- COVID-19. Этиология, патогенез

Содержание
- 2. В конце 2019 года в Китайской Народной Республике (КНР) произошла вспышка новой коронавирусной инфекции с эпицентром
- 3. Всемирная организация здравоохранения (ВОЗ) 11 февраля 2020 г. Присвоила официальное название инфекции, вызванной новым коронавирусом –
- 4. Определение Коронавирусная инфекция – острое вирусное заболевание с преимущественным поражением нижних дыхательных путей, вызываемое РНК-содержащим вирусом
- 5. Таксономическое положение Коронавирусы (Coronaviridae) – семейство, включающее на январь 2020 года 40 видов РНК-геномных сложноорганизованных вирусов,
- 6. В настоящее время известно о циркуляции среди населения четырёх коронавирусов (HCoV-229E, -OC43, -NL63 и –HKU1), которые
- 7. До 2002 г. коронавирусы рассматривались в качестве агентов, вызывающих нетяжёлые заболевания верхних дыхательных путей ( с
- 8. В конце 2002 г. появился коронавирус (SARS-CoV), возбудитель атипичной пневмонии, который вызывал ТОРС у людей. Данный
- 9. В 2012 г. зарегистрирован новый коронавирус MERS (MERS-CoV), возбудитель ближневосточного респираторного синдрома, также принадлежащий к роду
- 10. Международный комитет по таксономии вирусов 11 февраля 2020 г. присвоил официальное название возбудителю инфекции – SARS-CoV-2.
- 11. Таксономическое положение отряд Nidovirales семейство Coronaviridae подсемейство Letovirinae: род – Letovirus (содержит 1 вирус) подсемейство Orthocoronaviridae:
- 12. Морфология Размер вириона 80-220 нм. Тип симметрии – спиральный: нуклеокапсид представляет собой гибкую спираль, состоящую из
- 13. Строение коронавируса
- 15. Схема строения вириона Сферические частицы диаметром 120 нм; Оболочка вириона содержит булавовидные отростки (S, spike); Белок
- 16. Структура генома Геномная РНК содержит 2 основные, длинные рамки считывания, занимающие около 70% генома ORF1a и
- 17. Геном РНК плюс нить. Коронавирус SARS-CoV-2 предположительно является рекомбинантным вирусом между коронавирусом летучих мышей и неизвестным
- 18. Репликация коронавируса
- 19. Жизненный цикл коронавирусов Проникновение вируса в клетку с помощью S белка (рецептор для 2019-nCoV – ангиотензинсвязывающий
- 20. Репродукция Коронавирусы адсорбируются на клетке (1) при помощи гликопротеина S, проникают в клетку при слиянии оболочки
- 21. При трансляции каждой субгеномной иРНК образуется один белок (7). Белок N связывается в цитоплазме с геномной
- 22. Жизненный цикл Первым этапом жизненного цикла вируса является адсорбция вируса на поверхности клетки-мишени в результате специфического
- 23. Жизненный цикл АСЕ2 не является универсальным рецептором для всех коронавирусов: MERS-CoV использует в качестве рецептора дипептилпептидазу4
- 24. Рецепторный эндоцитоз завершается проникновением вируса в цитоплазму клетки-хозяина, где вирионная РНК выступает в качестве мРНК для
- 25. Важнейшим неструктурным белком является РНК-зависимая РНК-полимераза (RdRp – RNA-dependent RNA-polymerase), которая синтезирует комплементарную вирионной нить РНК
- 26. сгРНК являются матрицей для синтеза субгеномных матричных РНК позитивной полярности, с которых считываются структурные белки. Сборка
- 27. Вирус 2019-nCoV
- 28. Характеристика вируса Впервые обнаружен на оптовом рынке морепродуктов (в продаже змеи, летучие мыши и пр.); Имеет
- 29. Доказательства патогенности SARS-CoV-2 30 декабря 2019 г. 3 образца БАЛ от пациента с внебольничной пневмонией RT-PCR:
- 30. Культивирование Вирусы культивируют на культуре клеток: Перевиваемые клетки HeLa; Первично-трипсинизированная культура клеток почек эмбриона свиньи
- 31. Резистентность Устойчивость в окружающей среде низкая: Погибает под воздействием УФО. Дезинфекционных средств. При нагревании до 40°С
- 32. Сохранение коронавируса SARS-CoV-2 на различных поверхностях Аэрозоли – 3 часа Пластик – 5 дней Алюминий –
- 33. Патогенез Патогенез новой коронавирусной инфекции изучен недостаточно Данные о длительности и напряжённости иммунитета в отношении SARS-CoV-2
- 34. Клетки-мишени Входные ворота возбудителя – эпителий верхних дыхательных путей и эпителиоциты желудка и кишечника. Начальным этапом
- 35. Патогенез Развивается диффузное альвеолярное повреждение. Вирус вызывает повышение проницаемости мембран клеток и усиленный транспорт жидкости, богатой
- 36. Экссудативная (острая) стадия Повреждение альвеоцитов I типа →повышение проницаемости альвеоло-капиллярной мембраны клеток→интерстициальный и альвеолярный отёк→заполнение альвеол
- 37. Пролиферативная (подострая) стадия Повреждение альвеоцитов II типа→миграция фибробластов в альвеолярный экссудат→пролиферация альвеоцитов II типа→уменьшение отёка лёгких
- 38. Фибропролиферативная (хроническая) стадия Облитерация альвеол→выраженный фиброз лёгочной паренхимы
- 40. Скачать презентацию

