- Электронный парамагнитный резонанс

Содержание
- 2. Часть 1. Основы метода ЭПР Электронный Парамагнитный Резонанс
- 3. Электронный парамагнитный резонанс Электронный парамагнитный резонанс (ЭПР, EPR, ESR) - один из основных прямых методов обнаружения
- 4. Взаимосвязь механического и магнитного моментов Из классической механики известно, что момент количества движения электрона на круговой
- 5. Суммарный магнитный момент Таким образом, для орбитального движения μl/Pl=-e/2m Для спинового движения μs/Ps=-e/m Суммарный магнитный момент
- 6. Энергия электрона в магнитном поле равна E=μ*H*cos(μ,H), где μ - суммарный магнитный момент электрона, H -
- 7. Ориентация электронных спинов в магнитном поле В отсутствие внешнего магнитного поля все механические и магнитные спиновые
- 8. H E При наложении магнитного поля магнитные моменты электронов ориентируются по полю или против поля. Их
- 9. H E -1/2 gβH Система, где электроны распределены между двумя энергетическими уровнями, может поглощать электромагнитное излучение.
- 10. Почему ЭПР можно наблюдать только у парамагнетиков? Разность между числом электронов (заселенностью) на двух разных энергетических
- 11. Метод был открыт Евгением Константиновичем Завойским в Казанском Университете в 1944 году. Он заметил, что монокристалл
- 12. Усилитель Записывающее устройство Аттенюатор Микроволновой источник (клистрон) Микроволновой резонатор Детектор Образец Двойной Т-мост ЭПР спектрометр
- 13. В Казанском ГУ
- 14. Прибор Завойского для измерения ЯМР и ЭПР
- 15. Прибор Завойского для измерения ЭПР
- 16. Как получить сигнал ЭПР ? Поглощение электромагнитных волн в радиоспектрометре ЭПР можно определять двумя способами: 1
- 17. Что такое сигнал ЭПР ? ν = const H поглощение hν = gβH Кроме того, в
- 18. Часть 2. Спектры ЭПР Электронный Парамагнитный Резонанс
- 19. 77 K 300 K УФ облученный цистеин Примеры сигналов ЭПР g=4,3 g=2,25 Нитрозильные комплексы g=2,03 g=1,94
- 20. Основные характеристики сигнала ЭПР 4) сверхтонкая структура
- 21. Амплитуда сигнала ЭПР C = Cs(S / SS) Площадь S под линией поглощения прямо пропорциональна концентрации
- 22. Ширина сигнала ЭПР где Δ t - время измерения и ΔE - неопределенность энергии системы. Ширина
- 23. Спин-решеточная релаксация (T1) обусловлена взаимодействием магнитного момента электрона с решеткой (т.е. средой в которой находятся парамагнитные
- 24. Большое T Малое ΔH Малое T Большое ΔH Комплексы металлов - T1=1 нс ΔH=10 мТ Свободные
- 25. 2. Взаимодействие спинового магнитного момента электрона с орбитальным магнитным моментом электрона приводит к уширению сигнала в
- 26. g-фактор g-фактор сигнала ЭПР - это не только параметр, отражающий вклад орбитального и спинового моментов в
- 27. Сверхтонкое взаимодействие Взаимодействие магнитного момента неспаренного электрона с магнитным моментом ядра соседнего атома (например, H или
- 28. Сверхтонкая структура Неспаренный электрон в радикале может быть расположен близко к двум протонам, как в радикале
- 29. Сверхтонкое взаимодействие (продолжение) N=O H3C СH электрон протон Атом азота имеет магнитный момент который ориентирован во
- 30. Часть 3. Практическое применение в медико-биологических исследованиях Электронный Парамагнитный Резонанс
- 31. Применение ЭПР в медико-биологических исследованиях: естественные сигналы ЭПР метод спиновых меток и зондов метод спиновых ловушек
- 32. Применение ЭПР в медико-биологических исследованиях: естественные сигналы ЭПР метод спиновых меток и зондов метод спиновых ловушек
- 33. Если взглянуть на спектр ЭПР, например, печени крысы, то можно увидеть сигналы цитохрома Р-450, сигнал метгемоглобина
- 34. Радикал аскорбата аскорбат радикал аскорбата дегидроаскорбат Сигнал ЭПР радикала аскорбата можно наблюдать непосредственно в крови или
- 35. Спектр ЭПР облученного УФ светом цистеина Повысить концентрацию свободных радикалов, для избежания трудностей при их изучении,
- 36. Сигналы ЭПР семихинонных или феноксильных радикалов семихинонный радикал убихинона Q10 феноксильный радикал α-токоферола семихинонный радикал α-токоферола
- 37. Применение ЭПР в медико-биологических исследованиях: естественные сигналы ЭПР метод спиновых меток и зондов метод спиновых ловушек
- 38. Спектр ЭПР нитроксильных радикалов состоит, в простейшем случае, из трех линий равной интенсивности, благодаря взаимодействию неспаренного
- 39. Время корреляции τc Если нитроксильный радикал находится в водном растворе, то его вращение является изотропным и
- 40. Используя уравнение Стокса-Эйнштейна можно определить значение вязкости в соответствующей макроскопической системе где R - эффективный радиус
- 41. Представленные спектры ЭПР отражают поведение зонда ТЕМПО в средах с различной гидрофобностью. Определив отношения a /
- 42. Спектр ЭПР спинового зонда 5-доксил стеарата в мембране эритроцита При анизотропном вращении стеариновой кислоты вокруг длинной
- 43. Параметр упорядоченности S равен 1, есливращение зонда происходит только вокруг нормали к плоскости мембраны. В «замороженных»
- 44. Изменения сигнала ЭПР при уменьшении S и возрастании угла отклонения конуса вращения γ
- 45. Изменения сигнала ЭПР при удалении нитроксильного радикала от полярной карбоксильной группы жирной кислоты С-5 С-9 С-12
- 46. Cпектр ЭПР рН чувствительного зонда (pK=4,7) при разных рН Казалось бы, что измерение рН достаточно легко
- 47. aN=(aNR + aNRH+)/2 Зависимость параметра f и aN от рН f =(IR)/(IR + IRH+) aNR и
- 48. Спектр ЭПР нативного химотрипсина, помеченного спиновой меткой. I1 I0 Денатурация белка приводит к увеличению подвижности полипептидных
- 49. В основе метода спиновых меток лежит тот же принцип изменения спектра ЭПР нитроксильного радикала в зависимости
- 50. Аллостерические эффекты в гемоглобине, меченым иминоксольным радикалом в геминовой группе при 25C. a - спектры ЭПР,
- 51. Применение ЭПР в медико-биологических исследованиях: естественные сигналы ЭПР метод спиновых меток и зондов метод спиновых ловушек
- 52. Основы метода спиновых ловушек (spin trapping assay) Концентрация свободных радикалов в клетках и тканях не превышает
- 53. Характеристики спектра ЭПР спиновых аддуктов α-фенил-N-трет-бутил нитрон (PBN) PBN спиновый аддукт aN aβH ΔHpp
- 54. Основные виды спиновых ловушек нитрозосоединения нитроны Нитронил-нитроксилы PBN DMPO нитрозобензол трет-нитрозобутан DEPMPO NNR
- 55. Основные требования к спиновым ловушкам Быстрые реакции с радикалами Отсутствие побочных реакций Образование стабильных аддуктов Отличие
- 56. Кинетические и спектральные характеристики спиновых аддуктов
- 57. Идентификация радикалов по спектрам ЭПР спиновых аддуктов Если известны: (1) тип СЛ, (2) Р-ритель, (3) aN
- 59. Скачать презентацию

