- Клеточный иммунный ответ. Основные популяции клеток и механизмы их взаимодействия

Содержание
- 2. Зависимость типа иммунного ответа от типа патогена Гуморальный ответ Развивается в основном на внеклеточные паразиты (большинство
- 3. Внеклеточные патогены Внутриклеточные патогены эндоцитоз патогена, слияние фагосомы с лизосомой, деградация антигена, упаковка антигенных пептидов в
- 4. Зависимость типа иммунного ответа от типа презентации антигена Внеклеточные патогены Внеклеточные антигены представляются в молекулах MHC
- 5. Зависимость типа иммунного ответа от цитокинов АПК (ДК) Разные типы цитокинов, синтезируемые ДК, направляют дифференцировку Т
- 6. Цитокины ДК в регуляции функций Т-хелперов по Akdis M. et al., 2012 Bcl-6 сигнал поляризации антиген
- 7. Сравнение гуморального и клеточного иммунных ответов 1. Гуморальный иммунный ответ : Th0 (наивные) мигрируют в лимфоузлы
- 8. Этапы гуморального иммунного ответа: 1. Роль дендритных клеток (ДК) –презентация антигена Т-лимфоцитам, костимуляция Проникновение антигена в
- 9. 2.Контакт между антигенпрезентирующей ДК и наивным CD4+ лимфоцитом (Th0) в лимфоузле – образование антигенспецифичных клонов Tfn
- 10. 3. Наивный В лимфоцит тоже является АПК Наивные В- клетки распознают растворимый или связанный с клеточной
- 11. 4.«Моногамный контакт»: В лимфоцит - T fn На границе Т- и В-зон лимфоузла происходит первое распознавание
- 12. 5. Первичный иммунный ответ В результате контакта активированных В-лимфоцитов и Tfh происходит дальнейшая дифференцировка В клеток
- 13. «Моногамный контакт»: В лимоцит -T fn IL21R IL21 IL21R CXCR5 CXCL13 CXCL13 PD-1 PD-1L
- 14. Первичный иммунный ответ: роль Tfh (фолликулярных Т хелперов) На территории лимфоидного фолликула и развивающегося из него
- 15. Вторичный иммунный ответ (повторное попадание антигена, на который уже сформированы клетки памяти): роль Tfh памяти и
- 16. 6. Вторичный иммунный ответ: после вповторного проникновения антигена в организм «работают» клетки памяти - memory Tfn
- 17. Схема гуморального иммунного ответа Tfn IL-21,10,6 IL-21
- 18. Сравнение гуморального и клеточного иммунных ответов 2. Клеточный иммунный ответ : Th0 (наивные) мигрируют в лимфоузлы
- 19. Цитокины ДК в регуляции функций Т-хелперов по Akdis M. et al., 2012 Bcl-6 сигнал поляризации антиген
- 20. Разные типы иммунных ответов Th fn IL- 21 IL-21, IL-10, IL- 6 Синтез иммуноглобулинов IL-21 Гуморальный
- 21. Клеточная цитотоксичность - ответ CD8+ Т лимфоцитов на внутриклеточные патогены Клеточная цитотоксичность развивается в ответ на
- 22. Стадии развития цитотоксического (CD8+) иммунного ответа: 1.Индуктивная стадия (запуск ответа) роль ДК Дендритные клетки (ДК) после
- 23. Стадии развития цитотоксического (CD8+) иммунного ответа: 1.Индуктивная стадия (запуск ответа), роль ДК По градиенту хемокинов в
- 24. Строение иммунологического синапса: 3 зоны контактов между АПК и Т лимфоцитами СИГНАЛ 1 (антигенное распознавание): группа
- 25. Строение иммунологического синапса: 3 зоны контактов между АПК и Т лимфоцитами СИГНАЛЫ 2 И 3: Группа
- 26. Биологический смысл костимулирующих взаимодействий СИГНАЛ 2 (пролиферация):CD 80 или CD 86 (ДК) и CD 28 (Т-л)
- 27. Биологический смысл костимулирующих взаимодействий Взаимодействие молекул CD40 поверхностной мембране дендритных клеток с CD40 L(лигандами) на поверхностной
- 28. Презентация антигена - T клетки костимулированы Костимулирующие молекулы экспрессируются на большинстве АПК
- 29. экспрессия β и γ цепей IL-2 рецептора, но не α цепи Механизм костимуляции Т клеток сигналом
- 30. сигнал 2 активируются факторы AP-1 и NFκ-B и повышают транскрипцию гена IL-2 троекратно, стабилизируют и повышают
- 31. Анергия (неотвечаемость) развивается при отсутствии сигналов костимуляции Если Т клетка получает только СИГНАЛ 1 от АПК
- 32. Сигналы 1, 2 сигнал1 антиген и рецептор сигнал 2 B7 - CD28 костимуляция и 3 Сигналы
- 33. Поляризация ответа
- 34. Виды клеточного иммунного ответа в зависимости от типа внутриклеточного паразитирования патогенов (вирусы или внутриклеточно-паразитирующие бактерии) Вирусы
- 35. Этапы взаимодействия ДК с наивным Т-хелпером(Th0) в Т-клеточной зоне лимфоузла: образование Th1 1. Происходит взаимодействие: комплекса
- 36. Помощь Т-хелперов 1 типа CD8+ лимфоцитам в Т-клеточной зоне лимфоузла Роль ИЛ-2 Активированные Th 1 начинают
- 38. Стадии развития цитотоксического (CD8+) ответа: эффекторная стадия – механизмы киллинга пораженных вирусами клеток – мишеней Киллинг
- 39. Эффекторная стадия цитотоксического ответа: механизмы цитотоксичности CTL На мембране клетке-мишени эксперссируется рецептор «готовности к смерти» -
- 40. Т-киллер убивает клетку, на поверхности которой презентирован ассоциированный с MHC-I антиген, к которому комплементарен Т- клеточный
- 41. Заключительный этап цитотоксического (CD8+) иммунного ответа: клетки памяти Через 7-10 дней после распознавания антигена эффекторы цитотоксического
- 42. T h1 –опосредованный воспалительный ответ на внутриклеточно-паразитирующие в макрофагах патогены T h1 –опосредованный воспалительный ответ осуществляет
- 43. Краткая характеристика T h1 – опосредованного воспалительного ответа Презентация АГ дендритными клетками CD 4+Т –лимфоцитам, костиумляция,
- 44. Этапы воспалительного T h1 –опосредованного ответа(ГЗТ) В очаге инфицирования дендритные клетки поглощают патоген и или его
- 45. Этапы воспалительного T h1 –опосредованного ответа (ГЗТ) В макрофагах активируются гены, ответственные за активацию окислительного метаболизма.
- 46. Гранулемы: А- при туберкулезе; В- при лепре
- 47. Саркоидные гранулемы: неказеозные
- 48. Клеточный иммунный ответ NK не образуются из Tн 1, а АКТИВИРУЮТСЯ их цитокинами -IL-2 и IFN-γ
- 50. Типы иммунного ответа
- 51. Типы иммунного ответа
- 52. Типы иммунного ответа
- 53. Типы иммунного ответа
- 54. Типы иммунного ответа
- 55. Вопросы Типы иммунного ответа и факторы, их определяющие. Внутриклеточная локализация патогенна и тип клеточного иммунного ответа.
- 56. Тестовые вопросы Антиген в Т-зависимых зонах лимфатических узлов Т-лимфоцитам представляют: Плазматические клетки Дендритные клетки Эозинофилы Нейтрофилы
- 57. Тестовые вопросы Цитотоксические Т-лимфоциты своим корецептором вступают во взаимодействие с: Молекулами MHC I класса Молекулами MHC
- 58. Тестовые вопросы Цитотоксические Т-лимфоциты вызывают гибель клеток-мишеней с помощью: Перфоринов и гранзимов Fas-опосредованного апоптоза Цитокинового механизма
- 59. Тестовые вопросы Цитотоксичность - это: Способность вызывать гибель клеток-мишеней Способность к переключению изотипов иммуноглобулинов Многократное повышение
- 61. Скачать презентацию

