- Кроветворение (гемопоэз)

Содержание
- 2. Кроветворение (гемопоэз) – многостадийный процесс дифференцировки клеточных элементов, в результате которого образуются эритроциты, лейкоциты, тромбоциты, составляющие
- 3. Органы гемопоэза Центральные (костный мозг, тимус) Периферические (селезенка, лимфатические узлы, лимфоидная ткань, ассоциированная со слизистыми (MALT-система),
- 4. Эмбриональное кроветворение I период - внеэмбриональный (с 19 дня беременности) – возникновение первичных клеток-предшественников (примитивных эритробластов)
- 5. Эмбриональное кроветворение III период – проникновение ранних Т-лимфоцитов в тимус и формирование Т-клеточной иммунной системы (9-10
- 6. После рождения и в течение всей жизни человека костный мозг является единственным кроветворным органом. У ребенка
- 7. Теории кроветворения Первая теория кроветворения принадлежит выдающемуся гистологу А.А. Максимову, работавшему в начале ХХ века в
- 8. Теории кроветворения Дуалистическая теория кроветворения Негели. Согласно данной теории в постнатальный период признается существование двух самостоятельных
- 9. Теории кроветворения Полифилетическая теория. Сторонники ее утверждали, что для каждого ростка кроветворения существует свой отдельный предшественник.
- 10. Теории кроветворения Теория «умеренного унитаризма» - 30 гг. ХХ века. Разработана проф. Папенгеймом (Германия) и проф.
- 11. Современная теория кроветворения Начало положили опыты Till&McCallock, которые перелили в хвостовую вену облученной мыши смесь костномозговых
- 12. Современная теория кроветворения Далее было показано, что клетки одной колонии, будучи перелиты следующему облученному реципиенту, воспроизводят
- 13. Типы стволовых клеток оплодотворение Тотипотентная стволовая клетка Бластоциста Плюрипотентная клетка Эмбриональная стволовая клетка Фетальная ткань Мультипотентные
- 14. Современная теория кроветворения Родоначальной клеткой является тотипотентная (полипотентная) эмбриональная стволовая клетка (I отдел); Далее образуется пул
- 16. Современная теория кроветворения Клетки IV отдела – это монопотентные коммитированные предшественники, являются родоначальными для одного ростка
- 18. Современная теория кроветворения Нормальное кроветворение поликлональное, то есть осуществляется одновременно многими клонами. Продолжительность жизни одного клона
- 19. Свойства стволовых кроветворных клеток (СКК) Высокий пролиферативный потенциал, ограниченное самоподдержание; Полипотентность - способность к дифференцировке во
- 20. Стволовые кроветворные клетки Количество СКК в костном мозге– менее 0,01%. Маркер - CD34 (+) – антигенная
- 21. Закон клеточной кинетики В единицу времени рождается и умирает одно и то же количество клеток. Апоптоз
- 22. Механизмы клеточной смерти
- 23. Морфология клеток крови на поздних стадиях апоптоза
- 24. Регуляторы апоптоза Индукторы апоптоза- ИЛ-1, ФНО, γ-ИНФ, ИЛ-4, ИЛ-10, трансформирующий фактор роста, СД95-лиганд (FasL),. Большую роль
- 25. Основные пути апоптоза Повреждение ДНК АФК p53 Bax митохондрии освобождение из митоходрий цитохрома с и других
- 26. Белок р53 р53 представляет собой белок из 393 аминокислот, выделенный в 1979 году D. Lane; В
- 27. Классификация цитокинов: Позднедействующие линейно специфические факторы; Среднедействующие линейно неспецифические факторы; Факторы, влияющие на кинетику клеточного цикла
- 28. Позднедействующие линейно специфические факторы Эритропоэтин; М-КСФ; ИЛ-5; Тромбопоэтин. Поддерживают пролиферацию и созревание коммитированных клеток-предшественников.
- 29. Среднедействующие линейно неспецифические факторы ИЛ-3; ГМ-КСФ; ИЛ-4. Эти факторы поддерживают пролиферацию полипотентных предшественников, но только после
- 30. Факторы, влияющие на кинетику клеточного цикла дремлющих примитивных предшественников Фактор стволовых клеток; Г-КСФ; ИЛ-6; ИЛ-11; ИЛ-12.
- 31. Строение костного мозга Кроветворная ткань заключена в костный чехол, которая выполняет защитную и регулирующую функцию. Клеточные
- 32. Строение костного мозга
- 33. Строение костного мозга Предшественники и развивающиеся клетки располагаются следующим образом: в центре – делящиеся и незрелые
- 34. Микроокружение – клетки и внеклеточный матрикс Клетки микроокружения: Эндотелиальные клетки Адвентициальные клетки Жировые клетки (адипоциты) Фибробласты
- 35. Микроокружение Внеклеточный матрикс: Ламинин Фибронектин Гемопектин Коллаген Тромбосподин Гликозоаминогликаны Внеклеточный матрикс обеспечивает специфическое прилипание СКК, являясь
- 36. Функции системы микроокружения: Поддерживание клеток костного мозга; индукция пролиферации и дифференцировки стволовых клеток; передача информации о
- 37. Эритропоэз Эритропоэз преставляет собой постоянный и непрерывный процесс образования и обновления клеток эритрона, главной функцией которого
- 39. Эритропоэз Развитие эритрокариоцитов происходит в эритробластных островках, которые состоят из центрально расположенного макрофага и окружающих его
- 40. Эритробластный островок – в центре макрофаг, вокруг полихроматофильные нормобласты.
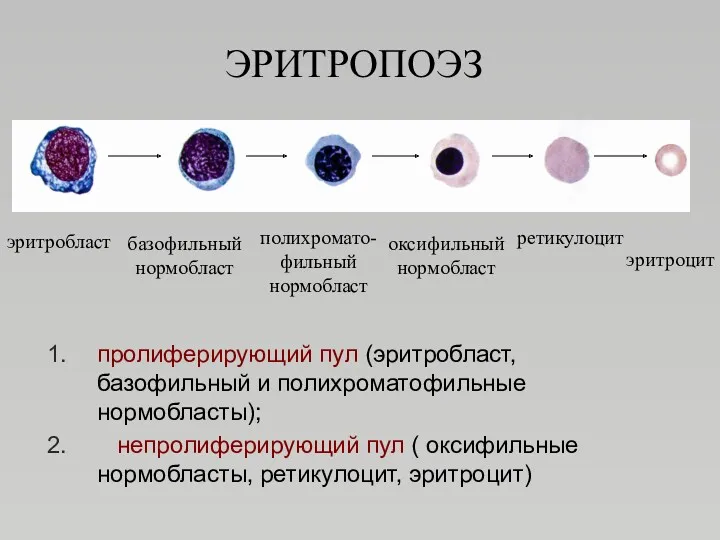
- 41. ЭРИТРОПОЭЗ эритробласт базофильный нормобласт полихромато- фильный нормобласт оксифильный нормобласт ретикулоцит эритроцит пролиферирующий пул (эритробласт, базофильный и
- 42. ЭРИТРОН Эритрон – это совокупность всех эритроидных клеток. 6% клеток системы эритрона находится в костном мозге,
- 43. Регуляция эритропоэза Регуляция эритропоэза осуществляется эритропоэтином (ЭПО). Гуморальная регуляция была доказана в 1950 году, но в
- 44. Продукция эритропоэтина Продукция ЭПО регулируется на уровне его гена и не зависит от его концентрации в
- 45. Масса эритроцитов Атмосферный кислород Сердечно-легочные функции Объем крови Концентрация гемоглобина Сродство к кислороду Эритропоэтин Почечное кровоснабжение
- 46. Место продукции эритропоэтина Основное место продукции эритропоэтина – это почки. Клетки, которые синтезируют ЭПО, располагаются в
- 47. Регуляторное действие эндокринной системы на эритропоэз Гипофиз – механизм действия модуляция продукции ЭПО в почках. Подавление
- 48. Регуляторное действие эндокринной системы на эритропоэз Надпочечники. Изменения эритропоэза при гипо- и гиперфункции коры надпочечников обусловлены
- 49. Половые железы. Андрогены стимулируют эритропоэз, а эстрогены ингибируют его. Влиянием гормонального фона объясняется разница в уровне
- 50. Эритробласт Это краеугольный камень в эритропоэзе, запрограммирован как для пролиферации, так и для синтеза, упаковки и
- 51. Кинетика эритропоэза Эритроидный костный мозг поддерживает на постоянном уровне популяцию в 25х циркулирующих эритроцитов, которые содержат
- 52. Кинетическая модель эритрона В 60 гг. 20 века была предложена кинетическая модель эритрона, которая легла в
- 53. Эритробласт Hb=21,6 пг Базоф нбл Hb= 10,8 пг Базоф нбл Hb=10,8 пг Базоф нбл Hb= 25,2
- 54. Неонатальный эритропоэз Здоровые новорожденные относительно полицитемичны. Концентрация гемоглобина менее 140 г/л в венозной крови – это
- 55. Функции эритроцитов Основная - осуществляют газообмен; определяют реологию крови; участвуют в гемостазе; транспортируют иммунные комплексы, токсины,
- 56. Тромбоцитопоэз Дифференцировка и созревание клеток мегакариоцитопоэза происходит в костном мозге, где из коммитированных, морфологически неидентифицируемых клеток-предшественников
- 57. Тромбоцитопоэз При созревании клетки проходят три морфологически дифференцируемые стадии: мегакариобласт, не превышающий 10% всей популяции, промегакариоцит
- 58. ТРОМБОЦИТОПОЭЗ А. мегакариобласт с одним овальным ядром и базофильной пенистой цитоплазмой, с характерными пузырьковидными выростами. В.
- 59. Тромбоцитопоэз Процесс преобразования мегакариобластов в мегакариоциты продолжается около 25 часов. Время созревания мегакариоцита составляет примерно 25
- 60. Тромбоцитопоэз Мегакариоциты располагаются в костном мозге вблизи костномозговых синусов и его цитоплазматические отростки через миграционные поры
- 61. Тромбоцитопоэз Отличительной чертой клеточных элементов мегакариоцитопоэза является их способность к эндомитозу (полиплоидизации) - делению ядра без
- 62. Значение плоидности мегакариоцитов Установлено, что образование тромбоцитов наблюдается в мегакариоцитах, начиная с плоидности 8N. Существует 2
- 63. Тромбоцитопоэз Созревание мегакариоцитарных элементов сопровождается накоплением в цитоплазме гранул. В гранулах мегакариоцитов содержится значительное количество белков:
- 64. Факторы, регулирующие тромбоцитопоэз Основными регуляторами, стимулирующими мегакариоцитопоэз, являются ИЛ-1, ИЛ-3, ИЛ-4, ИЛ-6, ИЛ-11, фактор стволовых клеток,
- 65. Кинетика тромбоцитов При нормальном содержании тромбоцитов в крови (200-400х10(9)/л в сутки разрушается 100х10(9)/л. Существует селезеночный пул
- 66. Связь числа циркулирующих тромбоцитов и их размера Существует обратное отношение числа тромбоцитов и их величины. При
- 67. Функции тромбоцитов Ангиотрофическая; Адгезивно-аггрегационная; Участие в процессах свертывания и фибринолиза; Ретракция кровяного сгустка; Участие в иммунной
- 68. Строение костного мозга Лимфоциты и моноциты располагаются вокруг ветвей артериальных сосудов. Гранулоциты располагаются в отдалении от
- 69. Гранулоцитопоэз КОЕ-Г (колониеобразующая единица гранулоцитопоэза) формирует пул пролиферирующих гранулоцитов, состоящий из миелобластов, промиелоцитов и миелоцитов. Все
- 70. Гранулоцитопоэз Другой пул, образующийся в костном мозге, - это непролиферирующие (созревающие) клетки - метамиелоциты, палочкоядерные и
- 71. Гранулоцитопоэз Процесс формирования зрелого гранулоцита из миелобласта осуществляется в костном мозге в течение 10-13 дней. Регуляция
- 72. ГРАНУЛОЦИТОПОЭЗ
- 73. Нейтрофилы составляют 60-70% общего числа лейкоцитов крови. После выхода нейтрофильных гранулоцитов из костного мозга в периферическую
- 74. Кинетика нейтрофилов В течение суток вырабатывается до 4х на 1 кг массы тела человека. Количество нейтрофилов
- 75. Выход нейтрофилов из резерва: Мобилизация и выход в циркуляцию из маргинального пула связаны с влиянием нейромедиаторов.
- 76. Функции нейтрофилов Нейтрофилы – это первая линия защиты от внешних и внутренних повреждающих агентов. Функцией нейтрофилов
- 77. Эозинофилы Образование эозинофилов из КОЕ-эозинофилов продолжается около 3-4 суток. Кинетика созревания эозинофилов в костном мозге сходна
- 78. Эозинофилы составляют 0,5-5% от всех лейкоцитов крови, циркулируют в течение 6-12 часов, после чего поступают в
- 79. Важнейшие белки эозинофилов: Большой основной белок – 9,2 кДа, составляет 50% больших гранул. Способен нейтрализовать гепарин,
- 80. Функции эозинофилов Обладая слабой фагоцитарной активностью, эозинофилы обуславливают внеклеточный цитолиз, тем самым участвуя в противогельминтном иммунитете.
- 81. Функции эозинофилов Эозинофилы предупреждают проникновение антигена в сосудистое русло, то есть генерализацию иммунного ответа. Эозинофилы завершают
- 82. Функции эозинофилов Другой функцией этих клеток является участие в реакциях гиперчувствительности немедленного типа. Эозинофилы выделяют инактивирующие
- 83. БазофилыБазофилы и тучные клетки имеют костномозговое происхождение и дифференцируются из колониеобразующих единиц тучных клеток – Mast-CFC.
- 84. Тучные клетки Тучные клетки обычно крупнее базофилов, имеют округлое ядро и много гранул, которые по составу
- 85. Функции базофилов В гранулах этих клеток содержатся гистамин, хондроитинсульфаты А и С, гепарин, серотонин, ферменты (трипсин,
- 86. Функции базофилов Активированные тучные клетки и базофилы в течение нескольких часов секретируют цитокины (ИЛ-4,13,5,3,6,8, ГМ-КСФ). Следствием
- 87. Лимфопоэз Все лимфоциты имеют единого предшественника - стволовую кроветворную клетку и проходят начальные этапы дифференцировки в
- 89. ЛИМФОЦИТЫ Созревание лимфоцитов можно подразделить на несколько этапов, причем на каждом существуют определенные клеточные антигены, называемые
- 90. Лимфопоэз Т-клеточная популяция лимфоцитов является наиболее многочисленной и нуждается в особых условиях развития, которые она находит,
- 91. Дифференцировка Т-лимфоцитов в тимусе Ранние тимоциты стадия I CD2+, CD7+, TdT+ Кортикальные тимоциты стадия II одновременная
- 92. Основные маркеры Т-лимфоцитов Основной и наиболее специфичный маркер – CD3 Менее специфичны, но свойственны Т-лимфоцитам –
- 94. Лимфопоэз Дифференцировка В-лимфоцитов осуществляется в костном мозге, где они проходят этап антигеннезависимой дифференцировки, который завершается экспрессией
- 95. Антигеннезависимый этап В-клеточной дифференцировки в костном мозге Про-В-лимфоцит: СD34+, TdT (терминальная дезоксинуклеотидилтрансфераза), CD22+ Пре-пре-В-лимфоцит CD34+, TdT,
- 96. Роль микроокружения: Большую роль в развитии В-лимфоцитов играет костномозговое микроокружение: клетки стромы; молекулы межклеточного матрикса, гуморальные
- 97. Лимфопоэз Зрелые В-лимфоциты покидают костный мозг, попадают в циркуляцию и заносятся в периферические лимфоидные органы, где
- 98. Результаты дифференцировки В-клеток: Образование наивных В-клеток; В-лимфоцитов фолликулярного центра (центроциты и центробласты) В-клеток постфолликулярной зоны (плазмоциты
- 99. В-лимфоциты Основной функцией В-лимфоцитов является реализация гуморального иммунного ответа. Основой его является активация В-клеток и их
- 101. Основные маркеры В-клеток: СD19, CD20, CD22, CD79a
- 102. Естественные киллеры (NK-клетки) фракция лимфоцитов, лишенных маркеров Т- и В-клеток. Наиболее важными факторами, способствующими дифференцировке естественных
- 103. Естественные киллеры (NK-клетки) В периферической крови на долю NK-клеток приходится от 5 до 25% лимфоцитов. Морфология
- 104. ЛИМФОПОЭЗ Лимфоидная стволовая клетка (?) TdT CD34 В-предшественник лимфобласт CD34 CD19 CD10 Наивная В-клетка CD19 CD20
- 105. Моноцитопоэз Костный мозг – монобласты, промоноциты (проходят 2-3 стадии деления), моноциты, которые сразу выходят в кровоток.
- 106. Моноцитопоэз Время циркуляции в периферической крови – 36-104 часа; существует 2 пула – циркулирующий и пристеночный,
- 107. В тканях МФ образуют 2 популяции клеток: Фагоцитирующие элементы (альвеолярные макрофаги, клетки Купфера, остеокласты) – обладают
- 108. СИСТЕМА МОНОНУКЛЕАРНЫХ ФАГОЦИТОВ
- 109. Функции моноцитов/макрофагов Макрофагальная система в целом рассматривается как своеобразный биологический фильтр крови и лимфы, удаляющий из
- 110. Функции моноцитов/макрофагов Макрофаги регулируют гемопоэз. Макрофаги участвуют в регуляции гемостаза – активированные клетки синтезируют прокоагулянтные факторы,
- 112. Скачать презентацию

