- ДНҚ репликациясы

Содержание
- 3. ДНК дупликациясы кезінде спиральды екі тізбекті тұрақты ұстап тұратын сутегілік байланыстар үзіледі. Кез келген тізбек басқа
- 4. Барлық екі тізбекті ДНК репликациясы жартылай консервативті болып келеді. Бірінші репликациядан кейін кез келген аналық молекуланың
- 5. Дуплексті ДНК қолданбалы репликация жобасы. Суреттің ортаңғы бөлігінде жартылай консервативті механизм көрсетілген. Соның ішінде – консервативті
- 6. ДНК репликациясы арнайы жерлерден басталады, оны репликация басталатын нүкте деп атаймыз. ДНК екі еселенгенше көшірілу процесі
- 7. ДНК синтезі ori нүктесінен екі бағытта жүреді. ДНК репликация синтезінің ерекшелігі 5'-ұшы жаңа синтезделген тізбектердің аяқталмаған
- 8. Эукариот жасушаларының геномына хромосомада 30-300 м.ж.н арақашықтығы бойынша таралған көптеген репликация басталатын нүктелер тән. Эукариот жасушаларының
- 9. Хромосомалық эукариот ДНК репликациясы. Репликация екі бағытта және әр түрлі нүктелерден бағытталады. Репликация басталатын нүкте (ori)
- 10. Электронды микрофотография арқылы ДНК Drosophila-да репликация жүріп жатқанын көре аламыз.
- 11. Геномдағы репликация жылдамдығы басты жағдайда инициация жиілігіне байланысты. Е. coli да әр бір айыршаның көшірілуі 1500
- 12. Кей жағдайларда репликация басталатын орын сондай бір нуклеотидтер кезектілігіне йе болады да кейін дуплекс ерекше конфирурацияға
- 13. ДНК тізбектері 5'-дезоксинуклеотидті дезоксирибонуклеозидтрифосфат бірлігінің 3'-гидроксильді ұшымен байланысуымен синтезеделеді. Матрицалық тізбек бойымен 5'–>3' бағытта олар синтезделеді,
- 14. ДНК тізбектері әрбір кезекті нуклеотид ұшындағы 3'-ОН-топқа дезоксинуклеотидтердің байланысуымен ұзарады. Жаңадан дезоксинуклеотид бірліктеріне 5'-дезоксинуклеозидтрифосфаттар донор болып
- 15. ДНК репликациясы. Праймер тізбегінің өсуі матрицалық тізбекке комлементарлы тізбектің көшірілуімен жүзеге асады. Жаңа дезоксинуклеотидтер 3'-ұшына жалғанса
- 16. ДНК екі тізбегінде де репликативті айырша синтезі 5'–>3' бағытта жүреді. Бір тізбек - бастаушы – толық
- 17. Оказаки фрагменттерін пайда болуы барысында репликацияның инициация механизмдері бірдей, бірақ кейбір ерекшеліктері бар. Екі жағдайда қысқа
- 18. ДНК репликациясының инициациясы Начало репликации Локальное расплетание спирали ДНК Синтез РНК-затравки Начало синтеза лидирующей цепи Синтез
- 19. Синтез барсында кейде матрицалық тізбек ішінде нуклеотидтердің қате байланысуы болады. Нуклеотидтердің қате қосылуынан тізбектің өсуі тоқтатылады.
- 20. ДНК-полимераза- праймер - қажетті, дезоксинуклеотидтетердің байланысуын ДНК матрицасымен детерминирленетін фермент. Оказаки фрагменттері ДНК тізбегін көшіру барыснда
- 21. Сирек кездесетін жұптар, жай және күрделілерді салыстырамыз
- 22. Қате байланысу кездейсоқтан пайда болады. ДНК тізбегі қате нуклеотидті қосып алады. Дұрыс байланысудың бұзылуы нәтижесінде праймер
- 23. ДНК-полимераза барлық прокариот және эукариот жасушаларында кездеседі. Көптеген жануарлар және бактерия вирустары вирус спецификалық ДНК полимераза
- 25. I (Pol I) E. coli ДНК полимеразасы толық зерттелген. Жалғыз полипептид мультиқызметтілігімен сипатталады. Pol I ДНК-полимераза
- 26. Нуклеотидердің полимеризациясынан басқа Pol I екі реакцияны катализдейді, биологиялық ролі өте маңызды. Солардың бірінде ДНК тізбегінің
- 27. Егер in vitro да трипсинмен Pol I байланыстырса, полипептидті тізбек үлкен және кіші бөлшектерге бөлінеді. Үлкен,
- 28. Pol I екі тізбекті ДНК ның бір тізбегінің 3'-ұшын ұзарта алады және 5'-ұшындағы нуклеотидтерді жоя алады
- 29. Pol III-холофермент – маңызды фермент, E. coli. хромосомалық ДНК репликациясына жауапты. Кез келген жасушада тек кана
- 30. ДНК-полимераза 3'–>5'- и 5'–>3'-экзонуклеазды реакцияларды катализдейді. 3'–>5'-экзонуклеаза бір тізбекті ДНК ны 3'-ОН-ұшынан ыдыратады, ал 5'–>3'-экзонуклеаза дуплексті
- 31. Эукариот жасушаларында көптеген ДНК-полимеразалар идентифицирленген. Прокариоттардың ферменттеріне қарағанда жақсы зерттелген. Сүт қоректілердің жасушаларынан төрт ДНК-полимераза алыған:
- 32. Ядролық хромосоманың репликациясына α ДНҚ полимеразамен бірге δ. ДНҚ полимераза қатысады. α ДНҚ полимераза–барлық эукариот жасушаларына
- 33. Тағы да бір полимераза ол- εДНК полимераза, δ ДНҚ полимеразаның кейбір жағдайда орынын басады, мысалы, ДНҚ
- 34. ДНК-полимераза хромосомалық ДНК репликациясына қатысады. Полимеразды белсенділігі үлкен полипептидтермен байланысты, бірақ муль-тисуббірлікті белок ретінде қызмет атқарады.
- 35. ДНК-лигаза ДНК тізбектерін байланыстырушы ретінде репликация, репарация және рекомбинацияға қатысады. Барлық белгілі лигазалар бұзылған ДНК орындарында
- 36. Барлық ДНК лигазалар дуплекті ДНК қарама қарсы ұштарында тұрған нуклеотидтердің 5'-фосфорильді және 3'-гидроксильді топтарын байланыстырады. Жаңа
- 37. Е. coli ДНК-лигаза, T4 және Т7 – жалғыз полипептидті тізбектер, ал сүт қоректілердің ДНК лигазаларының екі
- 38. Никотинамидадениндинуклеотид (NAD).
- 39. ДНК-лигазы (Enz) әсер ету механизмі. Қысқартылуы: Lys - лизин, лигазы құрамына кіреді, Rib - рибоза, Ad
- 40. Комплементарлы тізбектер көшірмелерін алу үшін екі тізбекті ДНК аздап шиыршықтануы қажет. Шиыршықтану немесе тарқатылу репликативті айырдың
- 41. ДНК дуплекстердің тарқатылуы екі белок қатысуымен жеңіл өтеді:геликазалар және белок. Олар біртізбекті ДНК байланыстырады. ДНК-геликаза Белки,
- 42. Егер де хроматин құрамындағы ДНК тарқатқымыз келсе, ДНК және гистондарды бұзу қажет. Кейін репликация аяқталған соң,
- 43. Шиыршықталудың шегіне қарай сегменттердің жоғары спиральдығы репликацияланатын айырша алдында тұрғанда біртіндеп төмендейді. Кейін айыршаны сақина ұшына
- 44. Екі спиральды ДНК айналуы механикалық өрісті азайтады. Здесь должно происходить быстрое вращение спирали ДНК Матрица для
- 45. Механикалық және топологиялық өрісті репликативті айыр алдында сақиналы дуплексте алып тастау қажет. Репликацияланбаған жоғары ДНК бөлігінде
- 46. ДНК-топоизомеразалар жоғары спиральдылықты және жоғары спираль типтерін өзгертеді. Олар репликативті айырдағы үздіксіз қозғалысты әкелетін шарнирлардың пайда
- 47. Әр түрлі ағзаларда топоизомеразалардың екі басты типтері идентифицирленген. I типті Топоизомеразалар жоғары айырларды ДНК да бірлікке
- 48. I типтегі топоизомераза арқылы катализденетін реакциялар ДНК-топоизомераза разрезает одну из цепей и коваяентно связывается с фосфатной
- 49. II типті топоизомеразалар екі сақиналы дуплекстерді катализдейді.
- 50. Топоизомераза II, гиразой тек қана бактерияларда табылған. Олар сақиналы ДНК теріс жоғары айырлардың индукциясын туғызады. Бұл
- 51. В бактерияларда II типтегі топоизомераза табылған, ол ДНК-гираза деп аталады А. гиразаның әсерімен релаксацияланған сақиналы ДНҚ
- 52. Жартылай консервативті Е.соli. ДНК репликация механизмі.
- 53. Артта қалған тізбектің инициация синтезі
- 55. Скачать презентацию