Зависимость типа иммунного ответа
от типа патогена
Гуморальный ответ
Развивается в основном
Зависимость типа иммунного ответа
от типа патогена
Гуморальный ответ
Развивается в основном
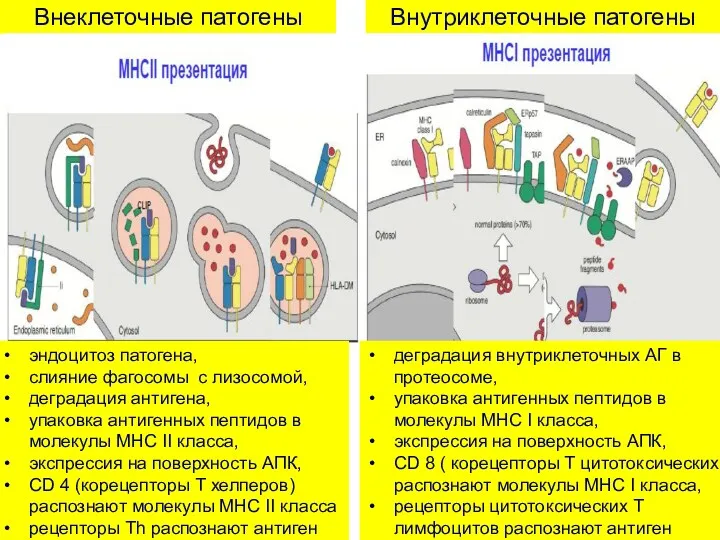
Внеклеточные патогены
Внутриклеточные патогены
эндоцитоз патогена,
слияние фагосомы с лизосомой,
деградация антигена,
упаковка антигенных пептидов
Внеклеточные патогены
Внутриклеточные патогены
эндоцитоз патогена,
слияние фагосомы с лизосомой,
деградация антигена,
упаковка антигенных пептидов

Зависимость типа иммунного ответа от типа презентации антигена
Внеклеточные патогены
Внеклеточные антигены представляются
Зависимость типа иммунного ответа от типа презентации антигена
Внеклеточные патогены
Внеклеточные антигены представляются

Зависимость типа иммунного ответа от цитокинов АПК (ДК)
Разные типы цитокинов, синтезируемые
Зависимость типа иммунного ответа от цитокинов АПК (ДК)
Разные типы цитокинов, синтезируемые
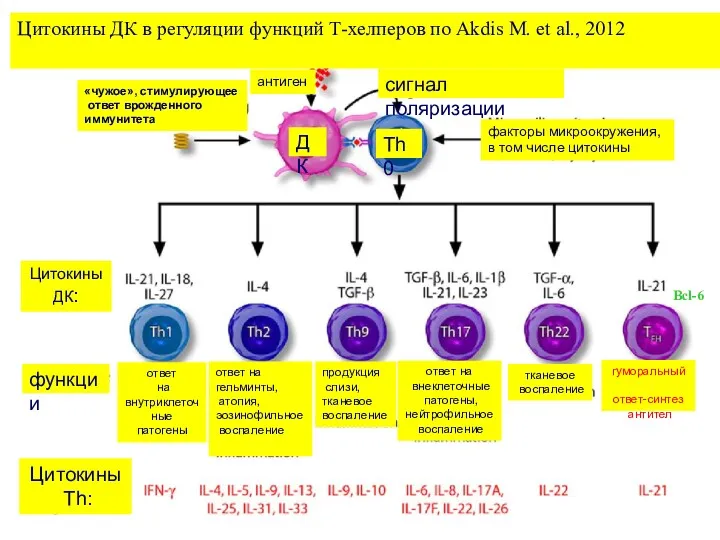
Цитокины ДК в регуляции функций Т-хелперов по Akdis M. et al.,
Цитокины ДК в регуляции функций Т-хелперов по Akdis M. et al.,
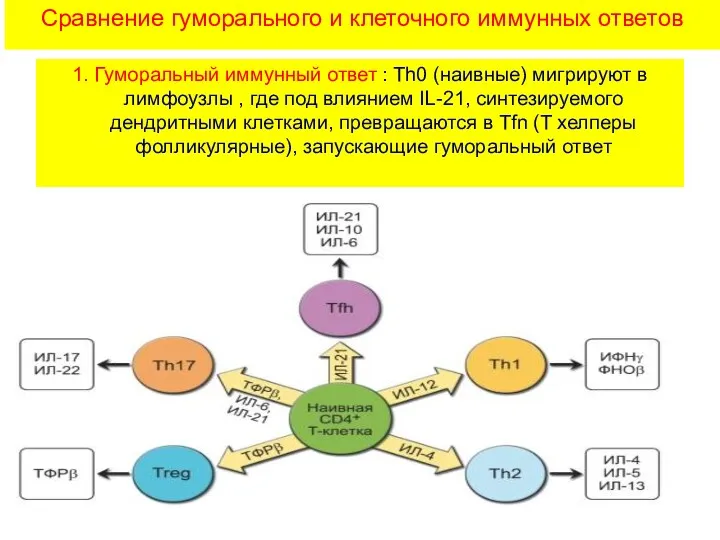
Сравнение гуморального и клеточного иммунных ответов
1. Гуморальный иммунный ответ : Th0
Сравнение гуморального и клеточного иммунных ответов
1. Гуморальный иммунный ответ : Th0

Этапы гуморального иммунного ответа:
1. Роль дендритных клеток (ДК) –презентация антигена Т-лимфоцитам,
Этапы гуморального иммунного ответа: 1. Роль дендритных клеток (ДК) –презентация антигена Т-лимфоцитам,
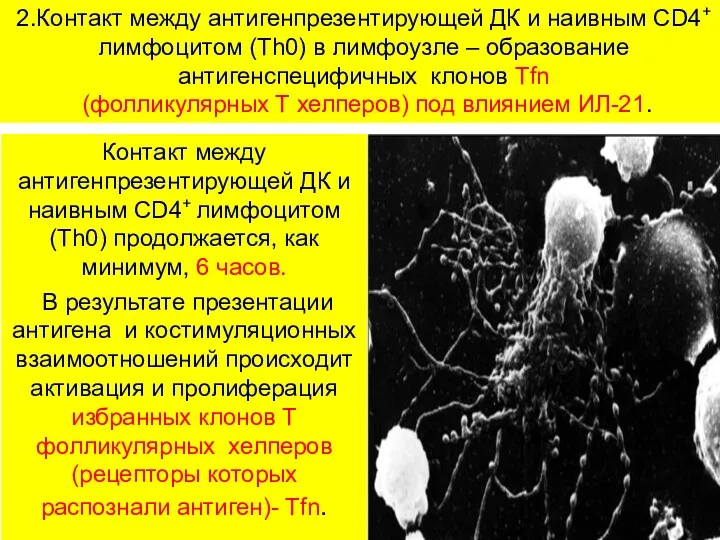
2.Контакт между антигенпрезентирующей ДК и наивным CD4+ лимфоцитом (Th0) в лимфоузле
2.Контакт между антигенпрезентирующей ДК и наивным CD4+ лимфоцитом (Th0) в лимфоузле