Кроветворение (гемопоэз) – многостадийный процесс дифференцировки клеточных элементов, в результате которого
Кроветворение (гемопоэз) – многостадийный процесс дифференцировки клеточных элементов, в результате которого
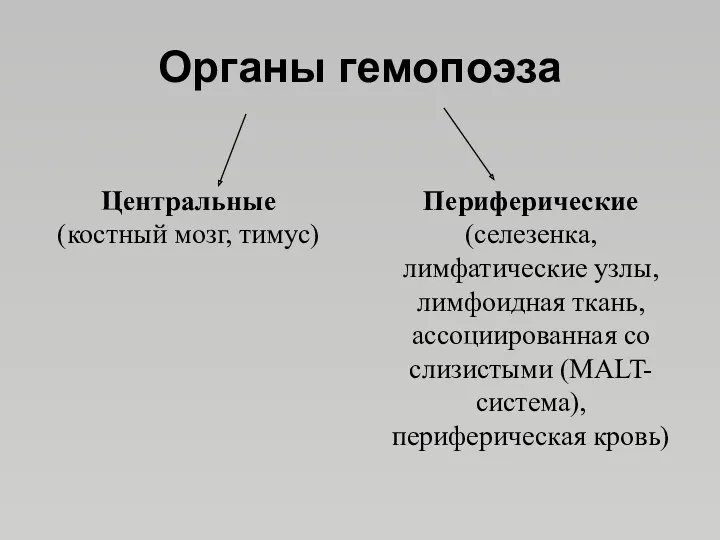
Органы гемопоэза
Центральные
(костный мозг, тимус)
Периферические
(селезенка, лимфатические узлы,
лимфоидная ткань, ассоциированная со слизистыми
Органы гемопоэза
Центральные
(костный мозг, тимус)
Периферические
(селезенка, лимфатические узлы,
лимфоидная ткань, ассоциированная со слизистыми

