают 2 механизма активации альфа-мотонейронов:
1. прямое влияние

СТРОЕНИЕ И ФУНКЦИЯ МЫШЕЧНЫХ ПРОПРИОРЕЦЕПТОРОВ
Ядерносумчатое интрафузальное мышечное волокно
СТРОЕНИЕ И ФУНКЦИЯ МЫШЕЧНЫХ ПРОПРИОРЕЦЕПТОРОВ
Ядерносумчатое интрафузальное мышечное волокно

Рецепторы ядерной сумки
возбуждаются тремя способами:
1. при расслаблении
Рецепторы ядерной сумки
возбуждаются тремя способами:
1. при расслаблении


ВЗАИМООТНОШЕНИЯ МЕЖДУ ЭКСТРАФУЗАЛЬНЫМИ И ИНТРАФУЗАЛЬНЫМИ МЫШЕЧНЫМИ ВОЛОКНАМИ:
При сокращении экстрафузальных волокон происходит
ВЗАИМООТНОШЕНИЯ МЕЖДУ ЭКСТРАФУЗАЛЬНЫМИ И ИНТРАФУЗАЛЬНЫМИ МЫШЕЧНЫМИ ВОЛОКНАМИ:
При сокращении экстрафузальных волокон происходит

Расслабление экстрафузальных волокон ведет к натяжению интрафу-зальных волокон, приводящее к раздражению
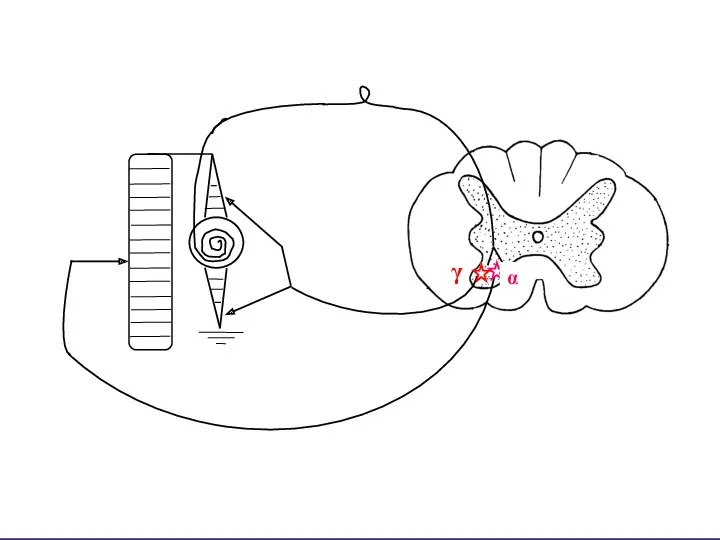

МЕХАНИЗМ ШАГАТЕЛЬНОГО РЕФЛЕКСА:
Когда необходимо повышение тонуса
мышц сгибателей
МЕХАНИЗМ ШАГАТЕЛЬНОГО РЕФЛЕКСА:
Когда необходимо повышение тонуса
мышц сгибателей
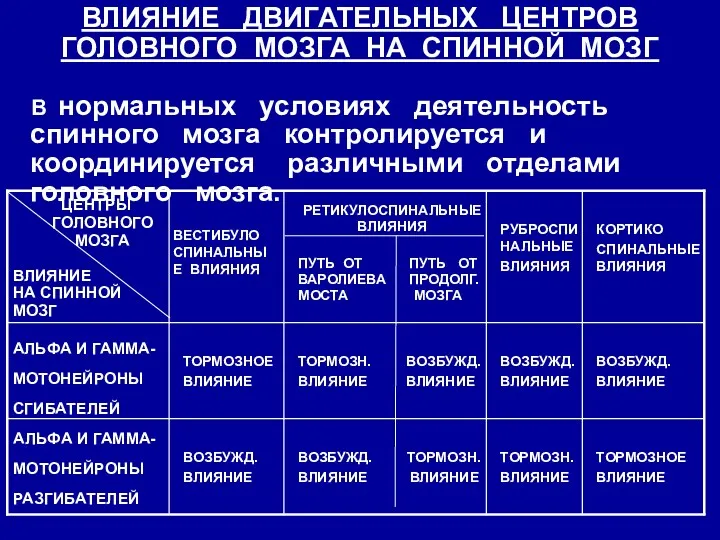
ВЛИЯНИЕ ДВИГАТЕЛЬНЫХ ЦЕНТРОВ ГОЛОВНОГО МОЗГА НА СПИННОЙ МОЗГ
В нормальных условиях
ВЛИЯНИЕ ДВИГАТЕЛЬНЫХ ЦЕНТРОВ ГОЛОВНОГО МОЗГА НА СПИННОЙ МОЗГ
В нормальных условиях

Нисходящие пути от супраспинальных отделов можно разделить на 2 группы, оказывающие

