- Гипоталамо-гипофизарная система. Стресс

Содержание
- 2. Гипоталамо-гипофизарная система — объединение структур гипофиза и гипоталамуса, выполняющее функции как нервной системы, так и эндокринной.
- 3. Гипофиз (питуитарная железа) расположен ниже гипоталамуса в турецком седле клиновидной кости основания черепа и состоит из
- 4. Клетки гипоталамуса обладают двойной функцией. Во-первых, они выполняют те же функции, что и любая другая нервная
- 5. Гормоны гипоталамо-гипофизарной системы Гипоталамические гормоны – либерины и статины, которые регулируют гормонпродуцирующую функцию гипофиза. Тиреолиберин –
- 6. Эти гормоны, синтезированные гипоталамусом, поступают в особую кровеносную систему, связывающую гипоталамус с передней долей гипофиза. Два
- 7. Гипофиз расположен на основании головного мозга и прикрепляется к мозгу тонким стеблем. По этому стеблю гипофиз
- 8. Гормоны, которые производит передняя доля гипофиза: Пролактин. Этот гормон стимулирует лактацию (образование материнского молока в молочных
- 9. Вазопрессин (АДГ, антидиуретический гормон) Синтез и выделение АДГ рефлекторно усиливаются при повышении осмотического давления крови более
- 10. Окситоцин Окситоцин— пептид, состоящий из 9 аминокислотных остатков. Он транспортируется в свободной форме, период полураспада 5—10
- 11. Меланоцитстимулирующий гормон У взрослого человека эта зона редуцирована и МСГ вырабатывается очень ограниченно. Его функции в
- 12. Гормон роста (ГР, соматотропин, соматотропный гормон) выделяется в кровь красными ацидофильными сома- тотрофами (50% от клеток
- 13. Пролактин (лактотропный гормон, ЛТГ) выделяется в кровь желтыми лактотрофами (10—25% клеток аденоги- пофиза, а во время
- 14. Тропные гормоны гипофиза.Регулируют функции периферических эндокринных желез и клеток, а также неэндокринных клеток.
- 15. Тиреотропный гормон (тиреотропин, ТТГ) синтезируется тиреотрофами, является сложным гликопротеином, действует через мембранные 7-ТМС-рецепторы (вторичный посредник цАМФ).
- 16. Гонадотропины: ФСГ (фоллитропин) и ЛГ (или лютропин) вырабатываются в одних и тех же базофильных клетках (гонадотрофах)
- 17. Адренокортикотропный гормон синтезируется кортикотрофами аденогипофиза, является пептидом (состоит из 39 аминокислотных остатков, первые 13 из которых
- 18. Основные группы биорегуляторов
- 19. Либерины (то есть нейроэндокринные стимуляторы продукции гипофизарных пептидов с идентифицированной структурой) и рилизинг-факторы (стимуляторы продукции гипофизарных
- 20. Статины (пептиды с установленной химической структурой, нейроэндокринным путём тормозящие продукцию гипофизарных гормонов) и ингибитинг-факторы (аналогичные пептиды,
- 21. Стресс
- 22. Стресс Стресс — совокупность всех неспецифических изменений, возникающих в организме под влиянием любых воздействий и включающих,
- 24. Первая стадия синдрома (реакция тревоги) характеризует остро протекающую, активную мобилизацию адаптационных процессов в организме в ответ
- 26. Гипофизэктомия или разрушение области срединного возвышения гипоталамуса снимает реакцию адреналовой коры на стресс. Эти воздействия, а
- 28. Скачать презентацию
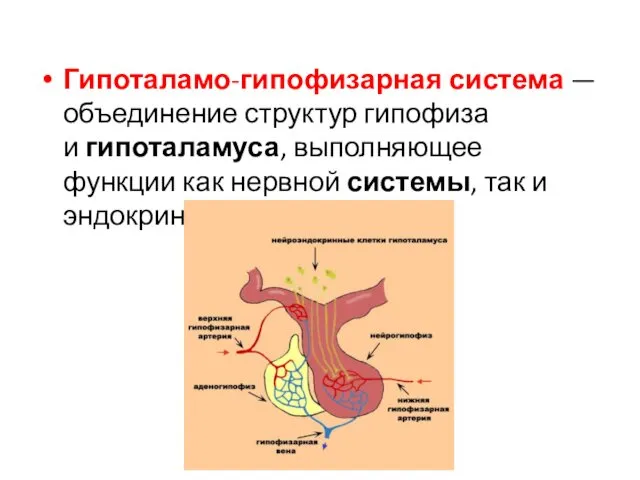
Гипоталамо-гипофизарная система — объединение структур гипофиза и гипоталамуса, выполняющее функции как нервной системы, так
Гипоталамо-гипофизарная система — объединение структур гипофиза и гипоталамуса, выполняющее функции как нервной системы, так