Эмбриональное кроветворение
I период - внеэмбриональный (с 19 дня беременности) – возникновение
Эмбриональное кроветворение
I период - внеэмбриональный (с 19 дня беременности) – возникновение

Эмбриональное кроветворение
III период – проникновение ранних Т-лимфоцитов в тимус и формирование
Эмбриональное кроветворение
III период – проникновение ранних Т-лимфоцитов в тимус и формирование
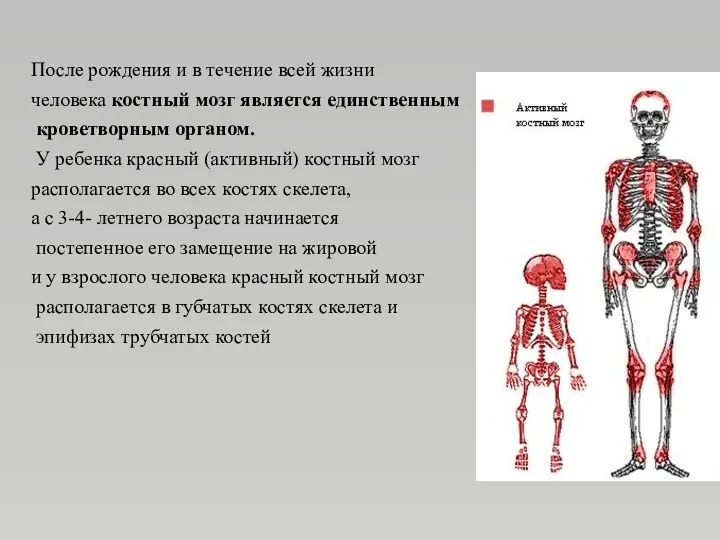
После рождения и в течение всей жизни
человека костный мозг
После рождения и в течение всей жизни
человека костный мозг

Теории кроветворения
Первая теория кроветворения принадлежит выдающемуся гистологу А.А. Максимову, работавшему в
Теории кроветворения
Первая теория кроветворения принадлежит выдающемуся гистологу А.А. Максимову, работавшему в

Теории кроветворения
Дуалистическая теория кроветворения Негели.
Согласно данной теории в постнатальный период
Теории кроветворения
Дуалистическая теория кроветворения Негели.
Согласно данной теории в постнатальный период

Теории кроветворения
Полифилетическая теория.
Сторонники ее утверждали, что для каждого ростка кроветворения существует
Теории кроветворения
Полифилетическая теория.
Сторонники ее утверждали, что для каждого ростка кроветворения существует

Теории кроветворения
Теория «умеренного унитаризма» - 30 гг. ХХ века. Разработана проф.
Теории кроветворения
Теория «умеренного унитаризма» - 30 гг. ХХ века. Разработана проф.

Современная теория кроветворения
Начало положили опыты Till&McCallock, которые перелили в хвостовую вену
Современная теория кроветворения
Начало положили опыты Till&McCallock, которые перелили в хвостовую вену

Современная теория кроветворения
Далее было показано, что клетки одной колонии, будучи перелиты
Современная теория кроветворения
Далее было показано, что клетки одной колонии, будучи перелиты
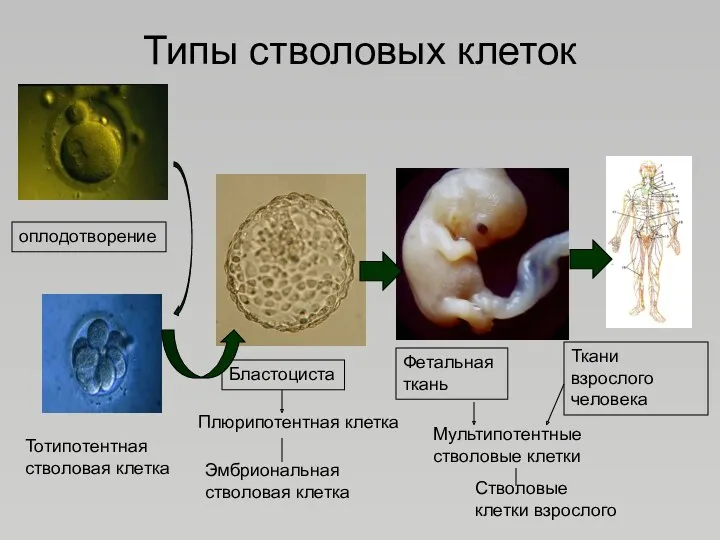
Типы стволовых клеток
оплодотворение
Тотипотентная
стволовая клетка
Бластоциста
Плюрипотентная клетка
Эмбриональная
стволовая клетка
Фетальная
ткань
Мультипотентные
стволовые клетки
Стволовые
клетки
Типы стволовых клеток
оплодотворение
Тотипотентная
стволовая клетка
Бластоциста
Плюрипотентная клетка
Эмбриональная
стволовая клетка
Фетальная
ткань
Мультипотентные
стволовые клетки
Стволовые
клетки

Современная теория кроветворения
Родоначальной клеткой является тотипотентная (полипотентная) эмбриональная стволовая клетка (I
Современная теория кроветворения
Родоначальной клеткой является тотипотентная (полипотентная) эмбриональная стволовая клетка (I
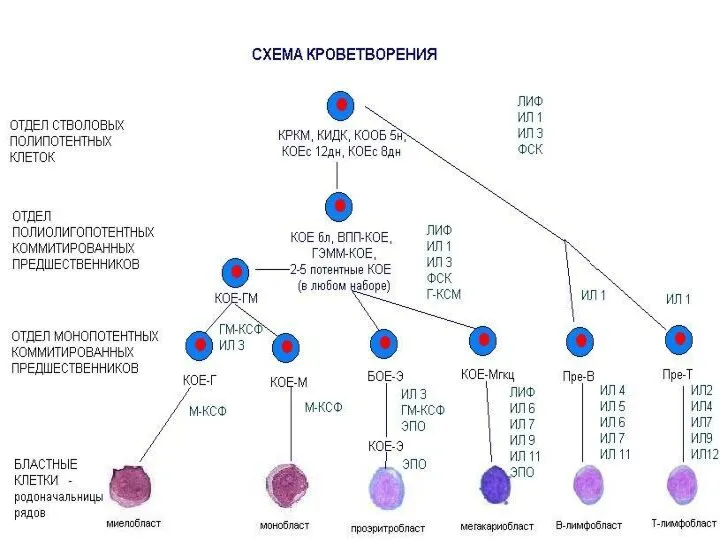

Современная теория кроветворения
Клетки IV отдела – это монопотентные коммитированные предшественники, являются
Современная теория кроветворения
Клетки IV отдела – это монопотентные коммитированные предшественники, являются
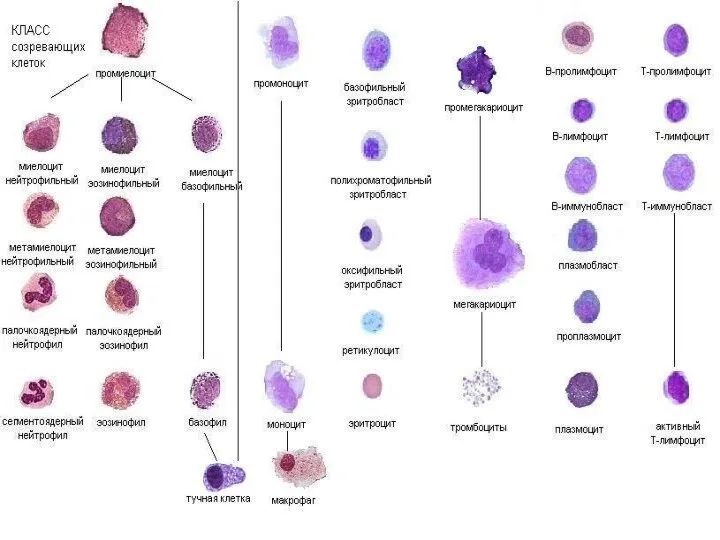

Современная теория кроветворения
Нормальное кроветворение поликлональное, то есть осуществляется одновременно многими клонами.
Продолжительность
Современная теория кроветворения
Нормальное кроветворение поликлональное, то есть осуществляется одновременно многими клонами.
Продолжительность

Свойства стволовых кроветворных клеток (СКК)
Высокий пролиферативный потенциал, ограниченное самоподдержание;
Полипотентность - способность
Свойства стволовых кроветворных клеток (СКК)
Высокий пролиферативный потенциал, ограниченное самоподдержание;
Полипотентность - способность

Стволовые кроветворные клетки
Количество СКК в костном мозге– менее 0,01%. Маркер -
Стволовые кроветворные клетки
Количество СКК в костном мозге– менее 0,01%. Маркер -