ФИЗИОЛОГИЯ ПРОДОЛГОВАТОГО МОЗГА
ПРОДОЛГОВАТЫЙ МОЗГ ТАКЖЕ ВЫПОЛНЯЕТ 2 ОСНОВНЫЕ ФУНКЦИИ:
ФИЗИОЛОГИЯ ПРОДОЛГОВАТОГО МОЗГА
ПРОДОЛГОВАТЫЙ МОЗГ ТАКЖЕ ВЫПОЛНЯЕТ 2 ОСНОВНЫЕ ФУНКЦИИ:

ФИЗИОЛОГИЯ СРЕДНЕГО МОЗГА
Главными образованиями среднего мозга являются: красное ядро, черная
ФИЗИОЛОГИЯ СРЕДНЕГО МОЗГА
Главными образованиями среднего мозга являются: красное ядро, черная

Они обеспечивают ориентировочные рефлексы на свет: поворот глаз, головы, туловища в

Черная субстанция выполняет следующие функции:
участвует в сложных координацион-ных движениях акта еды
участвует в сложных координацион-ных движениях акта еды

Красное ядро играет важную роль при осуществлении движений. Получая информацию от

Функция красного ядра была установлена в опытах с перерезкой ствола мозга
Функция красного ядра была установлена в опытах с перерезкой ствола мозга
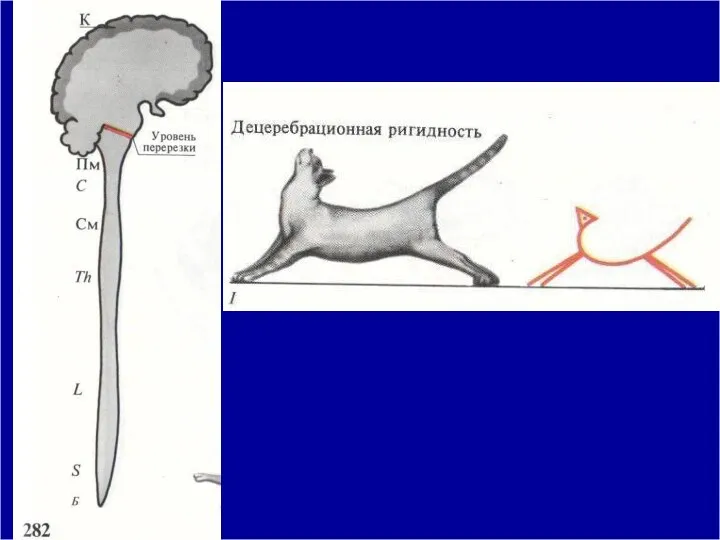

Механизм децеребрационной ригидности
Доказано, что нейроны красного ядра вызывают активацию альфа-мотонейронов
Механизм децеребрационной ригидности
Доказано, что нейроны красного ядра вызывают активацию альфа-мотонейронов

В норме красное ядро тормозит активность вестибулярных ядер. При перерезке ствола

Децеребрационная ригидность не возникает, если перерезка мозга проведена ниже латерального вестибулярного
Децеребрационная ригидность не возникает, если перерезка мозга проведена ниже латерального вестибулярного

Децеребрационная ригидность не наступает при перерезке ствола мозга выше красного ядра.
Децеребрационная ригидность не наступает при перерезке ствола мозга выше красного ядра.

ФИЗИОЛОГИЯ МОЗЖЕЧКА
Мозжечок как надсегментарный орган, входящий в систему регуляции движений,
ФИЗИОЛОГИЯ МОЗЖЕЧКА
Мозжечок как надсегментарный орган, входящий в систему регуляции движений,

Механизм работы мозжечка:
В мозжечок поступает информация от вестибулярного аппарата, мышеч-ных
Механизм работы мозжечка:
В мозжечок поступает информация от вестибулярного аппарата, мышеч-ных

Функции мозжечка были изучены в опытах с экстирпацией мозжечка. Лючиани в
Функции мозжечка были изучены в опытах с экстирпацией мозжечка. Лючиани в

Для поражения мозжечка характерна атаксия, т.е. нарушение координа-ции движений. Атаксия возникает
Для поражения мозжечка характерна атаксия, т.е. нарушение координа-ции движений. Атаксия возникает

Мозжечок является подкорковым центром координации движений, посредством которого кора головного мозга
Мозжечок является подкорковым центром координации движений, посредством которого кора головного мозга

Асинергия означает нарушение содружественной деятельности мышц, что ведет к расстройству движений,
Асинергия означает нарушение содружественной деятельности мышц, что ведет к расстройству движений,

Механизм адиадохокинеза:
при выполнении любого произ-вольного движения пирамидные клетки коры мозга посылают
Механизм адиадохокинеза:
при выполнении любого произ-вольного движения пирамидные клетки коры мозга посылают

Влияние мозжечка облегчает спин-ному мозгу легкую смену движе-ния на противоположное, т.е.
Влияние мозжечка облегчает спин-ному мозгу легкую смену движе-ния на противоположное, т.е.