В конце 2019 года в Китайской Народной Республике (КНР) произошла вспышка
В конце 2019 года в Китайской Народной Республике (КНР) произошла вспышка

Всемирная организация здравоохранения (ВОЗ) 11 февраля 2020 г. Присвоила официальное название
Всемирная организация здравоохранения (ВОЗ) 11 февраля 2020 г. Присвоила официальное название

Определение
Коронавирусная инфекция – острое вирусное заболевание с преимущественным поражением нижних дыхательных
Определение
Коронавирусная инфекция – острое вирусное заболевание с преимущественным поражением нижних дыхательных

Таксономическое положение
Коронавирусы (Coronaviridae) – семейство, включающее на январь 2020 года 40
Таксономическое положение
Коронавирусы (Coronaviridae) – семейство, включающее на январь 2020 года 40

В настоящее время известно о циркуляции среди населения четырёх коронавирусов (HCoV-229E,
В настоящее время известно о циркуляции среди населения четырёх коронавирусов (HCoV-229E,

До 2002 г. коронавирусы рассматривались в качестве агентов, вызывающих нетяжёлые заболевания
До 2002 г. коронавирусы рассматривались в качестве агентов, вызывающих нетяжёлые заболевания

В конце 2002 г. появился коронавирус (SARS-CoV), возбудитель атипичной пневмонии, который
В конце 2002 г. появился коронавирус (SARS-CoV), возбудитель атипичной пневмонии, который

В 2012 г. зарегистрирован новый коронавирус MERS (MERS-CoV), возбудитель ближневосточного респираторного
В 2012 г. зарегистрирован новый коронавирус MERS (MERS-CoV), возбудитель ближневосточного респираторного

Международный комитет по таксономии вирусов 11 февраля 2020 г. присвоил официальное
Международный комитет по таксономии вирусов 11 февраля 2020 г. присвоил официальное

Таксономическое положение
отряд Nidovirales
семейство Coronaviridae
подсемейство Letovirinae:
род – Letovirus (содержит 1 вирус)
подсемейство Orthocoronaviridae:
род
Таксономическое положение
отряд Nidovirales
семейство Coronaviridae
подсемейство Letovirinae:
род – Letovirus (содержит 1 вирус)
подсемейство Orthocoronaviridae:
род