Закон клеточной кинетики
В единицу времени рождается и умирает одно и то
Закон клеточной кинетики
В единицу времени рождается и умирает одно и то
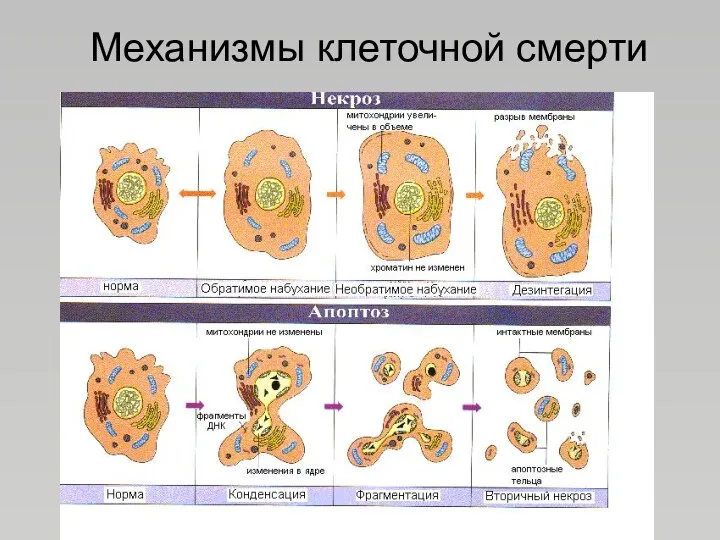
Механизмы клеточной смерти
Механизмы клеточной смерти
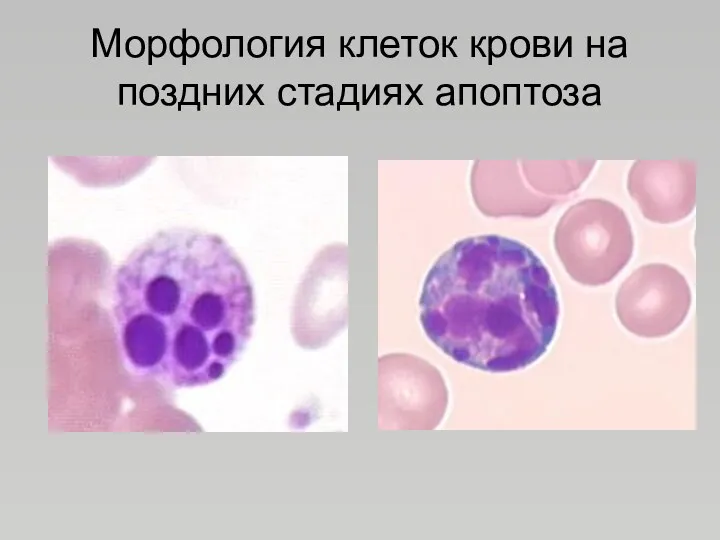
Морфология клеток крови на поздних стадиях апоптоза
Морфология клеток крови на поздних стадиях апоптоза

Регуляторы апоптоза
Индукторы апоптоза- ИЛ-1, ФНО, γ-ИНФ, ИЛ-4, ИЛ-10, трансформирующий фактор роста,
Регуляторы апоптоза
Индукторы апоптоза- ИЛ-1, ФНО, γ-ИНФ, ИЛ-4, ИЛ-10, трансформирующий фактор роста,
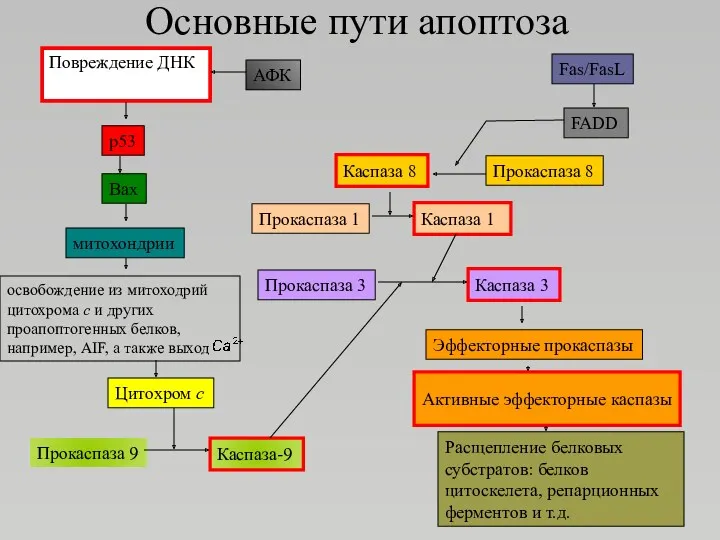
Основные пути апоптоза
Повреждение ДНК
АФК
p53
Bax
митохондрии
освобождение из митоходрий цитохрома с и других проапоптогенных
Основные пути апоптоза
Повреждение ДНК
АФК
p53
Bax
митохондрии
освобождение из митоходрий цитохрома с и других проапоптогенных

Белок р53
р53 представляет собой белок из 393 аминокислот, выделенный в 1979
Белок р53
р53 представляет собой белок из 393 аминокислот, выделенный в 1979

Классификация цитокинов:
Позднедействующие линейно специфические факторы;
Среднедействующие линейно неспецифические факторы;
Факторы, влияющие на кинетику
Классификация цитокинов:
Позднедействующие линейно специфические факторы;
Среднедействующие линейно неспецифические факторы;
Факторы, влияющие на кинетику

Позднедействующие линейно специфические факторы
Эритропоэтин;
М-КСФ;
ИЛ-5;
Тромбопоэтин.
Поддерживают пролиферацию и созревание коммитированных клеток-предшественников.
Позднедействующие линейно специфические факторы
Эритропоэтин;
М-КСФ;
ИЛ-5;
Тромбопоэтин.
Поддерживают пролиферацию и созревание коммитированных клеток-предшественников.

Среднедействующие линейно неспецифические факторы
ИЛ-3;
ГМ-КСФ;
ИЛ-4.
Эти факторы поддерживают пролиферацию полипотентных предшественников, но только
Среднедействующие линейно неспецифические факторы
ИЛ-3;
ГМ-КСФ;
ИЛ-4.
Эти факторы поддерживают пролиферацию полипотентных предшественников, но только

Факторы, влияющие на кинетику клеточного цикла дремлющих примитивных предшественников
Фактор стволовых клеток;
Г-КСФ;
ИЛ-6;
ИЛ-11;
ИЛ-12.
Механизм
Факторы, влияющие на кинетику клеточного цикла дремлющих примитивных предшественников
Фактор стволовых клеток;
Г-КСФ;
ИЛ-6;
ИЛ-11;
ИЛ-12.
Механизм

Строение костного мозга
Кроветворная ткань заключена в костный чехол, которая выполняет защитную
Строение костного мозга
Кроветворная ткань заключена в костный чехол, которая выполняет защитную
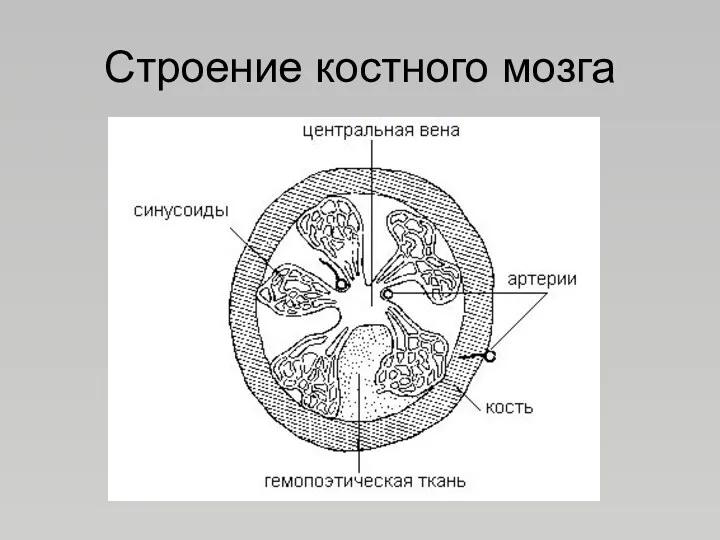
Строение костного мозга
Строение костного мозга

Строение костного мозга
Предшественники и развивающиеся клетки располагаются следующим образом: в центре
Строение костного мозга
Предшественники и развивающиеся клетки располагаются следующим образом: в центре
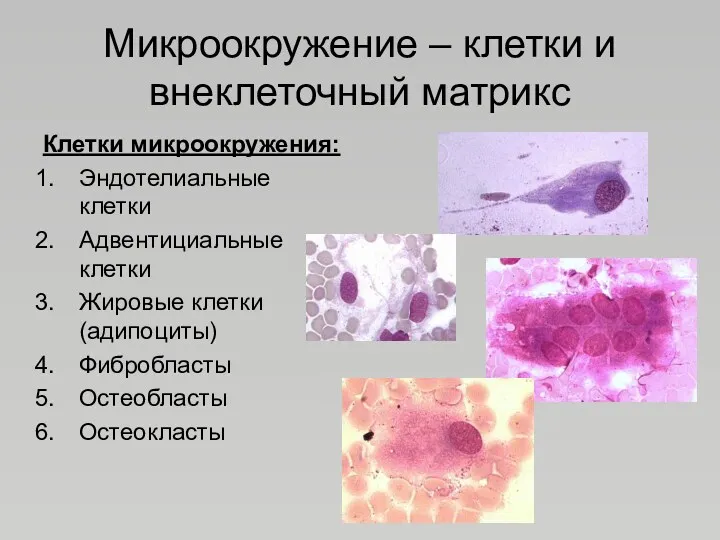
Микроокружение – клетки и внеклеточный матрикс
Клетки микроокружения:
Эндотелиальные клетки
Адвентициальные клетки
Жировые клетки (адипоциты)
Фибробласты
Остеобласты
Остеокласты
Микроокружение – клетки и внеклеточный матрикс
Клетки микроокружения:
Эндотелиальные клетки
Адвентициальные клетки
Жировые клетки (адипоциты)
Фибробласты
Остеобласты
Остеокласты

Микроокружение
Внеклеточный матрикс:
Ламинин
Фибронектин
Гемопектин
Коллаген
Тромбосподин
Гликозоаминогликаны
Внеклеточный матрикс обеспечивает специфическое прилипание СКК, являясь средой их обитания.
Микроокружение
Внеклеточный матрикс:
Ламинин
Фибронектин
Гемопектин
Коллаген
Тромбосподин
Гликозоаминогликаны
Внеклеточный матрикс обеспечивает специфическое прилипание СКК, являясь средой их обитания.

