Эмбриология. Оценка качества ооцитов. Факторы влияющие на качество ооцитов. Классическая методика эко. Оценка оплодотворения презентация
- Эмбриология. Оценка качества ооцитов. Факторы влияющие на качество ооцитов. Классическая методика эко. Оценка оплодотворения

Содержание
- 2. Подготовка к пункции Успех вспомогательных репродуктивных технологий зависит от правильной координации медицинского и научного подходов к
- 3. Номер эмбриологического протокола Номер медицинской карты пациента Дата и время проведения пункции ФИО супруги, супруга ФИО
- 4. Подготовка к проведению пункции Чашка с культуральной средой (среда для оплодотворения, с оптимальным состав для осуществления
- 5. Трансвагинальная пункция яичников Уровень отрицательного давления в вакуумном насосе 100–120 mmHG
- 6. Забор ооцитов Фолликулярный аспират исследуют немедленно под микроскопом на подогреваемой поверхности Ооциты отмываются в среде для
- 7. 25 мкл 0,5-0,7 мл 1,0 мл
- 8. Рис. Схематическое изображение строения яицеклетки человека. 1 – ядро с проядрышком; 2 – плазмолемма, оолемма; 3
- 9. Оболочки Зернистая оболочка – клетки фолликулярного эпителия (кумулюса). Лучистый венец (Zone radiate) – слой фолликулярных клеток
- 10. Органеллы Эндоплазмотическая сеть Аппарат Гольджи Лизосомы Вакуоли Митохондрии Рибосомы Цитоскелет (микротрубочки и микрофиламенты) Отсутствуют центриоли. В
- 11. Желточные мембранные гранулы – фосфо- и липопротеины. Обеспечивают развитие эмбриона в течение первой недели развития. Кортикальные
- 12. Основной фактор, определяющий результаты IVF – Морфо-функциональное состояние яйцеклетки Качество ооцита – фактор, ограничивающий фертильность женщины
- 13. Классификация ооцит-кумулюсного комплекса B. Метафаза I A. Профаза I Зародышевый пузырек Незрелый ооцит Клетки кумулюса, расположены
- 14. Около 20% ооцитов не зрелые – в стадии МI или GV Асперируемые фолликулы Дефицит рецепторов ЛГ
- 15. Почему на пункции не получают ооцитов или получают меньше ожидаемого количества Патогенез синдрома пустого фолликула На
- 16. Качество ооцитов (прогностические факторы) Характеристики Морфологические (аномальные морфологические характеристики отмечаются у 60-70% ооцитов). (Van Blerkom J,
- 17. Микроскопия в поляризованном свете Микроскопия с использованием модуляционнного контраста Хоффмана, Намарского Зрелость ядра Ооцит на стадии
- 18. Hum Reprod Update. 2011 Jan-Feb; 17(1): 34–45. Predictive value of oocyte morphology in human IVF: a
- 19. Таблица. Критерии качества ооцита
- 20. Продолжение таблицы. Критерии качества ооцита
- 22. применением поляризационной микроскопии: анализ зоны пеллюцида и веретена деления фиброгранулярная структура, три слоя высокие двупреломляющие свойства
- 23. Дисморфизм (гипотеза Blerkom, Henry, 1992) Раннее созревание Гигантские ооциты Центральная грануляция Веретено деления Позднее созревание Агрегация
- 24. Криоконсервированные ооциты Возможные последствия Изменение метаболизма Ультраструктуральные повреждения (оргенеллы, цитоскелет), деполимеризация микротрубочек MII веретена деления (степень
- 25. Кумулятивный показатель беременности (возраст пациенток Криоконсервация эмбрионов Криоконсервация ооцитов
- 26. Ооцит с хорошей морфологией «компетентный ооцит» правильной (сферической) формы с однородной, равномерно гранулированной цитоплазмой с маленьким
- 27. Созревание ооцитов в условиях in vitro (IVM – in vitro maturation) репродуктивная технология 1991 год –
- 28. IVM метод Недостатки метода Неоднородный пул ооцитов (GV, MI, MII) Дифференциальное расписание оплодотворения (ICSI) Дисинхронизация этапов
- 29. Факторы, влияющие на качество ооцита Клинические факторы Гинекологические заболевания (синдром поликистозных яичников, эндометриоз, аутоимунный оофорит) Стимуляция
- 30. Клинические факторы. Стимуляция суперовуляции в циклах ЭКО влияет на микроокружение ооцита и как следствие на качество
- 31. Возраст 1.Остановка эмбриогенеза может быть результатом событий, связанных с созреванием или несостоятельностью метаболизма ооцита. Цитоплазма ооцита
- 32. Рисунок. Уровень анеуплоидии в 3816 FISH диагностиках ооцитов относительно материнского возраста. Распределение хромосом зависело от возраста
- 33. «Маркеры апоптоза как предикторы эффективности вспомогательных репродуктивных технологий» Грищенко Н.Г. Харьковский национальный медицинский университет Международный медицинский
- 34. Факторы, влияющие на качество ооцита Клинические факторы Гинекологические заболевания (синдром поликистозных яичников, эндометриоз, аутоимунный оофорит) Стимуляция
- 35. pH культуральной среды pH культуральной среды находится в физиологическом интервале 7,2 – 7,4. Бикарбонатный буфер культ.
- 36. Температура При снижение температуры изменяется веретено деления. Деполимеризация веретена деления Нарушения метаболизма, стабильности клеточных мембран итранспортных
- 37. Осмолярность Важной характеристикой среды является ее осмолярность, определяемая концентрацией и константами диссоциации ее компонентов. Обычно осмолярность
- 38. Как обеспечить стабильность среды культивирования Использовать минеральное масло. При работе с гаметами и эмбрионами вне инкубатора
- 39. О2 Ооцитам и эмбрионам необходим кислород, в условиях in vitro они генерируют АТФ путем аэробного окислительного
- 40. Выводы Получение ооцита Трансвагинальная функция фолликулов под ультрозвуковым контролем Температура 37°С Стабильный рН-7,4 Качество ооцитов На
- 41. Классическая методика экстракорпорального оплодотворения
- 42. Подготовка сперматозоидов к оплодотворению Активация спермиев в придатках яичка (эпидидимусе) – обретение подвижности. Сперматозоид должен встретить
- 43. Сперматозоид человека in vivo Подготовка спермы к оплодотворению Удаление семенной плазмы и простагландинов Удаление мертвых, неподвижных
- 44. Капацитация Биохимически капацитация — это изменение строения некоторых молекул (стеролов, липидов и гликозилированных белков) цитоплазматической мембраны
- 45. Спермий в половых путях самки. Схема оплодотворения. Капацитация Гиперактивация Преодоление 3-х естественных барьеров: клетки кумулюса и
- 46. Реакция зоны Блок полиспермии необходим для предотвращения нарушения оплодотворения Индуцируется фактором активации фосфолипазы С-zeta сперматозоида. Медленный
- 47. Подготовка спермы для проведения ЭКО и внутриматочной инсеминации Спермальный профиль пациента Выбор метода подготовки спермы к
- 48. Метод «swim-up» Принцип метода основан на различной подвижности сперматозоидов Критерии выбора метода: - нормозооспрмия - легкие
- 49. Метод «градиент плотности» Критерии выбора метода: - олиго-, астено-, тератозооспермия - присутствие значительной клеточной контаминации и
- 50. Подготовка спермы к проведению ИКСИ Критерии выбора: - олигоспермия, объем ≤ 0,5 мл - выраженная олиго-астенозооспермия,
- 51. Инсеменация ООЦИТОВ in vitro Фактор времени Тригер овуляции (ХГЧ)→36 часов→пункция → → 2- 6 часов→инсеменация Способность
- 52. Инсеменация ООЦИТОВ in vitro Количественный фактор Культивирование в большом объеме Количество прогрессивно подвижных сперматозоидов обычно должно
- 53. Сперматозоид капацитация гиперактивация связывание с zona pellucida акросомный экзоцитоз проникновение сквозь zona pellucida сплавление мембран спермия
- 54. Оценка зиготы (1-й день) Параметры: Пронуклеусы Проядрышки Полярные тельца Формирование пронуклеусов Мужской пронуклеус формируется через 8
- 55. Оценка оплодотворения (количество пронуклеусов) 3PN Отсутствие оплодотворения ~ 20-30% ооцитов не оплодотворяется (0р) по причине: -
- 56. Качество пронуклеарных зигот, их морфология Форма зиготы Цитоплазматическое гало (органеллы клетки: митохондрии смещены к центру и
- 57. Рисунок. Различные конфигурации, используемые для оценки морфологии пронуклеусов (положение в цитоплазме, размер)
- 58. Рисунок.Ориентация пронуклеусов относительно PBII Figure 2: Computer-dependent measurements of morphological features in zygotes. Оолемма (зеленый), цитоплазматическое
- 59. Classification of PNs by Tesarik et al., 1999 Zygotes classified according to Tesarik's system and the
- 60. Classification of PNs by Scott et al., 2000 Developmental prognosis for zygotes based on pronuclear pattern:
- 61. ESHRE системы оценки морфологии зиготы
- 63. Скачать презентацию

