- Дифференцировка Т-лимфоцитов

Содержание
- 3. Относительное содержание Т-лимфоцитов в крови составляет в среднем около 73% (55-85%) от общего числа лимфоцитов; абсолютное
- 4. Т-лимфоциты — активно рециркулирующие клетки; Основное место локализации Т-лимфоцитов в лимфоидных органах — тимусзависимые зоны (паракортикальные
- 5. αβТ-лимфоциты
- 6. СD8+ Т-лимфоциты (ЦТЛ) Имеют цитотоксическую направленность против изменённых клеток собственного организма (инфицированные вирусом клетки, опухолевые клетки-мишени);
- 7. CD4+ Т-лимфоциты (Th) Поставляют вспомогательные сигналы при активации В-лимфоцитов и макрофагов; Взаимодействие Т-хелперов с дендритными клетками
- 8. Регуляторные Т-клетки (Treg) (CD4+CD25+FoxP3+) Некоторые CD4+ Т-клетки, экспрессирующие внутриклеточный фактор FoxP3 и мембранные молекулы CD25 и
- 9. γδT-лимфоциты
- 10. γδT-лимфоциты Вариабельность γδTCR ограничена и спектр антигенов, распознаваемых γδТ-клетками, узок. γδТ-клетки распознают антиген независимо от молекул
- 11. Этапы дифференцировки Т-лимфоцитов
- 12. Этапы дифференцировки Т-лимфоцитов
- 13. ELP: CD34+SCA-l+CD117(c-Kit)+Flt-3+ CCR9+ CD4lo Sca-1 — антиген стволовых клеток (Stem cell antigen); с-Kit — лиганд фактора
- 14. DN1 Тимоциты интенсивно пролиферируют (до 10 делений), что приводит к 1000-кратному увеличению их числа; Длительность периода
- 15. DN2 Тимоциты перемещаются в сторону наружных слоев коры по градиенту CXCL12, секретируемого эпителиальными клетками наружной коры;
- 16. DN3 Запускается перестройка V-генов TCR; Для раннего развития Т-клеток необходима экспрессия генов Notch, Runx-1, GATA-3, Ikaros,
- 17. Роль Notch-рецепторов в развитии Т-клеток в тимусе
- 18. Реаранжировка V-генов TCR
- 20. Выбор пути αβ/γδ-дифференцировки Т-клеток
- 21. DP Тимоциты приобретают фенотип CD44-CD25- одновременно с экспрессией корецепторов CD4 и CD8 (точнее, экспрессией молекулы CD8
- 22. Селекция тимоцитов и формирование субпопуляций CD4+ и CD8 + клеток Положительная селекция – обеспечивает отбор только
- 23. Положительная селекция «по умолчанию»
- 24. Отрицательная селекция
- 25. Дифференцировка CD4 + и CD8 + тимоцитов Дифференцировка Т-клеток регулируется как внутренними (дифференцировочные факторы), так и
- 26. Дифференцировка CD4 + и CD8 + тимоцитов
- 27. Проверка специфичности корецептора при дифференцировке CD4+ и CD8+ Т-лимфоцитов (селекционная модель).
- 28. Созревшие тимоциты довольно долго (7—14 сут) не покидают тимус, пребывая преимущественно в наружных слоях мозгового слоя;
- 29. Регуляторные Т-клетки (Treg) Имеют мембранный фенотип CD4+CD25hiCTLA-4+GITR+PD-l+ Сильно экспрессируют α-цепь рецептора для IL-2 (CD25); На их
- 30. Регуляторные Т-клетки (Treg)
- 31. Экспрессия генов CD25 и FOXP3 как ключевые события дифференцировки Treg
- 33. Скачать презентацию

Относительное содержание Т-лимфоцитов в крови составляет в среднем около 73% (55-85%)
Относительное содержание Т-лимфоцитов в крови составляет в среднем около 73% (55-85%)