Функции системы микроокружения:
Поддерживание клеток костного мозга;
индукция пролиферации и дифференцировки стволовых клеток;
передача
Функции системы микроокружения:
Поддерживание клеток костного мозга;
индукция пролиферации и дифференцировки стволовых клеток;
передача

Эритропоэз
Эритропоэз преставляет собой постоянный и непрерывный процесс образования и обновления клеток
Эритропоэз
Эритропоэз преставляет собой постоянный и непрерывный процесс образования и обновления клеток

Эритропоэз
Развитие эритрокариоцитов происходит в эритробластных островках, которые состоят из центрально расположенного
Эритропоэз
Развитие эритрокариоцитов происходит в эритробластных островках, которые состоят из центрально расположенного
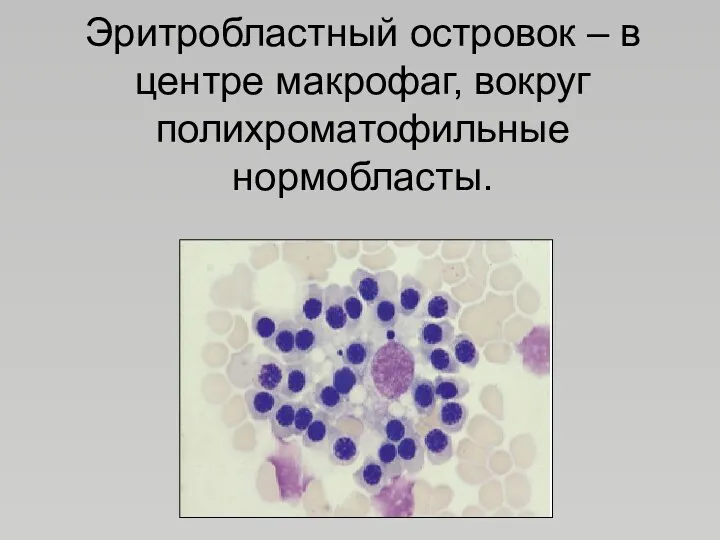
Эритробластный островок – в центре макрофаг, вокруг полихроматофильные нормобласты.
Эритробластный островок – в центре макрофаг, вокруг полихроматофильные нормобласты.
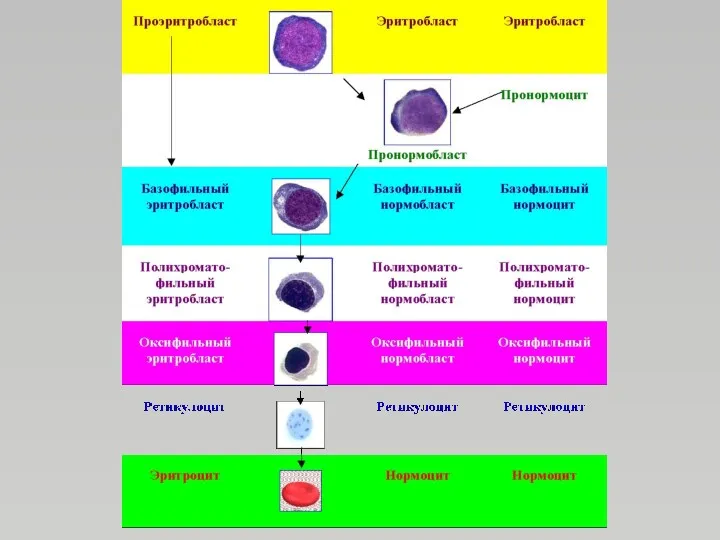
ЭРИТРОПОЭЗ
эритробласт
базофильный
нормобласт
полихромато-
фильный
нормобласт
оксифильный
нормобласт
ретикулоцит
эритроцит
пролиферирующий пул (эритробласт, базофильный и полихроматофильные нормобласты);
непролиферирующий пул (
ЭРИТРОПОЭЗ
эритробласт
базофильный
нормобласт
полихромато-
фильный
нормобласт
оксифильный
нормобласт
ретикулоцит
эритроцит
пролиферирующий пул (эритробласт, базофильный и полихроматофильные нормобласты);
непролиферирующий пул (

ЭРИТРОН
Эритрон – это совокупность всех эритроидных клеток.
6% клеток системы эритрона находится
ЭРИТРОН
Эритрон – это совокупность всех эритроидных клеток.
6% клеток системы эритрона находится

Регуляция эритропоэза
Регуляция эритропоэза осуществляется эритропоэтином (ЭПО). Гуморальная регуляция была доказана в
Регуляция эритропоэза
Регуляция эритропоэза осуществляется эритропоэтином (ЭПО). Гуморальная регуляция была доказана в

Продукция эритропоэтина
Продукция ЭПО регулируется на уровне его гена и не
Продукция эритропоэтина
Продукция ЭПО регулируется на уровне его гена и не
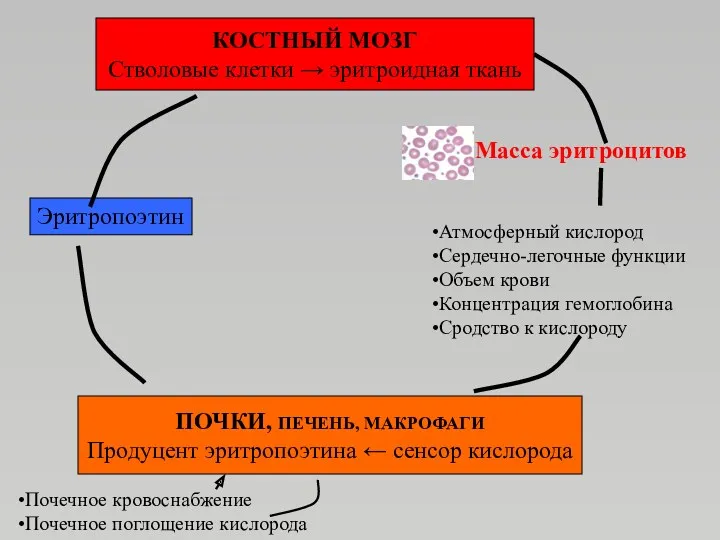
Масса эритроцитов
Атмосферный кислород
Сердечно-легочные функции
Объем крови
Концентрация гемоглобина
Сродство к кислороду
Эритропоэтин
Почечное кровоснабжение
Почечное поглощение кислорода
КОСТНЫЙ
Масса эритроцитов
Атмосферный кислород
Сердечно-легочные функции
Объем крови
Концентрация гемоглобина
Сродство к кислороду
Эритропоэтин
Почечное кровоснабжение
Почечное поглощение кислорода
КОСТНЫЙ

Место продукции эритропоэтина
Основное место продукции эритропоэтина – это почки. Клетки, которые
Место продукции эритропоэтина
Основное место продукции эритропоэтина – это почки. Клетки, которые

Регуляторное действие эндокринной системы на эритропоэз
Гипофиз – механизм действия модуляция продукции
Регуляторное действие эндокринной системы на эритропоэз
Гипофиз – механизм действия модуляция продукции

Регуляторное действие эндокринной системы на эритропоэз
Надпочечники.
Изменения эритропоэза при гипо- и
Регуляторное действие эндокринной системы на эритропоэз
Надпочечники.
Изменения эритропоэза при гипо- и

Половые железы.
Андрогены стимулируют эритропоэз, а эстрогены ингибируют его.
Влиянием гормонального
Половые железы.
Андрогены стимулируют эритропоэз, а эстрогены ингибируют его.
Влиянием гормонального

Эритробласт
Это краеугольный камень в эритропоэзе, запрограммирован как для пролиферации, так и
Эритробласт
Это краеугольный камень в эритропоэзе, запрограммирован как для пролиферации, так и

Кинетика эритропоэза
Эритроидный костный мозг поддерживает на постоянном уровне популяцию в 25х
Кинетика эритропоэза
Эритроидный костный мозг поддерживает на постоянном уровне популяцию в 25х

Кинетическая модель эритрона
В 60 гг. 20 века была предложена кинетическая модель
Кинетическая модель эритрона
В 60 гг. 20 века была предложена кинетическая модель
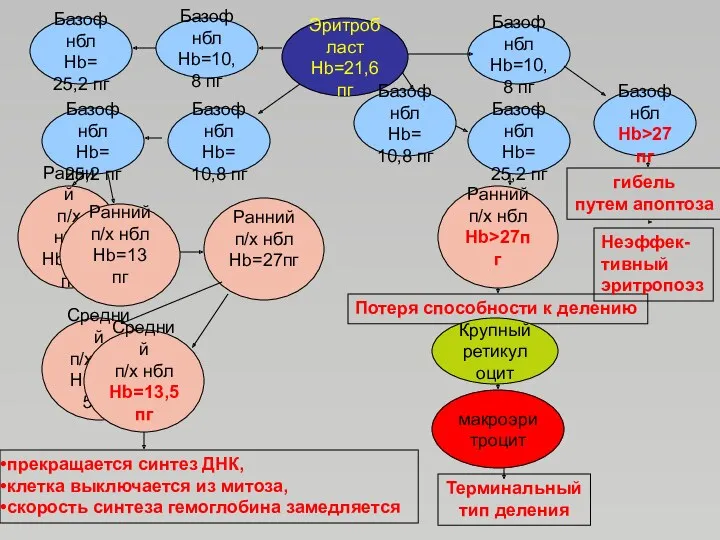
Эритробласт
Hb=21,6 пг
Базоф нбл
Hb= 10,8 пг
Базоф нбл
Hb=10,8 пг
Базоф нбл
Hb= 25,2 пг
Базоф нбл
Hb=
Эритробласт
Hb=21,6 пг
Базоф нбл
Hb= 10,8 пг
Базоф нбл
Hb=10,8 пг
Базоф нбл
Hb= 25,2 пг
Базоф нбл
Hb=