Гипофиз (питуитарная железа) расположен ниже гипоталамуса в турецком седле клиновидной
Гипофиз (питуитарная железа) расположен ниже гипоталамуса в турецком седле клиновидной

Клетки гипоталамуса обладают двойной функцией. Во-первых, они выполняют те же функции,
Клетки гипоталамуса обладают двойной функцией. Во-первых, они выполняют те же функции,

Гормоны гипоталамо-гипофизарной системы
Гипоталамические гормоны – либерины и статины, которые регулируют гормонпродуцирующую функцию
Гормоны гипоталамо-гипофизарной системы
Гипоталамические гормоны – либерины и статины, которые регулируют гормонпродуцирующую функцию
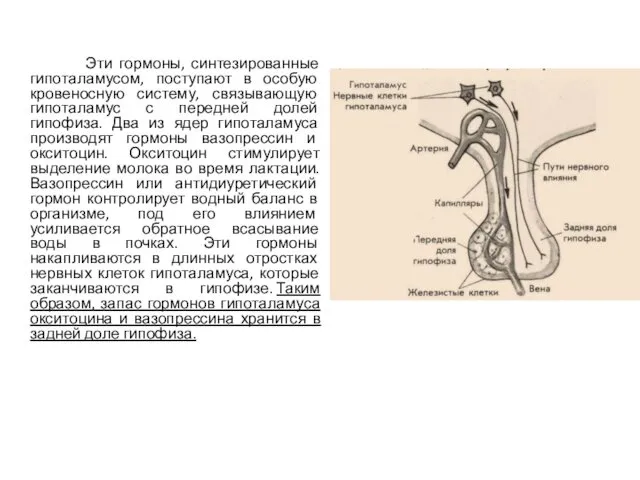
Эти гормоны, синтезированные гипоталамусом, поступают в особую кровеносную систему, связывающую
Эти гормоны, синтезированные гипоталамусом, поступают в особую кровеносную систему, связывающую
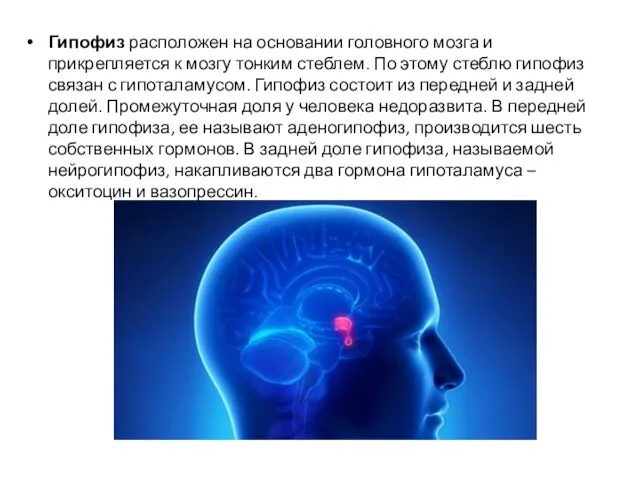
Гипофиз расположен на основании головного мозга и прикрепляется к мозгу тонким стеблем.
Гипофиз расположен на основании головного мозга и прикрепляется к мозгу тонким стеблем.