Т-лимфоциты — активно рециркулирующие клетки;
Основное место локализации Т-лимфоцитов в лимфоидных органах
Т-лимфоциты — активно рециркулирующие клетки;
Основное место локализации Т-лимфоцитов в лимфоидных органах
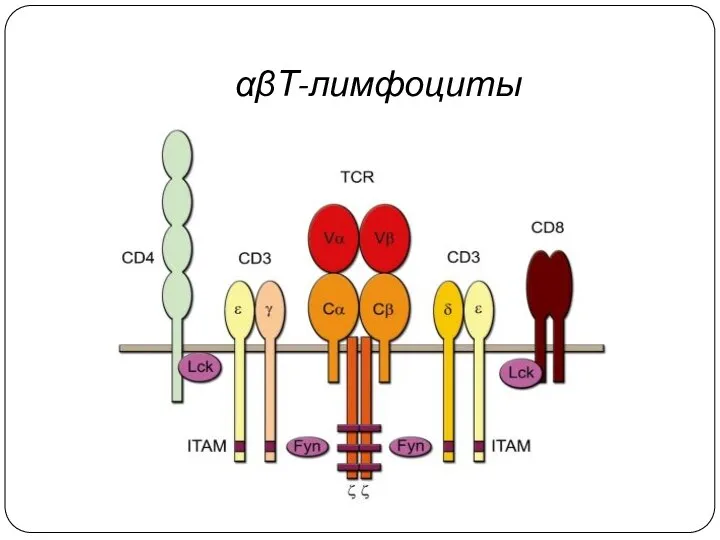
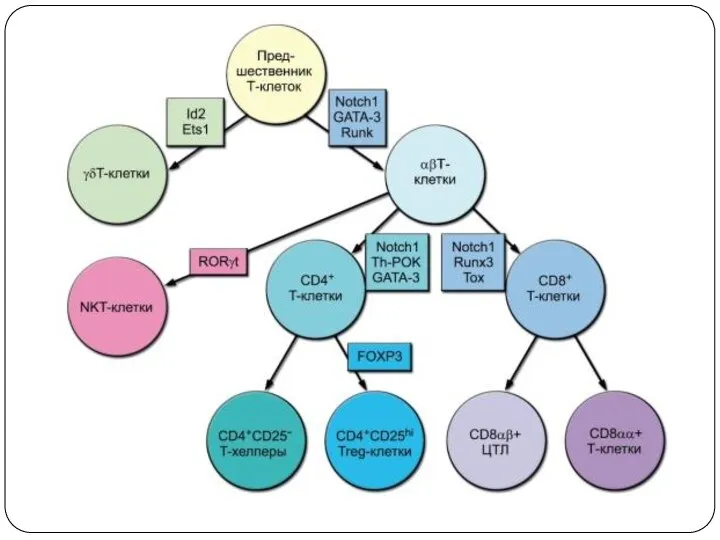
αβТ-лимфоциты
αβТ-лимфоциты

СD8+ Т-лимфоциты (ЦТЛ)
Имеют цитотоксическую направленность против изменённых клеток собственного организма (инфицированные
СD8+ Т-лимфоциты (ЦТЛ)
Имеют цитотоксическую направленность против изменённых клеток собственного организма (инфицированные

CD4+ Т-лимфоциты (Th)
Поставляют вспомогательные сигналы при активации В-лимфоцитов и макрофагов;
Взаимодействие Т-хелперов
CD4+ Т-лимфоциты (Th)
Поставляют вспомогательные сигналы при активации В-лимфоцитов и макрофагов;
Взаимодействие Т-хелперов

Регуляторные Т-клетки (Treg) (CD4+CD25+FoxP3+)
Некоторые CD4+ Т-клетки, экспрессирующие внутриклеточный фактор FoxP3 и
Регуляторные Т-клетки (Treg) (CD4+CD25+FoxP3+)
Некоторые CD4+ Т-клетки, экспрессирующие внутриклеточный фактор FoxP3 и
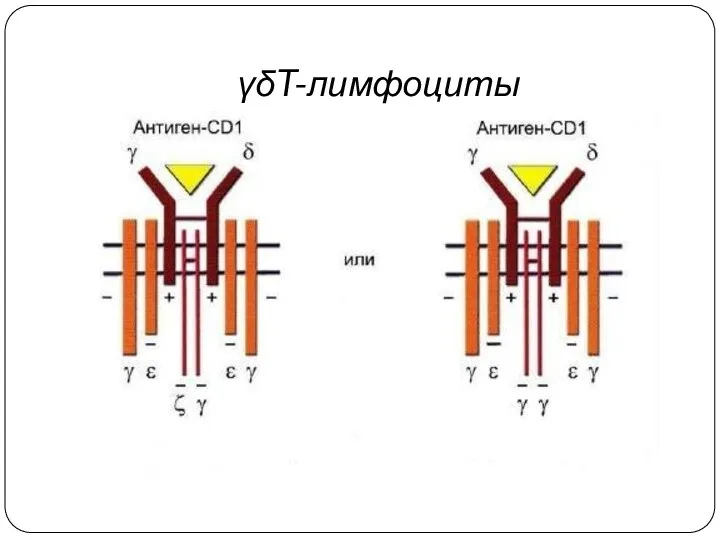
γδT-лимфоциты
γδT-лимфоциты