Неонатальный эритропоэз
Здоровые новорожденные относительно полицитемичны. Концентрация гемоглобина менее 140 г/л в
Неонатальный эритропоэз
Здоровые новорожденные относительно полицитемичны. Концентрация гемоглобина менее 140 г/л в

Функции эритроцитов
Основная - осуществляют газообмен;
определяют реологию крови;
участвуют в гемостазе;
транспортируют иммунные комплексы,
Функции эритроцитов
Основная - осуществляют газообмен;
определяют реологию крови;
участвуют в гемостазе;
транспортируют иммунные комплексы,

Тромбоцитопоэз
Дифференцировка и созревание клеток мегакариоцитопоэза происходит в костном мозге, где из
Тромбоцитопоэз
Дифференцировка и созревание клеток мегакариоцитопоэза происходит в костном мозге, где из

Тромбоцитопоэз
При созревании клетки проходят три морфологически дифференцируемые стадии:
мегакариобласт, не превышающий
Тромбоцитопоэз
При созревании клетки проходят три морфологически дифференцируемые стадии:
мегакариобласт, не превышающий
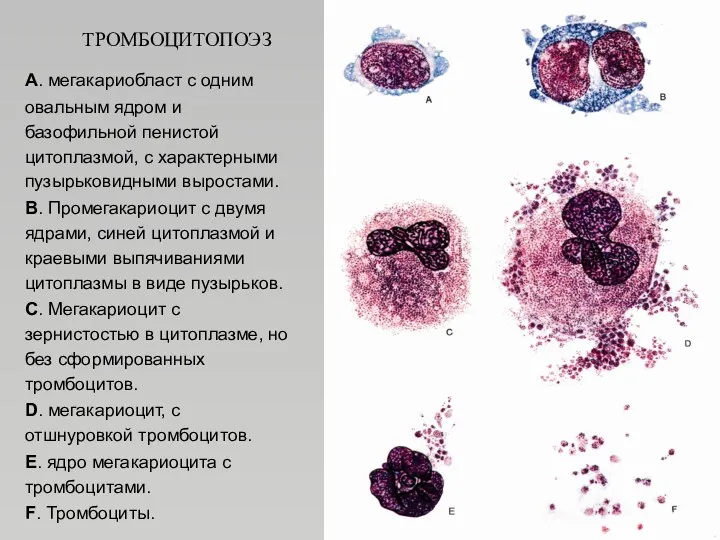
ТРОМБОЦИТОПОЭЗ
А. мегакариобласт с одним
овальным ядром и базофильной пенистой цитоплазмой, с
ТРОМБОЦИТОПОЭЗ
А. мегакариобласт с одним
овальным ядром и базофильной пенистой цитоплазмой, с

Тромбоцитопоэз
Процесс преобразования мегакариобластов в мегакариоциты продолжается около 25 часов.
Время созревания
Тромбоцитопоэз
Процесс преобразования мегакариобластов в мегакариоциты продолжается около 25 часов.
Время созревания

Тромбоцитопоэз
Мегакариоциты располагаются в костном мозге вблизи костномозговых синусов и его цитоплазматические
Тромбоцитопоэз
Мегакариоциты располагаются в костном мозге вблизи костномозговых синусов и его цитоплазматические

Тромбоцитопоэз
Отличительной чертой клеточных элементов мегакариоцитопоэза является их способность к эндомитозу (полиплоидизации)
Тромбоцитопоэз
Отличительной чертой клеточных элементов мегакариоцитопоэза является их способность к эндомитозу (полиплоидизации)

Значение плоидности мегакариоцитов
Установлено, что образование тромбоцитов наблюдается в мегакариоцитах, начиная с
Значение плоидности мегакариоцитов
Установлено, что образование тромбоцитов наблюдается в мегакариоцитах, начиная с

Тромбоцитопоэз
Созревание мегакариоцитарных элементов сопровождается накоплением в цитоплазме гранул.
В гранулах мегакариоцитов содержится
Тромбоцитопоэз
Созревание мегакариоцитарных элементов сопровождается накоплением в цитоплазме гранул.
В гранулах мегакариоцитов содержится

Факторы, регулирующие тромбоцитопоэз
Основными регуляторами, стимулирующими мегакариоцитопоэз, являются ИЛ-1, ИЛ-3, ИЛ-4, ИЛ-6,
Факторы, регулирующие тромбоцитопоэз
Основными регуляторами, стимулирующими мегакариоцитопоэз, являются ИЛ-1, ИЛ-3, ИЛ-4, ИЛ-6,

Кинетика тромбоцитов
При нормальном содержании тромбоцитов в крови (200-400х10(9)/л в сутки разрушается
Кинетика тромбоцитов
При нормальном содержании тромбоцитов в крови (200-400х10(9)/л в сутки разрушается

Связь числа циркулирующих тромбоцитов и их размера
Существует обратное отношение числа тромбоцитов
Связь числа циркулирующих тромбоцитов и их размера
Существует обратное отношение числа тромбоцитов

Функции тромбоцитов
Ангиотрофическая;
Адгезивно-аггрегационная;
Участие в процессах свертывания и фибринолиза;
Ретракция кровяного сгустка;
Участие в иммунной
Функции тромбоцитов
Ангиотрофическая;
Адгезивно-аггрегационная;
Участие в процессах свертывания и фибринолиза;
Ретракция кровяного сгустка;
Участие в иммунной

Строение костного мозга
Лимфоциты и моноциты располагаются вокруг ветвей артериальных сосудов.
Гранулоциты располагаются
Строение костного мозга
Лимфоциты и моноциты располагаются вокруг ветвей артериальных сосудов.
Гранулоциты располагаются

Гранулоцитопоэз
КОЕ-Г (колониеобразующая единица гранулоцитопоэза) формирует пул пролиферирующих гранулоцитов, состоящий из миелобластов,
Гранулоцитопоэз
КОЕ-Г (колониеобразующая единица гранулоцитопоэза) формирует пул пролиферирующих гранулоцитов, состоящий из миелобластов,

Гранулоцитопоэз
Другой пул, образующийся в костном мозге, - это непролиферирующие (созревающие) клетки
Гранулоцитопоэз
Другой пул, образующийся в костном мозге, - это непролиферирующие (созревающие) клетки

Гранулоцитопоэз
Процесс формирования зрелого гранулоцита из миелобласта осуществляется в костном мозге в
Гранулоцитопоэз
Процесс формирования зрелого гранулоцита из миелобласта осуществляется в костном мозге в
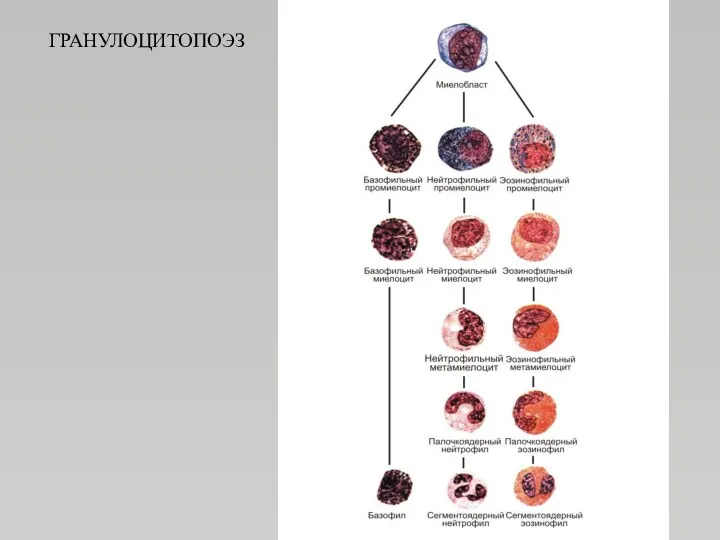
ГРАНУЛОЦИТОПОЭЗ
ГРАНУЛОЦИТОПОЭЗ

Нейтрофилы
составляют 60-70% общего числа лейкоцитов крови.
После выхода нейтрофильных гранулоцитов из
Нейтрофилы
составляют 60-70% общего числа лейкоцитов крови.
После выхода нейтрофильных гранулоцитов из

Кинетика нейтрофилов
В течение суток вырабатывается до 4х на 1 кг массы
Кинетика нейтрофилов
В течение суток вырабатывается до 4х на 1 кг массы

Выход нейтрофилов из резерва:
Мобилизация и выход в циркуляцию из маргинального пула
Выход нейтрофилов из резерва:
Мобилизация и выход в циркуляцию из маргинального пула

Функции нейтрофилов
Нейтрофилы – это первая линия защиты от внешних и внутренних
Функции нейтрофилов
Нейтрофилы – это первая линия защиты от внешних и внутренних