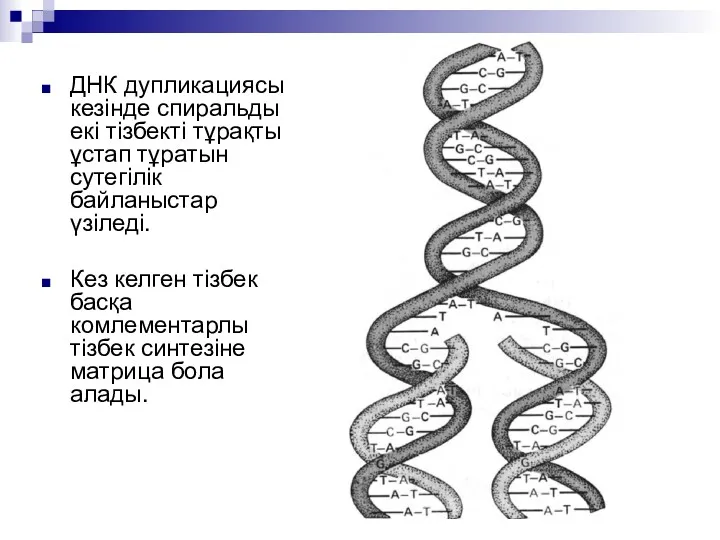
ДНК дупликациясы кезінде спиральды екі тізбекті тұрақты ұстап тұратын сутегілік байланыстар
ДНК дупликациясы кезінде спиральды екі тізбекті тұрақты ұстап тұратын сутегілік байланыстар

Барлық екі тізбекті ДНК репликациясы жартылай консервативті болып келеді. Бірінші репликациядан
Барлық екі тізбекті ДНК репликациясы жартылай консервативті болып келеді. Бірінші репликациядан
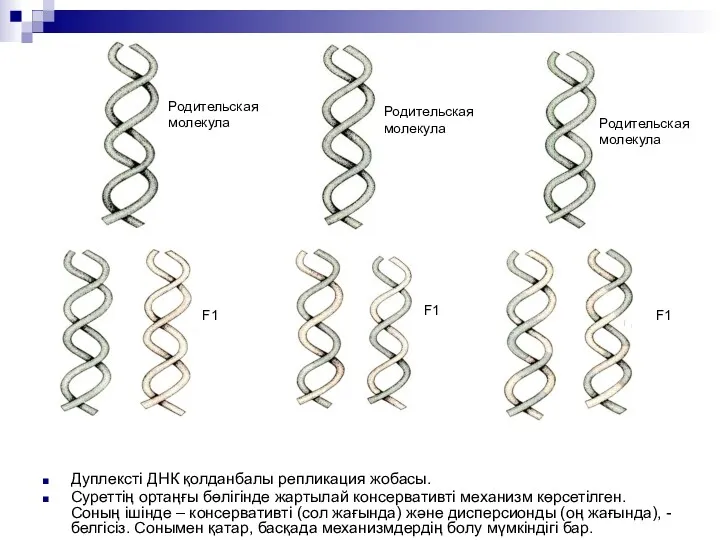
Дуплексті ДНК қолданбалы репликация жобасы.
Суреттің ортаңғы бөлігінде жартылай консервативті механизм
Дуплексті ДНК қолданбалы репликация жобасы.
Суреттің ортаңғы бөлігінде жартылай консервативті механизм