Часть 1. Основы метода ЭПР
Электронный Парамагнитный
Резонанс
Часть 1. Основы метода ЭПР
Электронный Парамагнитный
Резонанс
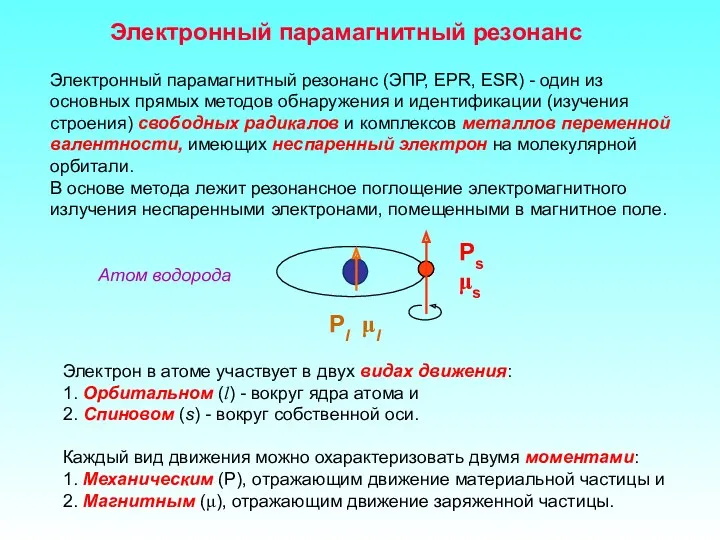
Электронный парамагнитный резонанс
Электронный парамагнитный резонанс (ЭПР, EPR, ESR) - один из
Электронный парамагнитный резонанс
Электронный парамагнитный резонанс (ЭПР, EPR, ESR) - один из
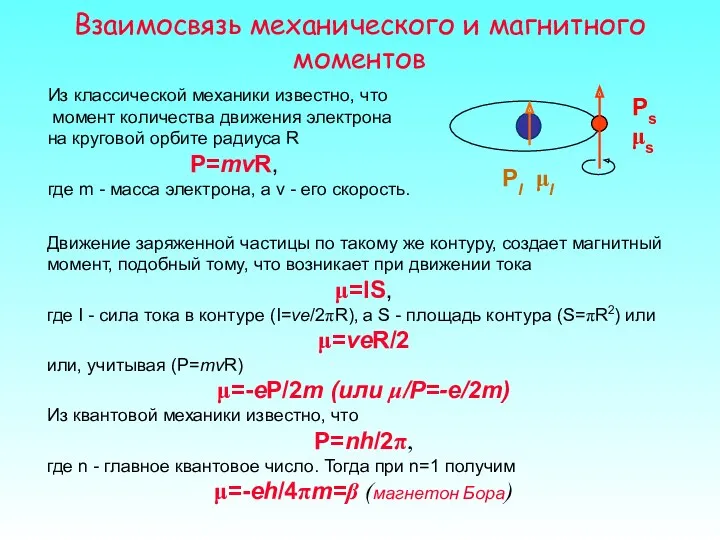
Взаимосвязь механического и магнитного моментов
Из классической механики известно, что
момент количества
Взаимосвязь механического и магнитного моментов
Из классической механики известно, что
момент количества

Суммарный магнитный момент
Таким образом, для орбитального движения
μl/Pl=-e/2m
Для спинового движения
μs/Ps=-e/m
Суммарный магнитный момент
Суммарный магнитный момент
Таким образом, для орбитального движения
μl/Pl=-e/2m
Для спинового движения
μs/Ps=-e/m
Суммарный магнитный момент

Энергия электрона в магнитном поле
равна
E=μ*H*cos(μ,H),
где μ - суммарный магнитный момент электрона,
Энергия электрона в магнитном поле
равна
E=μ*H*cos(μ,H),
где μ - суммарный магнитный момент электрона,
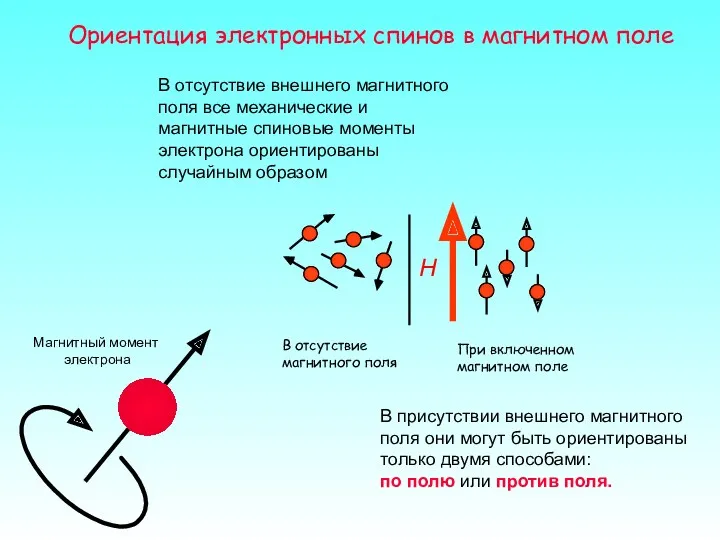
Ориентация электронных спинов в магнитном поле
В отсутствие внешнего магнитного поля все
Ориентация электронных спинов в магнитном поле
В отсутствие внешнего магнитного поля все
H
E
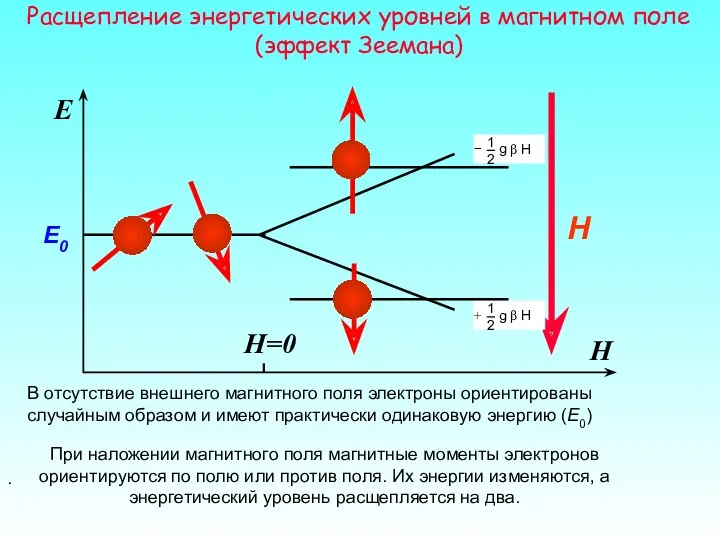
При наложении магнитного поля магнитные моменты электронов ориентируются по полю или
H
E
При наложении магнитного поля магнитные моменты электронов ориентируются по полю или
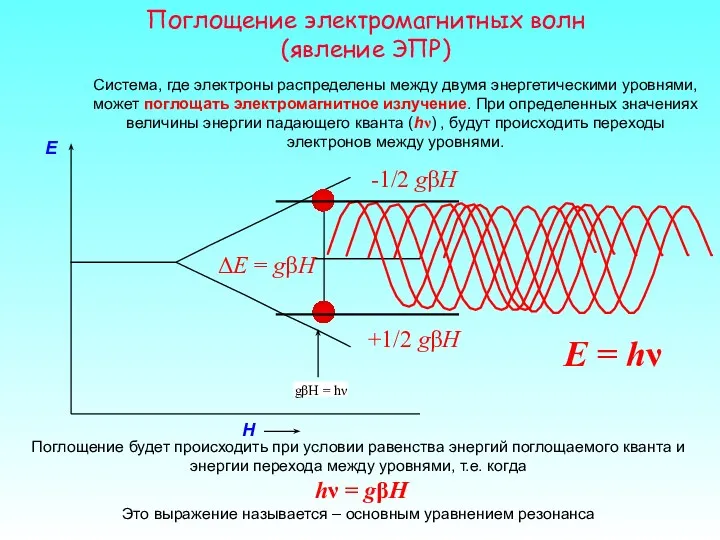
H
E
-1/2 gβH
Система, где электроны распределены между двумя энергетическими уровнями, может поглощать
H
E
-1/2 gβH
Система, где электроны распределены между двумя энергетическими уровнями, может поглощать