Эозинофилы
Образование эозинофилов из КОЕ-эозинофилов продолжается около 3-4 суток.
Кинетика созревания эозинофилов в
Эозинофилы
Образование эозинофилов из КОЕ-эозинофилов продолжается около 3-4 суток.
Кинетика созревания эозинофилов в

Эозинофилы
составляют 0,5-5% от всех лейкоцитов крови,
циркулируют в течение 6-12 часов,
Эозинофилы
составляют 0,5-5% от всех лейкоцитов крови,
циркулируют в течение 6-12 часов,

Важнейшие белки эозинофилов:
Большой основной белок – 9,2 кДа, составляет 50% больших
Важнейшие белки эозинофилов:
Большой основной белок – 9,2 кДа, составляет 50% больших

Функции эозинофилов
Обладая слабой фагоцитарной активностью, эозинофилы обуславливают внеклеточный цитолиз, тем самым
Функции эозинофилов
Обладая слабой фагоцитарной активностью, эозинофилы обуславливают внеклеточный цитолиз, тем самым

Функции эозинофилов
Эозинофилы предупреждают проникновение антигена в сосудистое русло, то есть генерализацию
Функции эозинофилов
Эозинофилы предупреждают проникновение антигена в сосудистое русло, то есть генерализацию

Функции эозинофилов
Другой функцией этих клеток является участие в реакциях гиперчувствительности немедленного
Функции эозинофилов
Другой функцией этих клеток является участие в реакциях гиперчувствительности немедленного

БазофилыБазофилы и тучные клетки
имеют костномозговое происхождение и дифференцируются из колониеобразующих единиц
БазофилыБазофилы и тучные клетки
имеют костномозговое происхождение и дифференцируются из колониеобразующих единиц
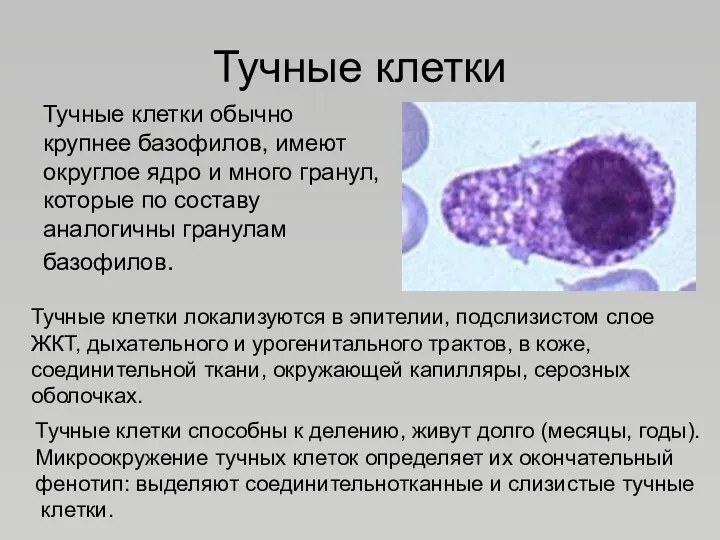
Тучные клетки
Тучные клетки обычно крупнее базофилов, имеют округлое ядро и много
Тучные клетки
Тучные клетки обычно крупнее базофилов, имеют округлое ядро и много

Функции базофилов
В гранулах этих клеток содержатся гистамин, хондроитинсульфаты А и С,
Функции базофилов
В гранулах этих клеток содержатся гистамин, хондроитинсульфаты А и С,

Функции базофилов
Активированные тучные клетки и базофилы в течение нескольких часов секретируют
Функции базофилов
Активированные тучные клетки и базофилы в течение нескольких часов секретируют

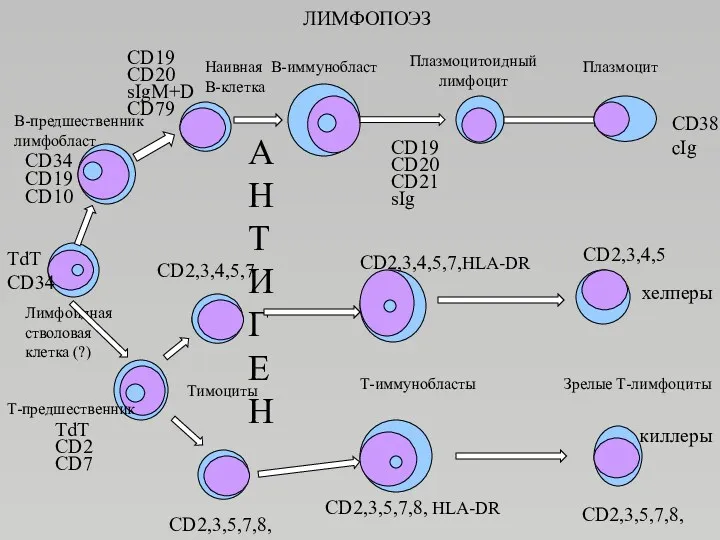
Лимфопоэз
Все лимфоциты имеют единого предшественника - стволовую кроветворную клетку и проходят
Лимфопоэз
Все лимфоциты имеют единого предшественника - стволовую кроветворную клетку и проходят
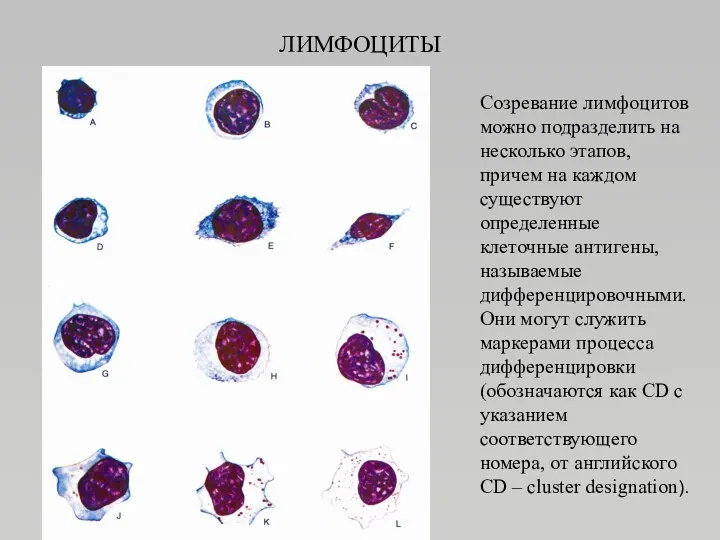
ЛИМФОЦИТЫ
Созревание лимфоцитов можно подразделить на несколько этапов, причем на каждом существуют
ЛИМФОЦИТЫ
Созревание лимфоцитов можно подразделить на несколько этапов, причем на каждом существуют

Лимфопоэз
Т-клеточная популяция лимфоцитов является наиболее многочисленной и нуждается в особых условиях
Лимфопоэз
Т-клеточная популяция лимфоцитов является наиболее многочисленной и нуждается в особых условиях

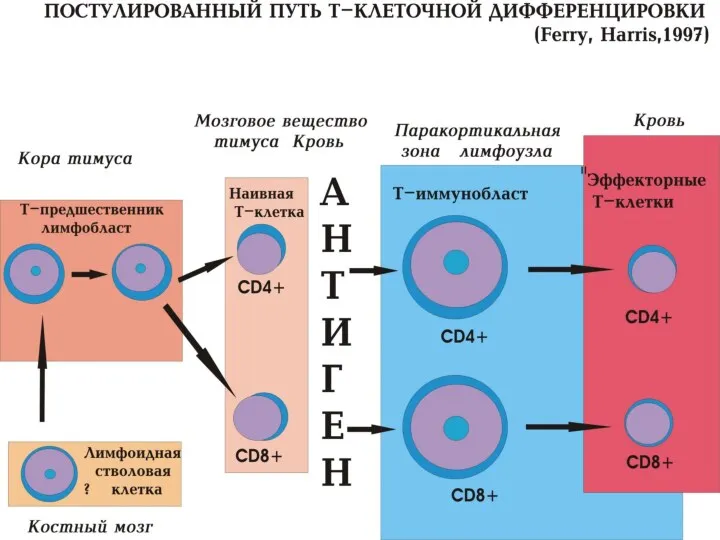
Дифференцировка Т-лимфоцитов в тимусе
Ранние тимоциты стадия I CD2+, CD7+, TdT+
Кортикальные тимоциты
Дифференцировка Т-лимфоцитов в тимусе
Ранние тимоциты стадия I CD2+, CD7+, TdT+
Кортикальные тимоциты

Основные маркеры Т-лимфоцитов
Основной и наиболее специфичный маркер – CD3
Менее специфичны, но
Основные маркеры Т-лимфоцитов
Основной и наиболее специфичный маркер – CD3
Менее специфичны, но

Лимфопоэз
Дифференцировка В-лимфоцитов осуществляется в костном мозге, где они проходят этап антигеннезависимой
Лимфопоэз
Дифференцировка В-лимфоцитов осуществляется в костном мозге, где они проходят этап антигеннезависимой

Антигеннезависимый этап В-клеточной дифференцировки в костном мозге
Про-В-лимфоцит: СD34+, TdT (терминальная дезоксинуклеотидилтрансфераза),
Антигеннезависимый этап В-клеточной дифференцировки в костном мозге
Про-В-лимфоцит: СD34+, TdT (терминальная дезоксинуклеотидилтрансфераза),

Роль микроокружения:
Большую роль в развитии В-лимфоцитов играет костномозговое микроокружение:
клетки стромы;
Роль микроокружения:
Большую роль в развитии В-лимфоцитов играет костномозговое микроокружение:
клетки стромы;