ДНК репликациясы арнайы жерлерден басталады, оны репликация басталатын нүкте деп атаймыз.
ДНК
ДНК репликациясы арнайы жерлерден басталады, оны репликация басталатын нүкте деп атаймыз.
ДНК
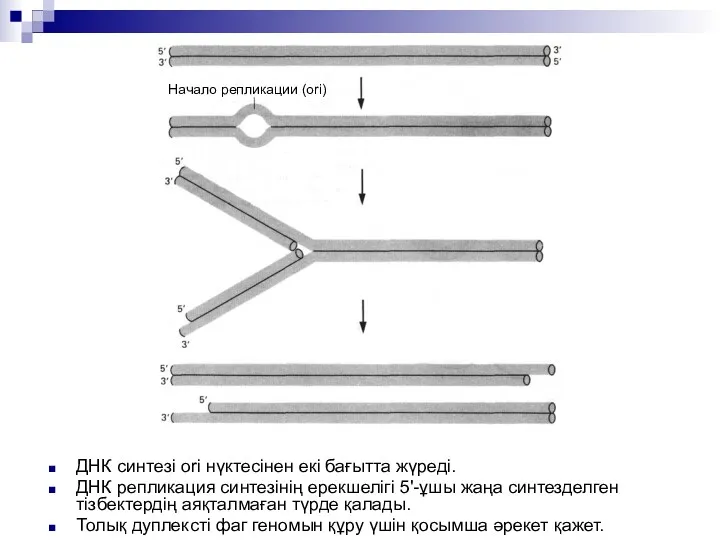
ДНК синтезі ori нүктесінен екі бағытта жүреді.
ДНК репликация синтезінің ерекшелігі
ДНК синтезі ori нүктесінен екі бағытта жүреді.
ДНК репликация синтезінің ерекшелігі

Эукариот жасушаларының геномына хромосомада 30-300 м.ж.н арақашықтығы бойынша таралған көптеген репликация
Эукариот жасушаларының геномына хромосомада 30-300 м.ж.н арақашықтығы бойынша таралған көптеген репликация
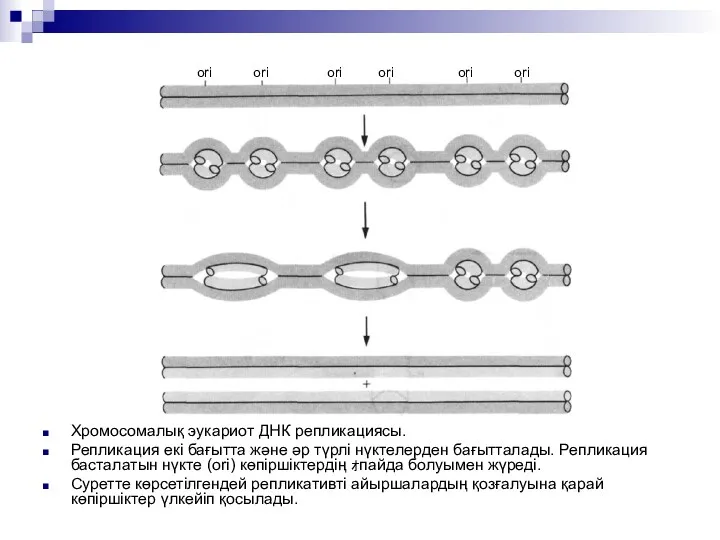
Хромосомалық эукариот ДНК репликациясы.
Репликация екі бағытта және әр түрлі нүктелерден
Хромосомалық эукариот ДНК репликациясы.
Репликация екі бағытта және әр түрлі нүктелерден

Электронды микрофотография арқылы ДНК Drosophila-да репликация жүріп жатқанын көре аламыз.
Электронды микрофотография арқылы ДНК Drosophila-да репликация жүріп жатқанын көре аламыз.

Геномдағы репликация жылдамдығы басты жағдайда инициация жиілігіне байланысты.
Е. coli да әр
Геномдағы репликация жылдамдығы басты жағдайда инициация жиілігіне байланысты.
Е. coli да әр

Кей жағдайларда репликация басталатын орын сондай бір нуклеотидтер кезектілігіне йе болады
Кей жағдайларда репликация басталатын орын сондай бір нуклеотидтер кезектілігіне йе болады

ДНК тізбектері 5'-дезоксинуклеотидті дезоксирибонуклеозидтрифосфат бірлігінің 3'-гидроксильді ұшымен байланысуымен синтезеделеді.
Матрицалық тізбек
ДНК тізбектері 5'-дезоксинуклеотидті дезоксирибонуклеозидтрифосфат бірлігінің 3'-гидроксильді ұшымен байланысуымен синтезеделеді.
Матрицалық тізбек
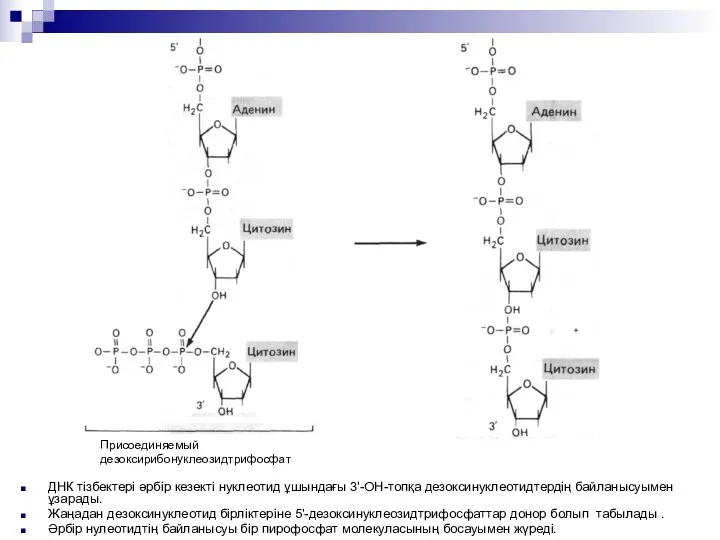
ДНК тізбектері әрбір кезекті нуклеотид ұшындағы 3'-ОН-топқа дезоксинуклеотидтердің байланысуымен ұзарады.
Жаңадан дезоксинуклеотид
ДНК тізбектері әрбір кезекті нуклеотид ұшындағы 3'-ОН-топқа дезоксинуклеотидтердің байланысуымен ұзарады.
Жаңадан дезоксинуклеотид
ДНК репликациясы. Праймер тізбегінің өсуі матрицалық тізбекке комлементарлы тізбектің көшірілуімен жүзеге
ДНК репликациясы. Праймер тізбегінің өсуі матрицалық тізбекке комлементарлы тізбектің көшірілуімен жүзеге
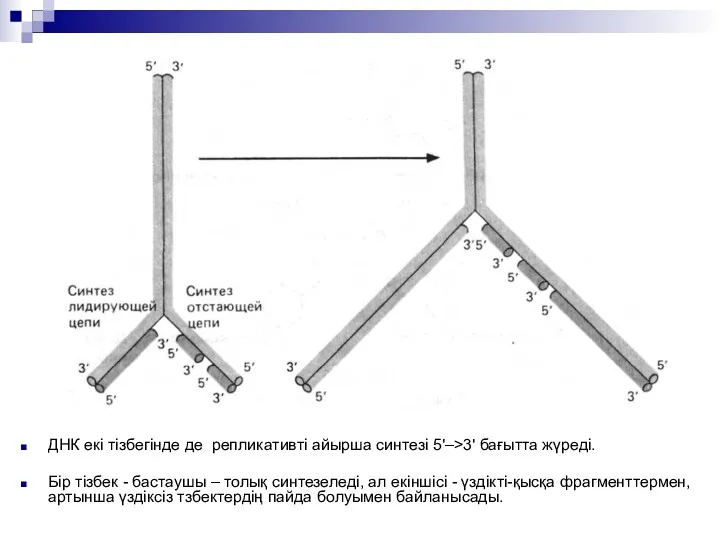
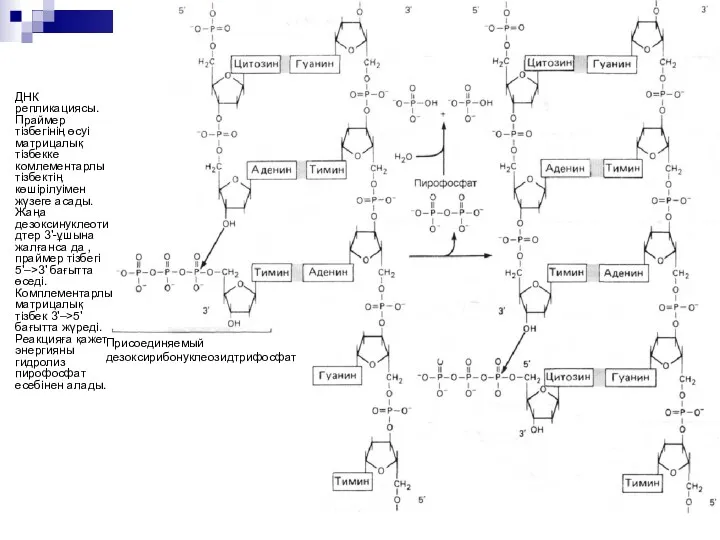
ДНК екі тізбегінде де репликативті айырша синтезі 5'–>3' бағытта жүреді.
Бір
ДНК екі тізбегінде де репликативті айырша синтезі 5'–>3' бағытта жүреді.
Бір

