- Иммунопоэз: созревание Т - и В - клеточных рецепторов

Содержание
- 2. ЦЕНТРАЛЬНЫЕ ОРГАНЫ ИММУНИТЕТА Центральные органы иммунитета - красный костный мозг и тимус. В центральных органах иммунитета
- 3. ЦЕНТРАЛЬНЫЕ ОРГАНЫ ИММУНИТЕТА Клетки становятся иммунокомпетентными – то есть способными различать разные молекулы чужеродных структур. Эта
- 4. Центральные органы иммунитета: тимус тимус
- 5. ТИМУС Дольчатая структура с эпителиальными клетками стромы и соединительной тканью Строма обеспечивает микроокружение для развития и
- 6. Клетки коры и мозгового вещества тимуса
- 7. Тимус - биологические часы: масса тимуса
- 8. Тимус - биологические часы Для пожилых характерно: большое число клеток памяти ( встречи со многими антигенами)
- 9. Созревание Т-лимфоцитов в тимусе: 1 этап
- 10. Созревание Т-лимфоцитов в тимусе: 2 этап
- 11. ЦЕНТРАЛЬНЫЕ ОРГАНЫ ИММУНИТЕТА: положительная и отрицательная селекция (отбор) клеток В центральных органах иммунитета происходят процессы селекции
- 12. Интенсивность отбора тимоцитов T клетки созревают в тимусе, но гораздо больше Т- клеток погибает в тимусе
- 13. T клетки созревают в тимусе, но гораздо больше Т- клеток погибает 98% клеток погибает в тимусе
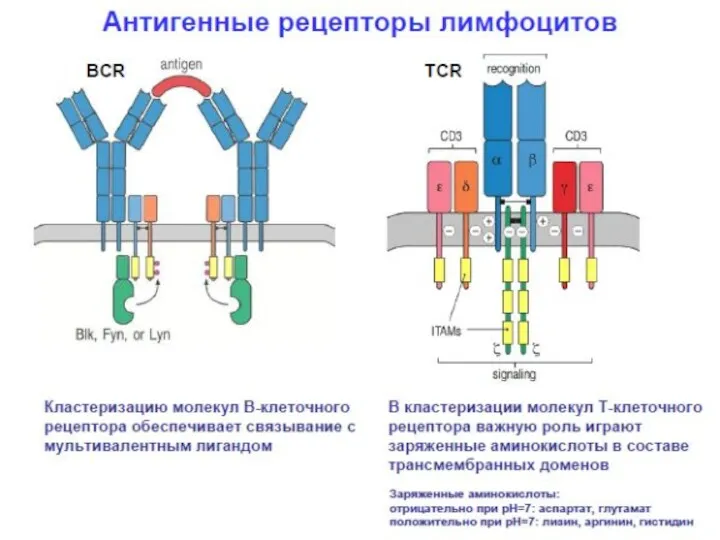
- 14. Строение Т рецепторов Т клеточный рецептор имеет α и β -цепи (есть альтернативные рецепторы, которые имеют
- 15. Т-клеточный рецептор Vα Vβ Cα Cβ карбогидраты моновалентность для сравнения:BCR - Ig Fab фрагмент Нет альтернативных
- 16. Какие клетки выходят на периферию из тимуса? В результате позитивной и негативной селекции в кровоток поступают
- 17. Строение Т рецепторного комплекса (TCR/ CD3) На клеточной поверхности αβ -Т клеточный рецептор (или γδ) расположен
- 19. Строение корецепторов (CD 4 или CD8) Корецепторы (CD 4 или CD8) расположены на мембране Т лимфоцита
- 20. Клетка-мишень Антигенпрезентирующая клетка
- 21. Процесс созревания тимоцитов: этапы формирования корецепторов Корецепторы: CD4 – распознает молекулы МНС II CD8 – распознает
- 23. Сигнал с CD4 отменяет экспрессию CD8 и наоборот ДВОЙНЫЕ ПОЗИТИВНЫЕ ТИМОЦИТЫ Эпителий тимуса Переход от двойных
- 24. Реаранжировка генов T-клеточных рецепторов При «созревании» рецепторов Т лимфоцитов в тимусе гены α- β- или γ-
- 25. Реаранжировка генов, кодирующих β -цепь Т рецептора Изначальная конфигурация генов, кодирующих β цепь: эти гены находятся
- 26. Реаранжировка генов Тклеточного рецептора (TCR) Рекомбинация ДНК происходит при объединении V-, D- и J-сегментов, катализируется комплексом
- 27. Реаранжировка генов, кодирующих Т рецептор, путем соматической рекомбинации 1 этап – слияние генов D-J 2 этап
- 28. Реаранжировка генов, кодирующих α -цепь Т рецептора, сборка Т рецептора При реаранжировке генов α -цепи Т
- 29. Каким образом развивается аутотолерантность к антигенам, отсутствующим в тимусе? T клетки, несущие TcR , и вступающие
- 30. Костимуляция (иммунный ответ) ; отсутствие костимуляции (анергия, толерантность). иммунного ответа). Процесс презентации антигенов сопровождается или не
- 31. Ко- стимулирующие молекулярные взаимодействия на АПК и Тлимфоците: CD40-CD 40L и комплекс B7-CD 28
- 32. Гипотезы повреждения и костимуляции Полная экспрессия функций Т лимфоцитов зависит от того, когда и где экспрессируются
- 33. Гипотеза угрозы АПК , определившие сигналы угрозы, экспрессируют костимулирующие молекулы, активируют Т – клетки и иммунный
- 34. Зрелые Т-лимфоциты: жизненный путь на периферии Выжившие в результате положительной и отрицательной селекции Т-клетки выходят из
- 35. Зрелые Т-лимфоциты: жизненный путь на периферии После встречи с антигеном в лимфоузле Т-клетка с помощью цитокинов,
- 36. Эмиграция эффекторной Т-клетки в ткань при вирусной инфекции
- 37. Зрелые Т-лимфоциты: рециркулирующие и резидентные Лимфоциты рециркулируют по лимфе и кровотоку в поисках антигена, который нужно
- 38. ОТКРЫТИЕ В - КЛЕТОЧНОГО ИММУНИТЕТА 1954 - Bruce Glick, США Изучение функции Фабрициевой сумки (bursa Fabricius),
- 39. Происхождение В клеток и орган, в котором созревают В-клетки После рождения их развитие продолжается в костном
- 40. Костный мозг
- 41. Стадии развития В лимфоцита 1). Стволовая клетка 2) Общий лимфоидный предшественник для B- и T-клеточного пути
- 42. Стадии развития В клеток На каждой стадии развития происходит реаранжировка генов тяжелых и легких цепей Ig,
- 43. Стадии развития В клеток Ранние этапы развития В-лимфоцитов зависят от прямого контактного взаимодействия со стромой. В
- 44. Стадии развития В клеток В дальнейшем ранние пре-В-клетки, трансформируются в малые пре-В-лимфоциты, у некоторых из которых
- 45. Развитие В клеток в костном мозге Более 75% созревающих в костном мозге В-клеток не попадает в
- 46. Развитие В клеток в костном мозге: «созревание» моноспецифичных рецепторов, «+» и «-» селекция Костный мозг обеспечивает
- 47. Клетки стромы Созревающие В клетки
- 48. B B Клетка стромы
- 49. X X X Схема развития В – клеток в костном мозге
- 50. Y Y Y Y Y Y Зрелый В-лимфоцит выходит на периферию Y Y В - клеточная
- 51. Постулаты теории клональной селекции Каждый В лимфоцит имеет рецептор уникальной специфичности. Высокоаффинное (прочное) взаимодействие рецептора с
- 53. Гены молекул иммуноглобулинов (Ig) Каждая молекула Ig состоит из 2 тяжелых (Н) и двух легких (L)
- 54. Стадии дифференцировки определяются реаранжировкой генов Ig Стадии созревания конфигурация генов IgH Стволовая клетка Ранний про-В Поздний
- 55. Реаранжировка генов, кодирующих легкие цепи молекулы Ig После завершения перестройки (реаранжировки) генов, кодирующих тяжелые цепи молекулы
- 58. Положительная и отрицательная селекция В лимфоцитов в костном мозге + селекция происходит при взаимодействии В -
- 59. Гены Ig молекул До встречи с антигеном: Рекомбинации ограниченного числа генных сегментов V, D и J
- 60. Дальнейшие этапы развития В-лимфоцитов Из костного мозга уже отобранные В-лимфоциты попадают с током крови в первичные
- 62. Выход зрелых В клеток на периферию Из костного мозга выходят только те В –лимфоциты, у которых
- 63. Рециркулирующие В – клетки встречаются со «своим» антигеном в лимфоузле
- 64. Распознал неауто-АГ на периферии Ig - секретирующая плазматическая клетка Дифференцировка В – клеток на периферии Зрелый
- 66. Схема ответа В-лимфоцитов на антиген
- 67. Субпопуляции В лимфоцитов: В1 и В2 В 2 (CD 5-) лимфоциты связывают белковые антигены, им нужна
- 68. Субпопуляция В1 лимфоцитов После активации В1 клетки секретируют анти-полисахаридные антитела класса М (IgM), которые присоединяются к
- 70. Взаимодействие АПК, Т- и В- лимфоцитов в ходе иммунного ответа на АГ
- 71. Вопросы к занятию № 3а Какова роль тимуса в процессе дифференцировки Т-лимфоцитов? Каков биологический смысл положительной
- 72. Тестовые задания к занятию №3 11. Антигеннезависимая дифференцировка Т лимфоцитов происходит в: щитовидной железе тимусе лимфатических
- 73. Тестовые задания к занятию №3 3. Общим маркером Т-лимфоцитов является молекула: CD 3 CD 4 CD
- 74. Тестовые задания к занятию №3 5. Для цитотоксических Т-лимфоцитов характерны маркеры: CD 3 ; CD 4
- 75. Тестовые задания к занятию № 3 7. Зародышевые гены тяжелых цепей иммуноглобулинов включают регионы: B DP
- 77. Скачать презентацию