3. Наивный В лимфоцит тоже является АПК
Наивные В- клетки распознают растворимый
3. Наивный В лимфоцит тоже является АПК
Наивные В- клетки распознают растворимый

4.«Моногамный контакт»: В лимфоцит - T fn
На границе Т- и В-зон
4.«Моногамный контакт»: В лимфоцит - T fn
На границе Т- и В-зон

5. Первичный иммунный ответ
В результате контакта активированных В-лимфоцитов и Tfh происходит
5. Первичный иммунный ответ
В результате контакта активированных В-лимфоцитов и Tfh происходит
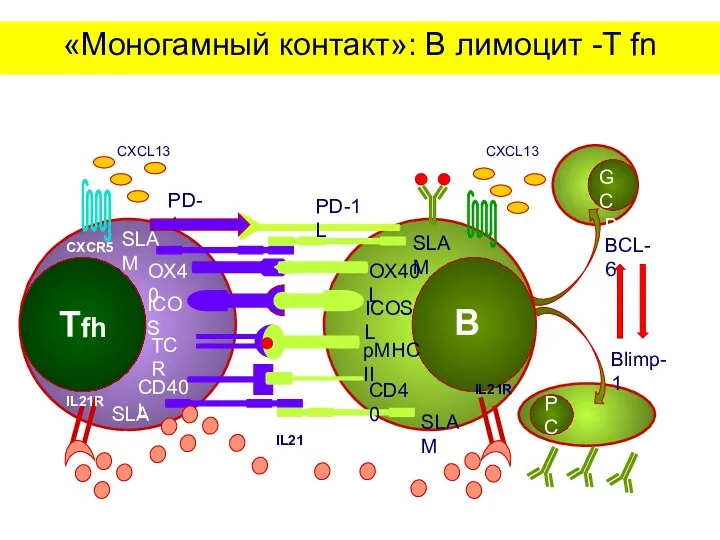
«Моногамный контакт»: В лимоцит -T fn
IL21R
IL21
IL21R
CXCR5
CXCL13
CXCL13
PD-1
PD-1L
«Моногамный контакт»: В лимоцит -T fn
IL21R
IL21
IL21R
CXCR5
CXCL13
CXCL13
PD-1
PD-1L
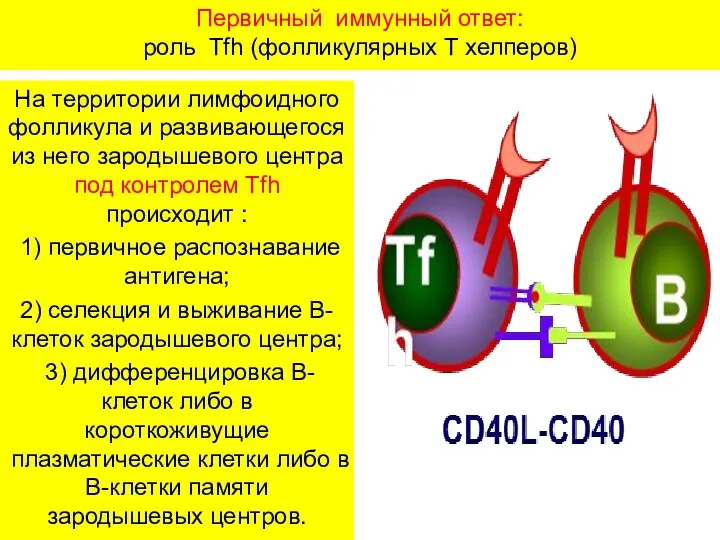
Первичный иммунный ответ:
роль Tfh (фолликулярных Т хелперов)
На территории лимфоидного
Первичный иммунный ответ:
роль Tfh (фолликулярных Т хелперов)
На территории лимфоидного
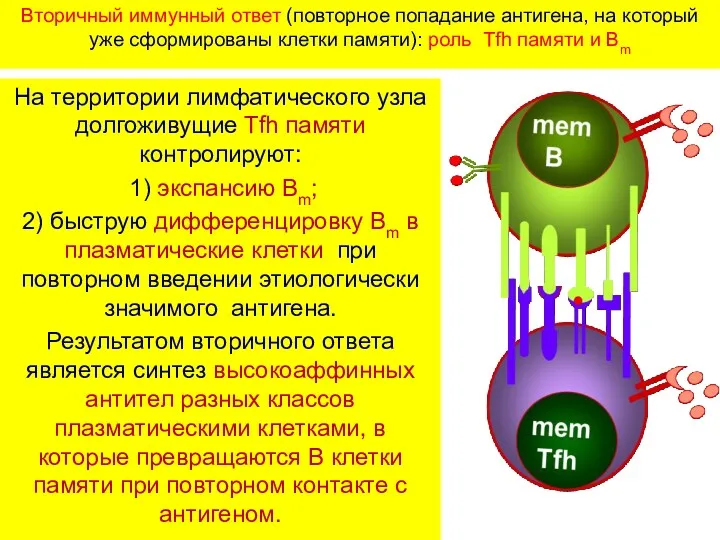
Вторичный иммунный ответ (повторное попадание антигена, на который уже сформированы клетки
Вторичный иммунный ответ (повторное попадание антигена, на который уже сформированы клетки
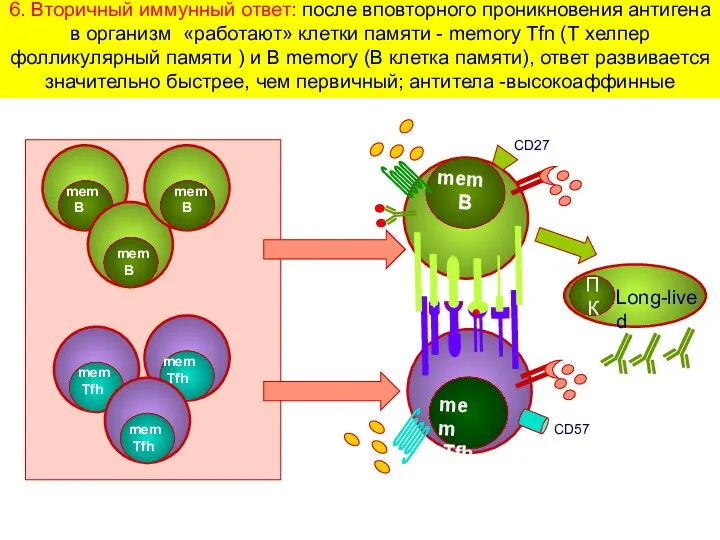
6. Вторичный иммунный ответ: после вповторного проникновения антигена в организм «работают»
6. Вторичный иммунный ответ: после вповторного проникновения антигена в организм «работают»
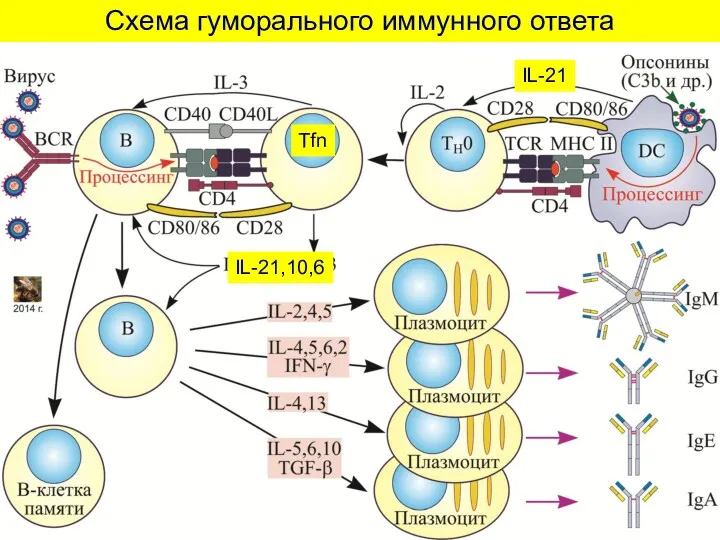
Схема гуморального иммунного ответа
Tfn
IL-21,10,6
IL-21
Схема гуморального иммунного ответа
Tfn
IL-21,10,6
IL-21
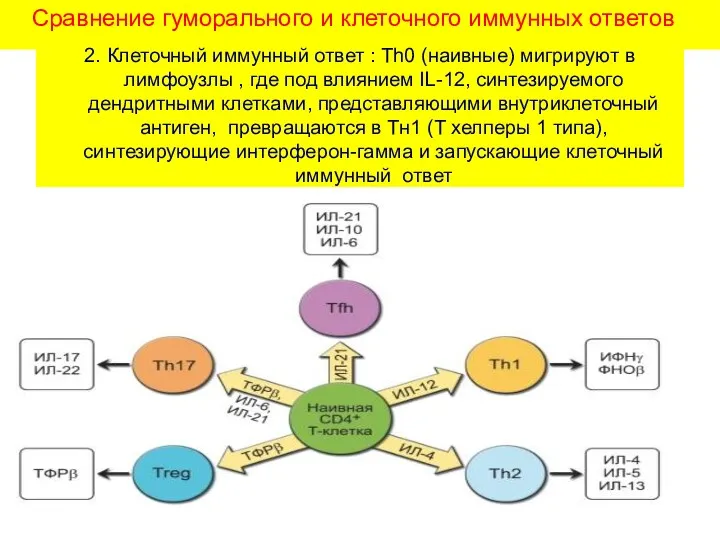
Сравнение гуморального и клеточного иммунных ответов
2. Клеточный иммунный ответ : Th0
Сравнение гуморального и клеточного иммунных ответов
2. Клеточный иммунный ответ : Th0
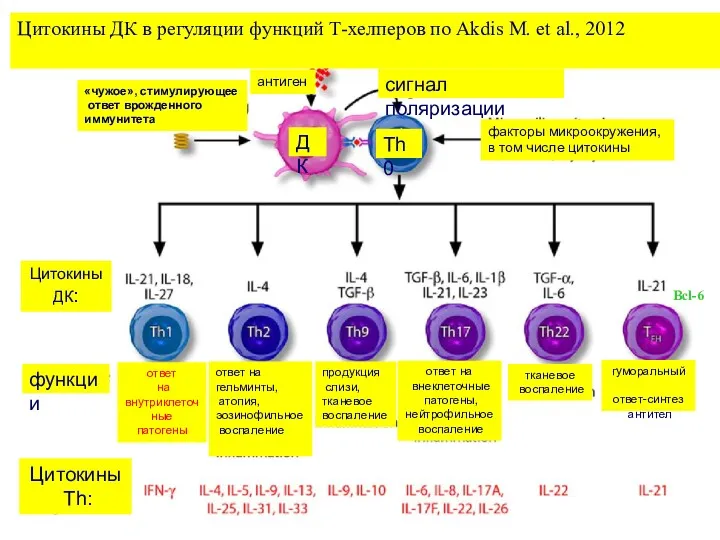
Цитокины ДК в регуляции функций Т-хелперов по Akdis M. et al.,
Цитокины ДК в регуляции функций Т-хелперов по Akdis M. et al.,
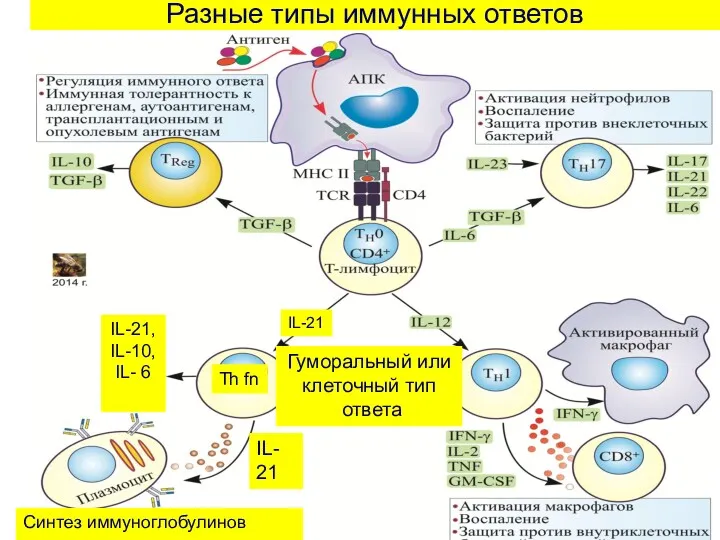
Разные типы иммунных ответов
Th fn
IL-
21
IL-21, IL-10, IL- 6
Синтез иммуноглобулинов
IL-21
Гуморальный или
клеточный
Разные типы иммунных ответов
Th fn
IL-
21
IL-21, IL-10, IL- 6
Синтез иммуноглобулинов
IL-21
Гуморальный или
клеточный