γδT-лимфоциты
Вариабельность γδTCR ограничена и спектр антигенов, распознаваемых γδТ-клетками, узок.
γδТ-клетки распознают антиген
γδT-лимфоциты
Вариабельность γδTCR ограничена и спектр антигенов, распознаваемых γδТ-клетками, узок.
γδТ-клетки распознают антиген
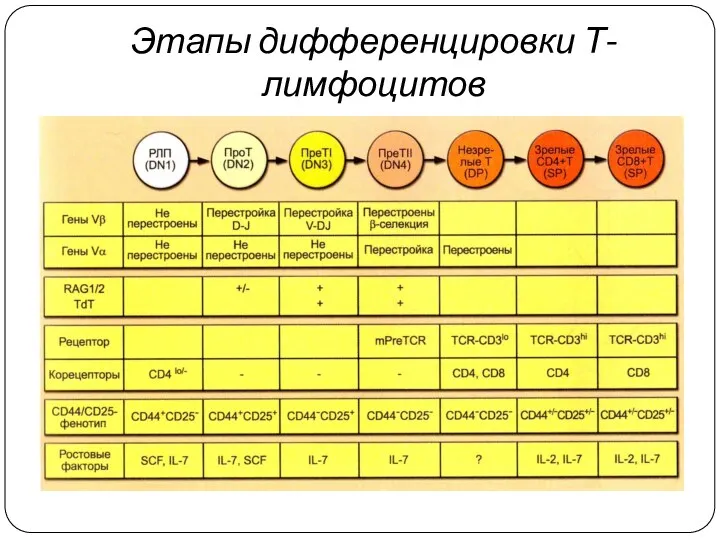
Этапы дифференцировки Т-лимфоцитов
Этапы дифференцировки Т-лимфоцитов
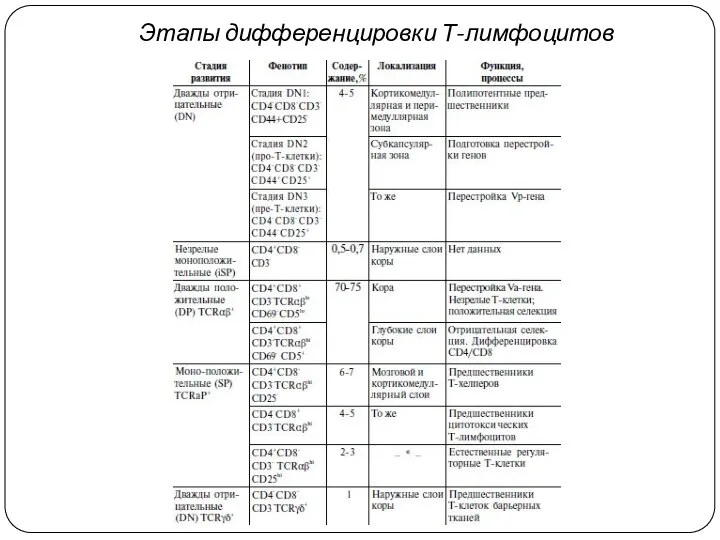
Этапы дифференцировки Т-лимфоцитов
Этапы дифференцировки Т-лимфоцитов
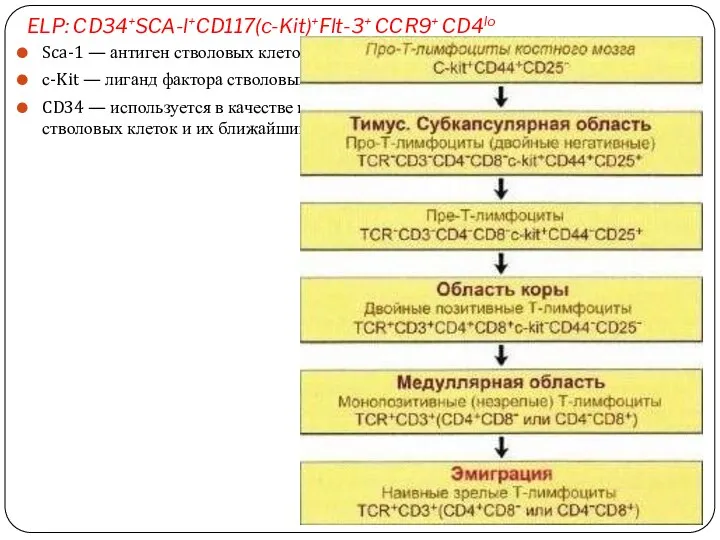
ELP: CD34+SCA-l+CD117(c-Kit)+Flt-3+ CCR9+ CD4lo
Sca-1 — антиген стволовых клеток (Stem cell antigen);
с-Kit
ELP: CD34+SCA-l+CD117(c-Kit)+Flt-3+ CCR9+ CD4lo
Sca-1 — антиген стволовых клеток (Stem cell antigen);
с-Kit