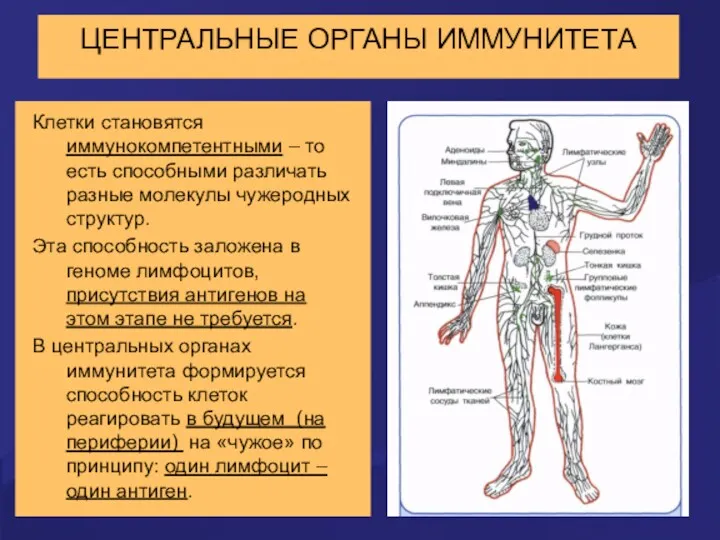
ЦЕНТРАЛЬНЫЕ ОРГАНЫ ИММУНИТЕТА
Центральные органы иммунитета - красный костный мозг и тимус.
ЦЕНТРАЛЬНЫЕ ОРГАНЫ ИММУНИТЕТА
Центральные органы иммунитета - красный костный мозг и тимус.
ЦЕНТРАЛЬНЫЕ ОРГАНЫ ИММУНИТЕТА
Клетки становятся иммунокомпетентными – то есть способными различать разные
ЦЕНТРАЛЬНЫЕ ОРГАНЫ ИММУНИТЕТА
Клетки становятся иммунокомпетентными – то есть способными различать разные
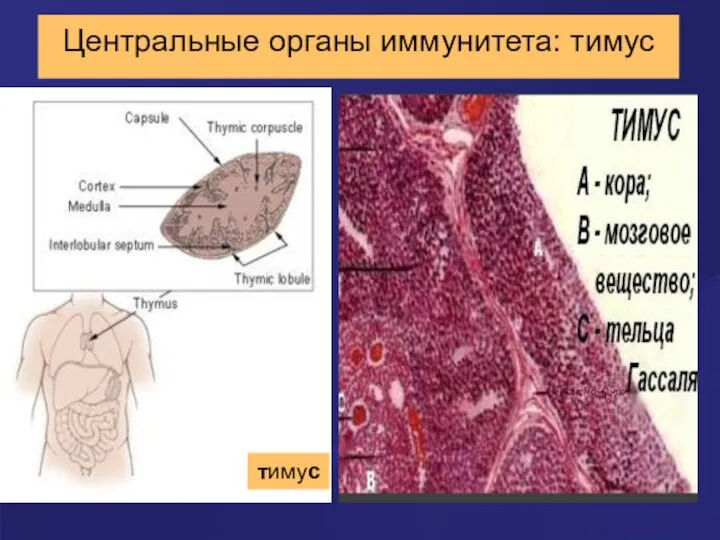
Центральные органы иммунитета: тимус
тимус
Центральные органы иммунитета: тимус
тимус
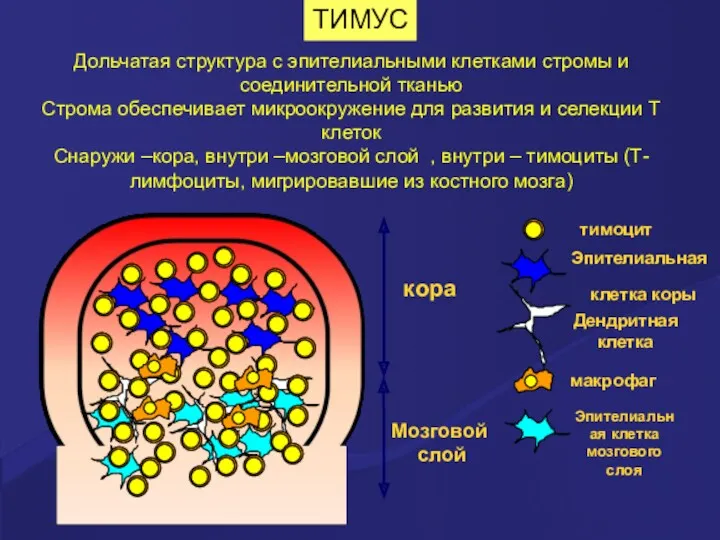
ТИМУС
Дольчатая структура с эпителиальными клетками стромы и соединительной тканью
Строма обеспечивает микроокружение
ТИМУС
Дольчатая структура с эпителиальными клетками стромы и соединительной тканью
Строма обеспечивает микроокружение
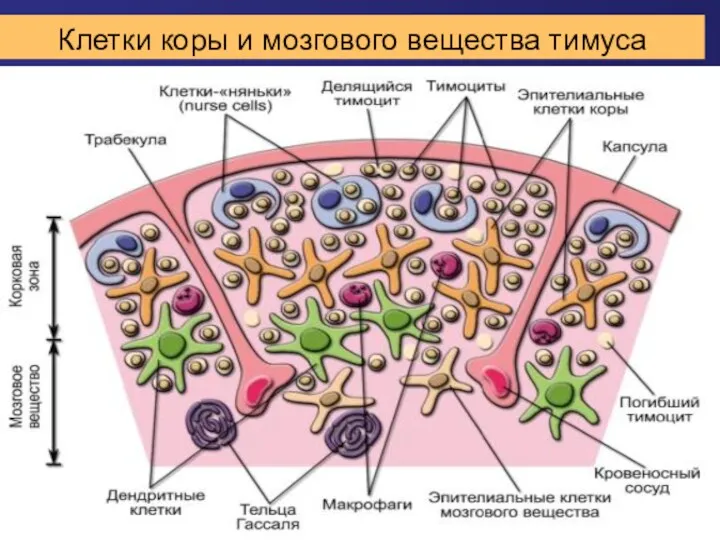
Клетки коры и мозгового вещества тимуса
Клетки коры и мозгового вещества тимуса
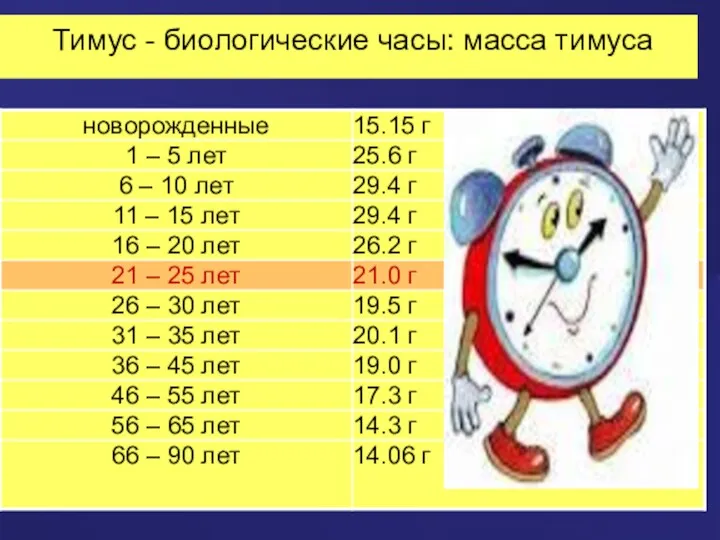
Тимус - биологические часы: масса тимуса
Тимус - биологические часы: масса тимуса
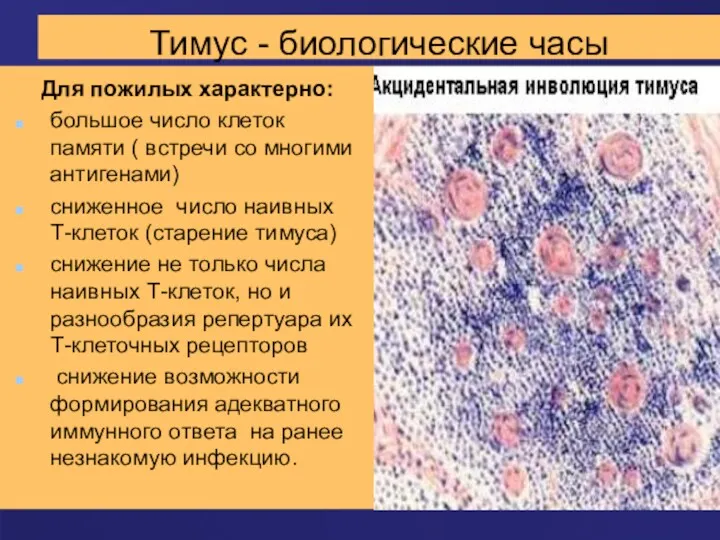
Тимус - биологические часы
Для пожилых характерно:
большое число клеток памяти ( встречи
Тимус - биологические часы
Для пожилых характерно:
большое число клеток памяти ( встречи
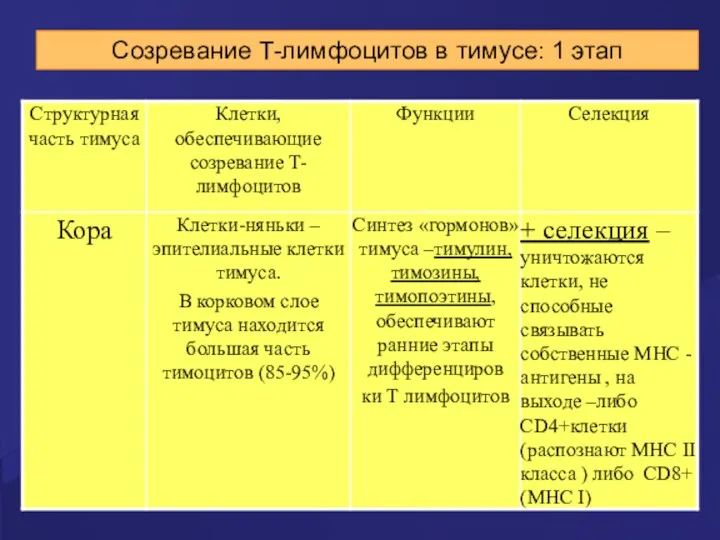
Созревание Т-лимфоцитов в тимусе: 1 этап
Созревание Т-лимфоцитов в тимусе: 1 этап
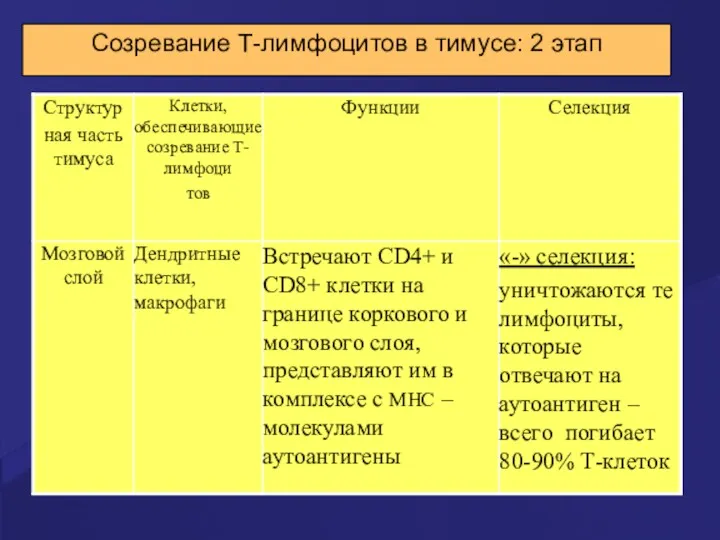
Созревание Т-лимфоцитов в тимусе: 2 этап
Созревание Т-лимфоцитов в тимусе: 2 этап