Подготовка к пункции
Успех вспомогательных репродуктивных технологий зависит от правильной координации медицинского
Подготовка к пункции
Успех вспомогательных репродуктивных технологий зависит от правильной координации медицинского
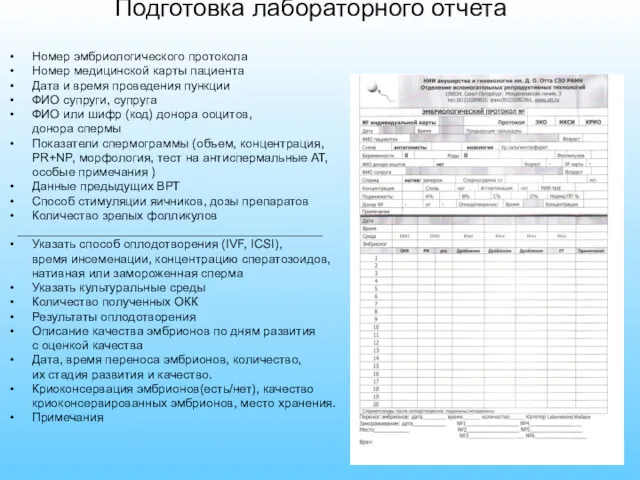
Номер эмбриологического протокола
Номер медицинской карты пациента
Дата и время проведения пункции
ФИО супруги,
Номер эмбриологического протокола
Номер медицинской карты пациента
Дата и время проведения пункции
ФИО супруги,

Подготовка к проведению пункции
Чашка с культуральной средой (среда для оплодотворения, с
Подготовка к проведению пункции
Чашка с культуральной средой (среда для оплодотворения, с
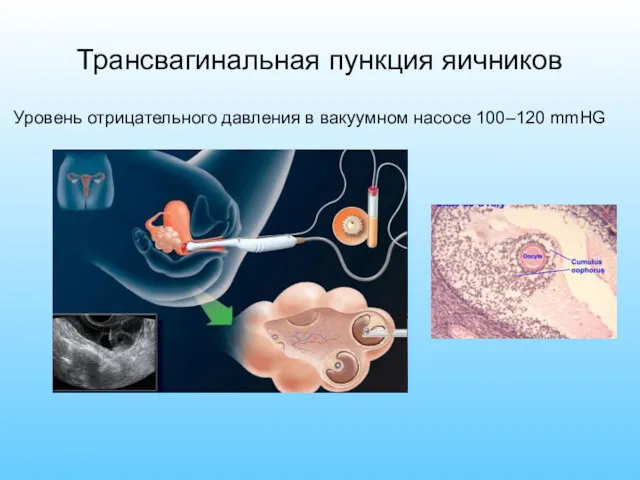
Трансвагинальная пункция яичников
Уровень отрицательного давления в вакуумном насосе 100–120 mmHG
Трансвагинальная пункция яичников
Уровень отрицательного давления в вакуумном насосе 100–120 mmHG
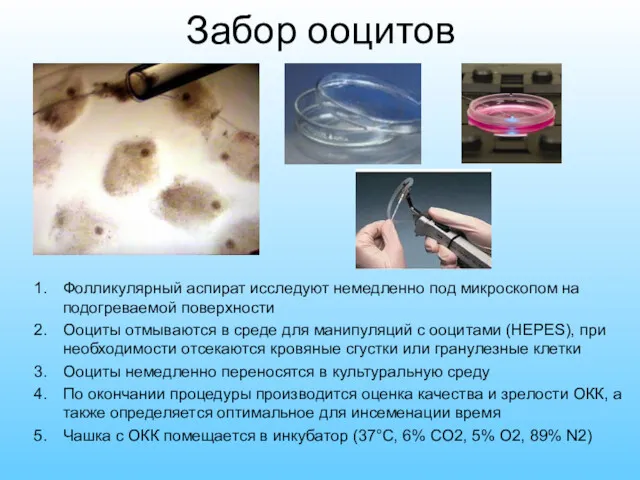
Забор ооцитов
Фолликулярный аспират исследуют немедленно под микроскопом на подогреваемой поверхности
Ооциты отмываются
Забор ооцитов
Фолликулярный аспират исследуют немедленно под микроскопом на подогреваемой поверхности
Ооциты отмываются
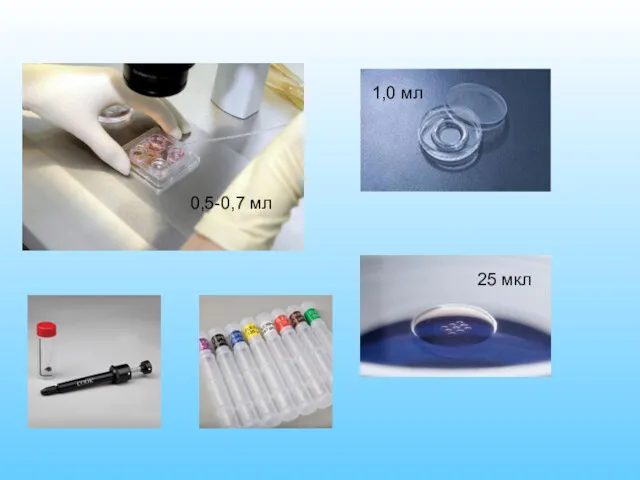
25 мкл
0,5-0,7 мл
1,0 мл
25 мкл
0,5-0,7 мл
1,0 мл