Оказаки фрагменттерін пайда болуы барысында репликацияның инициация механизмдері бірдей, бірақ кейбір
Оказаки фрагменттерін пайда болуы барысында репликацияның инициация механизмдері бірдей, бірақ кейбір
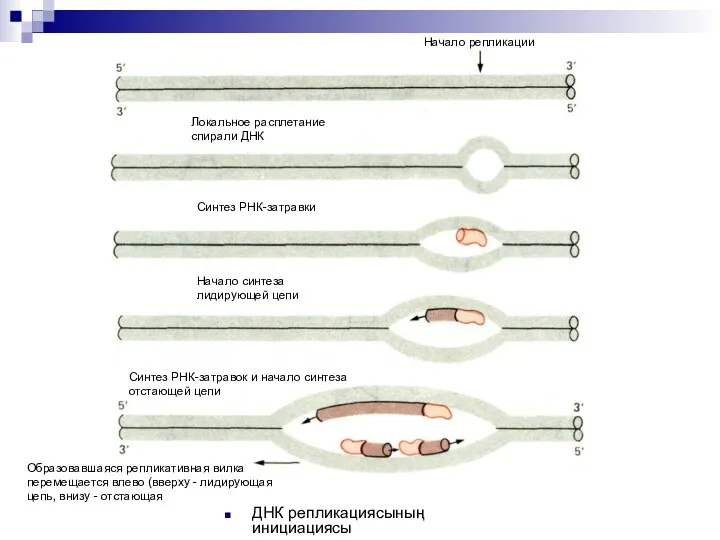
ДНК репликациясының инициациясы
Начало репликации
Локальное расплетание
спирали ДНК
Синтез РНК-затравки
Начало синтеза
лидирующей цепи
Синтез РНК-затравок и
ДНК репликациясының инициациясы
Начало репликации
Локальное расплетание
спирали ДНК
Синтез РНК-затравки
Начало синтеза
лидирующей цепи
Синтез РНК-затравок и

Синтез барсында кейде матрицалық тізбек ішінде нуклеотидтердің қате байланысуы болады.
Нуклеотидтердің
Синтез барсында кейде матрицалық тізбек ішінде нуклеотидтердің қате байланысуы болады.
Нуклеотидтердің

ДНК-полимераза- праймер - қажетті, дезоксинуклеотидтетердің байланысуын ДНК матрицасымен детерминирленетін фермент.
Оказаки фрагменттері
ДНК-полимераза- праймер - қажетті, дезоксинуклеотидтетердің байланысуын ДНК матрицасымен детерминирленетін фермент.
Оказаки фрагменттері
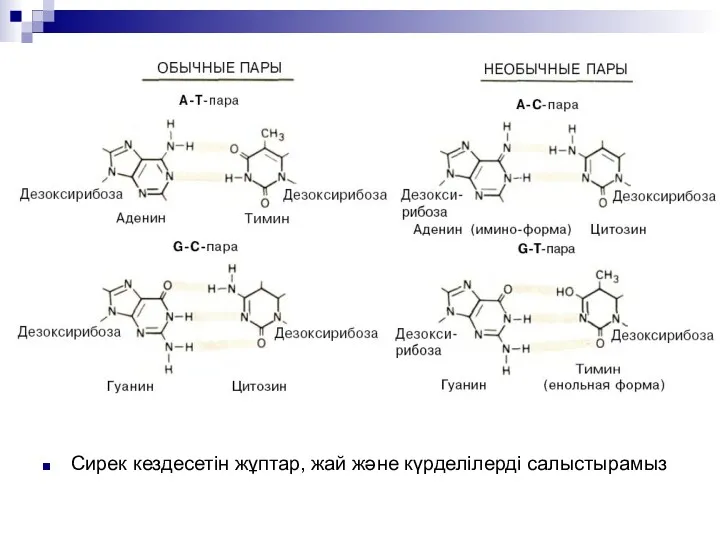
Сирек кездесетін жұптар, жай және күрделілерді салыстырамыз
Сирек кездесетін жұптар, жай және күрделілерді салыстырамыз
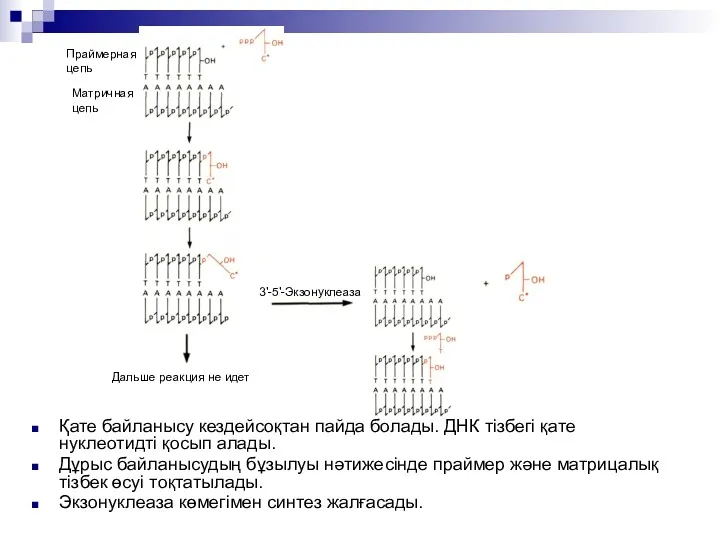
Қате байланысу кездейсоқтан пайда болады. ДНК тізбегі қате нуклеотидті қосып алады.
Қате байланысу кездейсоқтан пайда болады. ДНК тізбегі қате нуклеотидті қосып алады.