ЦЕНТРАЛЬНЫЕ ОРГАНЫ ИММУНИТЕТА: положительная и отрицательная селекция (отбор) клеток
В центральных органах
ЦЕНТРАЛЬНЫЕ ОРГАНЫ ИММУНИТЕТА: положительная и отрицательная селекция (отбор) клеток
В центральных органах
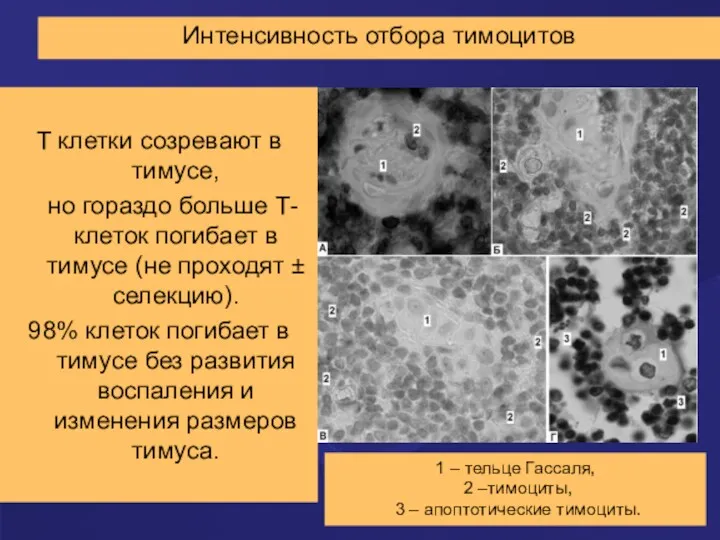
Интенсивность отбора тимоцитов
T клетки созревают в тимусе,
но гораздо больше
Интенсивность отбора тимоцитов
T клетки созревают в тимусе,
но гораздо больше
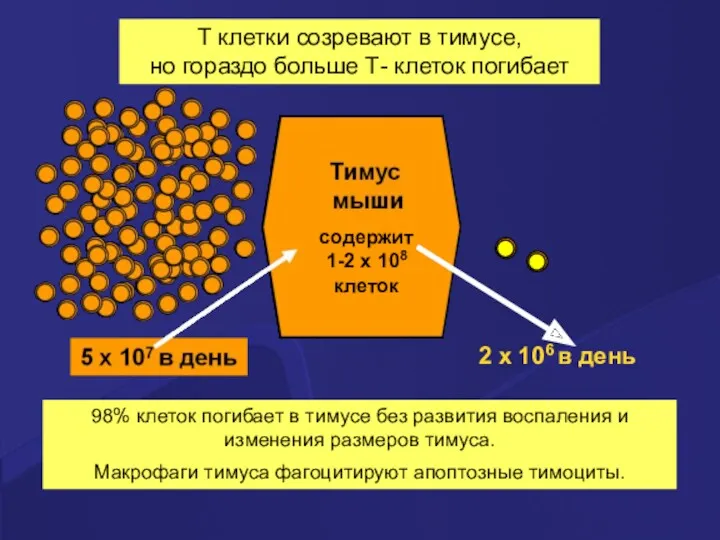
T клетки созревают в тимусе,
но гораздо больше Т- клеток погибает
98%
T клетки созревают в тимусе,
но гораздо больше Т- клеток погибает
98%

Строение Т рецепторов
Т клеточный рецептор имеет α и β -цепи (есть
Строение Т рецепторов
Т клеточный рецептор имеет α и β -цепи (есть
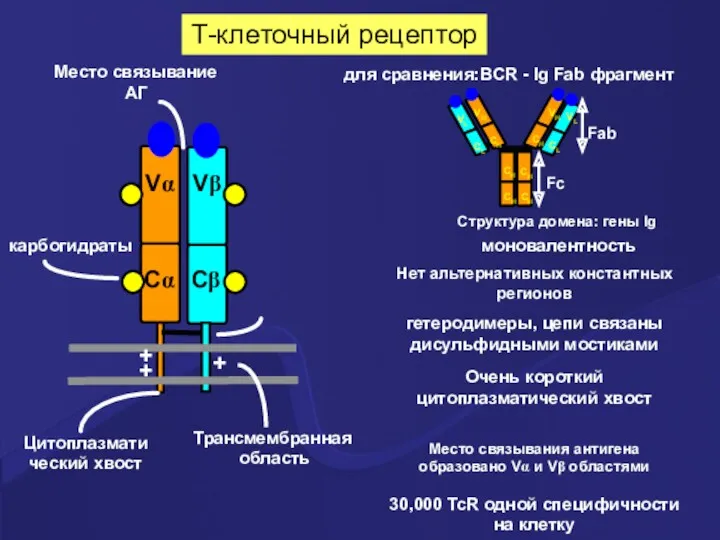
Т-клеточный рецептор
Vα
Vβ
Cα
Cβ
карбогидраты
моновалентность
для сравнения:BCR - Ig Fab фрагмент
Нет альтернативных константных регионов
Структура домена:
Т-клеточный рецептор
Vα
Vβ
Cα
Cβ
карбогидраты
моновалентность
для сравнения:BCR - Ig Fab фрагмент
Нет альтернативных константных регионов
Структура домена:
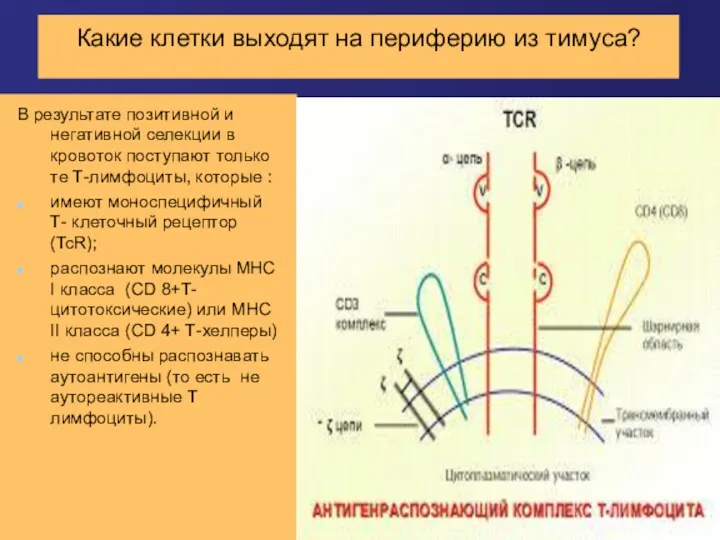
Какие клетки выходят на периферию из тимуса?
В результате позитивной и негативной
Какие клетки выходят на периферию из тимуса?
В результате позитивной и негативной
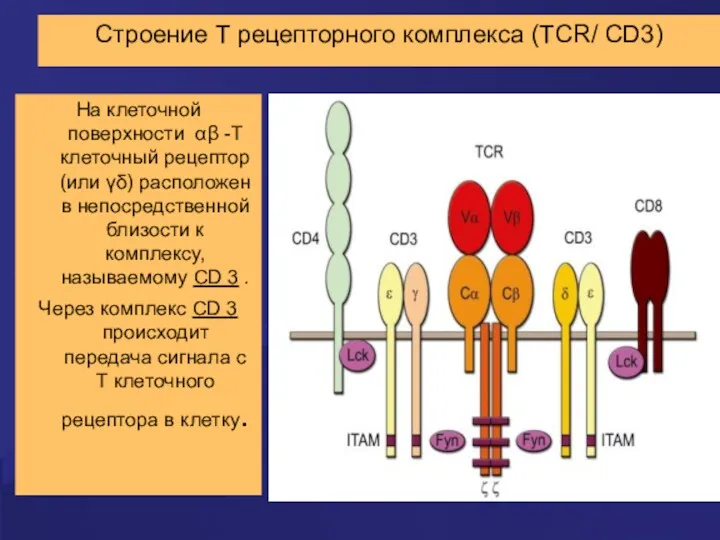
Строение Т рецепторного комплекса (TCR/ CD3)
На клеточной поверхности αβ -Т клеточный
Строение Т рецепторного комплекса (TCR/ CD3)
На клеточной поверхности αβ -Т клеточный
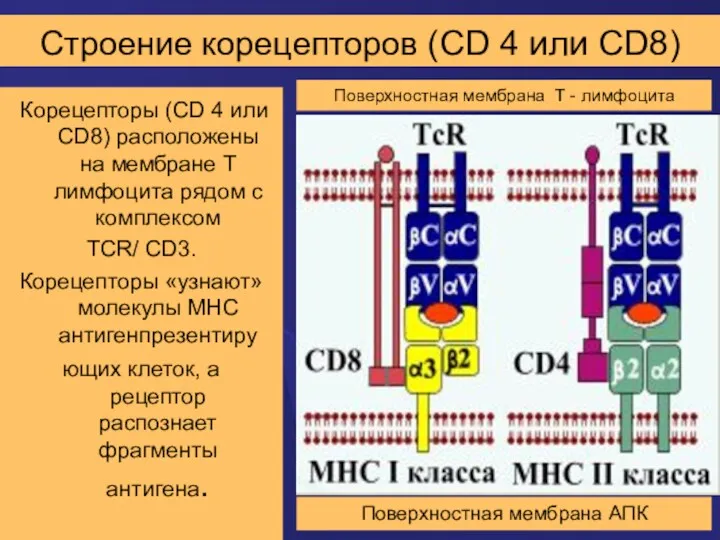
Строение корецепторов (CD 4 или CD8)
Корецепторы (CD 4 или CD8)
Строение корецепторов (CD 4 или CD8)
Корецепторы (CD 4 или CD8)
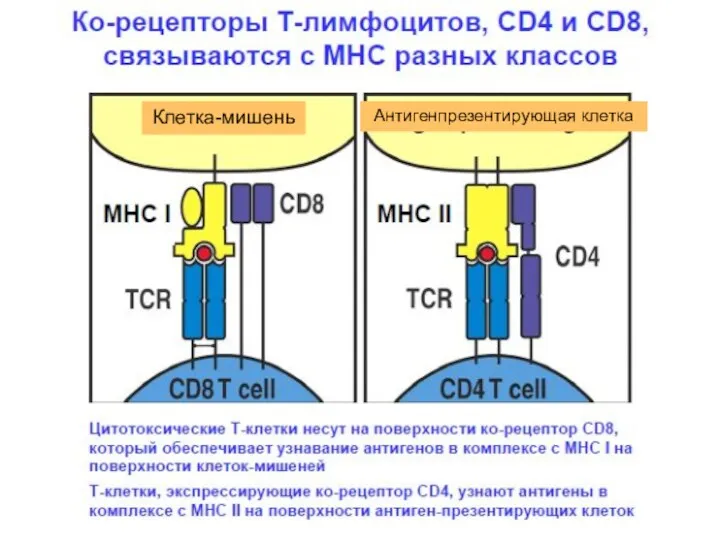
Клетка-мишень
Антигенпрезентирующая клетка
Клетка-мишень
Антигенпрезентирующая клетка