Рис. Схематическое изображение строения яицеклетки человека.
1 – ядро с проядрышком; 2 – плазмолемма, оолемма;
Рис. Схематическое изображение строения яицеклетки человека.
1 – ядро с проядрышком; 2 – плазмолемма, оолемма;
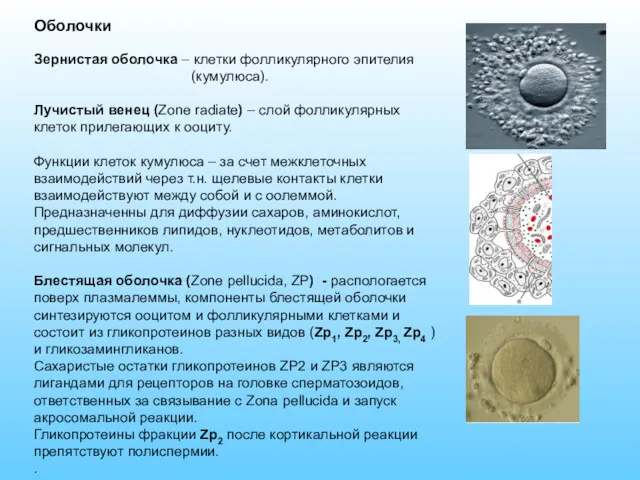
Оболочки
Зернистая оболочка – клетки фолликулярного эпителия
(кумулюса).
Лучистый венец (Zone radiate) –
Оболочки
Зернистая оболочка – клетки фолликулярного эпителия
(кумулюса).
Лучистый венец (Zone radiate) –

Органеллы
Эндоплазмотическая сеть
Аппарат Гольджи
Лизосомы
Вакуоли
Митохондрии
Рибосомы
Цитоскелет
Органеллы
Эндоплазмотическая сеть
Аппарат Гольджи
Лизосомы
Вакуоли
Митохондрии
Рибосомы
Цитоскелет
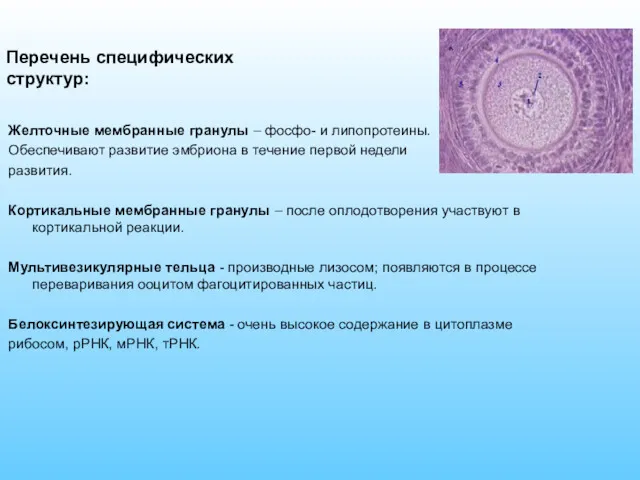
Желточные мембранные гранулы – фосфо- и липопротеины.
Обеспечивают развитие эмбриона в течение
Обеспечивают развитие эмбриона в течение

Основной фактор, определяющий результаты IVF –
Морфо-функциональное состояние яйцеклетки
Качество ооцита –
Основной фактор, определяющий результаты IVF –
Морфо-функциональное состояние яйцеклетки
Качество ооцита –
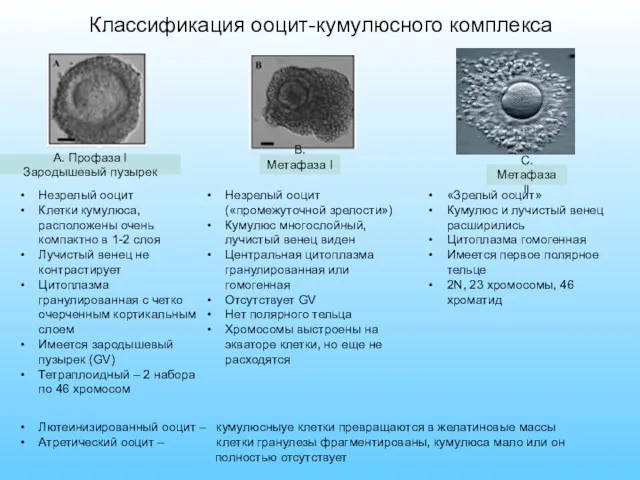
Классификация ооцит-кумулюсного комплекса
B. Метафаза I
A. Профаза I
Зародышевый пузырек
Незрелый ооцит
Клетки кумулюса, расположены
Классификация ооцит-кумулюсного комплекса
B. Метафаза I
A. Профаза I
Зародышевый пузырек
Незрелый ооцит
Клетки кумулюса, расположены