Клеточная цитотоксичность - ответ CD8+ Т лимфоцитов на внутриклеточные патогены
Клеточная цитотоксичность
Клеточная цитотоксичность - ответ CD8+ Т лимфоцитов на внутриклеточные патогены
Клеточная цитотоксичность

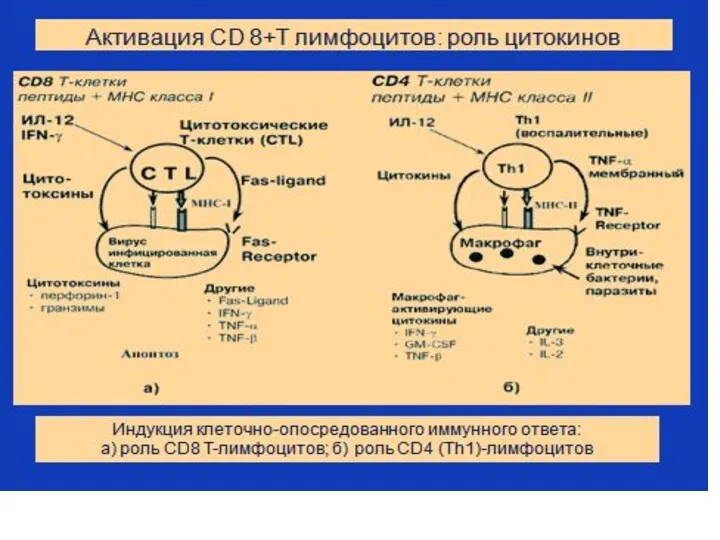
Стадии развития цитотоксического (CD8+) иммунного ответа: 1.Индуктивная стадия (запуск ответа) роль
Стадии развития цитотоксического (CD8+) иммунного ответа: 1.Индуктивная стадия (запуск ответа) роль

Стадии развития цитотоксического (CD8+) иммунного ответа: 1.Индуктивная стадия (запуск ответа), роль
Стадии развития цитотоксического (CD8+) иммунного ответа: 1.Индуктивная стадия (запуск ответа), роль