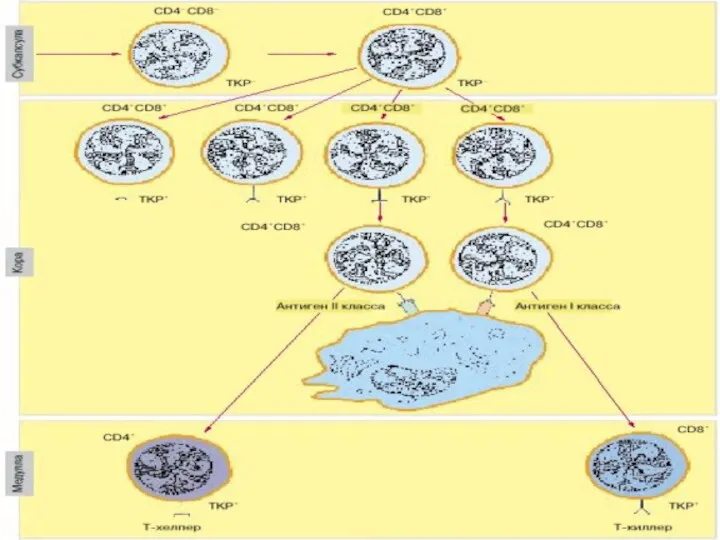
Процесс созревания тимоцитов:
этапы формирования корецепторов
Корецепторы:
CD4 – распознает молекулы МНС II
Процесс созревания тимоцитов:
этапы формирования корецепторов
Корецепторы:
CD4 – распознает молекулы МНС II
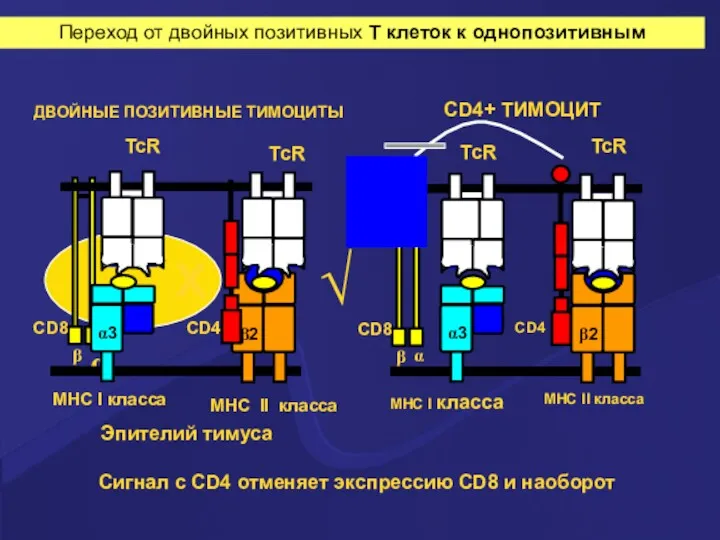
Сигнал с CD4 отменяет экспрессию CD8 и наоборот
ДВОЙНЫЕ ПОЗИТИВНЫЕ ТИМОЦИТЫ
Эпителий тимуса
Переход
Сигнал с CD4 отменяет экспрессию CD8 и наоборот
ДВОЙНЫЕ ПОЗИТИВНЫЕ ТИМОЦИТЫ
Эпителий тимуса
Переход

Реаранжировка генов T-клеточных рецепторов
При «созревании» рецепторов Т лимфоцитов в тимусе
Реаранжировка генов T-клеточных рецепторов
При «созревании» рецепторов Т лимфоцитов в тимусе
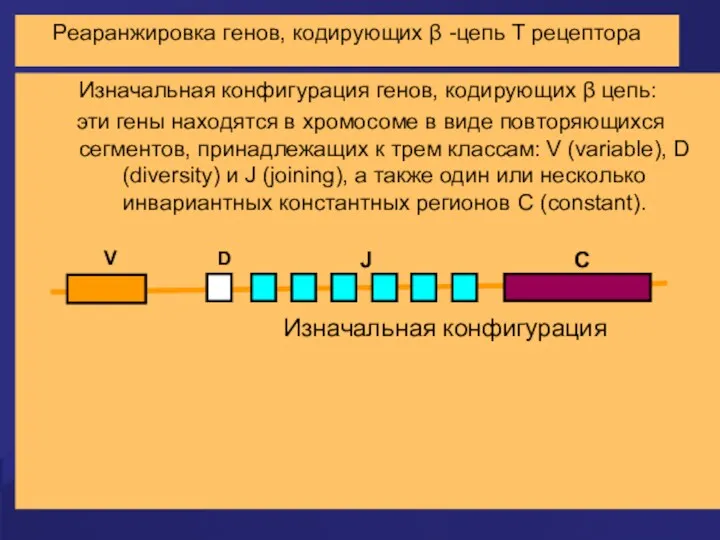
Реаранжировка генов, кодирующих β -цепь Т рецептора
Изначальная конфигурация генов, кодирующих β
Реаранжировка генов, кодирующих β -цепь Т рецептора
Изначальная конфигурация генов, кодирующих β
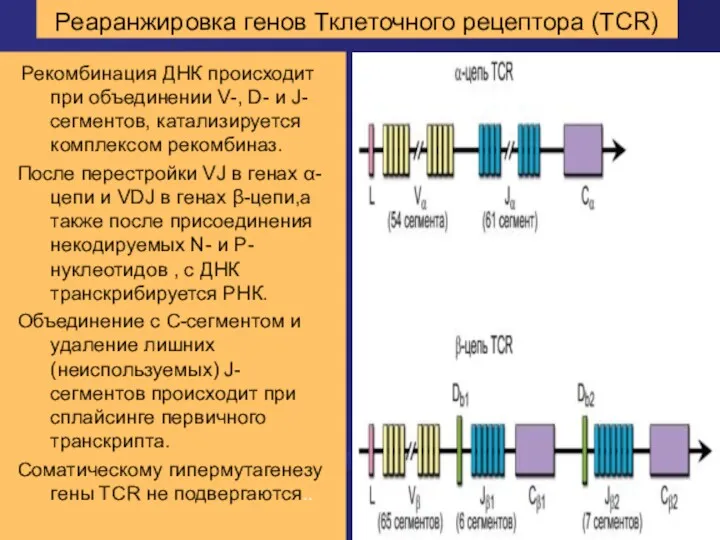
Реаранжировка генов Тклеточного рецептора (TCR)
Рекомбинация ДНК происходит при объединении V-, D-
Реаранжировка генов Тклеточного рецептора (TCR)
Рекомбинация ДНК происходит при объединении V-, D-
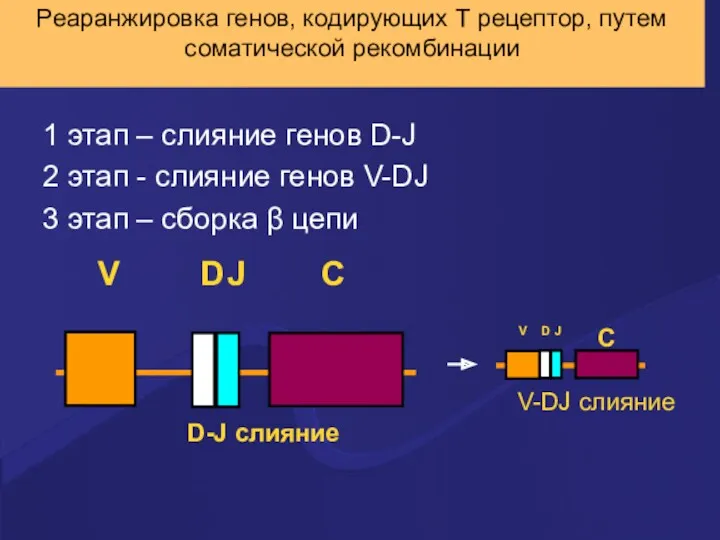
Реаранжировка генов, кодирующих Т рецептор, путем соматической рекомбинации
1 этап – слияние
Реаранжировка генов, кодирующих Т рецептор, путем соматической рекомбинации
1 этап – слияние