Почему ЭПР можно наблюдать только у парамагнетиков?
Разность между числом электронов (заселенностью)
Почему ЭПР можно наблюдать только у парамагнетиков?
Разность между числом электронов (заселенностью)

Метод был открыт Евгением Константиновичем Завойским в Казанском Университете в 1944
Метод был открыт Евгением Константиновичем Завойским в Казанском Университете в 1944
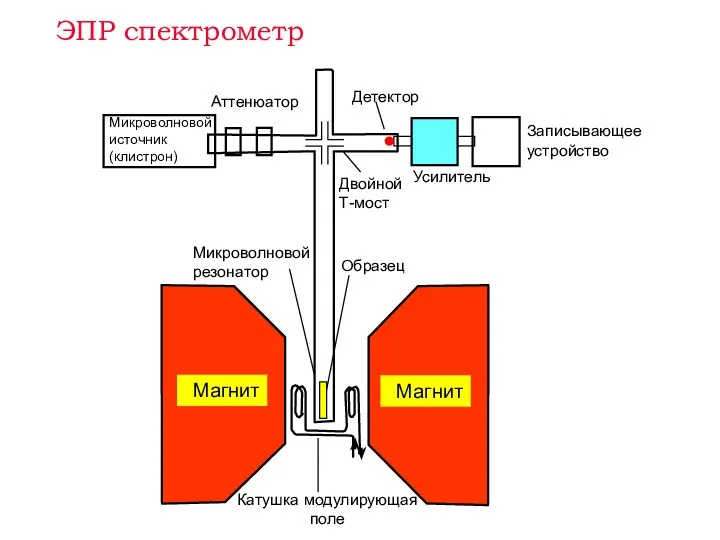
Усилитель
Записывающее устройство
Аттенюатор
Микроволновой источник
(клистрон)
Микроволновой резонатор
Детектор
Образец
Двойной Т-мост
ЭПР спектрометр
Усилитель
Записывающее устройство
Аттенюатор
Микроволновой источник
(клистрон)
Микроволновой резонатор
Детектор
Образец
Двойной Т-мост
ЭПР спектрометр

В Казанском ГУ
В Казанском ГУ

Прибор Завойского для измерения ЯМР и ЭПР
Прибор Завойского для измерения ЯМР и ЭПР

Прибор Завойского для измерения ЭПР
Прибор Завойского для измерения ЭПР
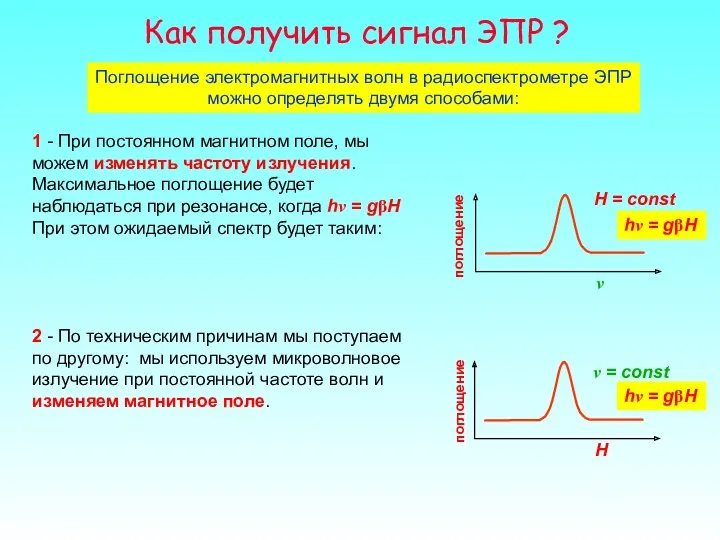
Как получить сигнал ЭПР ?
Поглощение электромагнитных волн в радиоспектрометре ЭПР можно
Как получить сигнал ЭПР ?
Поглощение электромагнитных волн в радиоспектрометре ЭПР можно
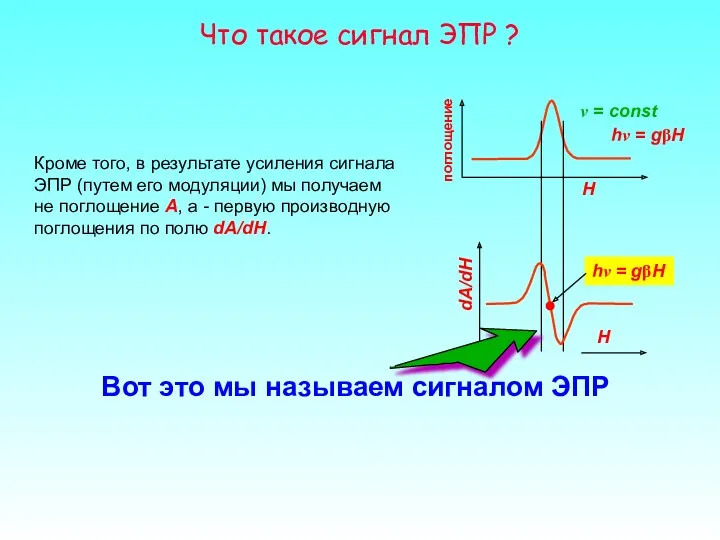
Что такое сигнал ЭПР ?
ν = const
H
поглощение
hν = gβH
Кроме того, в
Что такое сигнал ЭПР ?
ν = const
H
поглощение
hν = gβH
Кроме того, в

