- Механизмы врожденного иммунитета

Содержание
- 2. План лекции 1. КОМПОНЕНТЫ ВРОЖДЕННОГО ИММУНИТЕТА 2. ПОНЯТИЕ О РЕЦЕПТОРАХ РАСПОЗНАВАНИЯ ЧУЖОГО 3. РОЛЬ НАТУРАЛЬНЫХ КИЛЛЕРОВ
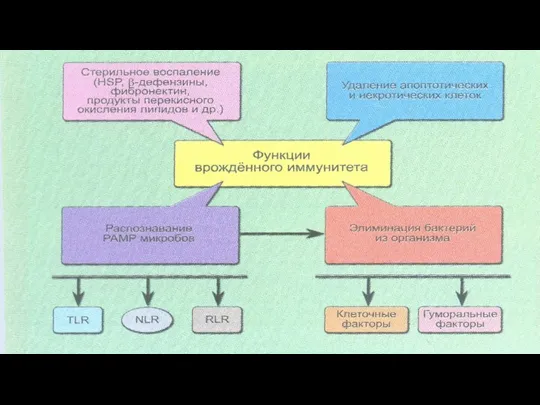
- 3. ФАКТОРЫ ВРОЖДЕННОГО ИММУНИТЕТА КЛЕТОЧНЫЕ ФАКТОРЫ: ФАГОЦИТОЗ НАТУРАЛЬНЫЕ КИЛЛЕРЫ ГУМОРАЛЬНЫЕ ФАКТОРЫ: СИСТЕМА КОМПЛЕМЕНТА СИСТЕМА ИНТЕРФЕРОНОВ
- 5. Факторы врожденного иммунитета Фагоциты Система комплемента Естественные киллеры Антитела – иммуноглобулины Интерфероны Цитокины Т-лимфоциты В-лимфоциты Факторы
- 6. ОСНОВНОЕ СВОЙСТВО ВРОЖДЕННОГО ИММУНИТЕТА НЕСПЕЦИФИЧНОСТЬ В РАСПОЗНАВАНИИ АНТИГЕНА МОЛЕКУЛЫ И КЛЕТКИ ВРОЖДЕННОГО ИММУНИТЕТА НЕ ВЫРАБАТЫВАЮТ ИММУНОЛОГИЧСКОЙ
- 7. Система врожденного иммунитета должна иметь возможность отличать свое от чужого Фагоциты могут различать сходные структурные компоненты
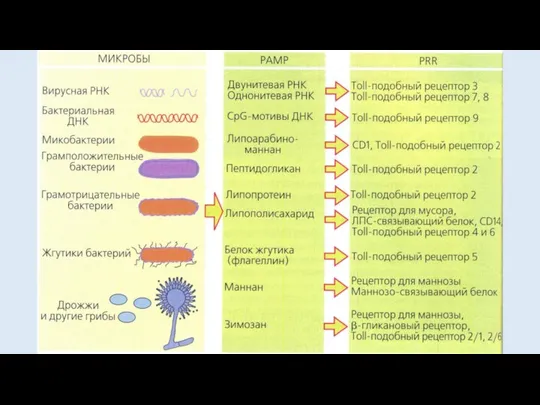
- 8. ПАТТЕРН-РАСПОЗНАЮЩИЕ РЕЦЕПТОРЫ PRR -расположены на клеточной мембране фагоцитов и дендритных клеток Примерами PAMP являются пептидогликаны клеточной
- 9. PRR можно разделить на 3 группы рецепторы передачи сигналов рецепторы эндоцитоза секретируемые рецепторы
- 10. РЕЦЕПТОРЫ ПЕРЕДАЧИ СИГНАЛОВ TOLL-ПОДОБНЫЕ РЕЦЕПТОРЫ (toll-like receptor) сигнализируют о появлении чужого имеются на мембранах фагоцитов, дендритных
- 11. ЭНДОЦИТОЗНЫЕ, МЕМБРАННЫЕ PRR Экспрессированы на поверхности фагоцитов Отвечают за поглощение и транспорт веществ, бактерий, вирусов к
- 12. СЕКРЕТИРУЕМЫЕ, РАСТВОРИМЫЕ, PRR Это опсонины, которые «обволакивают» поверхность антигена, облегчая фагоцитоз: Компонент С1 комплемента С-реактивный белок
- 14. У человека установлены типы TLR 1 – 11 типов Все известные TLR представляют собой одноцепочечные трансмембранные
- 15. После связывания Toll- подобных рецепторов с соответствующими лигандами микроорганизмов различного происхождения происходит цепь внутриклеточных событий.