Строение иммунологического синапса:
3 зоны контактов между АПК и Т лимфоцитами
СИГНАЛ
Строение иммунологического синапса:
3 зоны контактов между АПК и Т лимфоцитами
СИГНАЛ

Строение иммунологического синапса:
3 зоны контактов между АПК и Т лимфоцитами
СИГНАЛЫ
Строение иммунологического синапса:
3 зоны контактов между АПК и Т лимфоцитами
СИГНАЛЫ

Биологический смысл костимулирующих взаимодействий
СИГНАЛ 2 (пролиферация):CD 80 или CD 86 (ДК)
Биологический смысл костимулирующих взаимодействий
СИГНАЛ 2 (пролиферация):CD 80 или CD 86 (ДК)

Биологический смысл костимулирующих взаимодействий
Взаимодействие
молекул CD40 поверхностной мембране дендритных клеток с
Биологический смысл костимулирующих взаимодействий
Взаимодействие
молекул CD40 поверхностной мембране дендритных клеток с
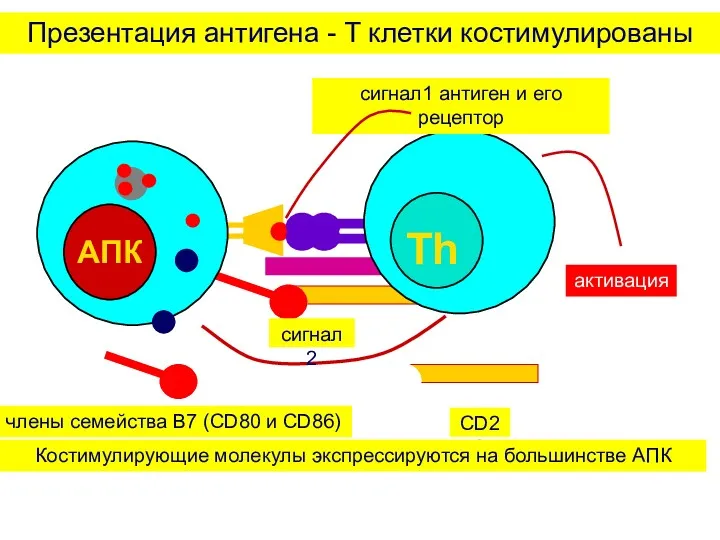
Презентация антигена - T клетки костимулированы
Костимулирующие молекулы экспрессируются на большинстве АПК
Презентация антигена - T клетки костимулированы
Костимулирующие молекулы экспрессируются на большинстве АПК
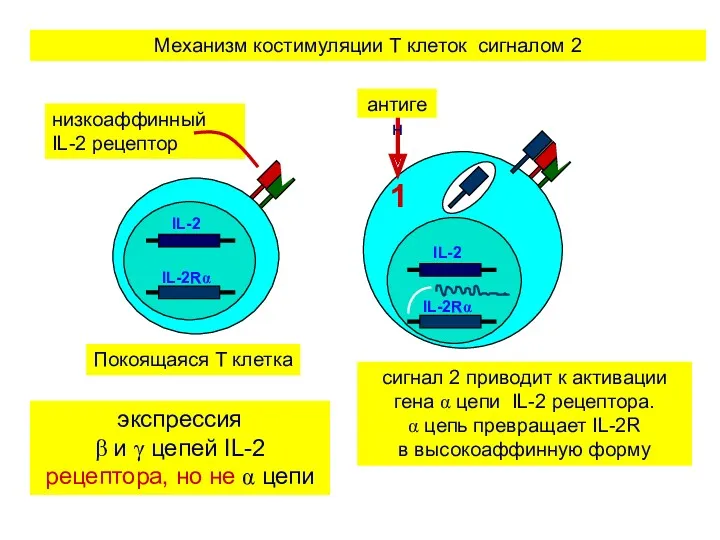
экспрессия
β и γ цепей IL-2 рецептора, но не α цепи
Механизм
экспрессия
β и γ цепей IL-2 рецептора, но не α цепи
Механизм
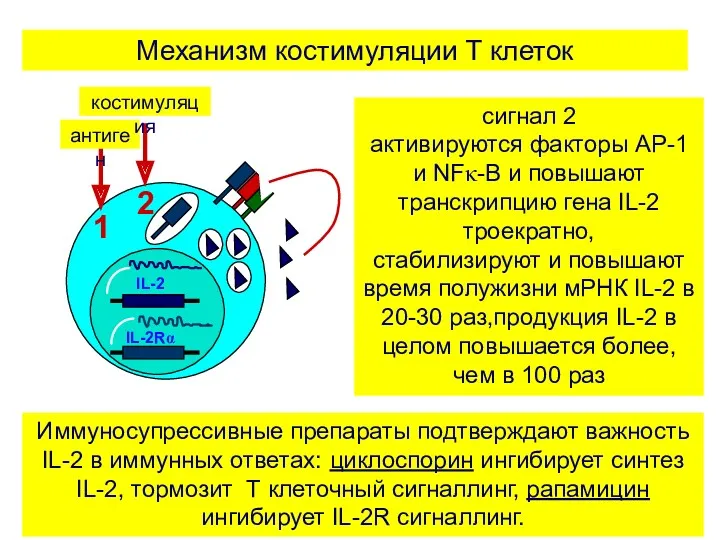
сигнал 2
активируются факторы AP-1 и NFκ-B и повышают транскрипцию гена IL-2
сигнал 2
активируются факторы AP-1 и NFκ-B и повышают транскрипцию гена IL-2

Анергия (неотвечаемость) развивается при отсутствии сигналов костимуляции
Если Т клетка получает только
Анергия (неотвечаемость) развивается при отсутствии сигналов костимуляции
Если Т клетка получает только
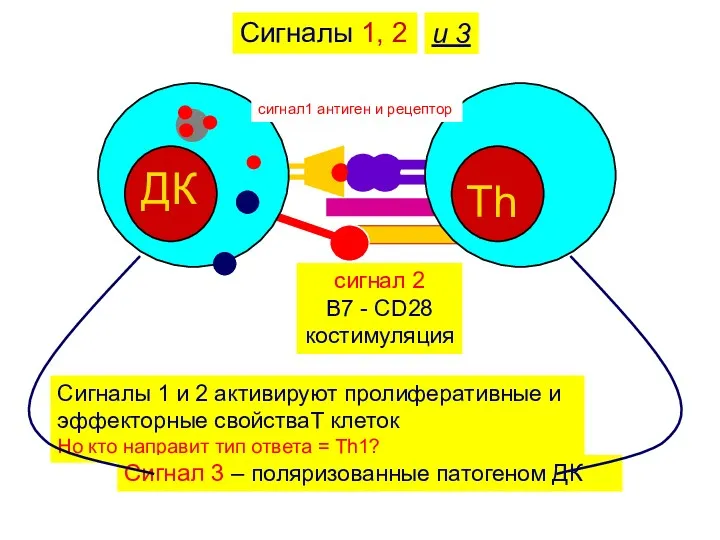
Сигналы 1, 2
сигнал1 антиген и рецептор
сигнал 2
B7 - CD28
костимуляция
и 3
Сигналы 1
Сигналы 1, 2
сигнал1 антиген и рецептор
сигнал 2
B7 - CD28
костимуляция
и 3
Сигналы 1
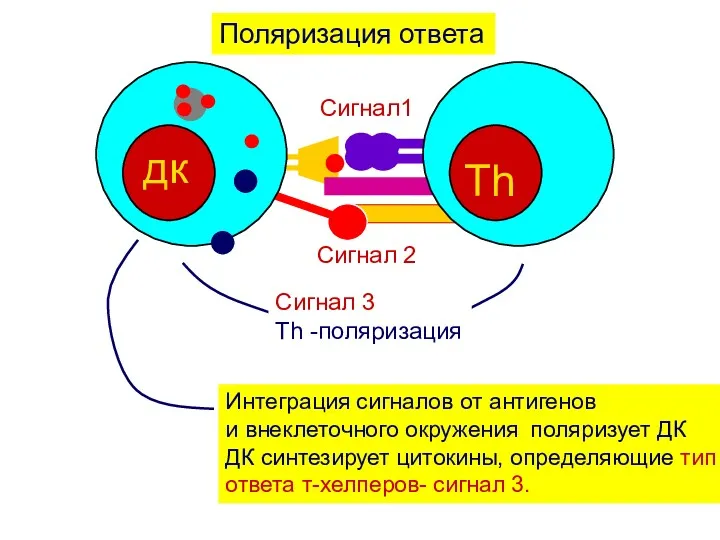
Поляризация ответа
Поляризация ответа