Часть 2. Спектры ЭПР
Электронный Парамагнитный
Резонанс
Часть 2. Спектры ЭПР
Электронный Парамагнитный
Резонанс
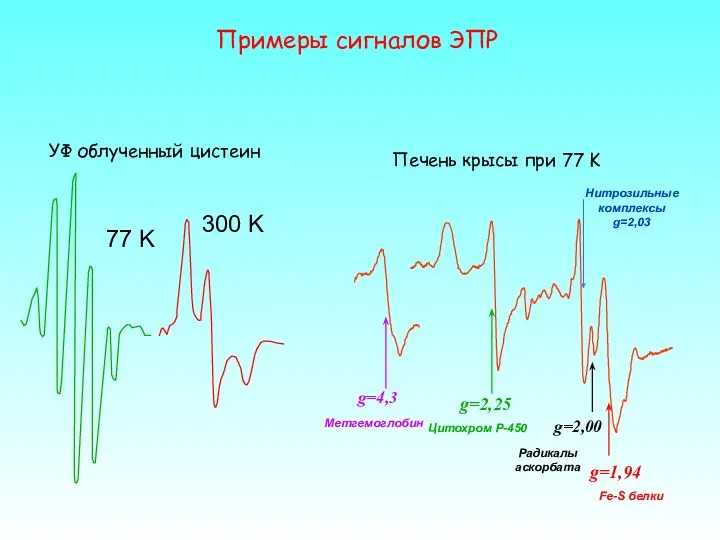
77 K
300 K
УФ облученный цистеин
Примеры сигналов ЭПР
g=4,3
g=2,25
Нитрозильные
комплексы
g=2,03
g=1,94
Fe-S
77 K
300 K
УФ облученный цистеин
Примеры сигналов ЭПР
g=4,3
g=2,25
Нитрозильные
комплексы
g=2,03
g=1,94
Fe-S
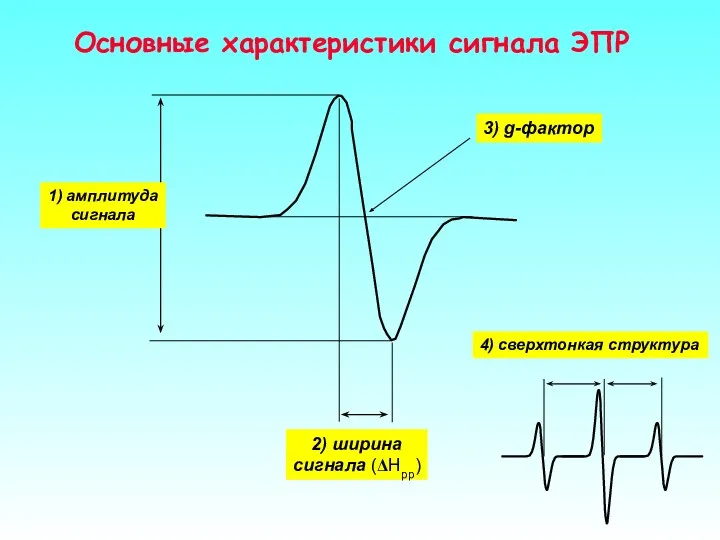
Основные характеристики сигнала ЭПР
4) сверхтонкая структура
Основные характеристики сигнала ЭПР
4) сверхтонкая структура
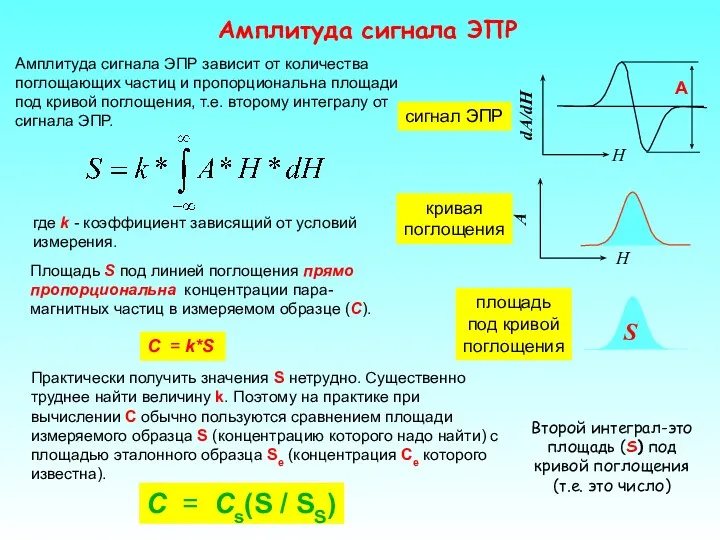
Амплитуда сигнала ЭПР
C = Cs(S / SS)
Площадь S под линией поглощения
Амплитуда сигнала ЭПР
C = Cs(S / SS)
Площадь S под линией поглощения

Ширина сигнала ЭПР
где Δ t - время измерения и ΔE -
Ширина сигнала ЭПР
где Δ t - время измерения и ΔE -
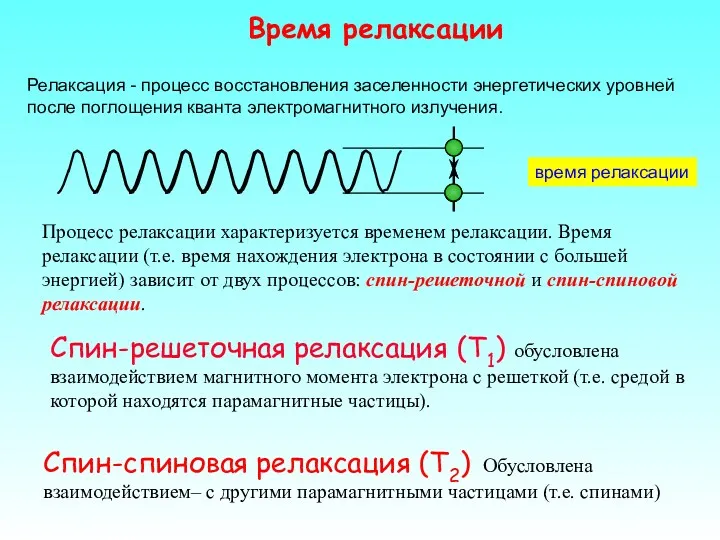
Спин-решеточная релаксация (T1) обусловлена взаимодействием магнитного момента электрона с решеткой (т.е.
Спин-решеточная релаксация (T1) обусловлена взаимодействием магнитного момента электрона с решеткой (т.е.
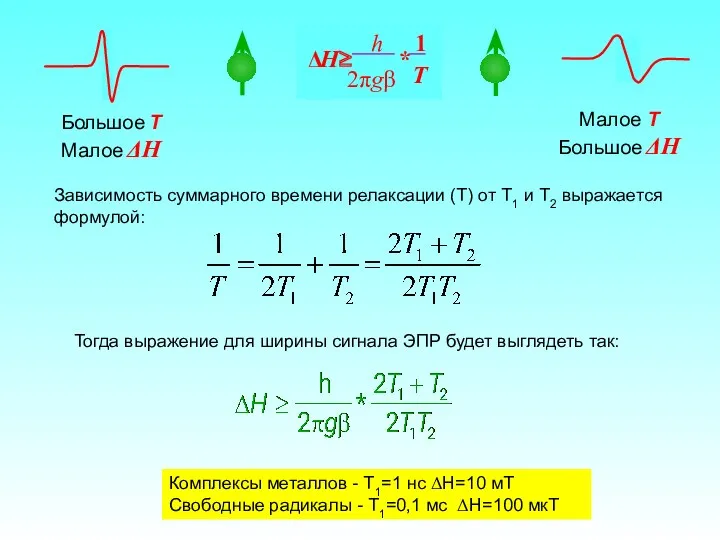
Большое T
Малое ΔH
Малое T
Большое ΔH
Комплексы металлов - T1=1 нс ΔH=10 мТ
Свободные
Большое T
Малое ΔH
Малое T
Большое ΔH
Комплексы металлов - T1=1 нс ΔH=10 мТ
Свободные
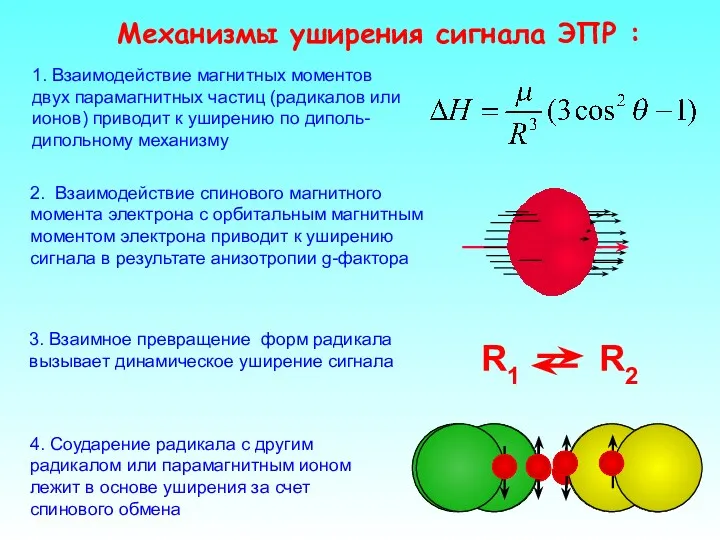
2. Взаимодействие спинового магнитного момента электрона с орбитальным магнитным моментом электрона
2. Взаимодействие спинового магнитного момента электрона с орбитальным магнитным моментом электрона
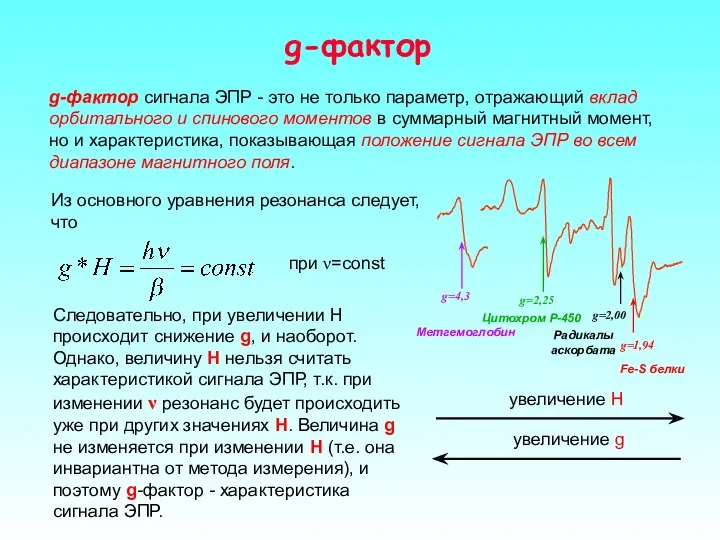
g-фактор
g-фактор сигнала ЭПР - это не только параметр, отражающий вклад орбитального
g-фактор
g-фактор сигнала ЭПР - это не только параметр, отражающий вклад орбитального
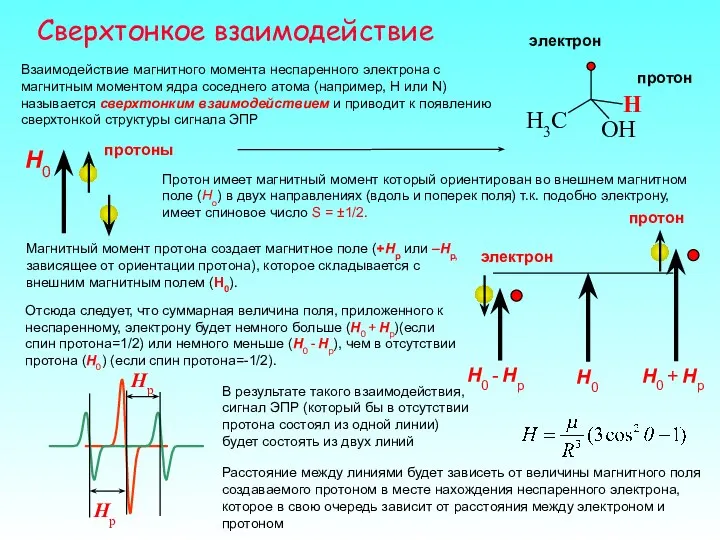
Сверхтонкое взаимодействие
Взаимодействие магнитного момента неспаренного электрона с магнитным моментом ядра соседнего
Сверхтонкое взаимодействие
Взаимодействие магнитного момента неспаренного электрона с магнитным моментом ядра соседнего