- 16. Эти структуры взаимодействуют со специализированными паттерн-распознающими (PRR) рецепторами на поверхности клеток доиммунного воспаления (макрофаг, дендритные клетки).
- 17. ФАГОЦИТОЗ ЭТО ПОГЛОЩЕНИЕ ГРАНУЛОЦИТАМИ (НЕЙТРОФИЛАМИ, ЭОЗИНОФИЛАМИ, БАЗОФИЛАМИ) И АГРАНУЛОЦИТАМИ (МОНОЦИТАМИ/МАКРОФАГАМИ) ЧАСТИЦ ДИАМЕТРОМ БОЛЕЕ 0,1 МКМ
- 18. Хемотаксис
- 19. Характеристики фагоцитирующих клеток
- 22. Условно весь процесс принято делить на несколько этапов. Первым из них считается хемотаксическое перемещение фагоцитирующей клетки
- 23. Наиболее типичными хемоаттрактантами собственного происхождения являются: медиаторы воспаления (лейкотриен В4, гистамин и др.), продукты активации системы
- 24. Для этих веществ на поверхности фагоцитирующих клеток имеются специфические рецепторы, присоединение к которым действующего агента вызывает
- 25. Клетка из округлой становится треугольной, обращенной в сторону движения. В части цитоплазмы уменьшается количество органелл и
- 26. Связывание фагоцита и чужеродного объекта. Стадия прилипания включает опсонизацию, распознавание и прикрепление фагоцита к объекту фагоцитоза.
- 27. Существует 2 основных варианта опсонизации: в одном случае опсонизирующие агенты — антитела класса IgG, а в
- 28. РЕЦЕПТОРЫ ДЛЯ РАСПОЗНАВАНИЯ ОПСОНИНОВ (FC- И C3-РЕЦЕПТОРЫ) Распознавание патогенов и других клеток, опсонизированных антите- лами класса
- 29. С3-рецепторы играют важную роль в контроле активации комплемента, ингибируя связывание С3 с поверхностью собственных клеток и
- 30. СТАДИЯ АКТИВАЦИИ МЕМБРАНЫ На начальных стадиях фагоцитоза основные события происходят на обращенном к мишени участке поляризованной
- 31. Следующий этап активации направлен на образование продуктов, участвующих в полимеризации актина — процесс, на котором основан
- 32. СТАДИИ ПОГРУЖЕНИЯ И ОБРАЗОВАНИЯ ФАГОСОМЫ. Погружение частицы обусловлено сокращением нитей актина, сконцентрированных вокруг фагоцитарной чаши. Погружение
- 33. Сразу после образования фагосома не несет бактерицидных веществ и ферментов, способных разрушить патоген. Показатель созревания фагосомы
- 34. СЛИЯНИЕ ФАГОСОМЫ И ЛИЗОСОМЫ Решающий вклад в созревание фагосом и обретение ими способности убивать и расщеплять
- 35. Стадия киллинга обеспечивается наличием в фагоцитах бактерицидных факторов, которые выделяются в фагосому или окружающей объект среду
- 36. Кислородзависимые механизмы связаны с образованием активных форм кислорода, обеспечивающих микробицидное действие К ним относят: Продукты «респираторного
- 37. Образующиеся активные радикалы обусловливают повреждение и деструкцию белков и липидов мембран, нуклеиновых кислот и других химических
- 38. ВЫБРОС ФАГОЦИТАМИ ПРОДУКТОВ ДЕГРАДАЦИИ (ДЕГРАНУЛЯЦИЯ) Заключительная фаза фагоцитоза — выброс содержимого фаголизосом путем дегрануляции. За счет
- 39. При незавершенном фагоцитозе чужеродные клетки, поглощенные фагоцитами, остаются жизнеспособными. Некоторые микроорганизмы, снабжённые капсулами или плотными гидрофобными
- 40. Незавершенный фагоцитоз Примером незавершенного фагоцитоза является поглощение гонококков в организме, не имеющем к ним иммунитета. При
- 41. Многие факультативные и облигатные внутриклеточные паразиты не только сохраняют жизнеспособность внутри клеток, но и способны размножаться.