DN1
Тимоциты интенсивно пролиферируют (до 10 делений), что приводит к 1000-кратному увеличению
DN1
Тимоциты интенсивно пролиферируют (до 10 делений), что приводит к 1000-кратному увеличению

DN2
Тимоциты перемещаются в сторону наружных слоев коры по градиенту CXCL12, секретируемого
DN2
Тимоциты перемещаются в сторону наружных слоев коры по градиенту CXCL12, секретируемого

DN3
Запускается перестройка V-генов TCR;
Для раннего развития Т-клеток необходима экспрессия генов Notch,
DN3
Запускается перестройка V-генов TCR;
Для раннего развития Т-клеток необходима экспрессия генов Notch,
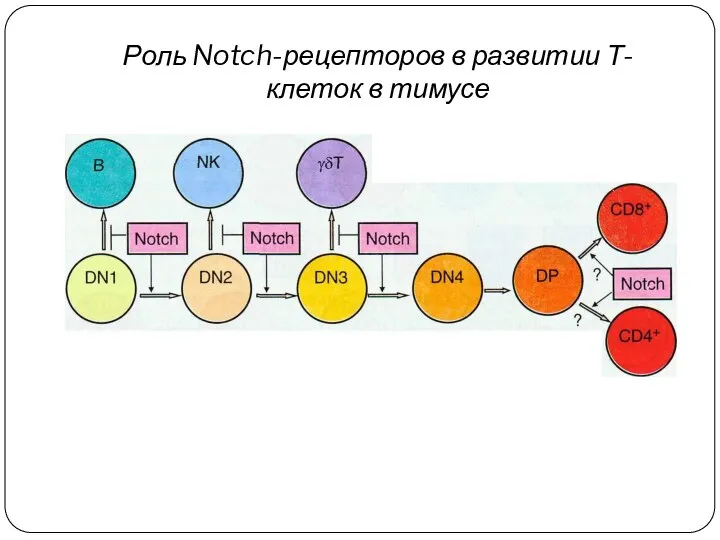
Роль Notch-рецепторов в развитии Т-клеток в тимусе
Роль Notch-рецепторов в развитии Т-клеток в тимусе
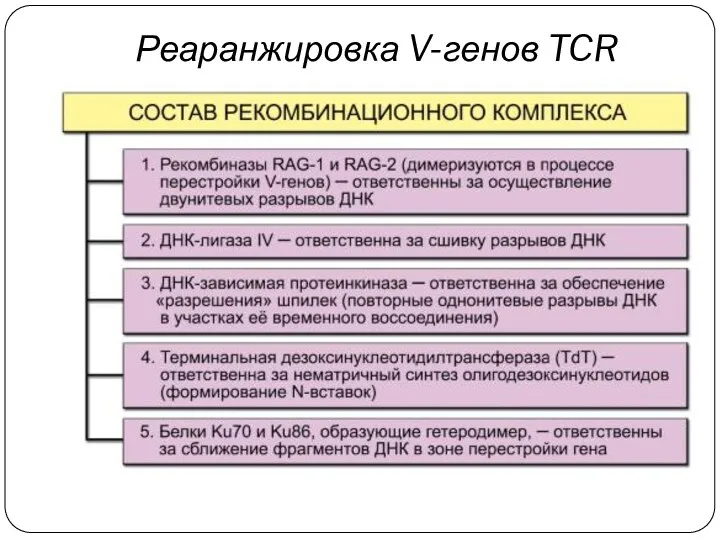
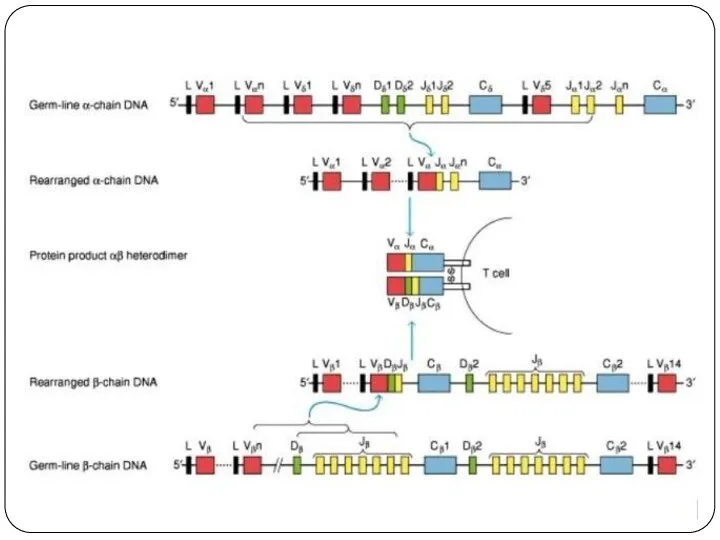
Реаранжировка V-генов TCR
Реаранжировка V-генов TCR
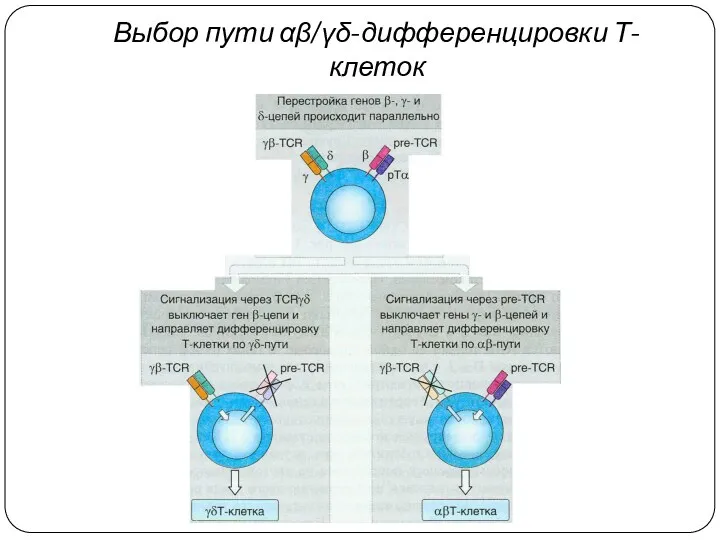
Выбор пути αβ/γδ-дифференцировки Т-клеток
Выбор пути αβ/γδ-дифференцировки Т-клеток

DP
Тимоциты приобретают фенотип CD44-CD25- одновременно с экспрессией корецепторов CD4 и CD8
DP
Тимоциты приобретают фенотип CD44-CD25- одновременно с экспрессией корецепторов CD4 и CD8