Сверхтонкая структура
Неспаренный электрон в радикале может быть расположен близко к двум
Сверхтонкая структура
Неспаренный электрон в радикале может быть расположен близко к двум
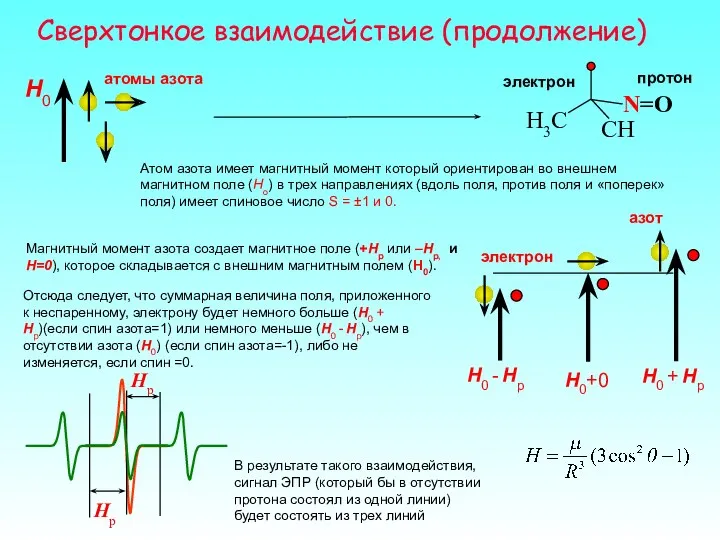
Сверхтонкое взаимодействие (продолжение)
N=O
H3C
СH
электрон
протон
Атом азота имеет магнитный момент который ориентирован во внешнем
Сверхтонкое взаимодействие (продолжение)
N=O
H3C
СH
электрон
протон
Атом азота имеет магнитный момент который ориентирован во внешнем
Часть 3. Практическое применение в медико-биологических исследованиях
Электронный Парамагнитный
Резонанс
Часть 3. Практическое применение в медико-биологических исследованиях
Электронный Парамагнитный
Резонанс

Применение ЭПР в медико-биологических исследованиях:
естественные сигналы ЭПР
метод спиновых меток и зондов
метод
Применение ЭПР в медико-биологических исследованиях:
естественные сигналы ЭПР
метод спиновых меток и зондов
метод