Около 20% ооцитов не зрелые – в стадии МI или GV
Асперируемые
Около 20% ооцитов не зрелые – в стадии МI или GV
Асперируемые
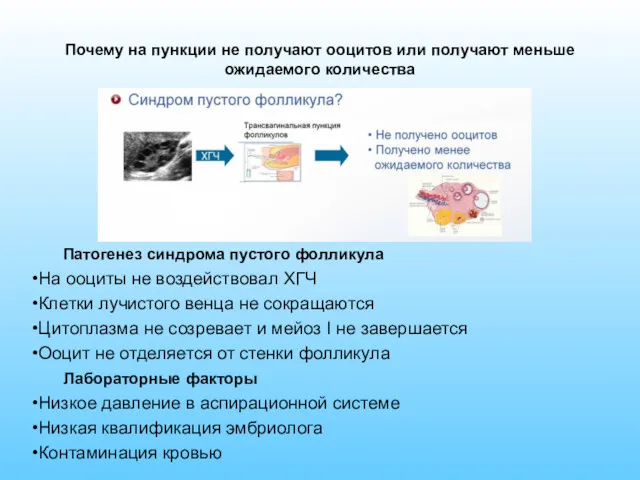
Почему на пункции не получают ооцитов или получают меньше ожидаемого количества
Почему на пункции не получают ооцитов или получают меньше ожидаемого количества

Качество ооцитов
(прогностические факторы)
Характеристики
Морфологические
(аномальные морфологические характеристики отмечаются у 60-70% ооцитов).
(Van Blerkom
Качество ооцитов
(прогностические факторы)
Характеристики
Морфологические
(аномальные морфологические характеристики отмечаются у 60-70% ооцитов).
(Van Blerkom
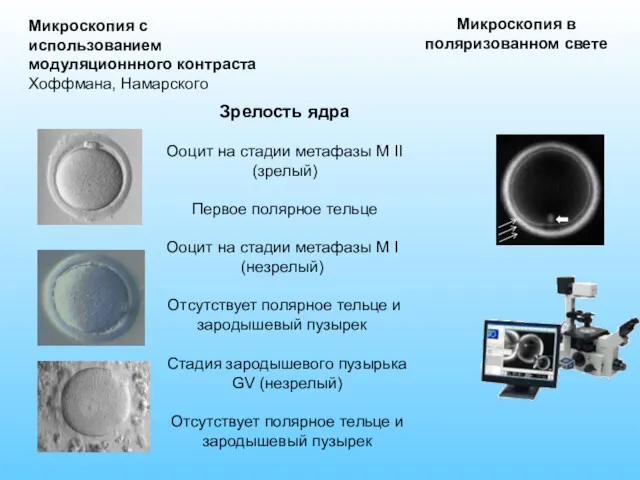
Микроскопия в
поляризованном свете
Микроскопия с использованием
модуляционнного контраста
Хоффмана, Намарского
Микроскопия в
поляризованном свете
Микроскопия с использованием
модуляционнного контраста
Хоффмана, Намарского

Hum Reprod Update. 2011 Jan-Feb; 17(1): 34–45.
Predictive value of oocyte
Hum Reprod Update. 2011 Jan-Feb; 17(1): 34–45.
Predictive value of oocyte
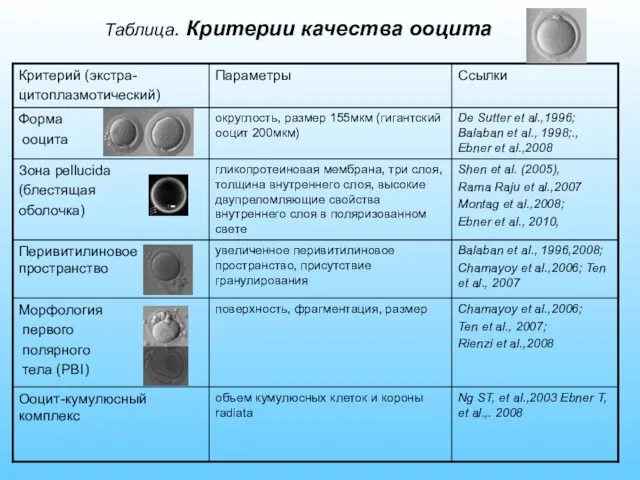
Таблица. Критерии качества ооцита
Таблица. Критерии качества ооцита
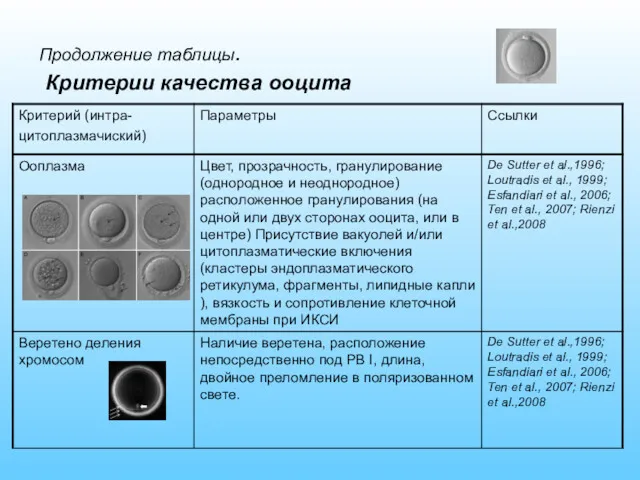
Продолжение таблицы.
Критерии качества ооцита
Продолжение таблицы.
Критерии качества ооцита
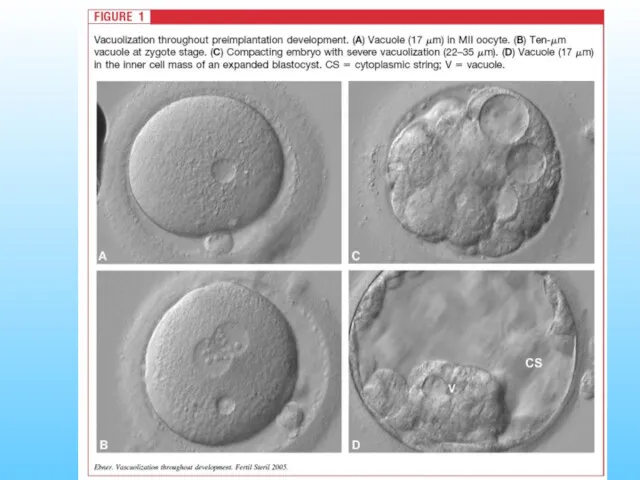
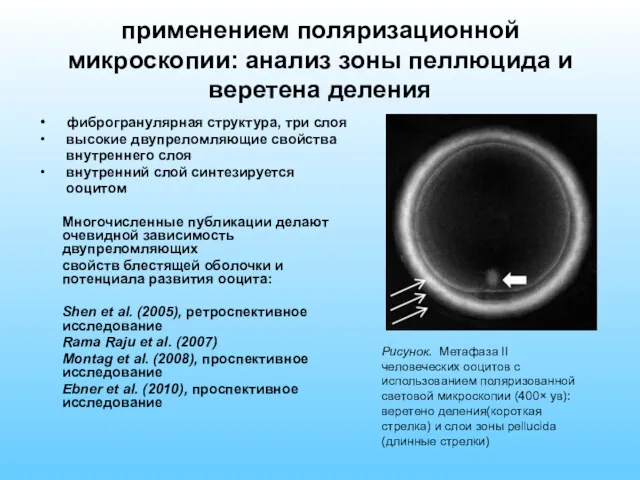
применением поляризационной
микроскопии: анализ зоны пеллюцида и веретена деления
фиброгранулярная структура, три
применением поляризационной
микроскопии: анализ зоны пеллюцида и веретена деления
фиброгранулярная структура, три
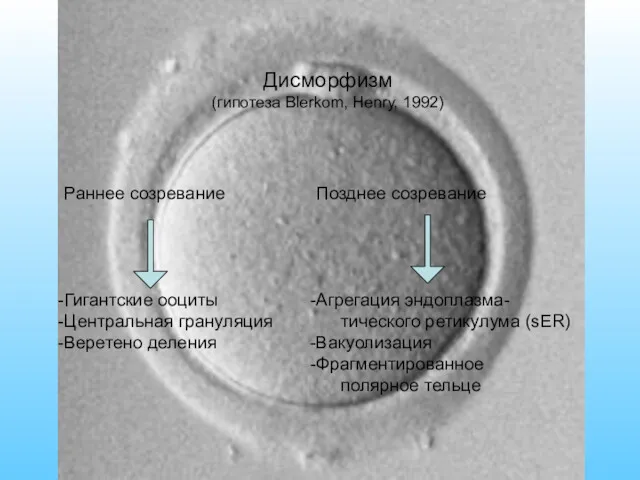
Дисморфизм
(гипотеза Blerkom, Henry, 1992)
Раннее созревание
Гигантские ооциты
Центральная грануляция
Веретено деления
Позднее созревание
Агрегация эндоплазма-
тического
Дисморфизм
(гипотеза Blerkom, Henry, 1992)
Раннее созревание
Гигантские ооциты
Центральная грануляция
Веретено деления
Позднее созревание
Агрегация эндоплазма-
тического