Селекция тимоцитов и формирование субпопуляций CD4+
и CD8 + клеток
Положительная селекция –
Селекция тимоцитов и формирование субпопуляций CD4+
и CD8 + клеток
Положительная селекция –
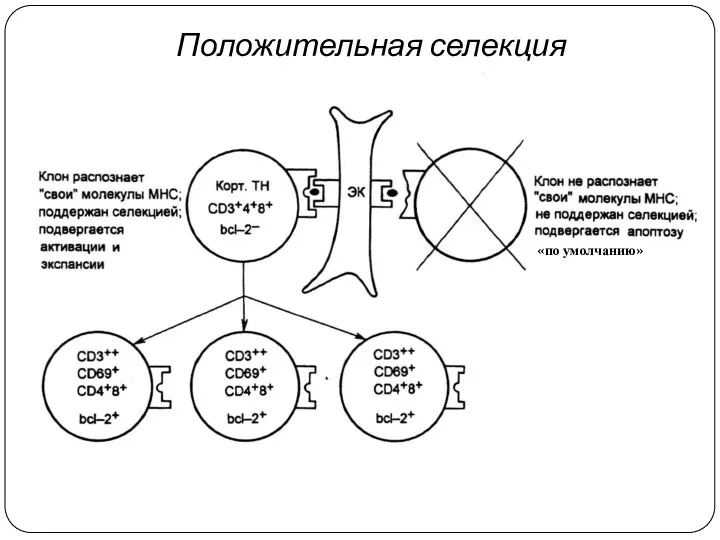
Положительная селекция
«по умолчанию»
Положительная селекция
«по умолчанию»
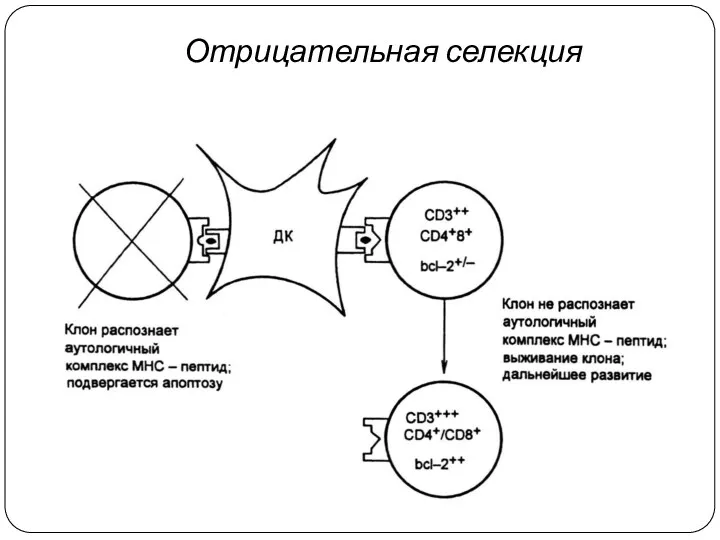
Отрицательная селекция
Отрицательная селекция