Гормоны, которые производит передняя доля гипофиза:
Пролактин. Этот гормон стимулирует лактацию (образование
Гормоны, которые производит передняя доля гипофиза:
Пролактин. Этот гормон стимулирует лактацию (образование

Вазопрессин (АДГ, антидиуретический гормон)
Синтез и выделение АДГ рефлекторно усиливаются при повышении
Вазопрессин (АДГ, антидиуретический гормон)
Синтез и выделение АДГ рефлекторно усиливаются при повышении

Окситоцин
Окситоцин— пептид, состоящий из 9 аминокислотных остатков. Он транспортируется в свободной
Окситоцин
Окситоцин— пептид, состоящий из 9 аминокислотных остатков. Он транспортируется в свободной

Меланоцитстимулирующий гормон
У взрослого человека эта зона редуцирована и МСГ вырабатывается очень
Меланоцитстимулирующий гормон
У взрослого человека эта зона редуцирована и МСГ вырабатывается очень

Гормон роста (ГР, соматотропин, соматотропный гормон)
выделяется в кровь красными ацидофильными сома-
Гормон роста (ГР, соматотропин, соматотропный гормон)
выделяется в кровь красными ацидофильными сома-

Пролактин (лактотропный гормон, ЛТГ)
выделяется в кровь желтыми лактотрофами (10—25% клеток аденоги- пофиза,
Пролактин (лактотропный гормон, ЛТГ)
выделяется в кровь желтыми лактотрофами (10—25% клеток аденоги- пофиза,

Тропные гормоны гипофиза.Регулируют функции периферических эндокринных желез и клеток, а также
Тропные гормоны гипофиза.Регулируют функции периферических эндокринных желез и клеток, а также

Тиреотропный гормон (тиреотропин, ТТГ)
синтезируется тиреотрофами, является сложным гликопротеином, действует через мембранные
Тиреотропный гормон (тиреотропин, ТТГ)
синтезируется тиреотрофами, является сложным гликопротеином, действует через мембранные

Гонадотропины: ФСГ (фоллитропин) и ЛГ (или лютропин)
вырабатываются в одних и тех
Гонадотропины: ФСГ (фоллитропин) и ЛГ (или лютропин)
вырабатываются в одних и тех

Адренокортикотропный гормон
синтезируется кортикотрофами аденогипофиза, является пептидом (состоит из 39 аминокислотных остатков,
Адренокортикотропный гормон
синтезируется кортикотрофами аденогипофиза, является пептидом (состоит из 39 аминокислотных остатков,

Основные группы биорегуляторов
Основные группы биорегуляторов

Либерины (то есть нейроэндокринные стимуляторы продукции гипофизарных пептидов с идентифицированной структурой) и рилизинг-факторы (стимуляторы
Либерины (то есть нейроэндокринные стимуляторы продукции гипофизарных пептидов с идентифицированной структурой) и рилизинг-факторы (стимуляторы

Статины (пептиды с установленной химической структурой, нейроэндокринным путём тормозящие продукцию гипофизарных гормонов)
Статины (пептиды с установленной химической структурой, нейроэндокринным путём тормозящие продукцию гипофизарных гормонов)

Стресс
Стресс

Стресс
Стресс — совокупность всех неспецифических изменений, возникающих в организме под влиянием
Стресс
Стресс — совокупность всех неспецифических изменений, возникающих в организме под влиянием
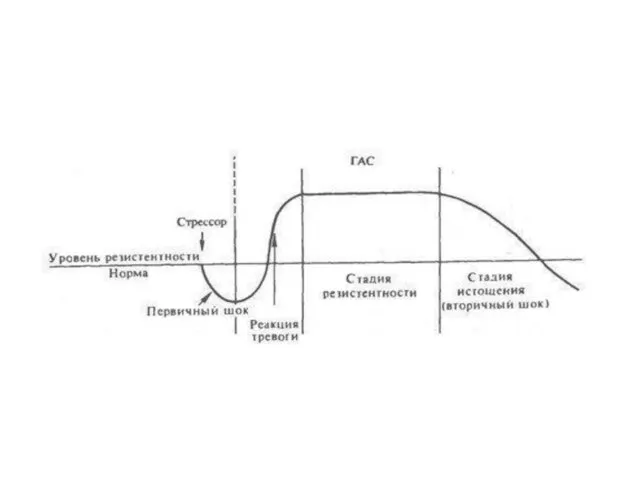