Виды клеточного иммунного ответа в зависимости от типа внутриклеточного паразитирования патогенов
Виды клеточного иммунного ответа в зависимости от типа внутриклеточного паразитирования патогенов

Этапы взаимодействия ДК с наивным Т-хелпером(Th0) в Т-клеточной зоне лимфоузла: образование
Этапы взаимодействия ДК с наивным Т-хелпером(Th0) в Т-клеточной зоне лимфоузла: образование

Помощь Т-хелперов 1 типа CD8+ лимфоцитам в Т-клеточной зоне лимфоузла
Роль ИЛ-2
Активированные
Помощь Т-хелперов 1 типа CD8+ лимфоцитам в Т-клеточной зоне лимфоузла
Роль ИЛ-2
Активированные

Стадии развития цитотоксического (CD8+) ответа: эффекторная стадия – механизмы киллинга пораженных
Стадии развития цитотоксического (CD8+) ответа: эффекторная стадия – механизмы киллинга пораженных

Эффекторная стадия цитотоксического ответа: механизмы цитотоксичности CTL
На мембране клетке-мишени эксперссируется
Эффекторная стадия цитотоксического ответа: механизмы цитотоксичности CTL
На мембране клетке-мишени эксперссируется
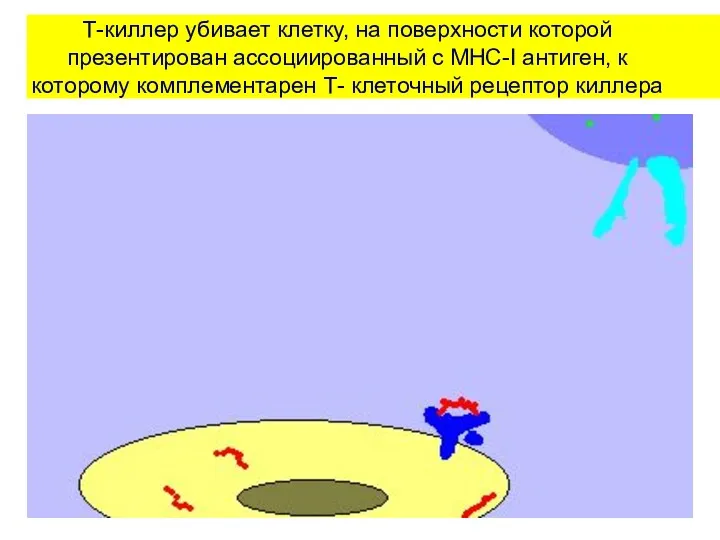
Т-киллер убивает клетку, на поверхности которой презентирован ассоциированный с MHC-I антиген,
Т-киллер убивает клетку, на поверхности которой презентирован ассоциированный с MHC-I антиген,

Заключительный этап цитотоксического (CD8+) иммунного ответа: клетки памяти
Через 7-10 дней после
Заключительный этап цитотоксического (CD8+) иммунного ответа: клетки памяти
Через 7-10 дней после

T h1 –опосредованный воспалительный ответ на внутриклеточно-паразитирующие в макрофагах патогены
T h1
T h1 –опосредованный воспалительный ответ на внутриклеточно-паразитирующие в макрофагах патогены
T h1

Краткая характеристика T h1 – опосредованного воспалительного ответа
Презентация АГ дендритными
Краткая характеристика T h1 – опосредованного воспалительного ответа
Презентация АГ дендритными