Реаранжировка генов, кодирующих α -цепь Т рецептора, сборка Т рецептора
При реаранжировке
Реаранжировка генов, кодирующих α -цепь Т рецептора, сборка Т рецептора
При реаранжировке

Каким образом развивается аутотолерантность к антигенам,
отсутствующим в тимусе?
T клетки, несущие
Каким образом развивается аутотолерантность к антигенам,
отсутствующим в тимусе?
T клетки, несущие
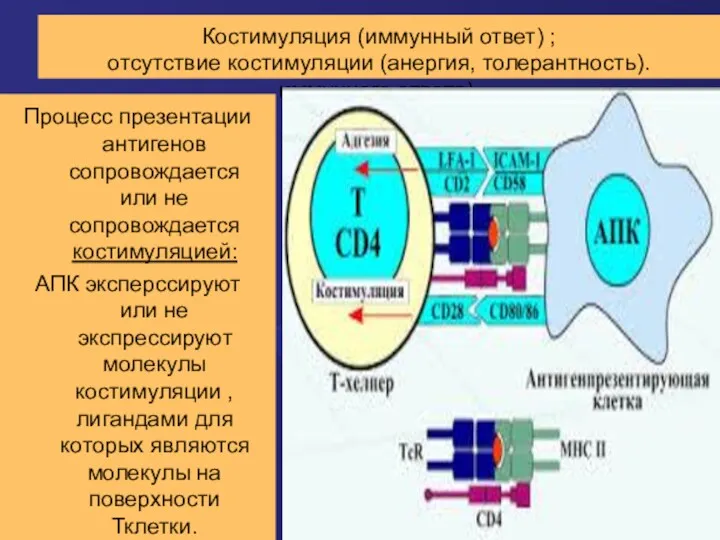
Костимуляция (иммунный ответ) ;
отсутствие костимуляции (анергия, толерантность). иммунного ответа).
Процесс презентации антигенов
Костимуляция (иммунный ответ) ;
отсутствие костимуляции (анергия, толерантность). иммунного ответа).
Процесс презентации антигенов
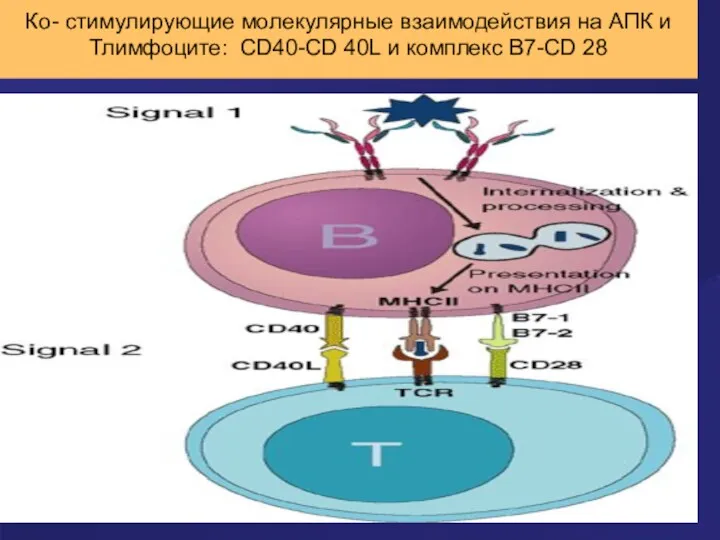
Ко- стимулирующие молекулярные взаимодействия на АПК и Тлимфоците: CD40-CD 40L и
Ко- стимулирующие молекулярные взаимодействия на АПК и Тлимфоците: CD40-CD 40L и
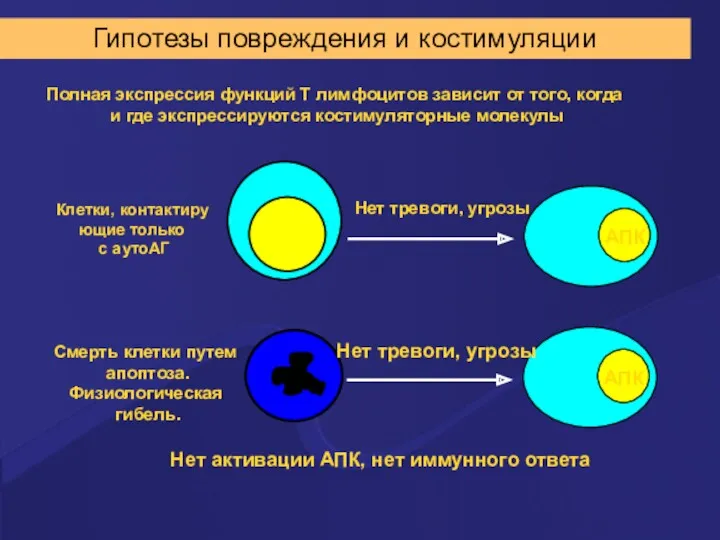
Гипотезы повреждения и костимуляции
Полная экспрессия функций Т лимфоцитов зависит от того,
Гипотезы повреждения и костимуляции
Полная экспрессия функций Т лимфоцитов зависит от того,
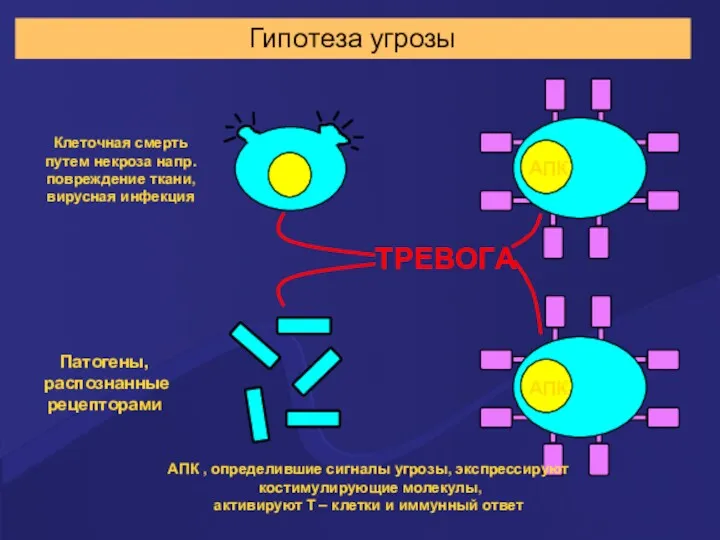
Гипотеза угрозы
АПК , определившие сигналы угрозы, экспрессируют
костимулирующие молекулы,
активируют Т
Гипотеза угрозы
АПК , определившие сигналы угрозы, экспрессируют
костимулирующие молекулы,
активируют Т

Зрелые Т-лимфоциты: жизненный путь на периферии
Выжившие в результате положительной и
Зрелые Т-лимфоциты: жизненный путь на периферии
Выжившие в результате положительной и

Зрелые Т-лимфоциты: жизненный путь на периферии
После встречи с антигеном в лимфоузле Т-клетка с
Зрелые Т-лимфоциты: жизненный путь на периферии
После встречи с антигеном в лимфоузле Т-клетка с
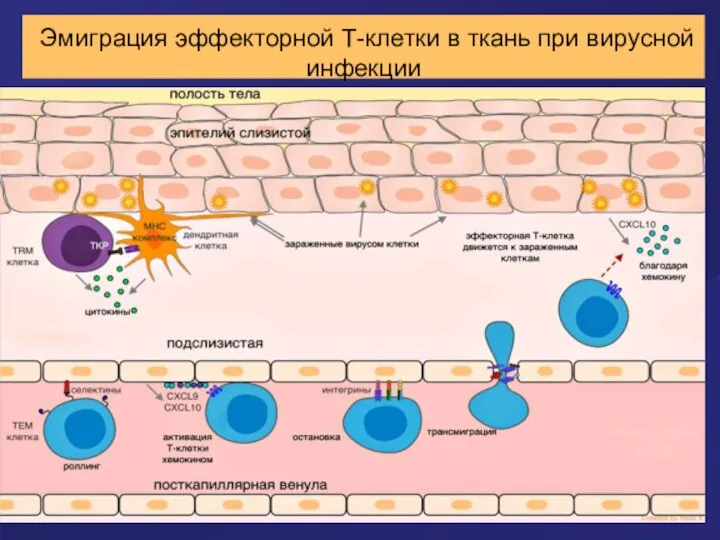
Эмиграция эффекторной Т-клетки в ткань при вирусной инфекции
Эмиграция эффекторной Т-клетки в ткань при вирусной инфекции