Дифференцировка CD4 + и CD8 + тимоцитов
Дифференцировка Т-клеток регулируется как внутренними
Дифференцировка CD4 + и CD8 + тимоцитов
Дифференцировка Т-клеток регулируется как внутренними
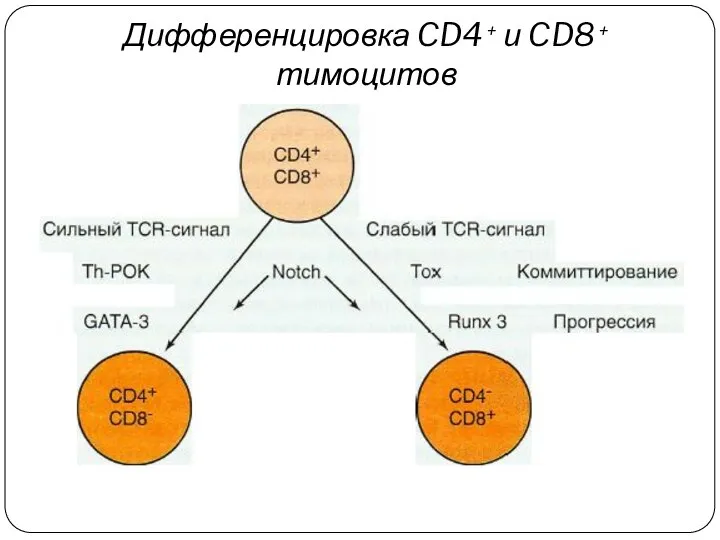
Дифференцировка CD4 + и CD8 + тимоцитов
Дифференцировка CD4 + и CD8 + тимоцитов
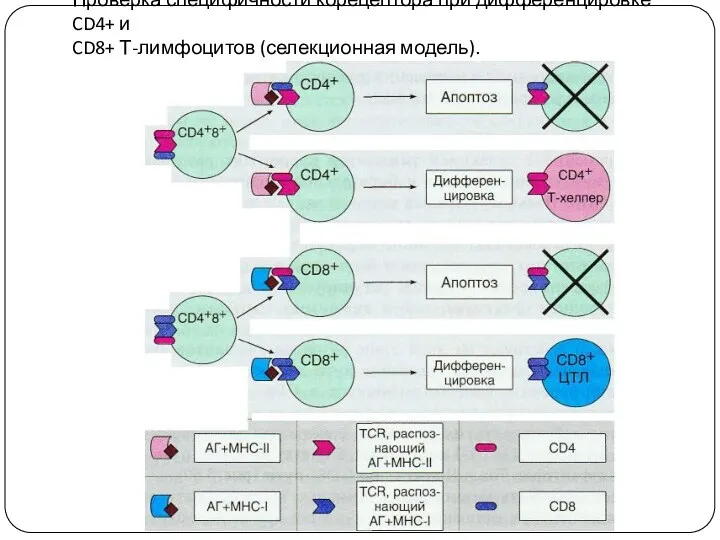
Проверка специфичности корецептора при дифференцировке CD4+ и
CD8+ Т-лимфоцитов (селекционная модель).
Проверка специфичности корецептора при дифференцировке CD4+ и
CD8+ Т-лимфоцитов (селекционная модель).

Созревшие тимоциты довольно долго (7—14 сут) не покидают тимус, пребывая преимущественно
Созревшие тимоциты довольно долго (7—14 сут) не покидают тимус, пребывая преимущественно