Морфология
Размер вириона 80-220 нм.
Тип симметрии – спиральный: нуклеокапсид представляет собой гибкую
Морфология
Размер вириона 80-220 нм.
Тип симметрии – спиральный: нуклеокапсид представляет собой гибкую
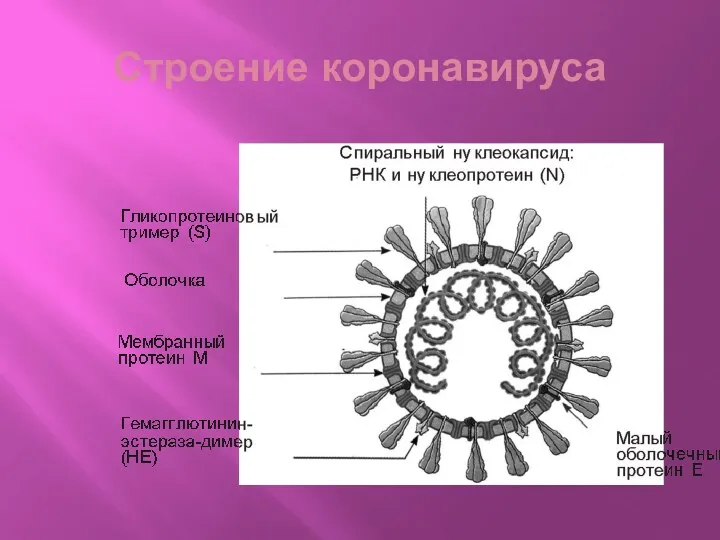
Строение коронавируса
Строение коронавируса
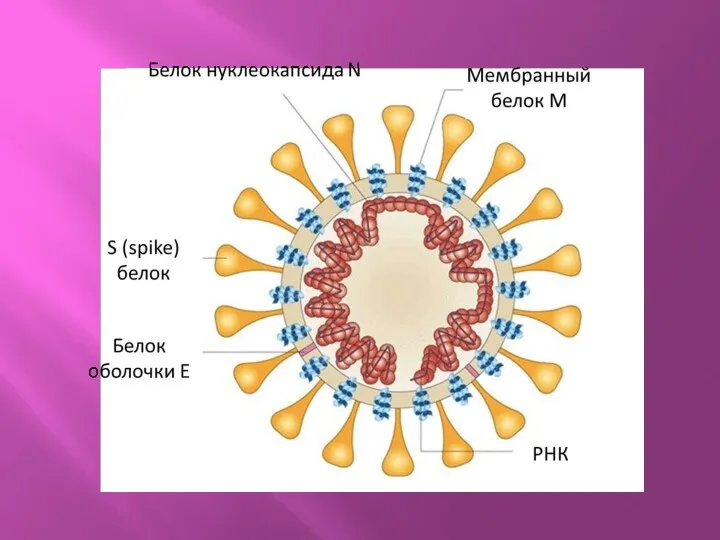

Схема строения вириона
Сферические частицы диаметром 120 нм;
Оболочка вириона содержит булавовидные отростки
Схема строения вириона
Сферические частицы диаметром 120 нм;
Оболочка вириона содержит булавовидные отростки

Структура генома
Геномная РНК содержит 2 основные, длинные рамки считывания, занимающие около
Структура генома
Геномная РНК содержит 2 основные, длинные рамки считывания, занимающие около

Геном
РНК плюс нить.
Коронавирус SARS-CoV-2 предположительно является рекомбинантным вирусом между коронавирусом летучих
Геном
РНК плюс нить.
Коронавирус SARS-CoV-2 предположительно является рекомбинантным вирусом между коронавирусом летучих
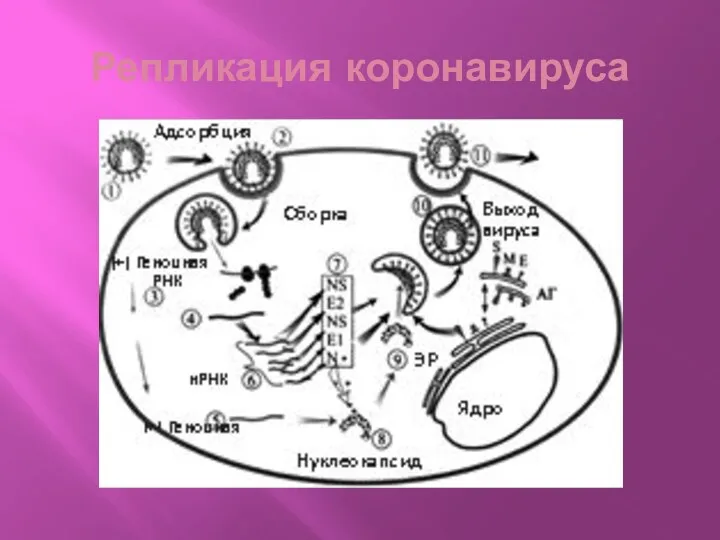
Репликация коронавируса
Репликация коронавируса

Жизненный цикл коронавирусов
Проникновение вируса в клетку с помощью S белка
Жизненный цикл коронавирусов
Проникновение вируса в клетку с помощью S белка