ДНК-полимераза барлық прокариот және эукариот жасушаларында кездеседі.
Көптеген жануарлар және бактерия
ДНК-полимераза барлық прокариот және эукариот жасушаларында кездеседі.
Көптеген жануарлар және бактерия
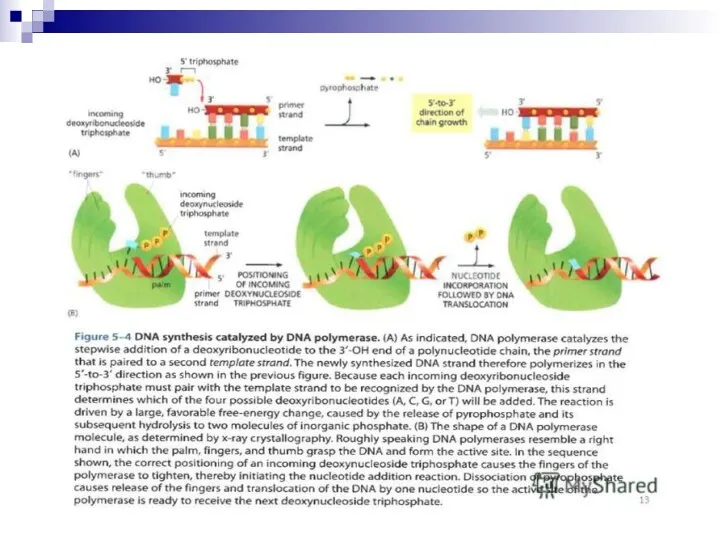

I (Pol I) E. coli ДНК полимеразасы толық зерттелген. Жалғыз полипептид
I (Pol I) E. coli ДНК полимеразасы толық зерттелген. Жалғыз полипептид

Нуклеотидердің полимеризациясынан басқа Pol I екі реакцияны катализдейді, биологиялық ролі өте
Нуклеотидердің полимеризациясынан басқа Pol I екі реакцияны катализдейді, биологиялық ролі өте

Егер in vitro да трипсинмен Pol I байланыстырса, полипептидті тізбек үлкен
Егер in vitro да трипсинмен Pol I байланыстырса, полипептидті тізбек үлкен

Pol I екі тізбекті ДНК ның бір тізбегінің 3'-ұшын ұзарта алады
Pol I екі тізбекті ДНК ның бір тізбегінің 3'-ұшын ұзарта алады

Pol III-холофермент – маңызды фермент, E. coli. хромосомалық ДНК репликациясына жауапты.
Pol III-холофермент – маңызды фермент, E. coli. хромосомалық ДНК репликациясына жауапты.
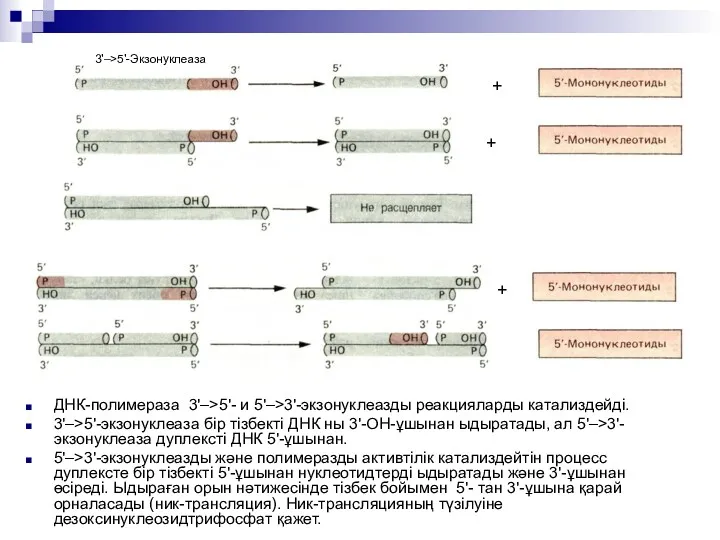
ДНК-полимераза 3'–>5'- и 5'–>3'-экзонуклеазды реакцияларды катализдейді.
3'–>5'-экзонуклеаза бір тізбекті ДНК ны
ДНК-полимераза 3'–>5'- и 5'–>3'-экзонуклеазды реакцияларды катализдейді.
3'–>5'-экзонуклеаза бір тізбекті ДНК ны

Эукариот жасушаларында көптеген ДНК-полимеразалар идентифицирленген. Прокариоттардың ферменттеріне қарағанда жақсы зерттелген.
Сүт қоректілердің
Эукариот жасушаларында көптеген ДНК-полимеразалар идентифицирленген. Прокариоттардың ферменттеріне қарағанда жақсы зерттелген.
Сүт қоректілердің

Ядролық хромосоманың репликациясына α ДНҚ полимеразамен бірге δ. ДНҚ полимераза қатысады.
Ядролық хромосоманың репликациясына α ДНҚ полимеразамен бірге δ. ДНҚ полимераза қатысады.