Криоконсервированные ооциты
Возможные последствия
Изменение метаболизма
Ультраструктуральные повреждения (оргенеллы, цитоскелет),
деполимеризация микротрубочек MII
Криоконсервированные ооциты
Возможные последствия
Изменение метаболизма
Ультраструктуральные повреждения (оргенеллы, цитоскелет),
деполимеризация микротрубочек MII

Кумулятивный показатель беременности
(возраст пациенток <39 лет)
Криоконсервация эмбрионов
Криоконсервация ооцитов
Кумулятивный показатель беременности
(возраст пациенток <39 лет)
Криоконсервация эмбрионов
Криоконсервация ооцитов
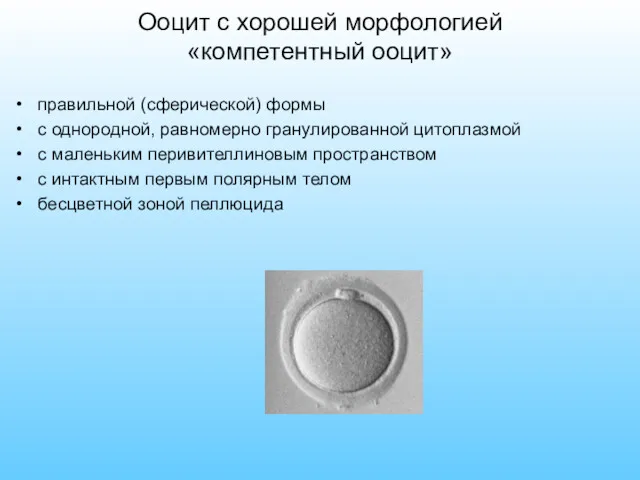
Ооцит с хорошей морфологией
«компетентный ооцит»
правильной (сферической) формы
с однородной, равномерно гранулированной цитоплазмой
с
Ооцит с хорошей морфологией
«компетентный ооцит»
правильной (сферической) формы
с однородной, равномерно гранулированной цитоплазмой
с

Созревание ооцитов в условиях in vitro
(IVM – in vitro maturation)
репродуктивная
Созревание ооцитов в условиях in vitro (IVM – in vitro maturation) репродуктивная
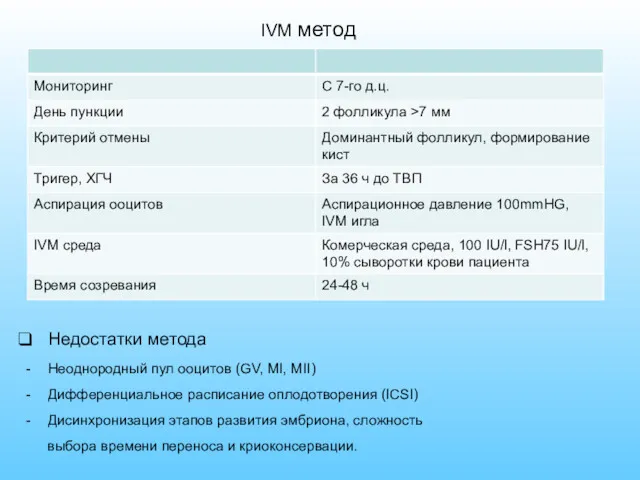
IVM метод
Недостатки метода
Неоднородный пул ооцитов (GV, MI, MII)
Дифференциальное расписание оплодотворения (ICSI)
Дисинхронизация
IVM метод
Недостатки метода
Неоднородный пул ооцитов (GV, MI, MII)
Дифференциальное расписание оплодотворения (ICSI)
Дисинхронизация

Факторы, влияющие на качество ооцита
Клинические
факторы
Гинекологические заболевания (синдром поликистозных яичников,
Факторы, влияющие на качество ооцита
Клинические
факторы
Гинекологические заболевания (синдром поликистозных яичников,

Клинические факторы. Стимуляция суперовуляции в циклах ЭКО
влияет на микроокружение ооцита и
Клинические факторы. Стимуляция суперовуляции в циклах ЭКО
влияет на микроокружение ооцита и

Возраст
1.Остановка эмбриогенеза может быть результатом событий, связанных с созреванием или
Возраст
1.Остановка эмбриогенеза может быть результатом событий, связанных с созреванием или

Рисунок. Уровень анеуплоидии в 3816 FISH диагностиках ооцитов относительно материнского возраста.
Рисунок. Уровень анеуплоидии в 3816 FISH диагностиках ооцитов относительно материнского возраста.