Мозжечок контролирует висцераль-ные функции организма. При раздражении мозжечка возникают расстройства, характерные
Мозжечок контролирует висцераль-ные функции организма. При раздражении мозжечка возникают расстройства, характерные

Нарушение функций мозжечка влияет на высшую нервную деятельность: ослабляется сила процессов
Нарушение функций мозжечка влияет на высшую нервную деятельность: ослабляется сила процессов

Р Е Т И К У Л Я Р Н А

Нейронная организация РФ
Ретикулярная формация берет начало в шейной части спинного
Нейронная организация РФ
Ретикулярная формация берет начало в шейной части спинного

Морфологические особенности РФ:
1. Нейроны РФ имеют слабоветвящиеся дендриты. Аксоны делятся на
Морфологические особенности РФ:
1. Нейроны РФ имеют слабоветвящиеся дендриты. Аксоны делятся на

Связи РФ с другими отделами ЦНС
РФ является структурой, расположен-ной
Связи РФ с другими отделами ЦНС
РФ является структурой, расположен-ной

Нисходящие ретикуло-спинальные пути.
Они берут начало от ядер заднего мозга и
Нисходящие ретикуло-спинальные пути.
Они берут начало от ядер заднего мозга и

Нисходящие влияния РФ
По нисходящим путям РФ оказывает на спинной
Нисходящие влияния РФ
По нисходящим путям РФ оказывает на спинной

Тормозное влияние РФ на спинномозговые центры осуществляется двояким путем:
1. за
Тормозное влияние РФ на спинномозговые центры осуществляется двояким путем:
1. за

Активирующее влияние РФ на рефлекторную деятельность спинного мозга было обнаружено при
Активирующее влияние РФ на рефлекторную деятельность спинного мозга было обнаружено при

Активирующее влияние на спинной мозг осуществляется двумя путями:
1. путем
Активирующее влияние на спинной мозг осуществляется двумя путями:
1. путем

РФ регулирует возбудимость альфа-мотонейронов частично по моносинап-тическим путям, частично через систему

Регуляция мышечного тонуса происходит, в основном, при участии покрышки
Регуляция мышечного тонуса происходит, в основном, при участии покрышки

Восходящие ретикулярные влияния
Восходящие ретикуло-кортикальные влияния были открыты Моруцци и Мегуном
Восходящие ретикулярные влияния
Восходящие ретикуло-кортикальные влияния были открыты Моруцци и Мегуном

Регистрируется α-ритм в положении лежа или сидя с закрытыми глазами
Регистрируется α-ритм в положении лежа или сидя с закрытыми глазами
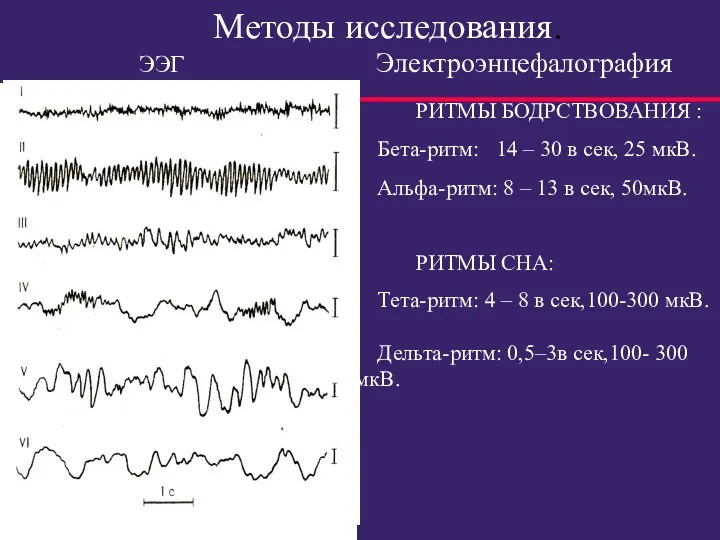
Методы исследования.
РИТМЫ БОДРСТВОВАНИЯ :
Бета-ритм: 14 – 30 в
Методы исследования.
РИТМЫ БОДРСТВОВАНИЯ :
Бета-ритм: 14 – 30 в

Анализ частотного спектра ЭЭГ позволяет судить о функциональном состоянии коры головного
Анализ частотного спектра ЭЭГ позволяет судить о функциональном состоянии коры головного

Реакцию активации в ЭЭГ можно вызвать при раздражении различных отделов продолговатого
Реакцию активации в ЭЭГ можно вызвать при раздражении различных отделов продолговатого

Активирующее восходящее влияние РФ на кору головного мозга является непременным условием
Активирующее восходящее влияние РФ на кору головного мозга является непременным условием