Этапы воспалительного T h1 –опосредованного ответа(ГЗТ)
В очаге инфицирования дендритные клетки поглощают
Этапы воспалительного T h1 –опосредованного ответа(ГЗТ)
В очаге инфицирования дендритные клетки поглощают
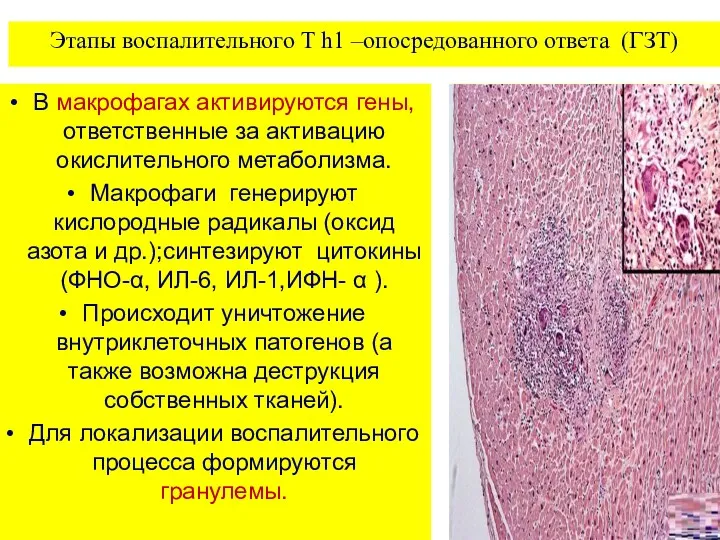
Этапы воспалительного T h1 –опосредованного ответа (ГЗТ)
В макрофагах активируются гены, ответственные
Этапы воспалительного T h1 –опосредованного ответа (ГЗТ)
В макрофагах активируются гены, ответственные
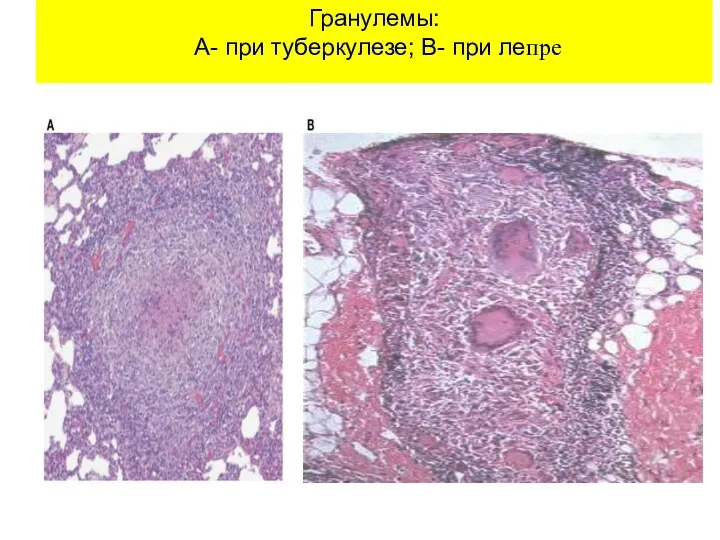
Гранулемы:
А- при туберкулезе; В- при лепре
Гранулемы:
А- при туберкулезе; В- при лепре
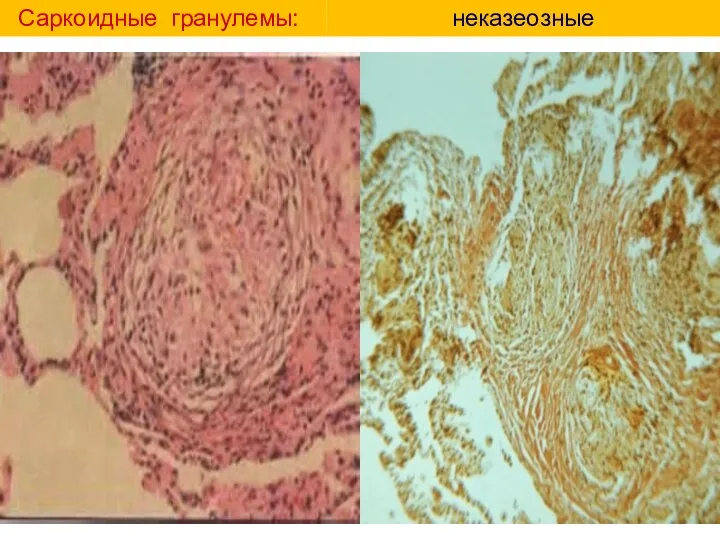
Саркоидные гранулемы:
неказеозные
Саркоидные гранулемы:
неказеозные
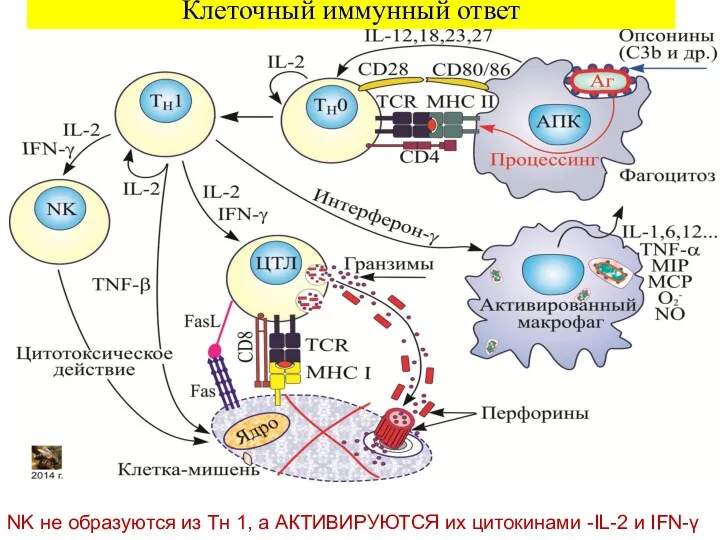
Клеточный иммунный ответ
NK не образуются из Tн 1, а АКТИВИРУЮТСЯ их
Клеточный иммунный ответ
NK не образуются из Tн 1, а АКТИВИРУЮТСЯ их
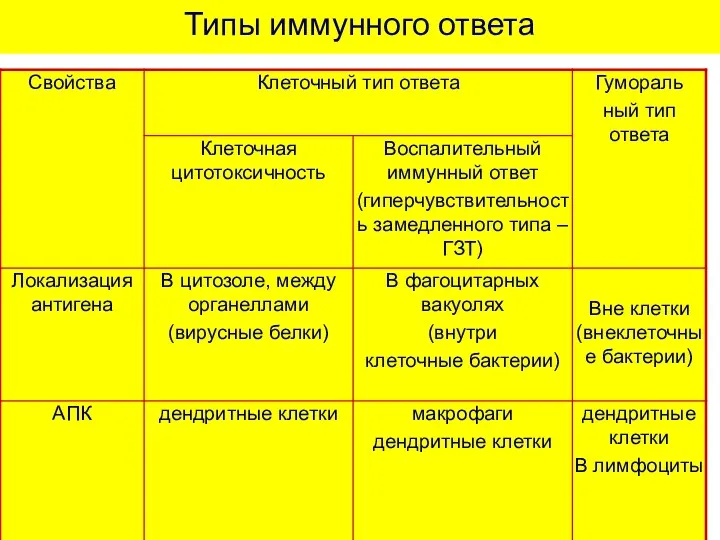
Типы иммунного ответа
Типы иммунного ответа
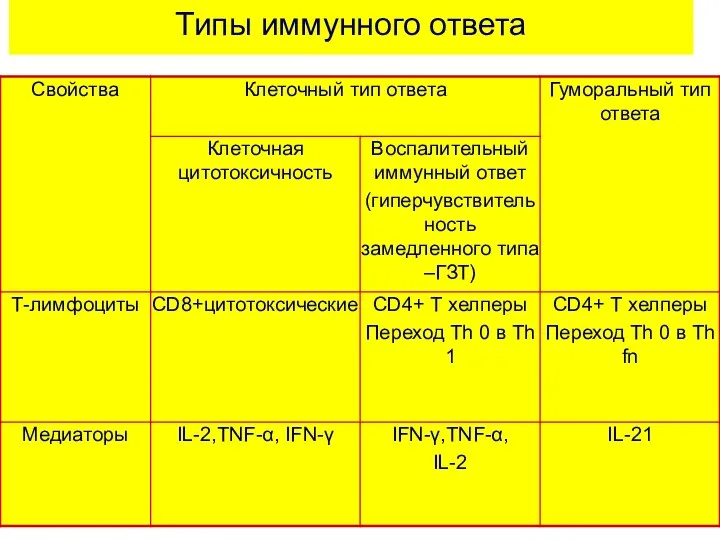
Типы иммунного ответа
Типы иммунного ответа
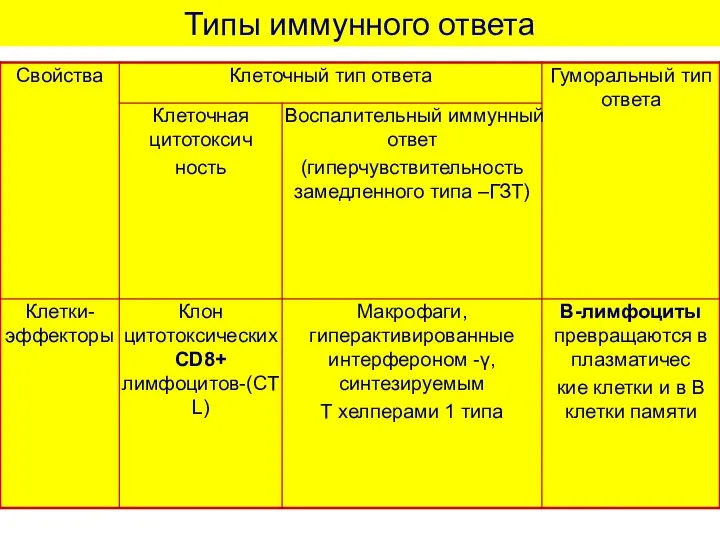
Типы иммунного ответа
Типы иммунного ответа
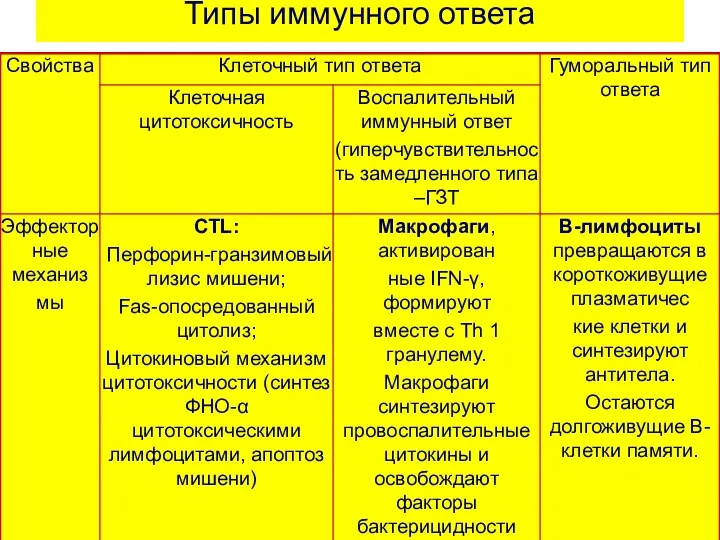
Типы иммунного ответа
Типы иммунного ответа
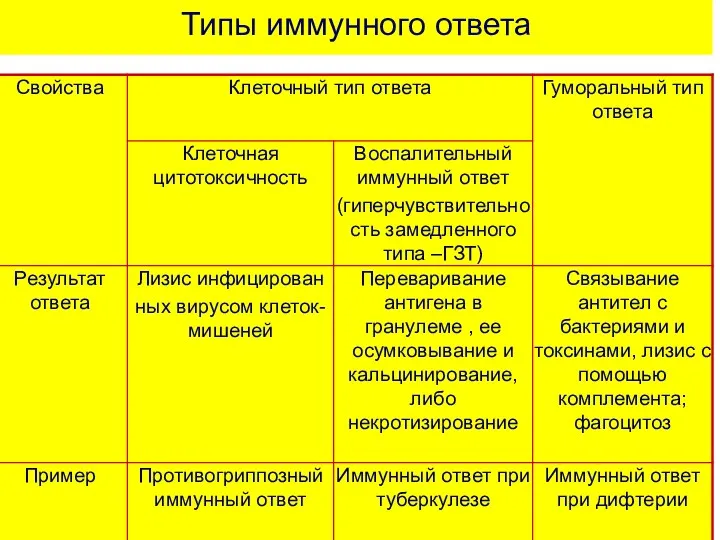
Типы иммунного ответа
Типы иммунного ответа