Применение ЭПР в медико-биологических исследованиях:
естественные сигналы ЭПР
метод спиновых меток и зондов
метод
Применение ЭПР в медико-биологических исследованиях:
естественные сигналы ЭПР
метод спиновых меток и зондов
метод
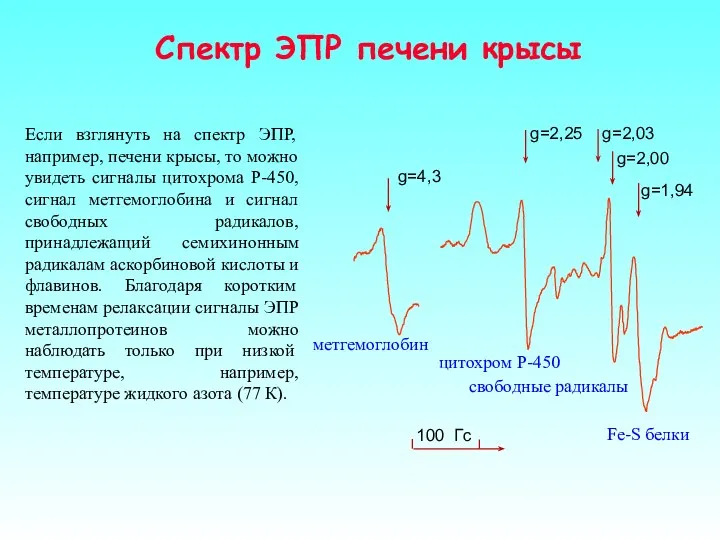
Если взглянуть на спектр ЭПР, например, печени крысы, то можно увидеть
Если взглянуть на спектр ЭПР, например, печени крысы, то можно увидеть
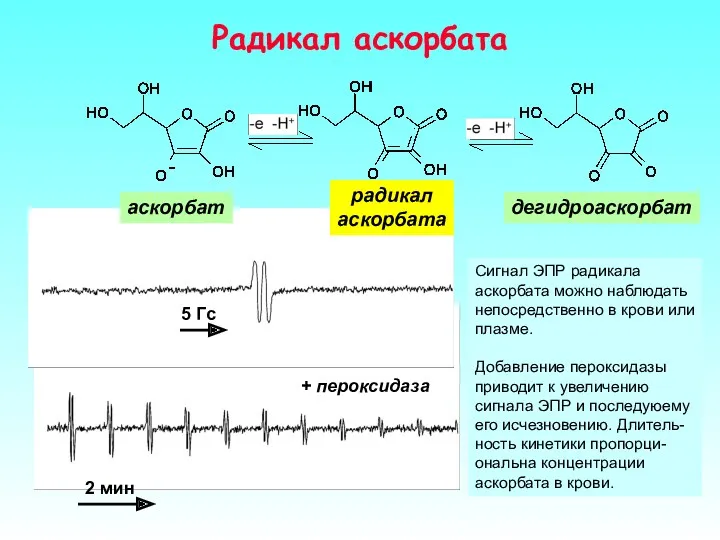
Радикал аскорбата
аскорбат
радикал
аскорбата
дегидроаскорбат
Сигнал ЭПР радикала аскорбата можно наблюдать непосредственно в крови или
Радикал аскорбата
аскорбат
радикал
аскорбата
дегидроаскорбат
Сигнал ЭПР радикала аскорбата можно наблюдать непосредственно в крови или
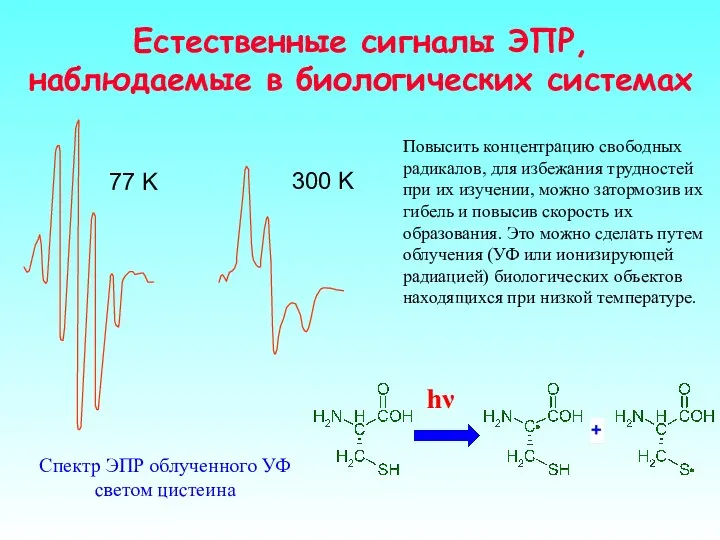
Спектр ЭПР облученного УФ светом цистеина
Повысить концентрацию свободных радикалов, для
Спектр ЭПР облученного УФ светом цистеина
Повысить концентрацию свободных радикалов, для
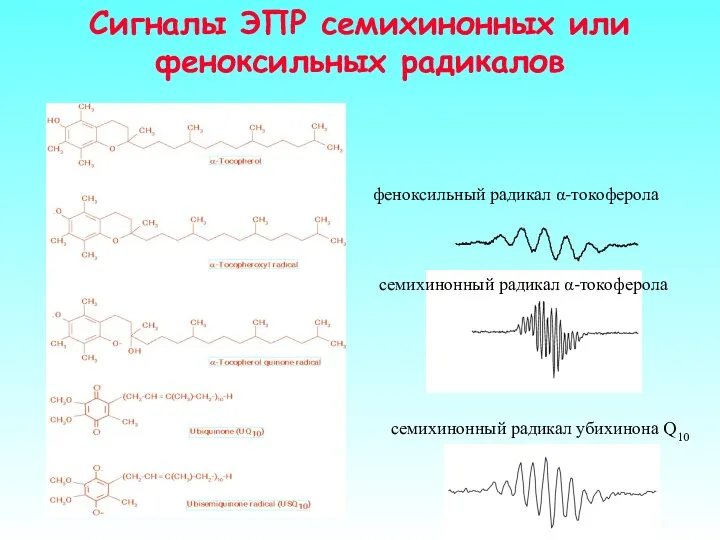
Сигналы ЭПР семихинонных или феноксильных радикалов
семихинонный радикал убихинона Q10
феноксильный радикал α-токоферола
семихинонный
Сигналы ЭПР семихинонных или феноксильных радикалов
семихинонный радикал убихинона Q10
феноксильный радикал α-токоферола
семихинонный

Применение ЭПР в медико-биологических исследованиях:
естественные сигналы ЭПР
метод спиновых меток и зондов
метод
Применение ЭПР в медико-биологических исследованиях:
естественные сигналы ЭПР
метод спиновых меток и зондов
метод

Спектр ЭПР нитроксильных радикалов состоит, в простейшем случае, из трех линий
Спектр ЭПР нитроксильных радикалов состоит, в простейшем случае, из трех линий

Время корреляции τc
Если нитроксильный радикал находится в водном растворе, то его
Время корреляции τc
Если нитроксильный радикал находится в водном растворе, то его

Используя уравнение Стокса-Эйнштейна можно определить значение вязкости в соответствующей макроскопической системе
Используя уравнение Стокса-Эйнштейна можно определить значение вязкости в соответствующей макроскопической системе

Представленные спектры ЭПР отражают поведение зонда ТЕМПО в средах с различной
Представленные спектры ЭПР отражают поведение зонда ТЕМПО в средах с различной
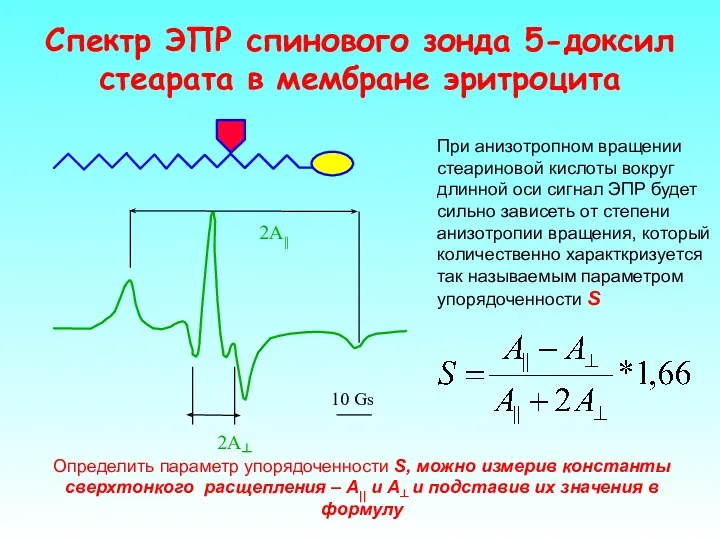
Спектр ЭПР спинового зонда 5-доксил стеарата в мембране эритроцита
При анизотропном вращении
Спектр ЭПР спинового зонда 5-доксил стеарата в мембране эритроцита
При анизотропном вращении
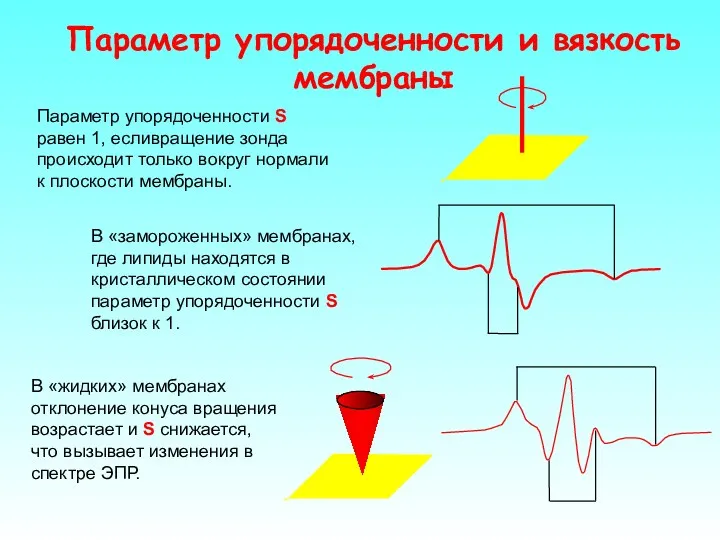
Параметр упорядоченности S равен 1, есливращение зонда происходит только вокруг нормали
Параметр упорядоченности S равен 1, есливращение зонда происходит только вокруг нормали
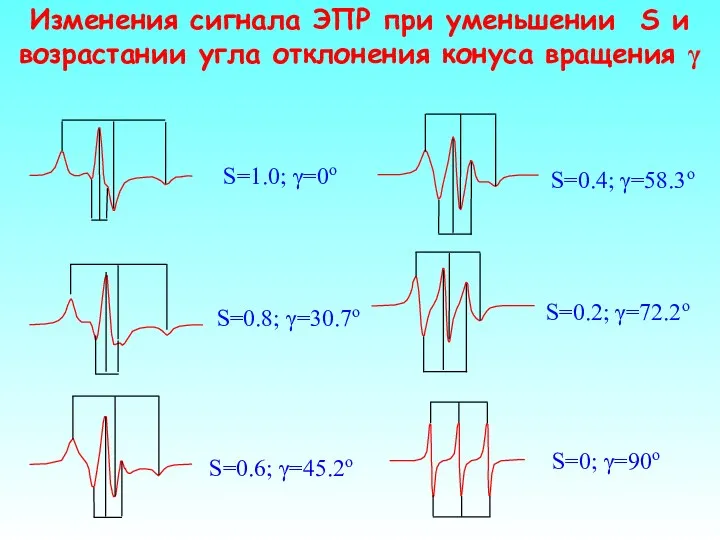
Изменения сигнала ЭПР при уменьшении S и возрастании угла отклонения конуса
Изменения сигнала ЭПР при уменьшении S и возрастании угла отклонения конуса
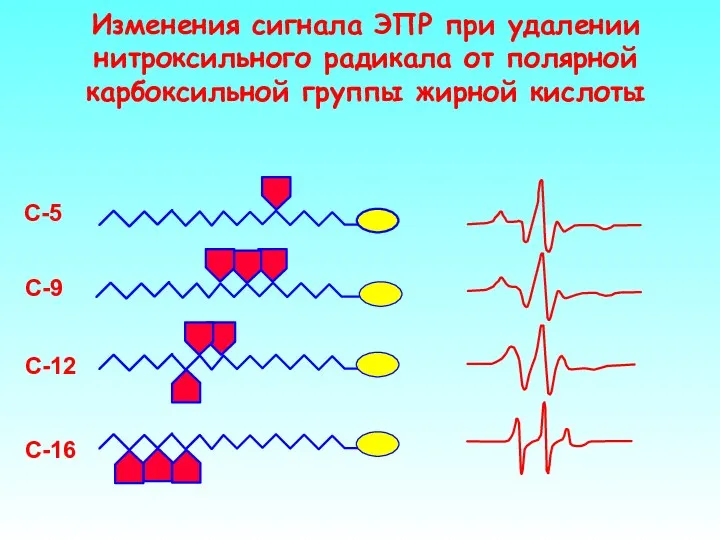
Изменения сигнала ЭПР при удалении нитроксильного радикала от полярной карбоксильной группы
Изменения сигнала ЭПР при удалении нитроксильного радикала от полярной карбоксильной группы