Репродукция
Коронавирусы адсорбируются на клетке (1) при помощи гликопротеина S, проникают в
Репродукция
Коронавирусы адсорбируются на клетке (1) при помощи гликопротеина S, проникают в

При трансляции каждой субгеномной иРНК образуется один белок (7). Белок N
При трансляции каждой субгеномной иРНК образуется один белок (7). Белок N

Жизненный цикл
Первым этапом жизненного цикла вируса является адсорбция вируса на поверхности
Жизненный цикл
Первым этапом жизненного цикла вируса является адсорбция вируса на поверхности

Жизненный цикл
АСЕ2 не является универсальным рецептором для всех коронавирусов:
MERS-CoV использует в
Жизненный цикл
АСЕ2 не является универсальным рецептором для всех коронавирусов:
MERS-CoV использует в

Рецепторный эндоцитоз завершается проникновением вируса в цитоплазму клетки-хозяина, где вирионная РНК
Рецепторный эндоцитоз завершается проникновением вируса в цитоплазму клетки-хозяина, где вирионная РНК

Важнейшим неструктурным белком является РНК-зависимая РНК-полимераза (RdRp – RNA-dependent RNA-polymerase), которая
Важнейшим неструктурным белком является РНК-зависимая РНК-полимераза (RdRp – RNA-dependent RNA-polymerase), которая

сгРНК являются матрицей для синтеза субгеномных матричных РНК позитивной полярности, с
сгРНК являются матрицей для синтеза субгеномных матричных РНК позитивной полярности, с
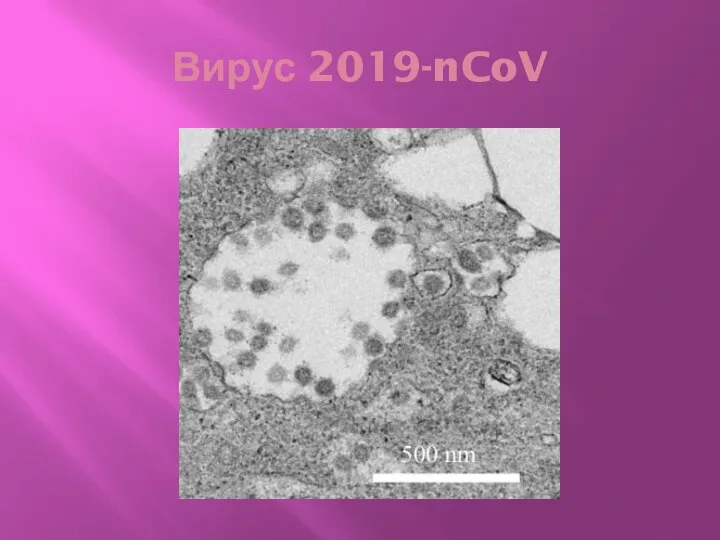
Вирус 2019-nCoV
Вирус 2019-nCoV

Характеристика вируса
Впервые обнаружен на оптовом рынке морепродуктов (в продаже змеи, летучие
Характеристика вируса
Впервые обнаружен на оптовом рынке морепродуктов (в продаже змеи, летучие

Доказательства патогенности SARS-CoV-2
30 декабря 2019 г.
3 образца БАЛ от пациента
Доказательства патогенности SARS-CoV-2
30 декабря 2019 г.
3 образца БАЛ от пациента

Культивирование
Вирусы культивируют на культуре клеток:
Перевиваемые клетки HeLa;
Первично-трипсинизированная культура клеток почек эмбриона
Культивирование
Вирусы культивируют на культуре клеток:
Перевиваемые клетки HeLa;
Первично-трипсинизированная культура клеток почек эмбриона

Резистентность
Устойчивость в окружающей среде низкая:
Погибает под воздействием УФО.
Дезинфекционных средств.
При нагревании до
Резистентность
Устойчивость в окружающей среде низкая:
Погибает под воздействием УФО.
Дезинфекционных средств.
При нагревании до

Сохранение коронавируса SARS-CoV-2 на различных поверхностях
Аэрозоли – 3 часа
Пластик –
Сохранение коронавируса SARS-CoV-2 на различных поверхностях
Аэрозоли – 3 часа
Пластик –