Вопросы
Типы иммунного ответа и факторы, их определяющие.
Внутриклеточная локализация патогенна и тип
Вопросы
Типы иммунного ответа и факторы, их определяющие.
Внутриклеточная локализация патогенна и тип

Тестовые вопросы
Антиген в Т-зависимых зонах лимфатических узлов Т-лимфоцитам представляют:
Плазматические клетки
Дендритные клетки
Эозинофилы
Нейтрофилы
Эпителиальные
Тестовые вопросы
Антиген в Т-зависимых зонах лимфатических узлов Т-лимфоцитам представляют:
Плазматические клетки
Дендритные клетки
Эозинофилы
Нейтрофилы
Эпителиальные

Тестовые вопросы
Цитотоксические Т-лимфоциты своим корецептором вступают во взаимодействие с:
Молекулами MHC I
Тестовые вопросы
Цитотоксические Т-лимфоциты своим корецептором вступают во взаимодействие с:
Молекулами MHC I

Тестовые вопросы
Цитотоксические Т-лимфоциты вызывают гибель клеток-мишеней с помощью:
Перфоринов и гранзимов
Fas-опосредованного
Тестовые вопросы
Цитотоксические Т-лимфоциты вызывают гибель клеток-мишеней с помощью:
Перфоринов и гранзимов
Fas-опосредованного

Тестовые вопросы
Цитотоксичность - это:
Способность вызывать гибель клеток-мишеней
Способность к переключению изотипов иммуноглобулинов
Многократное
Тестовые вопросы
Цитотоксичность - это:
Способность вызывать гибель клеток-мишеней
Способность к переключению изотипов иммуноглобулинов
Многократное
 Балалардағы гельминтоздар (аскаридоз, энтеробиоз, трихинеллез)
Балалардағы гельминтоздар (аскаридоз, энтеробиоз, трихинеллез) Пиодермии. Этиология. Классификация. Клиника
Пиодермии. Этиология. Классификация. Клиника Новые наркотики: эффект и побочные действия
Новые наркотики: эффект и побочные действия Естественное вскармливание. Преимущества. Питание, режим и гигиена кормящей матери
Естественное вскармливание. Преимущества. Питание, режим и гигиена кормящей матери Правовое обеспечение медицинской деятельности
Правовое обеспечение медицинской деятельности Гельмінтози. Механізми зараження
Гельмінтози. Механізми зараження Современные подходы к антибактериальной терапии
Современные подходы к антибактериальной терапии Основы ЭКГ
Основы ЭКГ Тиреотоксикоз жаңа туған балалардан бастап
Тиреотоксикоз жаңа туған балалардан бастап Аллергический ринит. Острый экзогенный аллергический альвеолит
Аллергический ринит. Острый экзогенный аллергический альвеолит Дія електричного струму на організм людини. Електротравма. Надання домедичної допомоги при ураженні електричним струмом
Дія електричного струму на організм людини. Електротравма. Надання домедичної допомоги при ураженні електричним струмом Миксовирусы. Вирусологическая диагностика гриппа, парагриппа, кори
Миксовирусы. Вирусологическая диагностика гриппа, парагриппа, кори Здоровый образ жизни
Здоровый образ жизни Грипп
Грипп Азаматтарға еңбекке жарамсыздық парақтарын беру ережесi. Жалпы ережелер
Азаматтарға еңбекке жарамсыздық парақтарын беру ережесi. Жалпы ережелер Сестринский процесс при гнойных заболеваниях легких
Сестринский процесс при гнойных заболеваниях легких Остеопороз и имплантация
Остеопороз и имплантация Пухлини гемопоетичної і лімфоїдної тканини. Лейкози і лімфоми
Пухлини гемопоетичної і лімфоїдної тканини. Лейкози і лімфоми ВИЧ-инфекция. Основные моменты
ВИЧ-инфекция. Основные моменты Жыныс функциясының бұзылыстары. Климакс
Жыныс функциясының бұзылыстары. Климакс Медицинская консультация 2010. 2 часть
Медицинская консультация 2010. 2 часть Тканевые паразиты человека. Трихомониаз, лямблиоз, балантидиаз
Тканевые паразиты человека. Трихомониаз, лямблиоз, балантидиаз Механизмы памяти
Механизмы памяти Балалардың жақ-бет аймағындағы деформациялар мен ақауларды емдеудің негізгі әдістері.Ортодонтиялық емге жастық көрсеткіштер
Балалардың жақ-бет аймағындағы деформациялар мен ақауларды емдеудің негізгі әдістері.Ортодонтиялық емге жастық көрсеткіштер Гипер – IgE синдромы (Джоб синдромы)
Гипер – IgE синдромы (Джоб синдромы) ТШҚҰ – ның негізгі клиникалық белгілері
ТШҚҰ – ның негізгі клиникалық белгілері Корь. Симптомы кори
Корь. Симптомы кори Топографическая перкуссия легких методика и техника. Границы легких в норме и патологии
Топографическая перкуссия легких методика и техника. Границы легких в норме и патологии