Регуляторные Т-клетки (Treg)
Имеют мембранный фенотип CD4+CD25hiCTLA-4+GITR+PD-l+
Сильно экспрессируют α-цепь рецептора для IL-2
Регуляторные Т-клетки (Treg)
Имеют мембранный фенотип CD4+CD25hiCTLA-4+GITR+PD-l+
Сильно экспрессируют α-цепь рецептора для IL-2
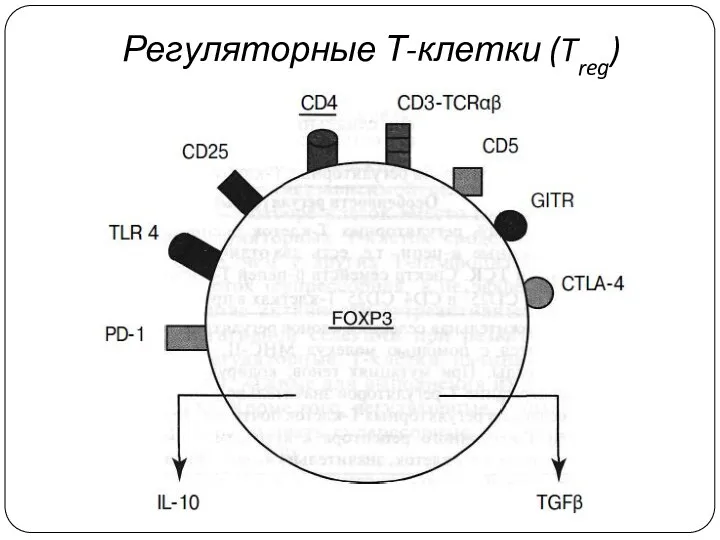
Регуляторные Т-клетки (Treg)
Регуляторные Т-клетки (Treg)
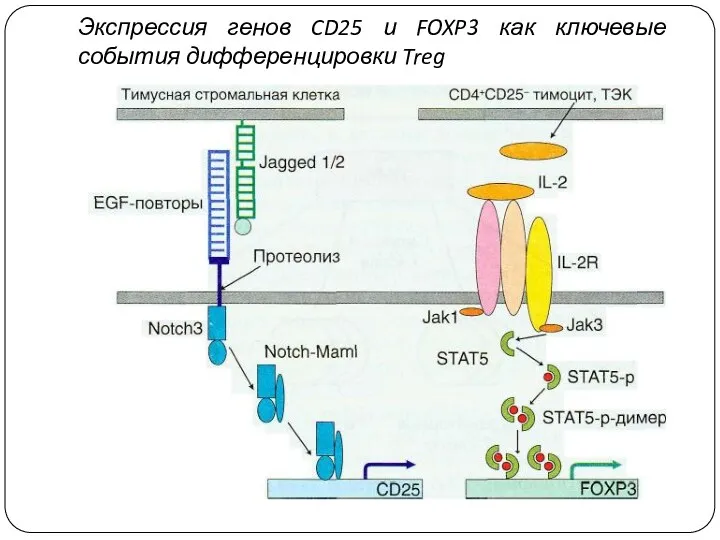
Экспрессия генов CD25 и FOXP3 как ключевые события дифференцировки Treg
Экспрессия генов CD25 и FOXP3 как ключевые события дифференцировки Treg
 Общие вопросы миологии. Мышцы головы и шеи. (Лекция 7)
Общие вопросы миологии. Мышцы головы и шеи. (Лекция 7) Грыжи головного мозга у детей
Грыжи головного мозга у детей Содержание глюкозы в крови
Содержание глюкозы в крови Жұтқыншақ ағзасының құрылымы мен тапографиясының жасқа байланысты ерекшеліктері және олардың жұтқыншақтағы патологиясының маңызы
Жұтқыншақ ағзасының құрылымы мен тапографиясының жасқа байланысты ерекшеліктері және олардың жұтқыншақтағы патологиясының маңызы Слуховые аппараты. Виды слуховых аппаратов
Слуховые аппараты. Виды слуховых аппаратов Гиперкортицизм. Синдромының асқынуы
Гиперкортицизм. Синдромының асқынуы Терапия. СД. Задача. Диагноз: гипертоническая болезнь 3 стадии
Терапия. СД. Задача. Диагноз: гипертоническая болезнь 3 стадии Основы микрохирургии
Основы микрохирургии Гнойные заболевания уха. Болезнь Меньера. Отосклероз. Нейросенсорная тугоухость
Гнойные заболевания уха. Болезнь Меньера. Отосклероз. Нейросенсорная тугоухость Сальмонеллёз: диагностика, лечение
Сальмонеллёз: диагностика, лечение Медицина без границ. Центр добровольного медицинского страхования