Зрелые Т-лимфоциты: рециркулирующие и резидентные
Лимфоциты рециркулируют по лимфе и кровотоку в поисках антигена, который
Зрелые Т-лимфоциты: рециркулирующие и резидентные
Лимфоциты рециркулируют по лимфе и кровотоку в поисках антигена, который
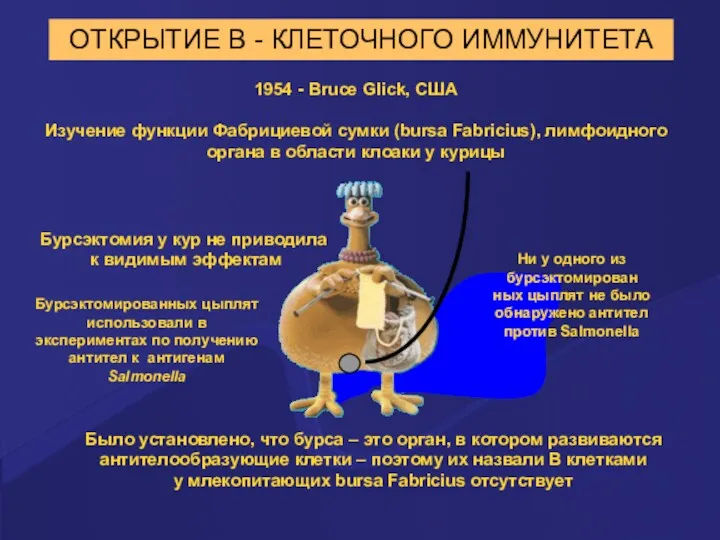
ОТКРЫТИЕ В - КЛЕТОЧНОГО ИММУНИТЕТА
1954 - Bruce Glick, США
Изучение функции Фабрициевой
ОТКРЫТИЕ В - КЛЕТОЧНОГО ИММУНИТЕТА
1954 - Bruce Glick, США
Изучение функции Фабрициевой
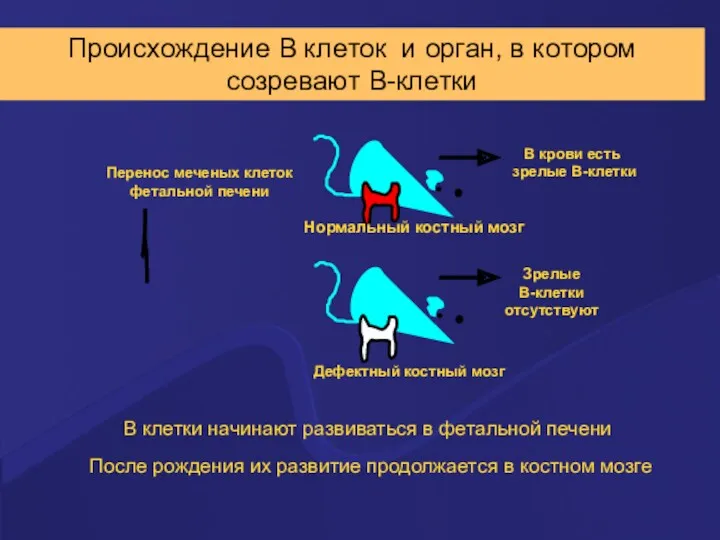
Происхождение В клеток и орган, в котором
созревают В-клетки
После рождения их
Происхождение В клеток и орган, в котором
созревают В-клетки
После рождения их
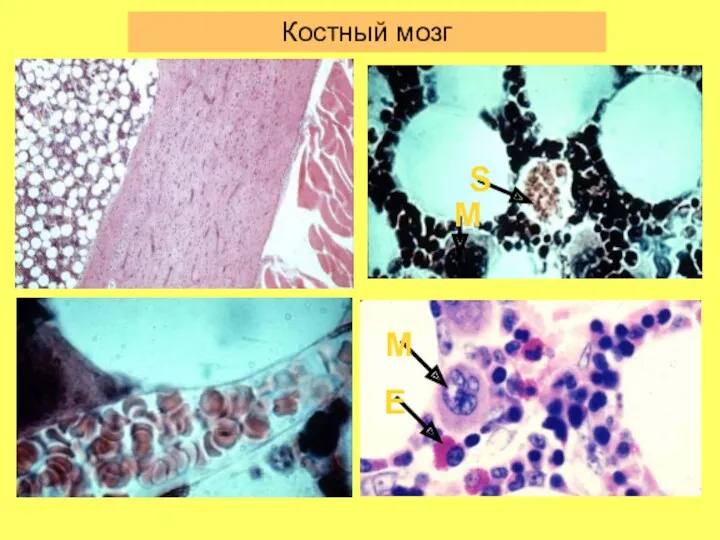
Костный мозг
Костный мозг

Стадии развития В лимфоцита
1). Стволовая клетка
2) Общий лимфоидный предшественник для B-
Стадии развития В лимфоцита
1). Стволовая клетка
2) Общий лимфоидный предшественник для B-
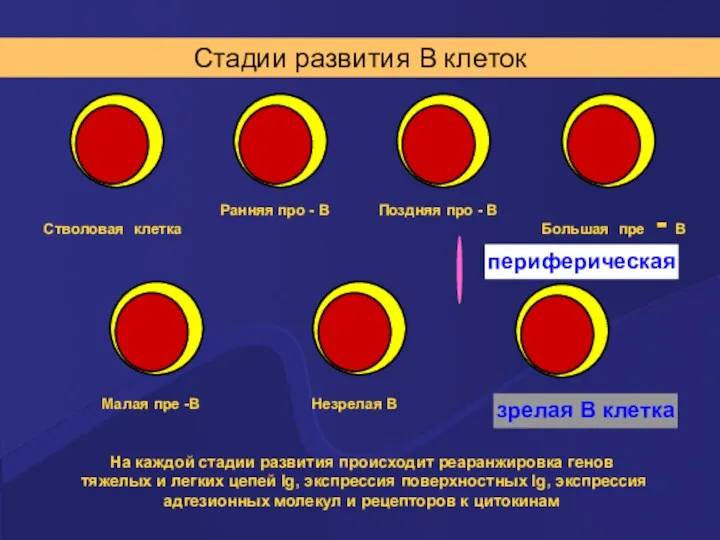
Стадии развития В клеток
На каждой стадии развития происходит реаранжировка генов тяжелых
Стадии развития В клеток
На каждой стадии развития происходит реаранжировка генов тяжелых

Стадии развития В клеток
Ранние этапы развития В-лимфоцитов зависят от прямого контактного
Стадии развития В клеток
Ранние этапы развития В-лимфоцитов зависят от прямого контактного

Стадии развития В клеток
В дальнейшем ранние пре-В-клетки, трансформируются в малые пре-В-лимфоциты,
Стадии развития В клеток
В дальнейшем ранние пре-В-клетки, трансформируются в малые пре-В-лимфоциты,

Развитие В клеток в костном мозге
Более 75% созревающих в костном мозге
Развитие В клеток в костном мозге
Более 75% созревающих в костном мозге
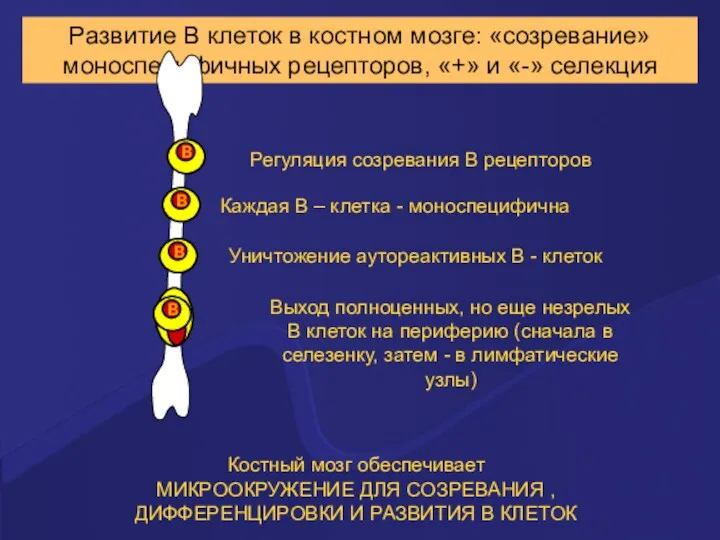
Развитие В клеток в костном мозге: «созревание» моноспецифичных рецепторов, «+» и
Развитие В клеток в костном мозге: «созревание» моноспецифичных рецепторов, «+» и
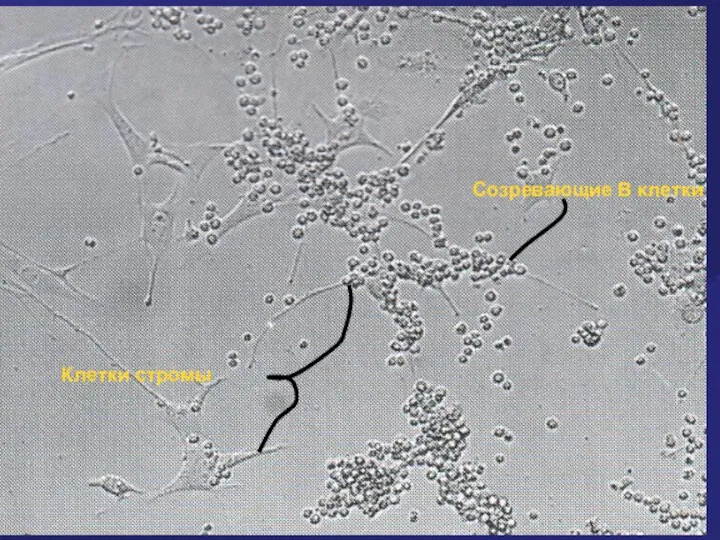
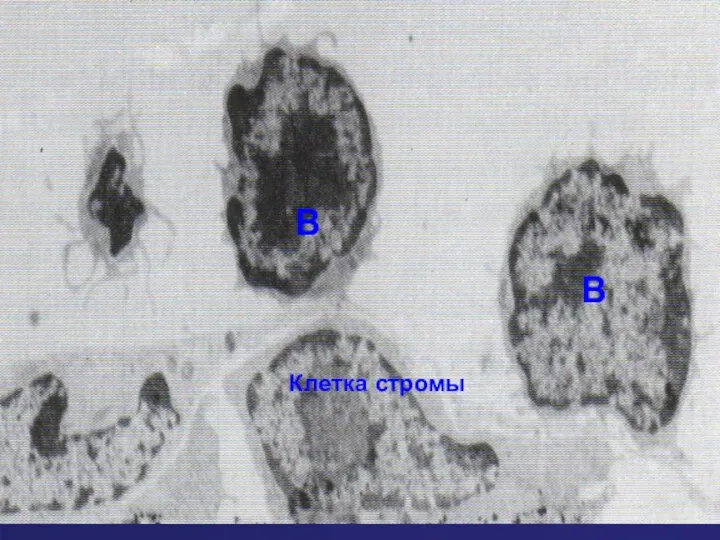
Клетки стромы
Созревающие В клетки
Клетки стромы
Созревающие В клетки
B
B
Клетка стромы
B
B
Клетка стромы
X
X
X
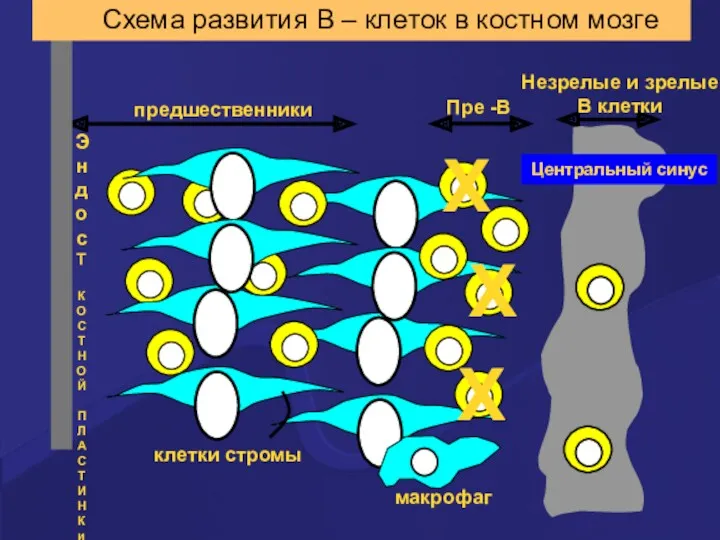
Схема развития В – клеток в костном мозге
X
X
X
Схема развития В – клеток в костном мозге
Y
Y
Y
Y
Y
Y
Зрелый В-лимфоцит
выходит
на периферию
Y
Y
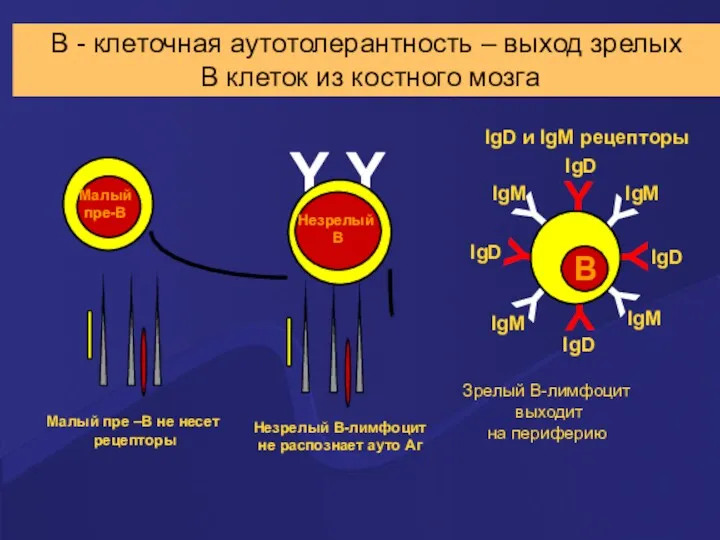
В - клеточная аутотолерантность – выход зрелых
Y
Y
Y
Y
Y
Y
Зрелый В-лимфоцит
выходит
на периферию
Y
Y
В - клеточная аутотолерантность – выход зрелых