Активация коры мозга может возникать при любом афферентном воздействии. На уровне
Активация коры мозга может возникать при любом афферентном воздействии. На уровне

Пример: классический лемнисковый путь, передающий в мозг сигналы о прикосновении к
Пример: классический лемнисковый путь, передающий в мозг сигналы о прикосновении к

От ретикулярной формации возбуж-дение в виде мощных восходящих потоков направляется к
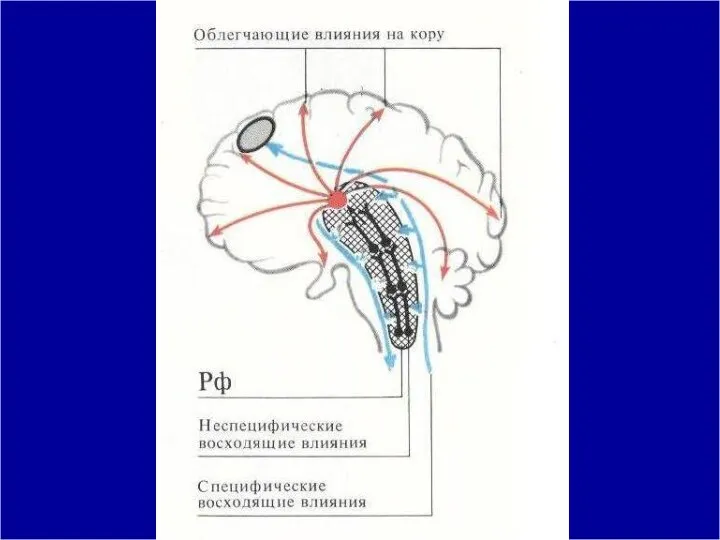

Если раздражение ретикулярных ядер ствола мозга вызывает генерализован-ное активирующее действие, то

Существует корковый контроль над деятельностью РФ. Воздействуя на нейроны РФ, кора

Таким образом, ретикулярная формация является структурой, отвечающей за состояние бодрствования, структурой,
Таким образом, ретикулярная формация является структурой, отвечающей за состояние бодрствования, структурой,









 Гормондар туралы жалпы түсінік
Гормондар туралы жалпы түсінік Сочетанные и комбинированные ранения лица
Сочетанные и комбинированные ранения лица Эффект Допплера
Эффект Допплера Introduction in Medical informatics
Introduction in Medical informatics Нарушения мозгового кровообращения на догоспитальном этапе
Нарушения мозгового кровообращения на догоспитальном этапе Егде жастағы және қарттардың тамақтануы
Егде жастағы және қарттардың тамақтануы Первая помощь при обморожении, ожогах. Профилактика
Первая помощь при обморожении, ожогах. Профилактика Синдром поликистозных яичников
Синдром поликистозных яичников Вступ до хірургії. Базові хірургічні навички
Вступ до хірургії. Базові хірургічні навички Мочевыделительная система
Мочевыделительная система Методы исследования и синдромы патологии эндокринных органов
Методы исследования и синдромы патологии эндокринных органов Воспалительные заболевания слюнных желез у детей. Клиника, диагностика и лечение
Воспалительные заболевания слюнных желез у детей. Клиника, диагностика и лечение Сколиоз, мүсіннің бұзылыстары
Сколиоз, мүсіннің бұзылыстары Abnormalities in the number of teeth
Abnormalities in the number of teeth Введение в фармакологию. Развитие лекарствоведения и история фармакологии. Общая фармакология
Введение в фармакологию. Развитие лекарствоведения и история фармакологии. Общая фармакология Здравницы Казахстана
Здравницы Казахстана Лептоспироз. Возбудители болезни
Лептоспироз. Возбудители болезни Нарушения памяти в неврологической практике
Нарушения памяти в неврологической практике Цереброваскулярные заболевания: ОНМК
Цереброваскулярные заболевания: ОНМК Радиационные поражения
Радиационные поражения Помощь умирающему человеку
Помощь умирающему человеку Я то, что я ем
Я то, что я ем Рак предстательной железы
Рак предстательной железы Симптоматология заболеваний эндокринной системы
Симптоматология заболеваний эндокринной системы Принципы и современные методы лечения переломов. Несросшиеся переломы, ложные суставы
Принципы и современные методы лечения переломов. Несросшиеся переломы, ложные суставы Противоэпидемические мероприятия в эпидемическом очаге инфекционного заболевания
Противоэпидемические мероприятия в эпидемическом очаге инфекционного заболевания Гравидограмманы түсіндіру
Гравидограмманы түсіндіру Токсикология, токсикокинетика, токсикодинамика, токсикометрия. (Лекция 2)
Токсикология, токсикокинетика, токсикодинамика, токсикометрия. (Лекция 2)