Первая стадия синдрома (реакция тревоги) характеризует остро протекающую, активную мобилизацию адаптационных
Первая стадия синдрома (реакция тревоги) характеризует остро протекающую, активную мобилизацию адаптационных
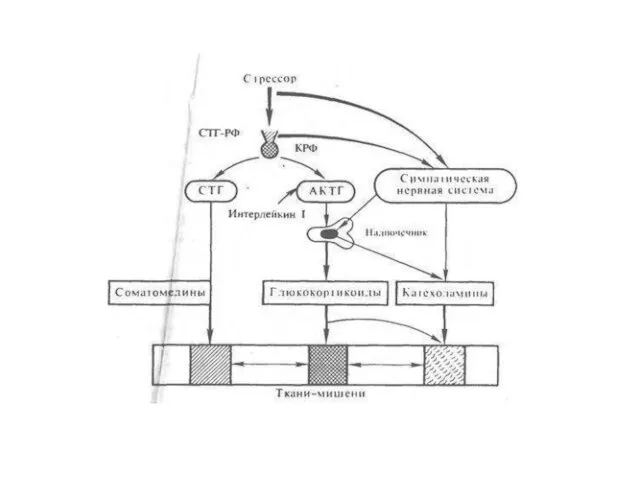

Гипофизэктомия или разрушение области срединного возвышения гипоталамуса снимает реакцию адреналовой коры
Гипофизэктомия или разрушение области срединного возвышения гипоталамуса снимает реакцию адреналовой коры
 Наследственные болезни обмена веществ: клиника, диагностика, лечение
Наследственные болезни обмена веществ: клиника, диагностика, лечение Красный плоский лишай
Красный плоский лишай Заболевания СОПР. Воспалительные заболевания челюстно-лицевой области
Заболевания СОПР. Воспалительные заболевания челюстно-лицевой области Асептика. Антисептика
Асептика. Антисептика Особенности эпидемического процесса при сибирской язве
Особенности эпидемического процесса при сибирской язве Pathologie de la cornée
Pathologie de la cornée Железы внутренней секреции
Железы внутренней секреции История хирургии. Асптика. Антисептика. (лекция 1). (1)
История хирургии. Асптика. Антисептика. (лекция 1). (1) Дифференциальная диагностика носовых кровотечений
Дифференциальная диагностика носовых кровотечений Эхокардиография в диагностике ИБС
Эхокардиография в диагностике ИБС Воспаление. Факторы, вызывающие воспаление
Воспаление. Факторы, вызывающие воспаление Балалардағы дермо-респираторлық синдром
Балалардағы дермо-респираторлық синдром Лучевое исследование костей и суставов
Лучевое исследование костей и суставов Туберкулезді менингит
Туберкулезді менингит Бұлшықет тіндері
Бұлшықет тіндері Бронхіальна астма. (Лекція 4)
Бронхіальна астма. (Лекція 4) Комплексный подход к лечению миопии
Комплексный подход к лечению миопии ботулизм
ботулизм Эпидемиологическая ситуация по гриппу, ОРВИ и внебольничным пневмониям среди населения Новосибирской области
Эпидемиологическая ситуация по гриппу, ОРВИ и внебольничным пневмониям среди населения Новосибирской области Терінің вирусты аурулары
Терінің вирусты аурулары Болезнь Ауески
Болезнь Ауески Острые респираторные вирусные инфекции у детей
Острые респираторные вирусные инфекции у детей Сүйек. Эктопиялық дамуы
Сүйек. Эктопиялық дамуы Кровь. Состав крови, строение и функции форменных элементов крови
Кровь. Состав крови, строение и функции форменных элементов крови Электрофорез. Электротерапия
Электрофорез. Электротерапия ВВЕДЕНИЕ В ПСИХИАТРИЮ
ВВЕДЕНИЕ В ПСИХИАТРИЮ Функциональная диспепсия: клиника и лечение
Функциональная диспепсия: клиника и лечение Острые пневмонии, ХНЗЛ, пневмоканиозы. Макропрепараты
Острые пневмонии, ХНЗЛ, пневмоканиозы. Макропрепараты