«Маркеры апоптоза как предикторы эффективности вспомогательных репродуктивных технологий»
Грищенко Н.Г.
Харьковский национальный медицинский
«Маркеры апоптоза как предикторы эффективности вспомогательных репродуктивных технологий»
Грищенко Н.Г.
Харьковский национальный медицинский

Факторы, влияющие на качество ооцита
Клинические
факторы
Гинекологические заболевания (синдром поликистозных яичников,
Факторы, влияющие на качество ооцита
Клинические
факторы
Гинекологические заболевания (синдром поликистозных яичников,

pH культуральной среды
pH культуральной среды находится в физиологическом интервале 7,2 –
pH культуральной среды
pH культуральной среды находится в физиологическом интервале 7,2 –

Температура
При снижение температуры изменяется
веретено деления.
Деполимеризация веретена деления
Нарушения
Температура
При снижение температуры изменяется
веретено деления.
Деполимеризация веретена деления
Нарушения

Осмолярность
Важной характеристикой среды является ее осмолярность, определяемая концентрацией и константами
Осмолярность
Важной характеристикой среды является ее осмолярность, определяемая концентрацией и константами

Как обеспечить стабильность среды культивирования
Использовать минеральное масло.
При работе с гаметами и
Как обеспечить стабильность среды культивирования
Использовать минеральное масло.
При работе с гаметами и

О2
Ооцитам и эмбрионам необходим кислород, в условиях in vitro
они генерируют
О2
Ооцитам и эмбрионам необходим кислород, в условиях in vitro
они генерируют

Выводы
Получение ооцита
Трансвагинальная функция фолликулов под ультрозвуковым контролем
Температура 37°С
Стабильный рН-7,4
Качество ооцитов
На способность
Выводы
Получение ооцита
Трансвагинальная функция фолликулов под ультрозвуковым контролем
Температура 37°С
Стабильный рН-7,4
Качество ооцитов
На способность

Классическая методика
экстракорпорального
оплодотворения
Классическая методика
экстракорпорального
оплодотворения

Подготовка сперматозоидов к оплодотворению
Активация спермиев в придатках яичка (эпидидимусе) –
Подготовка сперматозоидов к оплодотворению
Активация спермиев в придатках яичка (эпидидимусе) –
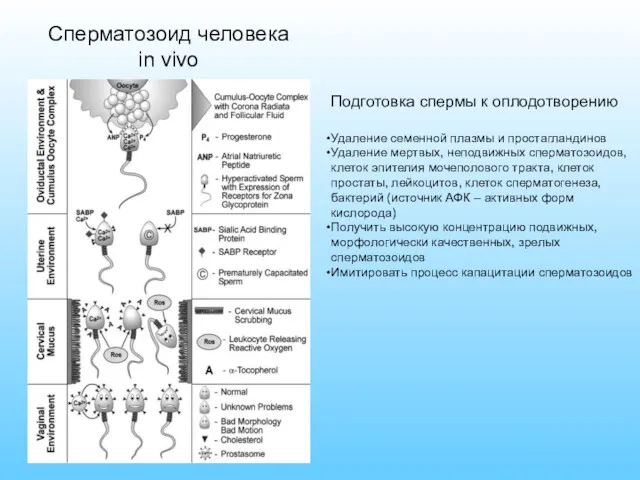
Сперматозоид человека in vivo
Подготовка спермы к оплодотворению
Удаление семенной плазмы и простагландинов
Удаление
Сперматозоид человека in vivo
Подготовка спермы к оплодотворению
Удаление семенной плазмы и простагландинов
Удаление

Капацитация
Биохимически капацитация — это изменение строения некоторых молекул (стеролов, липидов и
Капацитация
Биохимически капацитация — это изменение строения некоторых молекул (стеролов, липидов и

Спермий в половых путях самки.
Схема оплодотворения.
Капацитация
Гиперактивация
Преодоление 3-х естественных барьеров:
клетки
Спермий в половых путях самки.
Схема оплодотворения.
Капацитация
Гиперактивация
Преодоление 3-х естественных барьеров:
клетки
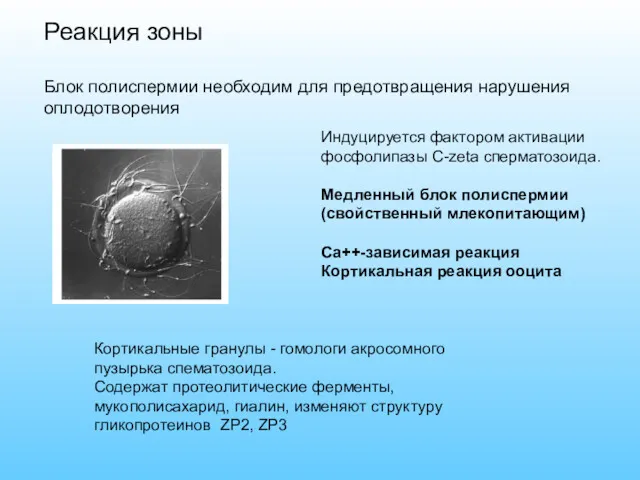
Реакция зоны
Блок полиспермии необходим для предотвращения нарушения оплодотворения
Индуцируется фактором активации фосфолипазы
Реакция зоны
Блок полиспермии необходим для предотвращения нарушения оплодотворения
Индуцируется фактором активации фосфолипазы
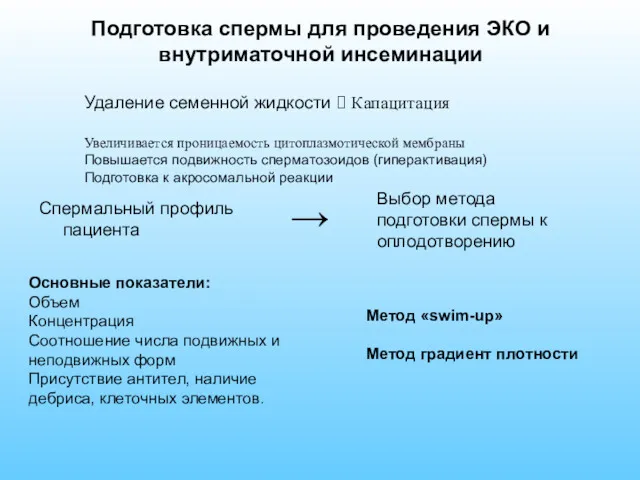
Подготовка спермы для проведения ЭКО и внутриматочной инсеминации
Спермальный профиль пациента
Выбор метода
Подготовка спермы для проведения ЭКО и внутриматочной инсеминации
Спермальный профиль пациента
Выбор метода