Постулаты теории клональной селекции
Каждый В лимфоцит имеет рецептор уникальной специфичности.
Высокоаффинное (прочное)
Постулаты теории клональной селекции
Каждый В лимфоцит имеет рецептор уникальной специфичности.
Высокоаффинное (прочное)
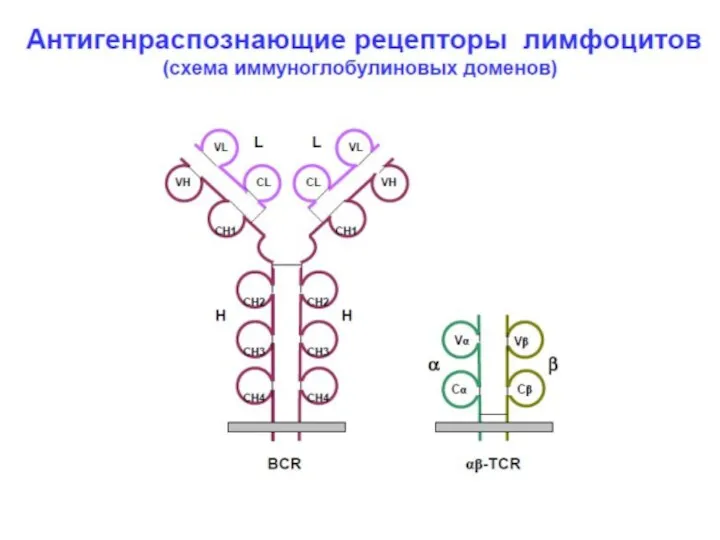

Гены молекул иммуноглобулинов (Ig)
Каждая молекула Ig состоит из 2 тяжелых (Н)
Гены молекул иммуноглобулинов (Ig)
Каждая молекула Ig состоит из 2 тяжелых (Н)
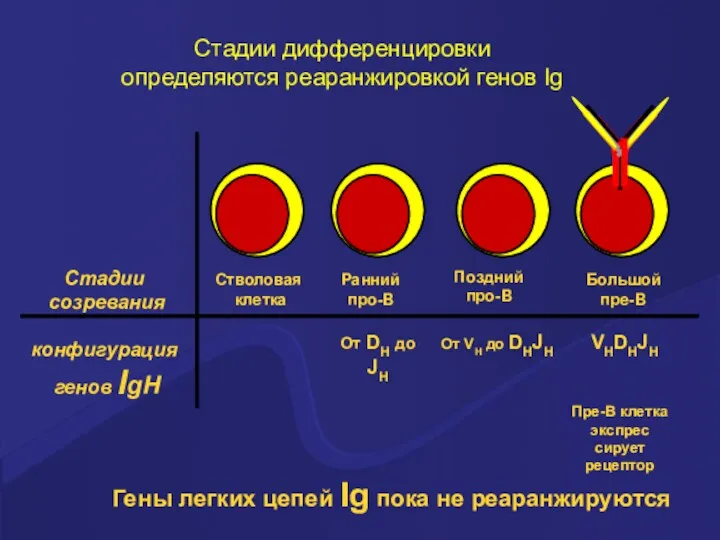
Стадии дифференцировки
определяются реаранжировкой генов Ig
Стадии
созревания
конфигурация
генов IgH
Стволовая
клетка
Ранний
про-В
Поздний
Стадии дифференцировки
определяются реаранжировкой генов Ig
Стадии
созревания
конфигурация
генов IgH
Стволовая
клетка
Ранний
про-В
Поздний

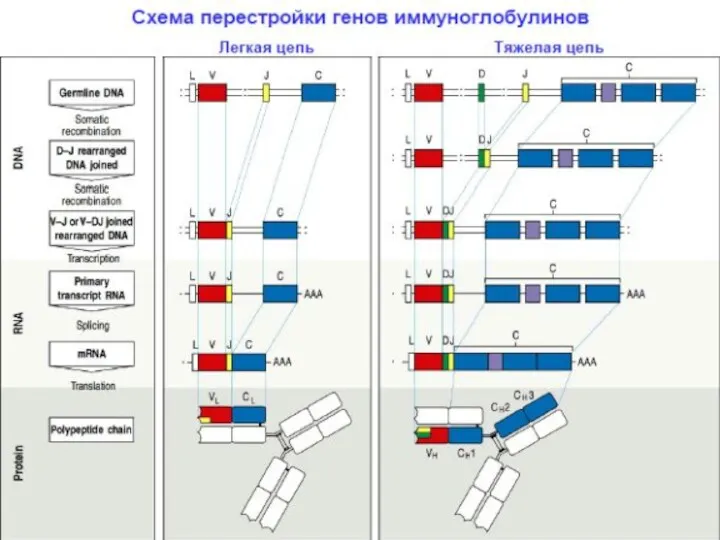
Реаранжировка генов, кодирующих легкие цепи молекулы Ig
После завершения перестройки (реаранжировки) генов,
Реаранжировка генов, кодирующих легкие цепи молекулы Ig
После завершения перестройки (реаранжировки) генов,
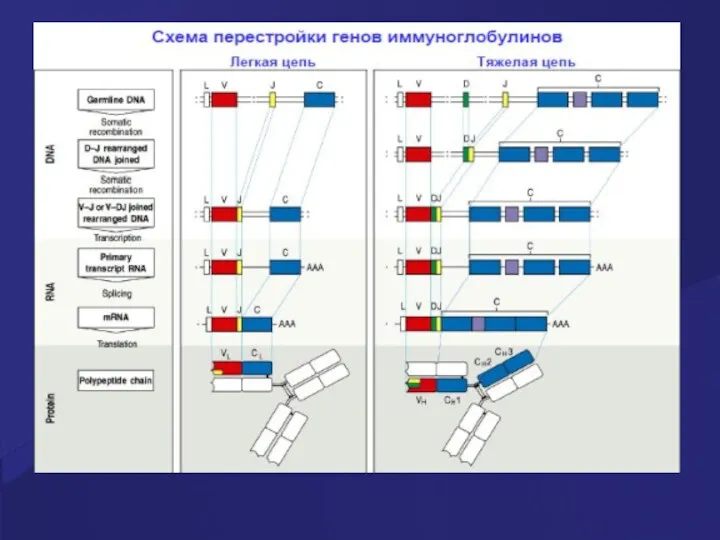

Положительная и отрицательная селекция В лимфоцитов в костном мозге
+ селекция происходит
Положительная и отрицательная селекция В лимфоцитов в костном мозге
+ селекция происходит

Гены Ig молекул
До встречи с антигеном:
Рекомбинации ограниченного числа генных сегментов
V,
Гены Ig молекул
До встречи с антигеном:
Рекомбинации ограниченного числа генных сегментов
V,

Дальнейшие этапы развития В-лимфоцитов
Из костного мозга уже отобранные В-лимфоциты попадают с
Дальнейшие этапы развития В-лимфоцитов
Из костного мозга уже отобранные В-лимфоциты попадают с