Медицина без границ. Центр добровольного медицинского страхования Методы обследования больных с бронхо-легочной патологией. Аускультация легких
Методы обследования больных с бронхо-легочной патологией. Аускультация легких Темір жетіспеушілік анемияның клиникалық көріністері
Темір жетіспеушілік анемияның клиникалық көріністері Torch-инфекции: клиническая диагностика и этиологическая верификация
Torch-инфекции: клиническая диагностика и этиологическая верификация Расщелины губы и неба
Расщелины губы и неба Эндодонтические материалы
Эндодонтические материалы Ароматерапия в жизни человека
Ароматерапия в жизни человека Патология шейки матки
Патология шейки матки Жасөспірімдік азық
Жасөспірімдік азық Болезни птиц. Классификация. Диагностические экспертизы
Болезни птиц. Классификация. Диагностические экспертизы Трансплантологиядағы жаңалықтар
Трансплантологиядағы жаңалықтар Цели и задачи микробиологии, вирусологии, иммунологии и их значение в практической деятельности врача
Цели и задачи микробиологии, вирусологии, иммунологии и их значение в практической деятельности врача Бас сүйегі мен Ми жарақаттары
Бас сүйегі мен Ми жарақаттары Мезиальный прикус. Этиология, патогенез, клиника, диагностика, профилактика и лечение
Мезиальный прикус. Этиология, патогенез, клиника, диагностика, профилактика и лечение Жедел бүйрек жеткіліксіздігі
Жедел бүйрек жеткіліксіздігі Вступ до спортивної морфології
Вступ до спортивної морфології Рентгенологические признаки заболеваний легких: как читать рентгенограммы и КТ органов грудной клетки
Рентгенологические признаки заболеваний легких: как читать рентгенограммы и КТ органов грудной клетки Особенности раннего послеоперационного периода
Особенности раннего послеоперационного периода