Лимфопоэз
Зрелые В-лимфоциты покидают костный мозг, попадают в циркуляцию и заносятся в
Лимфопоэз
Зрелые В-лимфоциты покидают костный мозг, попадают в циркуляцию и заносятся в

Результаты дифференцировки В-клеток:
Образование наивных В-клеток;
В-лимфоцитов фолликулярного центра (центроциты и центробласты)
В-клеток постфолликулярной
Результаты дифференцировки В-клеток:
Образование наивных В-клеток;
В-лимфоцитов фолликулярного центра (центроциты и центробласты)
В-клеток постфолликулярной

В-лимфоциты
Основной функцией В-лимфоцитов является реализация гуморального иммунного ответа. Основой его является
В-лимфоциты
Основной функцией В-лимфоцитов является реализация гуморального иммунного ответа. Основой его является
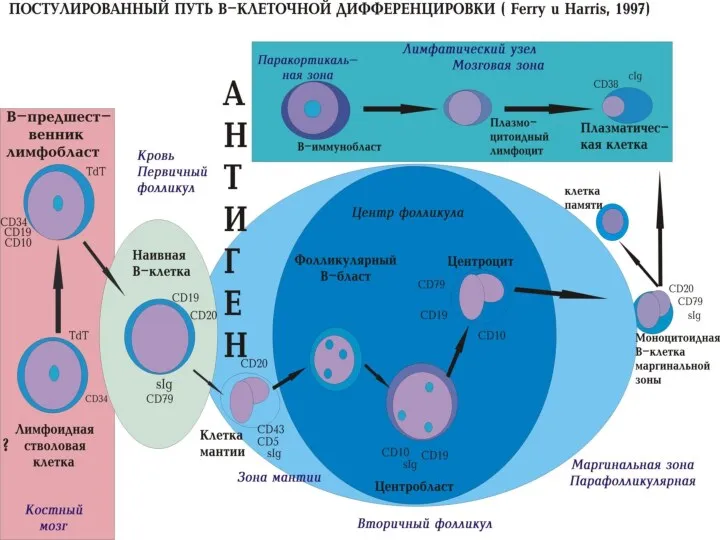

Основные маркеры В-клеток:
СD19, CD20, CD22, CD79a
Основные маркеры В-клеток:
СD19, CD20, CD22, CD79a

Естественные киллеры (NK-клетки)
фракция лимфоцитов, лишенных маркеров Т- и В-клеток.
Наиболее важными
Естественные киллеры (NK-клетки)
фракция лимфоцитов, лишенных маркеров Т- и В-клеток.
Наиболее важными

Естественные киллеры (NK-клетки)
В периферической крови на долю NK-клеток приходится от 5
Естественные киллеры (NK-клетки)
В периферической крови на долю NK-клеток приходится от 5
ЛИМФОПОЭЗ
Лимфоидная
стволовая
клетка (?)
TdT
CD34
В-предшественник
лимфобласт
CD34
CD19
CD10
Наивная
В-клетка
CD19
CD20
sIgM+D
CD79
А
Н
Т
И
Г
Е
Н
В-иммунобласт
Плазмоцитоидный
лимфоцит
Плазмоцит
CD38
cIg
CD19
CD20
CD21
sIg
Т-предшественник
TdT
CD2
CD7
Тимоциты
Т-иммунобласты
Зрелые Т-лимфоциты
CD2,3,4,5,7
CD2,3,5,7,8,
CD2,3,4,5
CD2,3,4,5,7,HLA-DR
CD2,3,5,7,8, HLA-DR
CD2,3,5,7,8,
киллеры
хелперы
ЛИМФОПОЭЗ
Лимфоидная
стволовая
клетка (?)
TdT
CD34
В-предшественник
лимфобласт
CD34
CD19
CD10
Наивная
В-клетка
CD19
CD20
sIgM+D
CD79
А
Н
Т
И
Г
Е
Н
В-иммунобласт
Плазмоцитоидный
лимфоцит
Плазмоцит
CD38
cIg
CD19
CD20
CD21
sIg
Т-предшественник
TdT
CD2
CD7
Тимоциты
Т-иммунобласты
Зрелые Т-лимфоциты
CD2,3,4,5,7
CD2,3,5,7,8,
CD2,3,4,5
CD2,3,4,5,7,HLA-DR
CD2,3,5,7,8, HLA-DR
CD2,3,5,7,8,
киллеры
хелперы
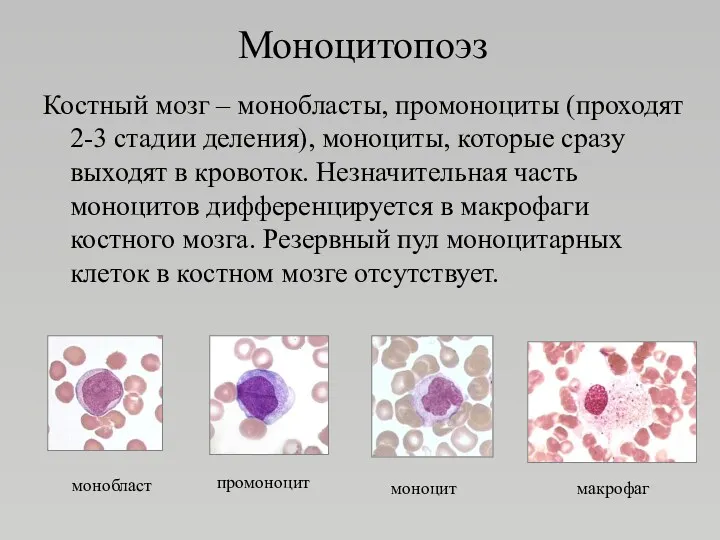
Моноцитопоэз
Костный мозг – монобласты, промоноциты (проходят 2-3 стадии деления), моноциты, которые
Моноцитопоэз
Костный мозг – монобласты, промоноциты (проходят 2-3 стадии деления), моноциты, которые

Моноцитопоэз
Время циркуляции в периферической крови – 36-104 часа; существует 2 пула
Моноцитопоэз
Время циркуляции в периферической крови – 36-104 часа; существует 2 пула

В тканях МФ образуют 2 популяции клеток:
Фагоцитирующие элементы (альвеолярные макрофаги, клетки
В тканях МФ образуют 2 популяции клеток:
Фагоцитирующие элементы (альвеолярные макрофаги, клетки

СИСТЕМА МОНОНУКЛЕАРНЫХ ФАГОЦИТОВ
СИСТЕМА МОНОНУКЛЕАРНЫХ ФАГОЦИТОВ

Функции моноцитов/макрофагов
Макрофагальная система в целом рассматривается как своеобразный биологический фильтр крови
Функции моноцитов/макрофагов
Макрофагальная система в целом рассматривается как своеобразный биологический фильтр крови

Функции моноцитов/макрофагов
Макрофаги регулируют гемопоэз.
Макрофаги участвуют в регуляции гемостаза – активированные клетки
Функции моноцитов/макрофагов
Макрофаги регулируют гемопоэз.
Макрофаги участвуют в регуляции гемостаза – активированные клетки
 Особенности реализации плана системного ухода за больными взрослыми и детьми терапевтического профиля
Особенности реализации плана системного ухода за больными взрослыми и детьми терапевтического профиля Патофизиология боли
Патофизиология боли Сифіліс. Класифікація сифілісу
Сифіліс. Класифікація сифілісу Психологические особенности лиц с нарушениями речи
Психологические особенности лиц с нарушениями речи Диагностика анемий
Диагностика анемий Артериальная гипертензия при беременности
Артериальная гипертензия при беременности Визначення чинників розвитку безпліддя у пацієнтів та аналіз факторів, які його викликають
Визначення чинників розвитку безпліддя у пацієнтів та аналіз факторів, які його викликають Методы исследования детей с различными заболеваниями сердца
Методы исследования детей с различными заболеваниями сердца Структура эпидемиологического метода исследования
Структура эпидемиологического метода исследования Врачебные ошибки и иные формы причинения вреда здоровью пациента
Врачебные ошибки и иные формы причинения вреда здоровью пациента Анатомо-физиологические особенности кожи у детей
Анатомо-физиологические особенности кожи у детей Лимфатическая система
Лимфатическая система Регуляция менструального цикла
Регуляция менструального цикла Основы организации стационарной помощи населению в РК
Основы организации стационарной помощи населению в РК WBC pathology. (Subject 11)
WBC pathology. (Subject 11) Рациональное питание, его роль в жизни человека. Гигиена питания
Рациональное питание, его роль в жизни человека. Гигиена питания Нарушения ритма сердца
Нарушения ритма сердца Процессный анализ в деятельности врача. Обеспечение безопасности пациентов
Процессный анализ в деятельности врача. Обеспечение безопасности пациентов Организация лечебно-эвакуационных мероприятий
Организация лечебно-эвакуационных мероприятий Приобретенная апластическая анемия
Приобретенная апластическая анемия Пульс. Микроциркуляция
Пульс. Микроциркуляция Электрокардиография. Методика записи ЭКГ
Электрокардиография. Методика записи ЭКГ Тактика ведения ВИЧ-инфицированных пациентов с туберкулезом. Лечение туберкулеза у ВИЧ-инфицированных
Тактика ведения ВИЧ-инфицированных пациентов с туберкулезом. Лечение туберкулеза у ВИЧ-инфицированных Заболевания шеи, гортани, пищевода
Заболевания шеи, гортани, пищевода Заболевания эндокринной системы
Заболевания эндокринной системы Сосудистая деменция
Сосудистая деменция Туберкулезді менингит
Туберкулезді менингит Общие правила оказания первой помощи
Общие правила оказания первой помощи