Метод «swim-up»
Принцип метода основан на различной подвижности сперматозоидов
Критерии выбора метода:
- нормозооспрмия
-
Метод «swim-up»
Принцип метода основан на различной подвижности сперматозоидов
Критерии выбора метода:
- нормозооспрмия
-

Метод «градиент плотности»
Критерии выбора метода:
- олиго-, астено-, тератозооспермия
- присутствие значительной клеточной
Метод «градиент плотности»
Критерии выбора метода:
- олиго-, астено-, тератозооспермия
- присутствие значительной клеточной

Подготовка спермы к проведению ИКСИ
Критерии выбора:
- олигоспермия, объем ≤ 0,5 мл
Подготовка спермы к проведению ИКСИ
Критерии выбора:
- олигоспермия, объем ≤ 0,5 мл

Инсеменация ООЦИТОВ in vitro
Фактор времени
Тригер овуляции (ХГЧ)→36 часов→пункция → →
Инсеменация ООЦИТОВ in vitro
Фактор времени
Тригер овуляции (ХГЧ)→36 часов→пункция → →

Инсеменация ООЦИТОВ in vitro
Количественный фактор
Культивирование в большом объеме
Количество прогрессивно подвижных сперматозоидов
обычно
Инсеменация ООЦИТОВ in vitro
Количественный фактор
Культивирование в большом объеме
Количество прогрессивно подвижных сперматозоидов
обычно

Сперматозоид
капацитация
гиперактивация
связывание с zona pellucida
акросомный экзоцитоз
проникновение сквозь zona pellucida
сплавление мембран спермия и
Сперматозоид
капацитация
гиперактивация
связывание с zona pellucida
акросомный экзоцитоз
проникновение сквозь zona pellucida
сплавление мембран спермия и
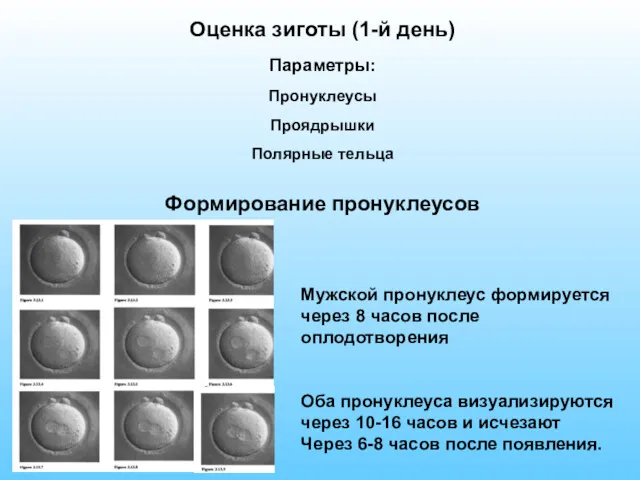
Оценка зиготы (1-й день)
Параметры:
Пронуклеусы
Проядрышки
Полярные тельца
Формирование пронуклеусов
Мужской пронуклеус формируется через 8 часов
Оценка зиготы (1-й день)
Параметры:
Пронуклеусы
Проядрышки
Полярные тельца
Формирование пронуклеусов
Мужской пронуклеус формируется через 8 часов
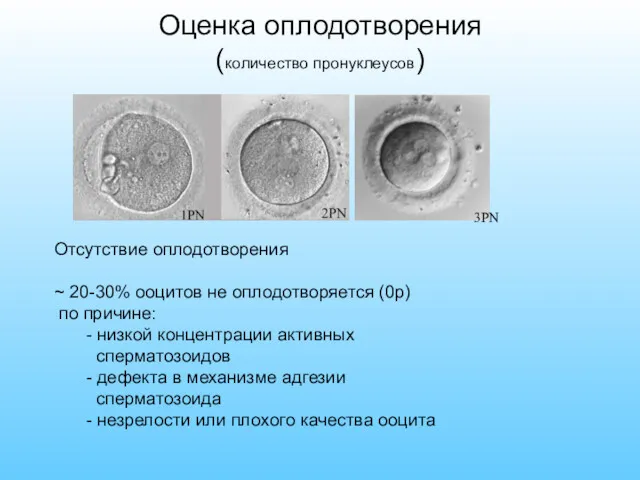
Оценка оплодотворения
(количество пронуклеусов)
3PN
Отсутствие оплодотворения
~ 20-30% ооцитов не оплодотворяется (0р)
по причине:
-
Оценка оплодотворения
(количество пронуклеусов)
3PN
Отсутствие оплодотворения
~ 20-30% ооцитов не оплодотворяется (0р)
по причине:
-