Тағы да бір полимераза ол- εДНК полимераза, δ ДНҚ полимеразаның кейбір
Тағы да бір полимераза ол- εДНК полимераза, δ ДНҚ полимеразаның кейбір

ДНК-полимераза хромосомалық ДНК репликациясына қатысады. Полимеразды белсенділігі үлкен полипептидтермен байланысты, бірақ
ДНК-полимераза хромосомалық ДНК репликациясына қатысады. Полимеразды белсенділігі үлкен полипептидтермен байланысты, бірақ

ДНК-лигаза ДНК тізбектерін байланыстырушы ретінде репликация, репарация және рекомбинацияға қатысады.
Барлық
ДНК-лигаза ДНК тізбектерін байланыстырушы ретінде репликация, репарация және рекомбинацияға қатысады.
Барлық

Барлық ДНК лигазалар дуплекті ДНК қарама қарсы ұштарында тұрған нуклеотидтердің 5'-фосфорильді
Барлық ДНК лигазалар дуплекті ДНК қарама қарсы ұштарында тұрған нуклеотидтердің 5'-фосфорильді

Е. coli ДНК-лигаза, T4 және Т7 – жалғыз полипептидті тізбектер, ал
Е. coli ДНК-лигаза, T4 және Т7 – жалғыз полипептидті тізбектер, ал
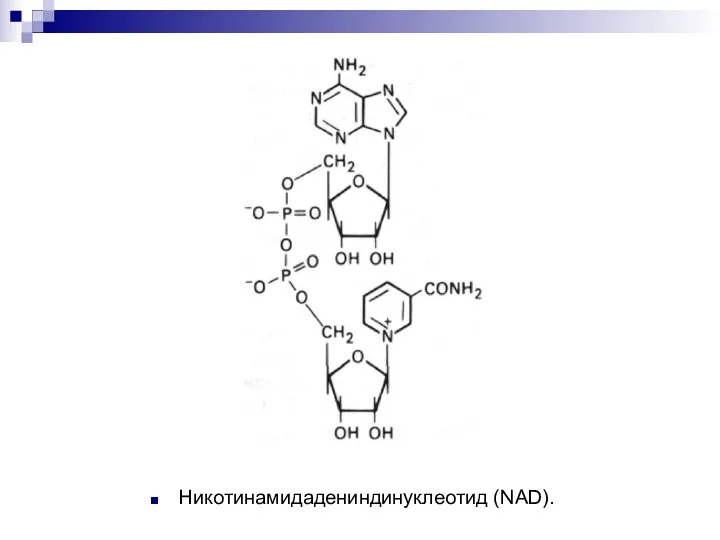
Никотинамидадениндинуклеотид (NAD).
Никотинамидадениндинуклеотид (NAD).
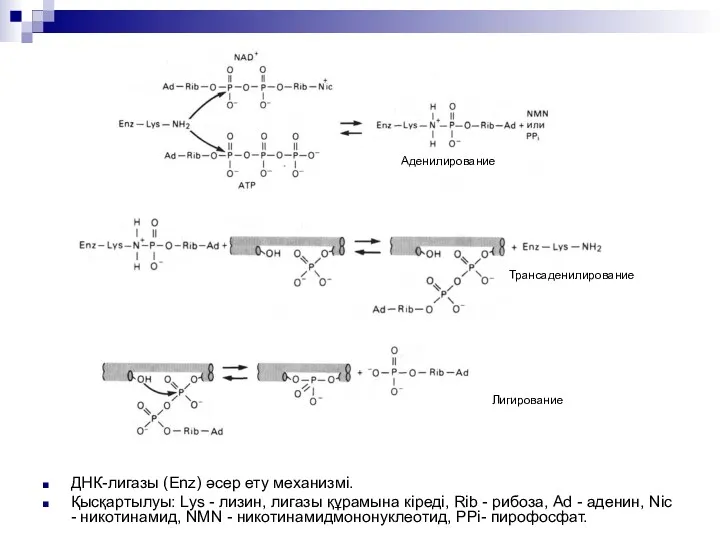
ДНК-лигазы (Enz) әсер ету механизмі.
Қысқартылуы: Lys - лизин, лигазы құрамына кіреді,
ДНК-лигазы (Enz) әсер ету механизмі.
Қысқартылуы: Lys - лизин, лигазы құрамына кіреді,

Комплементарлы тізбектер көшірмелерін алу үшін екі тізбекті ДНК аздап шиыршықтануы қажет.
Комплементарлы тізбектер көшірмелерін алу үшін екі тізбекті ДНК аздап шиыршықтануы қажет.
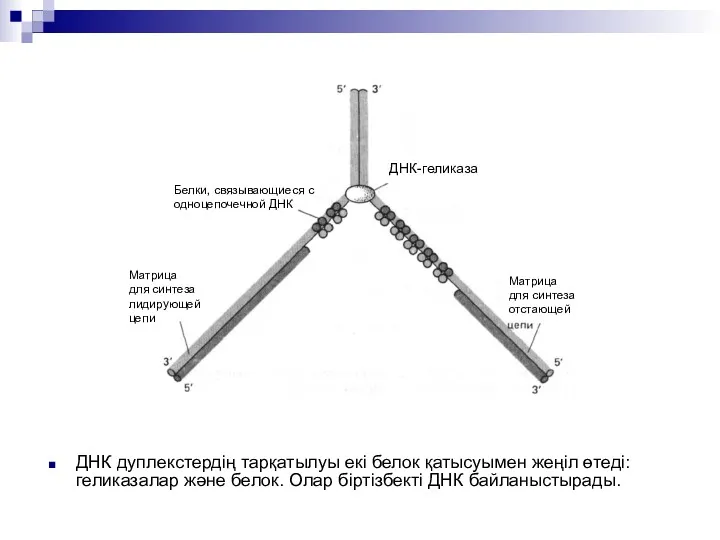
ДНК дуплекстердің тарқатылуы екі белок қатысуымен жеңіл өтеді:геликазалар және белок. Олар
ДНК дуплекстердің тарқатылуы екі белок қатысуымен жеңіл өтеді:геликазалар және белок. Олар

Егер де хроматин құрамындағы ДНК тарқатқымыз келсе, ДНК және гистондарды бұзу
Егер де хроматин құрамындағы ДНК тарқатқымыз келсе, ДНК және гистондарды бұзу