Патогенез
Патогенез новой коронавирусной инфекции изучен недостаточно
Данные о длительности и напряжённости иммунитета
Патогенез
Патогенез новой коронавирусной инфекции изучен недостаточно
Данные о длительности и напряжённости иммунитета

Клетки-мишени
Входные ворота возбудителя – эпителий верхних дыхательных путей и эпителиоциты желудка
Клетки-мишени
Входные ворота возбудителя – эпителий верхних дыхательных путей и эпителиоциты желудка

Патогенез
Развивается диффузное альвеолярное повреждение. Вирус вызывает повышение проницаемости мембран клеток и
Патогенез
Развивается диффузное альвеолярное повреждение. Вирус вызывает повышение проницаемости мембран клеток и

Экссудативная (острая) стадия
Повреждение альвеоцитов I типа →повышение проницаемости альвеоло-капиллярной мембраны клеток→интерстициальный
Экссудативная (острая) стадия
Повреждение альвеоцитов I типа →повышение проницаемости альвеоло-капиллярной мембраны клеток→интерстициальный

Пролиферативная (подострая) стадия
Повреждение альвеоцитов II типа→миграция фибробластов в альвеолярный экссудат→пролиферация альвеоцитов
Пролиферативная (подострая) стадия
Повреждение альвеоцитов II типа→миграция фибробластов в альвеолярный экссудат→пролиферация альвеоцитов

Фибропролиферативная (хроническая) стадия
Облитерация альвеол→выраженный фиброз лёгочной паренхимы
Фибропролиферативная (хроническая) стадия
Облитерация альвеол→выраженный фиброз лёгочной паренхимы
 Общая фармакология. Фармакодинамика
Общая фармакология. Фармакодинамика Острая лучевая болезнь
Острая лучевая болезнь Омыртқа жотасының қисаюы
Омыртқа жотасының қисаюы Болезнь Паркинсона. Редкие симптомы
Болезнь Паркинсона. Редкие симптомы Алгоритм відновлення прохідності верхніх дихальних шляхів. Прийом Геймліха
Алгоритм відновлення прохідності верхніх дихальних шляхів. Прийом Геймліха Инфекционная безопасность. Профилактика внутрибольничной инфекции
Инфекционная безопасность. Профилактика внутрибольничной инфекции Пероральные сахароснижающие препараты
Пероральные сахароснижающие препараты Техника и методика инъекций
Техника и методика инъекций Нормы питания
Нормы питания Диуретические средства
Диуретические средства Жүйке жүйесінің қызметі
Жүйке жүйесінің қызметі Доврачебная помощь при острых аллергических заболеваниях
Доврачебная помощь при острых аллергических заболеваниях Пастереллез
Пастереллез Доброкачественная опухоль феохромоцитома
Доброкачественная опухоль феохромоцитома Стандартизация лекарственных средств в зависимости от формы випуска. (Лекция 5)
Стандартизация лекарственных средств в зависимости от формы випуска. (Лекция 5) Зубы и уход за ними (школьники младших классов)
Зубы и уход за ними (школьники младших классов) Ерік. Балада ерікті дамыту жолдары
Ерік. Балада ерікті дамыту жолдары Місцеве знеболювання
Місцеве знеболювання Мануальная медицина
Мануальная медицина Отдельные санитарно-эпидемиологические требования к перевозке скоропортящихся грузов
Отдельные санитарно-эпидемиологические требования к перевозке скоропортящихся грузов The problem of iodine deficiency: an epidemiological, clinical, social values. Solutions
The problem of iodine deficiency: an epidemiological, clinical, social values. Solutions Гестационный сахарный диабет
Гестационный сахарный диабет Атрезия пищевода и пилоростеноз
Атрезия пищевода и пилоростеноз Особо опасные инфекции (1)
Особо опасные инфекции (1) Жүрек кардиостимуляторы
Жүрек кардиостимуляторы Роль фельдшера в диагностике и лечении бронхиальной астмы у детей
Роль фельдшера в диагностике и лечении бронхиальной астмы у детей Инфекция мочевой системы у детей
Инфекция мочевой системы у детей Урбоэкология. Экологические и гигиенические проблемы жилища, ЛПУ. Лекция 16
Урбоэкология. Экологические и гигиенические проблемы жилища, ЛПУ. Лекция 16