Качество пронуклеарных зигот, их морфология
Форма зиготы
Цитоплазматическое гало (органеллы клетки: митохондрии смещены
Качество пронуклеарных зигот, их морфология
Форма зиготы
Цитоплазматическое гало (органеллы клетки: митохондрии смещены
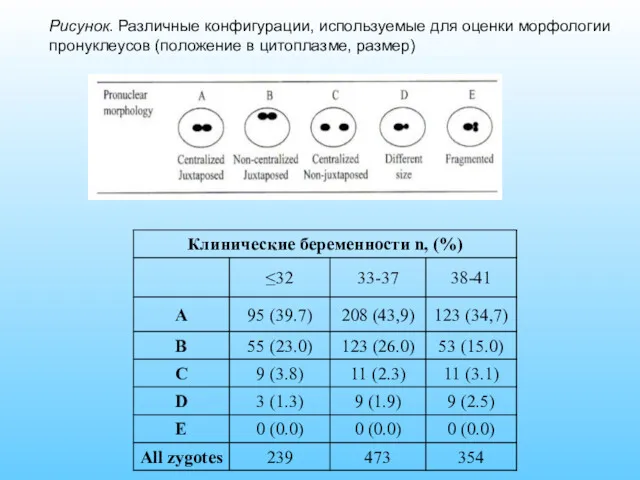
Рисунок. Различные конфигурации, используемые для оценки морфологии пронуклеусов (положение в цитоплазме,
Рисунок. Различные конфигурации, используемые для оценки морфологии пронуклеусов (положение в цитоплазме,

Рисунок.Ориентация пронуклеусов
относительно PBII
Figure 2: Computer-dependent measurements of morphological features
in zygotes.
Оолемма (зеленый),
Рисунок.Ориентация пронуклеусов
относительно PBII
Figure 2: Computer-dependent measurements of morphological features
in zygotes.
Оолемма (зеленый),

Classification of PNs by Tesarik et al., 1999
Zygotes classified according to
Classification of PNs by Tesarik et al., 1999
Zygotes classified according to
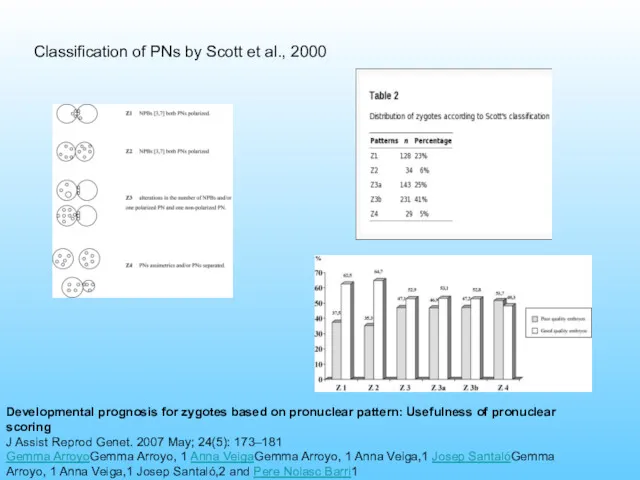
Classification of PNs by Scott et al., 2000
Developmental prognosis for zygotes based
Classification of PNs by Scott et al., 2000
Developmental prognosis for zygotes based
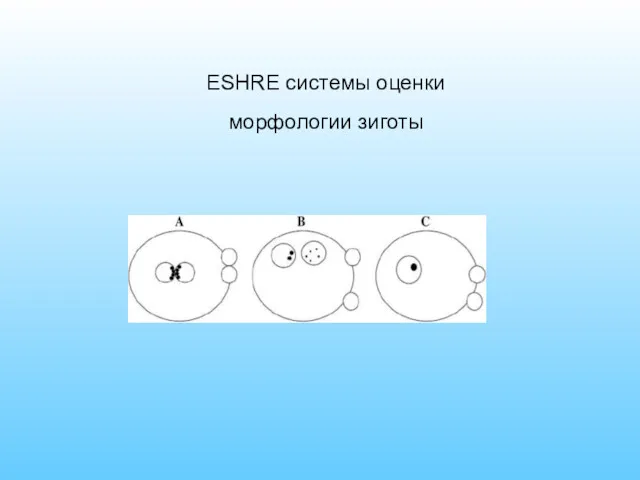
ESHRE системы оценки
морфологии зиготы
ESHRE системы оценки
морфологии зиготы
 Фармацевтична опіка при симптоматичному лікуванні болю у горлі, кашлю, лихоманки
Фармацевтична опіка при симптоматичному лікуванні болю у горлі, кашлю, лихоманки Вскармливание детей первого года жизни
Вскармливание детей первого года жизни Prophylaxis as the main task of a family doctor
Prophylaxis as the main task of a family doctor Патофизиология опухолевого роста (механизмы канцерогенеза)
Патофизиология опухолевого роста (механизмы канцерогенеза) Применение методов статистического анализа для оценки состояния общественного здоровья и здравоохранения (тема № 1)
Применение методов статистического анализа для оценки состояния общественного здоровья и здравоохранения (тема № 1) ВИЧ-инфекция. Общие положения. Истинные и ложные пути передачи. Диагностика. Меры профилактики
ВИЧ-инфекция. Общие положения. Истинные и ложные пути передачи. Диагностика. Меры профилактики Пневмония у детей
Пневмония у детей Організація фармацевтичного забезпечення населення в умовах медичного страхування. Організація проведення аудиторської перевірки
Організація фармацевтичного забезпечення населення в умовах медичного страхування. Організація проведення аудиторської перевірки Охрана материнства и детства
Охрана материнства и детства Основы микрохирургии
Основы микрохирургии Болевой синдром в онкологии
Болевой синдром в онкологии Гестационный сахарный диабет
Гестационный сахарный диабет Медицинская и санитарная техника
Медицинская и санитарная техника Пероральные лекарственные формы с модифицированным высвобождением
Пероральные лекарственные формы с модифицированным высвобождением Профилактика инфекционных заболеваний
Профилактика инфекционных заболеваний Органы кроветворения и иммунной защиты
Органы кроветворения и иммунной защиты Речевые нарушения у детей
Речевые нарушения у детей Удаление зубов
Удаление зубов Диспепсия. Классификация
Диспепсия. Классификация Вступление в систему непрерывного медицинского и фармацевтического образования. Пошаговые инструкции
Вступление в систему непрерывного медицинского и фармацевтического образования. Пошаговые инструкции Экстрапирамидная система
Экстрапирамидная система Кардиомиопатия такоцубо: уникальная кардиомиопатия с переменной морфологией желудочка
Кардиомиопатия такоцубо: уникальная кардиомиопатия с переменной морфологией желудочка Афазия. Характеристика семантической афазии
Афазия. Характеристика семантической афазии Лицензирование медицинской деятельности
Лицензирование медицинской деятельности Анемии. Клиника, диагностика, лечение
Анемии. Клиника, диагностика, лечение Мастер-класс. Травматологический конкурс
Мастер-класс. Травматологический конкурс Кариес и его профилактика
Кариес и его профилактика Порядок проведения диспансеризации определенных групп взрослого населения
Порядок проведения диспансеризации определенных групп взрослого населения