Cпектр ЭПР рН чувствительного зонда (pK=4,7) при разных рН
Казалось бы, что
Cпектр ЭПР рН чувствительного зонда (pK=4,7) при разных рН
Казалось бы, что

aN=(aNR + aNRH+)/2
Зависимость параметра f и aN от рН
f =(IR)/(IR +
aN=(aNR + aNRH+)/2
Зависимость параметра f и aN от рН
f =(IR)/(IR +
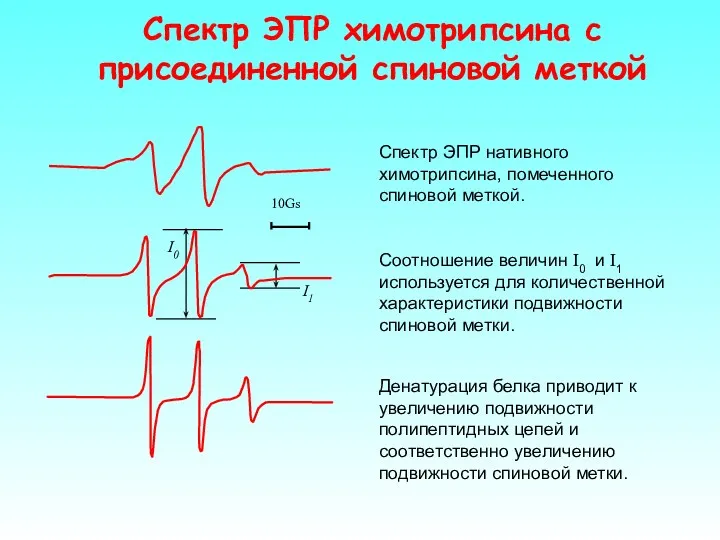
Спектр ЭПР нативного химотрипсина, помеченного спиновой меткой.
I1
I0
Денатурация белка приводит к
Спектр ЭПР нативного химотрипсина, помеченного спиновой меткой.
I1
I0
Денатурация белка приводит к
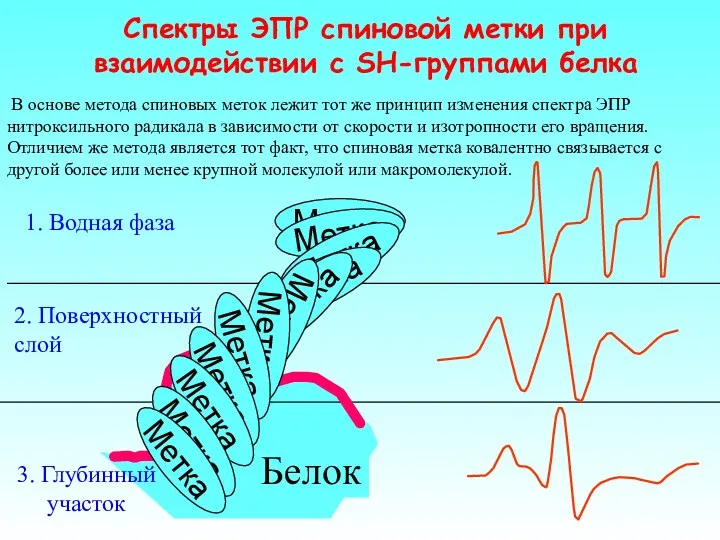
В основе метода спиновых меток лежит тот же принцип изменения
В основе метода спиновых меток лежит тот же принцип изменения

Аллостерические эффекты в гемоглобине, меченым иминоксольным радикалом в геминовой группе при
Аллостерические эффекты в гемоглобине, меченым иминоксольным радикалом в геминовой группе при

Применение ЭПР в медико-биологических исследованиях:
естественные сигналы ЭПР
метод спиновых меток и зондов
метод
Применение ЭПР в медико-биологических исследованиях:
естественные сигналы ЭПР
метод спиновых меток и зондов
метод