Шиыршықталудың шегіне қарай сегменттердің жоғары спиральдығы репликацияланатын айырша алдында тұрғанда біртіндеп
Шиыршықталудың шегіне қарай сегменттердің жоғары спиральдығы репликацияланатын айырша алдында тұрғанда біртіндеп
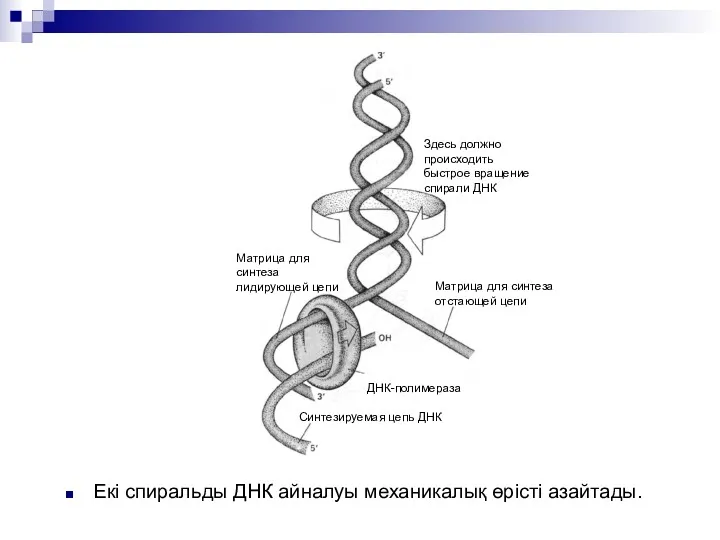
Екі спиральды ДНК айналуы механикалық өрісті азайтады.
Здесь должно
происходить
быстрое вращение
спирали ДНК
Матрица для
Екі спиральды ДНК айналуы механикалық өрісті азайтады.
Здесь должно
происходить
быстрое вращение
спирали ДНК
Матрица для

Механикалық және топологиялық өрісті репликативті айыр алдында сақиналы дуплексте алып тастау
Механикалық және топологиялық өрісті репликативті айыр алдында сақиналы дуплексте алып тастау

ДНК-топоизомеразалар жоғары спиральдылықты және жоғары спираль типтерін өзгертеді.
Олар репликативті айырдағы
ДНК-топоизомеразалар жоғары спиральдылықты және жоғары спираль типтерін өзгертеді.
Олар репликативті айырдағы

Әр түрлі ағзаларда топоизомеразалардың екі басты типтері идентифицирленген.
I типті
Әр түрлі ағзаларда топоизомеразалардың екі басты типтері идентифицирленген.
I типті
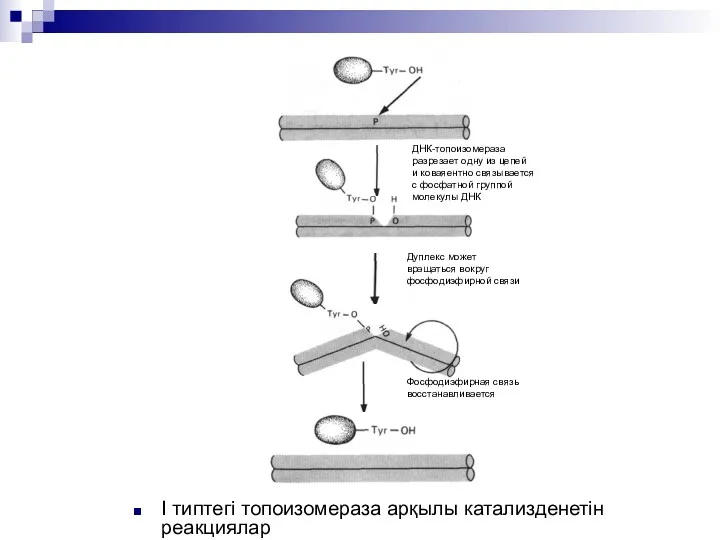
I типтегі топоизомераза арқылы катализденетін реакциялар
ДНК-топоизомераза
разрезает одну из цепей
и коваяентно связывается
с
I типтегі топоизомераза арқылы катализденетін реакциялар
ДНК-топоизомераза
разрезает одну из цепей
и коваяентно связывается
с
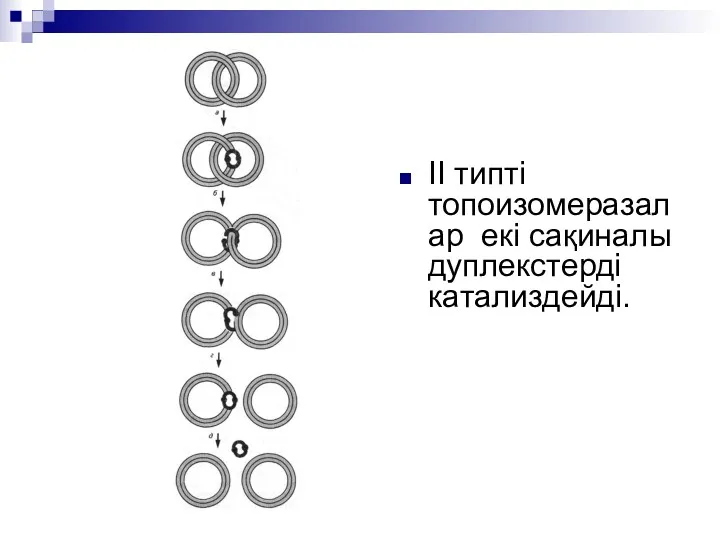
II типті топоизомеразалар екі сақиналы дуплекстерді катализдейді.
II типті топоизомеразалар екі сақиналы дуплекстерді катализдейді.