Выход зрелых В клеток на периферию
Из костного мозга выходят только те
Выход зрелых В клеток на периферию
Из костного мозга выходят только те
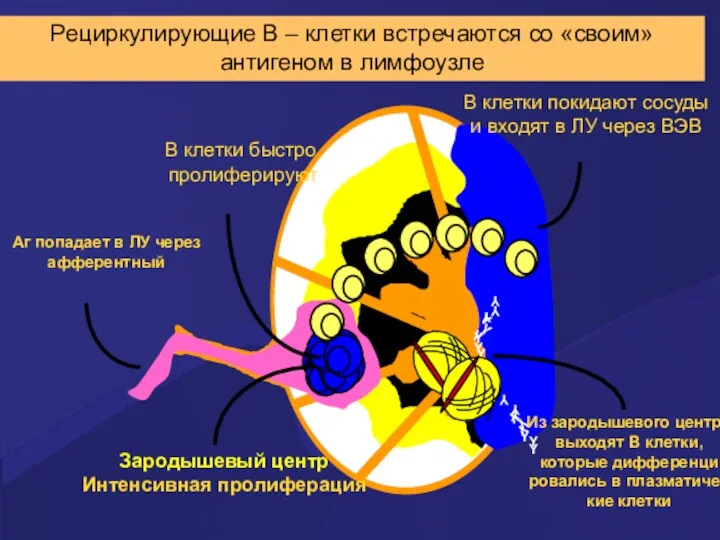
Рециркулирующие В – клетки встречаются со «своим» антигеном в лимфоузле
Рециркулирующие В – клетки встречаются со «своим» антигеном в лимфоузле
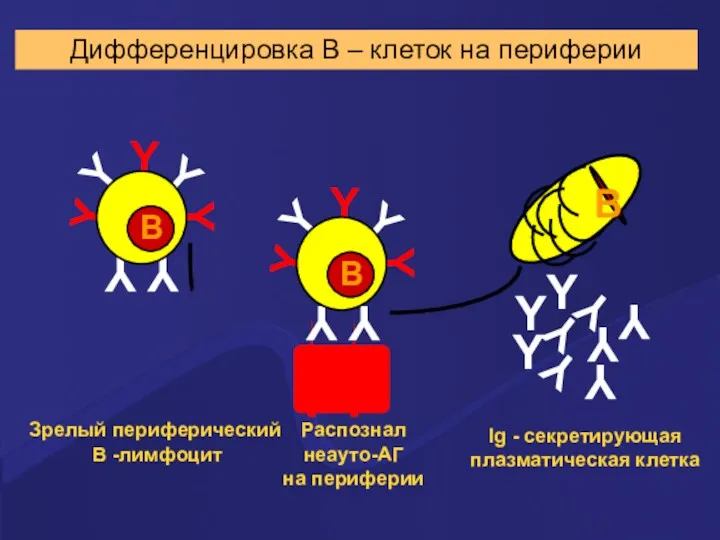
Распознал
неауто-АГ
на периферии
Ig - секретирующая
плазматическая клетка
Дифференцировка В –
Распознал
неауто-АГ
на периферии
Ig - секретирующая
плазматическая клетка
Дифференцировка В –

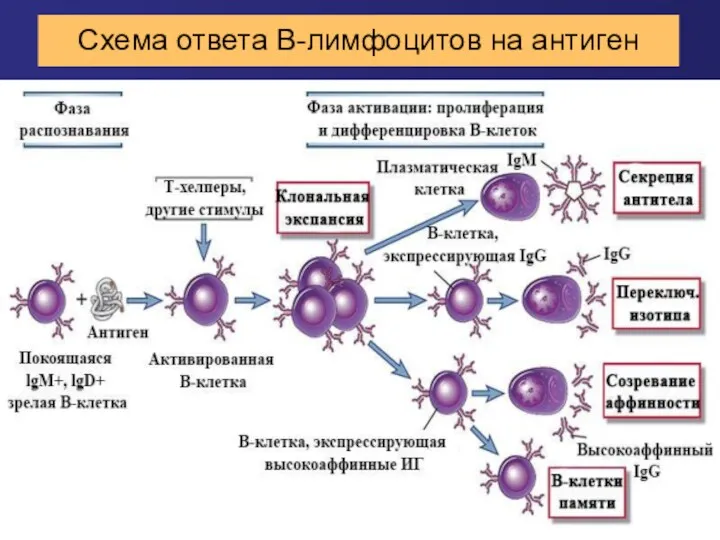
Схема ответа В-лимфоцитов на антиген
Схема ответа В-лимфоцитов на антиген

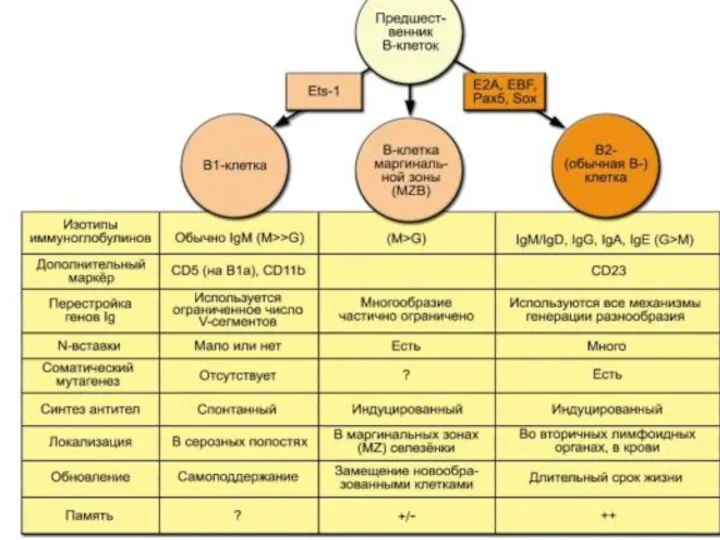
Субпопуляции В лимфоцитов: В1 и В2
В 2 (CD 5-) лимфоциты связывают
Субпопуляции В лимфоцитов: В1 и В2
В 2 (CD 5-) лимфоциты связывают

Субпопуляция В1 лимфоцитов
После активации В1 клетки секретируют анти-полисахаридные антитела класса М
Субпопуляция В1 лимфоцитов
После активации В1 клетки секретируют анти-полисахаридные антитела класса М
Взаимодействие АПК, Т- и В- лимфоцитов в ходе иммунного ответа на
Взаимодействие АПК, Т- и В- лимфоцитов в ходе иммунного ответа на

Вопросы к занятию № 3а
Какова роль тимуса в процессе дифференцировки Т-лимфоцитов?
Каков
Вопросы к занятию № 3а
Какова роль тимуса в процессе дифференцировки Т-лимфоцитов?
Каков

Тестовые задания к занятию №3
11. Антигеннезависимая дифференцировка Т лимфоцитов происходит в:
щитовидной
Тестовые задания к занятию №3
11. Антигеннезависимая дифференцировка Т лимфоцитов происходит в:
щитовидной

Тестовые задания к занятию №3
3. Общим маркером Т-лимфоцитов является молекула:
CD 3
CD
Тестовые задания к занятию №3
3. Общим маркером Т-лимфоцитов является молекула:
CD 3
CD

Тестовые задания к занятию №3
5. Для цитотоксических Т-лимфоцитов характерны маркеры:
CD 3
Тестовые задания к занятию №3
5. Для цитотоксических Т-лимфоцитов характерны маркеры:
CD 3

Тестовые задания к занятию № 3
7. Зародышевые гены тяжелых цепей иммуноглобулинов
Тестовые задания к занятию № 3
7. Зародышевые гены тяжелых цепей иммуноглобулинов
 Гемофилия. Клиника. Принципы лечения
Гемофилия. Клиника. Принципы лечения Etiologiya. Аллергендер сырттан енетін экзогендік және ағзаның өзінде болатын эндогендік болып ажыратылады
Etiologiya. Аллергендер сырттан енетін экзогендік және ағзаның өзінде болатын эндогендік болып ажыратылады Нарушение ритма сердца. Экстренная медицинская помощь. Показания для госпитализации
Нарушение ритма сердца. Экстренная медицинская помощь. Показания для госпитализации Риски и осложнения эндоваскулярного лечения аневризм торакоабдоминального отдела аорты
Риски и осложнения эндоваскулярного лечения аневризм торакоабдоминального отдела аорты Потовые железы. Ориентировочная реакция и привыкание
Потовые железы. Ориентировочная реакция и привыкание Creation of radiographs and sonograms for cardiovascular system
Creation of radiographs and sonograms for cardiovascular system Эндокриндік жүйенің бұзылыстары
Эндокриндік жүйенің бұзылыстары Телемедицина
Телемедицина Опухоли. Определение понятия. Номенклатура. Биология опухолевого роста
Опухоли. Определение понятия. Номенклатура. Биология опухолевого роста Профессиональная деятельность медицинской сестры в школе здоровья для больных артериальной гипертензией
Профессиональная деятельность медицинской сестры в школе здоровья для больных артериальной гипертензией Нейрофиброматоз Реклингхаузена
Нейрофиброматоз Реклингхаузена Болезнь Лайма
Болезнь Лайма Гнойно-воспалительные заболевания мягких тканей. Медиастинит
Гнойно-воспалительные заболевания мягких тканей. Медиастинит О работе Солигорской районной организации Белорусского общества Красного Креста в 2017 году
О работе Солигорской районной организации Белорусского общества Красного Креста в 2017 году Недоношенные дети
Недоношенные дети Универсальный алгоритм оказания первой помощи
Универсальный алгоритм оказания первой помощи Акушерский сепсис – диагностика и терапия
Акушерский сепсис – диагностика и терапия Совершенствование диспансеризации населения на современном этапе модернизации первичного звена здравоохранения
Совершенствование диспансеризации населения на современном этапе модернизации первичного звена здравоохранения ДЦП как нарушение опорно-двигательного аппарата
ДЦП как нарушение опорно-двигательного аппарата Учаскелік терапевт диспансеризацияны
Учаскелік терапевт диспансеризацияны Деконгестанты: как выбрать оптимальный препарат
Деконгестанты: как выбрать оптимальный препарат Пациент және оның отбасына реабилитация шараларына үйрету
Пациент және оның отбасына реабилитация шараларына үйрету Ауруханадан тыс екі жақты төменгі бөліктік пневмония, орташа ауыр жағдайда. ДН II
Ауруханадан тыс екі жақты төменгі бөліктік пневмония, орташа ауыр жағдайда. ДН II Язвенная болезнь
Язвенная болезнь Заболевания околощитовидных желез
Заболевания околощитовидных желез “Жедел көмек” дәрігер тәжірбиесіндегі антиаритмиялық препараттар
“Жедел көмек” дәрігер тәжірбиесіндегі антиаритмиялық препараттар Дәрігердің жұмысындағы қарым-қатынас кедергілері
Дәрігердің жұмысындағы қарым-қатынас кедергілері Инфекционная безопасность
Инфекционная безопасность