Основы метода спиновых ловушек
(spin trapping assay)
Концентрация свободных радикалов в клетках и
Основы метода спиновых ловушек
(spin trapping assay)
Концентрация свободных радикалов в клетках и
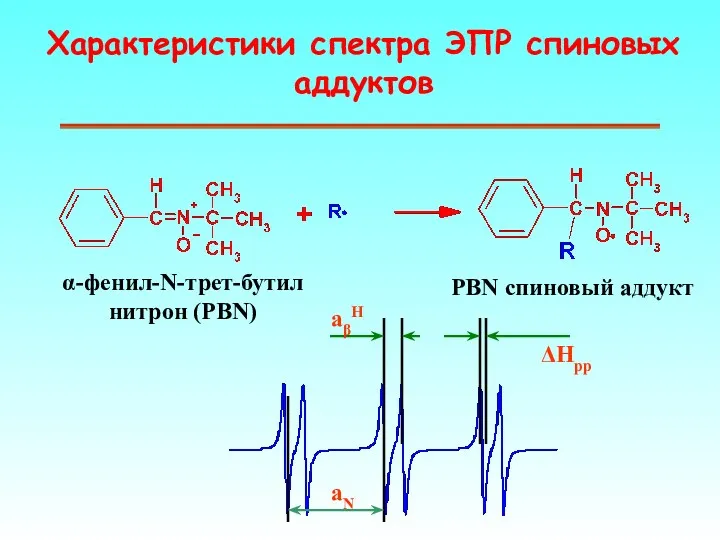
Характеристики спектра ЭПР спиновых аддуктов
α-фенил-N-трет-бутил нитрон (PBN)
PBN спиновый аддукт
aN
aβH
ΔHpp
Характеристики спектра ЭПР спиновых аддуктов
α-фенил-N-трет-бутил нитрон (PBN)
PBN спиновый аддукт
aN
aβH
ΔHpp
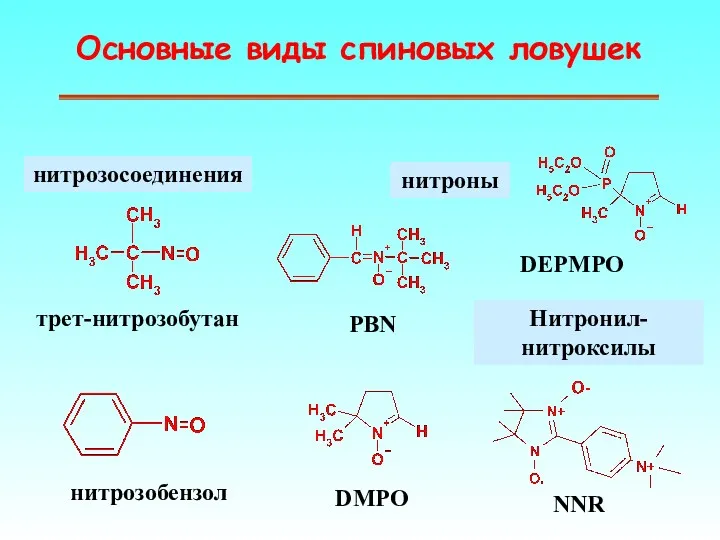
Основные виды спиновых ловушек
нитрозосоединения
нитроны
Нитронил-нитроксилы
PBN
DMPO
нитрозобензол
трет-нитрозобутан
DEPMPO
NNR
Основные виды спиновых ловушек
нитрозосоединения
нитроны
Нитронил-нитроксилы
PBN
DMPO
нитрозобензол
трет-нитрозобутан
DEPMPO
NNR

Основные требования к спиновым ловушкам
Быстрые реакции с радикалами
Отсутствие побочных реакций
Образование стабильных
Основные требования к спиновым ловушкам
Быстрые реакции с радикалами
Отсутствие побочных реакций
Образование стабильных

Кинетические и спектральные характеристики спиновых аддуктов
Кинетические и спектральные характеристики спиновых аддуктов
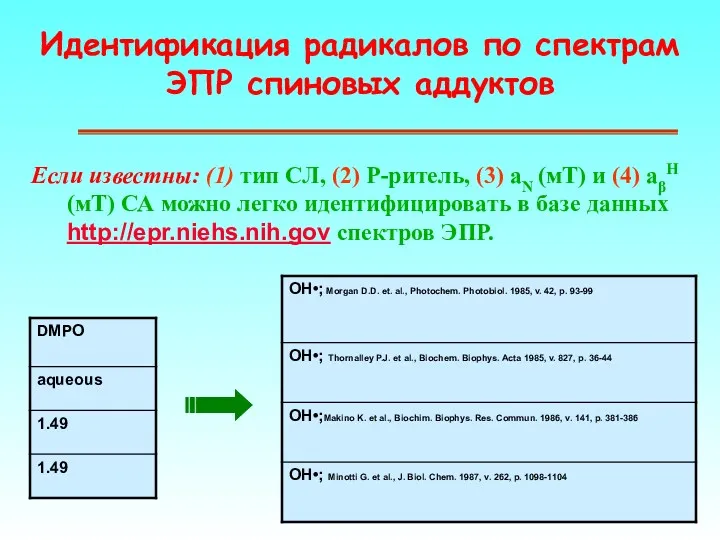
Идентификация радикалов по спектрам ЭПР спиновых аддуктов
Если известны: (1) тип СЛ,
Идентификация радикалов по спектрам ЭПР спиновых аддуктов
Если известны: (1) тип СЛ,
 Утомление, переутомление, перетренированность. Хроническое физическое перенапряжение систем организма. Внезапная смерть в спорте
Утомление, переутомление, перетренированность. Хроническое физическое перенапряжение систем организма. Внезапная смерть в спорте Принципы лучевой терапии ЗНО. Брахитерапия и сочетано-лучевая терапия
Принципы лучевой терапии ЗНО. Брахитерапия и сочетано-лучевая терапия Микробиологические методы лабораторной диагностики инфекционных заболеваний
Микробиологические методы лабораторной диагностики инфекционных заболеваний Особиста профілактика інфекційних захворюваннь
Особиста профілактика інфекційних захворюваннь Хронический пиелонефрит в практике участкового терапевта
Хронический пиелонефрит в практике участкового терапевта Геморрагическая лихорадка с почечным синдромом (ГЛПС)
Геморрагическая лихорадка с почечным синдромом (ГЛПС) Теория рационального питания. Гигиенические требования к рациональному питанию человека
Теория рационального питания. Гигиенические требования к рациональному питанию человека Виды ущемлений грыж
Виды ущемлений грыж Острые кишечные инфекции. Шигеллез. Сальмонеллез. (Лекция 4)
Острые кишечные инфекции. Шигеллез. Сальмонеллез. (Лекция 4) Микробиология чумы
Микробиология чумы Спортивное питание
Спортивное питание Лечение псориаза средней тяжести и тяжелого течения в эру биологической терапии
Лечение псориаза средней тяжести и тяжелого течения в эру биологической терапии Морально-этические проблемы трансплантологии
Морально-этические проблемы трансплантологии Нәрестенің гемолитикалық ауруы
Нәрестенің гемолитикалық ауруы Инфекции, передаваемые половым путём. Пути передачи, основные заболевания, меры профилактики
Инфекции, передаваемые половым путём. Пути передачи, основные заболевания, меры профилактики Жақ - бет аймағының одонтогенді емес ісіктері
Жақ - бет аймағының одонтогенді емес ісіктері Гипертензия, ишемическая болезнь сердца
Гипертензия, ишемическая болезнь сердца Острая жировая дистрофия печени. Холестатический гепатоз беременных
Острая жировая дистрофия печени. Холестатический гепатоз беременных Afecțiunile frecvente în neurorecuperare
Afecțiunile frecvente în neurorecuperare Основные симптомы неврологических расстройств
Основные симптомы неврологических расстройств Болезни органов дыхания. Гигиена дыхания
Болезни органов дыхания. Гигиена дыхания Стабільна ішемічна хвороба серця
Стабільна ішемічна хвороба серця Florence Nightingale - Pioneer of nursing
Florence Nightingale - Pioneer of nursing Клиническая анатомия позвоночника и спинного мозга
Клиническая анатомия позвоночника и спинного мозга Методы лучевой диагностики при исследовании тонкой кишки
Методы лучевой диагностики при исследовании тонкой кишки Курация больного с оформлением учебной истории болезни
Курация больного с оформлением учебной истории болезни Культура питания. Правила рационального питания
Культура питания. Правила рационального питания Санаторий-профилакторий ООО СО АРУ Саянская благодать
Санаторий-профилакторий ООО СО АРУ Саянская благодать