Топоизомераза II, гиразой тек қана бактерияларда табылған. Олар сақиналы ДНК теріс
Топоизомераза II, гиразой тек қана бактерияларда табылған. Олар сақиналы ДНК теріс
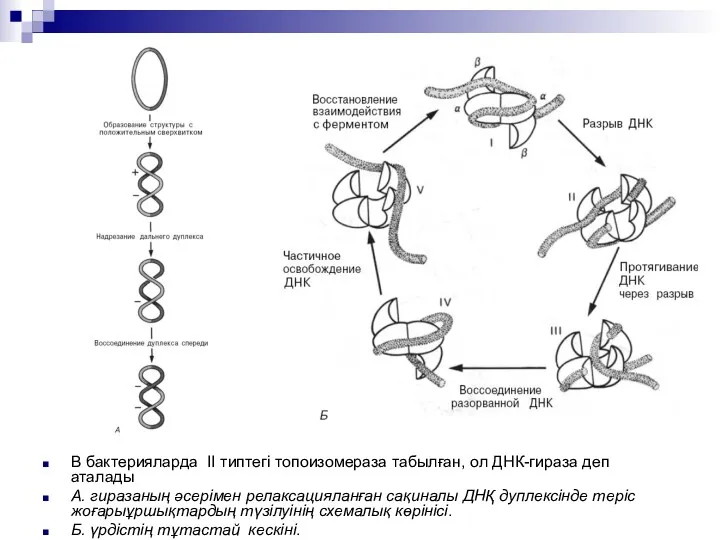
В бактерияларда II типтегі топоизомераза табылған, ол ДНК-гираза деп аталады
А.
В бактерияларда II типтегі топоизомераза табылған, ол ДНК-гираза деп аталады
А.
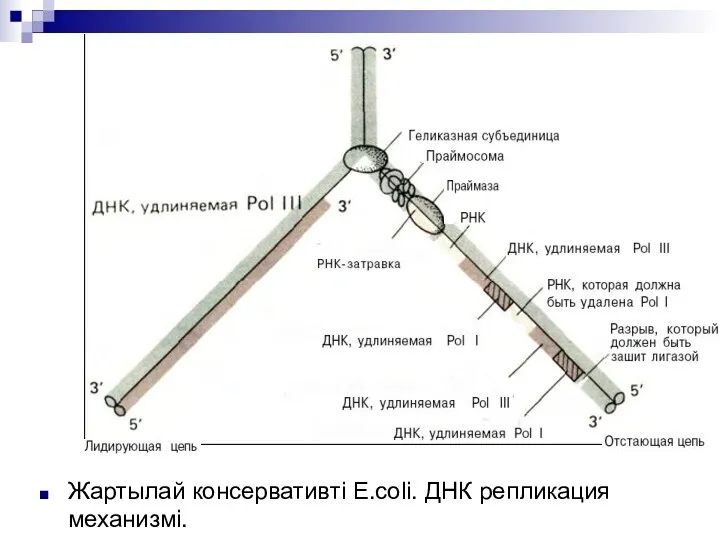
Жартылай консервативті Е.соli. ДНК репликация механизмі.
Жартылай консервативті Е.соli. ДНК репликация механизмі.
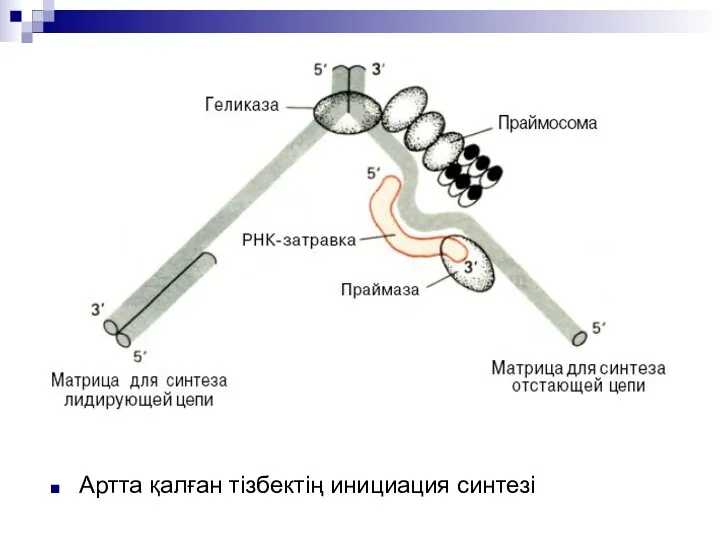
Артта қалған тізбектің инициация синтезі
Артта қалған тізбектің инициация синтезі
 Панкреатит. Ранние операции при панкреатите
Панкреатит. Ранние операции при панкреатите Эндодонтический инструментарий
Эндодонтический инструментарий Типовые нарушения функций печени
Типовые нарушения функций печени Домедична допомога в умовах бойових дій (тактична медицина). Тема №3.5
Домедична допомога в умовах бойових дій (тактична медицина). Тема №3.5 Схема истории болезни
Схема истории болезни Расходный материал в медицине
Расходный материал в медицине ВИЧ инфекция. Клиника, лечение, профилактика
ВИЧ инфекция. Клиника, лечение, профилактика Травмы. Повреждения головы, груди, живота
Травмы. Повреждения головы, груди, живота Жегі қуысын егеу принциптері мен ерекшеліктері: 1.тоннельді егеу, 2.атипиялық қуыстар
Жегі қуысын егеу принциптері мен ерекшеліктері: 1.тоннельді егеу, 2.атипиялық қуыстар Организация государственного ветеринарного надзора
Организация государственного ветеринарного надзора ЛФК при варикозной болезни
ЛФК при варикозной болезни ВИЧ-инфекция. Истинные и ложные пути передачи. Диагностика. Меры профилактики
ВИЧ-инфекция. Истинные и ложные пути передачи. Диагностика. Меры профилактики John Hunter
John Hunter Нейропсихология позднего возраста. Старение (лекция 7)
Нейропсихология позднего возраста. Старение (лекция 7) ЭКГ қалыпты жағдайдағы
ЭКГ қалыпты жағдайдағы Дәлелді медицинаға кіріспе
Дәлелді медицинаға кіріспе Оценка новорожденного ребенка сразу после рождения и принципы ухода за новорожденным
Оценка новорожденного ребенка сразу после рождения и принципы ухода за новорожденным Телемедицина - использование компьютерных и телекоммуникационных технологий для обмена медицинской информацией
Телемедицина - использование компьютерных и телекоммуникационных технологий для обмена медицинской информацией Гостра печінкова недостатність
Гостра печінкова недостатність Острые лейкозы у детей
Острые лейкозы у детей Жүрек қан тамырлар жүйесін зерттеу
Жүрек қан тамырлар жүйесін зерттеу Тениаринхоз. Возбудитель тениаринхоза. Осложнения. Лечение
Тениаринхоз. Возбудитель тениаринхоза. Осложнения. Лечение Препарирование зуба
Препарирование зуба Электромагниттік терапия
Электромагниттік терапия Доброкачественные опухоли желудка
Доброкачественные опухоли желудка Хронические лимфопролиферативные заболевания
Хронические лимфопролиферативные заболевания Физическая реабилитация при нарушениях осанки
Физическая реабилитация при нарушениях осанки Методология консультирования женщин, идущих на прерывание беременности
Методология консультирования женщин, идущих на прерывание беременности