- 42. Механизмы незавершенного фагоцитоза Блокада фагосомо–лизосомального слияния. Этот феномен обнаружен у вирусов (например, у вируса гриппа), бактерий
- 44. СИСТЕМА КОМПЛЕМЕНТА СИСТЕМА СЫВОРОТОЧНЫХ БЕЛКОВ (БОЛЕЕ 20 КОМПОНЕНТОВ), КОТОРЫЕ НАХОДЯТСЯ В КРОВИ В НЕАКТИВНОМ СОСТОЯНИИ И
- 46. КЛАССИЧЕСКИЙ ПУТЬ АКТИВАЦИИ КОМПЛЕМЕНТА АНТИТЕЛОЗАВИСИМЫЙ НАЧИНАЕТСЯ ПОСЛЕ ПРИСОЕДИНЕНИЯ С1 К КОМПЛЕКСУ АНТИГЕН-АНТИТЕЛО С5-КОНВЕРТАЗА СИНТЕЗИРУЕТСЯ КАК КОНЕЧНЫЙ
- 47. АЛЬТЕРНАТИВНЫЙ ПУТЬ АКТИВАЦИИ КОМПЛЕМЕНТА АКТИВАТОРЫ – КОМПОНЕНТЫ МИКРОБОВ, АГРЕГАТЫ РАЗЛИЧНЫХ БЕЛКОВ ПРОИСХОДИТ БЕЗ УЧАСТИЯ АНТИТЕЛ БЕЗ
- 48. ЛЕКТИНОВЫЙ ПУТЬ АКТИВАЦИИ КОМПЛЕМЕНТА ИНИЦИИРУЕТСЯ МАННОЗОСВЯЗЫВАЮЩИМ БЕЛКОМ – ЛЕКТИНОМ КРОВИ, СТРУКТУРНЫМ АНАЛОГОМ С1q МСБ СВЯЗЫВАЕТСЯ С
- 49. ЭФФЕКТОРНЫЕ ФУНКЦИИ КОМПЛЕМЕНТА
- 50. Антимикробные пептиды Лизоцим. Содержится в сыворотке, крови, слюне, слезах и других тканевых жидкостей человека. По биологической
- 51. Антимикробные пептиды Пропердин Пропердин или фактор Р-белок, содержащийся в сыворотке крови. Система пропердина состоит из самого
- 52. Антимикробные пептиды Цитокины Под термином “цитокины” объединяются так называемые ростовые факторы, которые регулируют пролиферацию, дифференцировку и
- 53. Белки острой фазы Пентраксины: - СРБ - Сывороточный амилоид Р Транспортные /связывающие белки - MBL (
- 56. Скачать презентацию

План лекции
1. КОМПОНЕНТЫ ВРОЖДЕННОГО ИММУНИТЕТА
2. ПОНЯТИЕ О РЕЦЕПТОРАХ РАСПОЗНАВАНИЯ ЧУЖОГО
3. РОЛЬ
План лекции
1. КОМПОНЕНТЫ ВРОЖДЕННОГО ИММУНИТЕТА
2. ПОНЯТИЕ О РЕЦЕПТОРАХ РАСПОЗНАВАНИЯ ЧУЖОГО
3. РОЛЬ

ФАКТОРЫ ВРОЖДЕННОГО ИММУНИТЕТА
КЛЕТОЧНЫЕ ФАКТОРЫ:
ФАГОЦИТОЗ
НАТУРАЛЬНЫЕ КИЛЛЕРЫ
ГУМОРАЛЬНЫЕ ФАКТОРЫ:
СИСТЕМА КОМПЛЕМЕНТА
СИСТЕМА
ФАКТОРЫ ВРОЖДЕННОГО ИММУНИТЕТА
КЛЕТОЧНЫЕ ФАКТОРЫ:
ФАГОЦИТОЗ
НАТУРАЛЬНЫЕ КИЛЛЕРЫ
ГУМОРАЛЬНЫЕ ФАКТОРЫ:
СИСТЕМА КОМПЛЕМЕНТА
СИСТЕМА
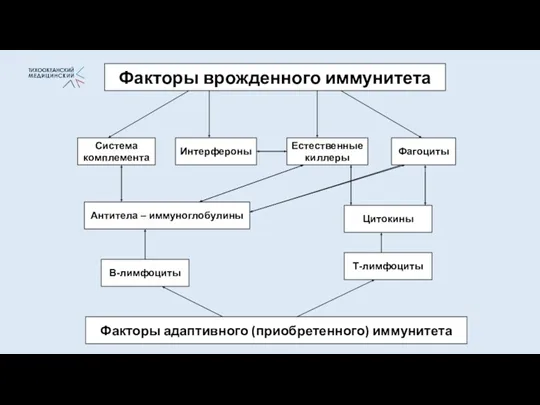
Факторы врожденного иммунитета
Фагоциты
Система
комплемента
Естественные
киллеры
Антитела – иммуноглобулины
Интерфероны
Факторы врожденного иммунитета
Фагоциты
Система
комплемента
Естественные
киллеры
Антитела – иммуноглобулины
Интерфероны

ОСНОВНОЕ СВОЙСТВО ВРОЖДЕННОГО ИММУНИТЕТА
НЕСПЕЦИФИЧНОСТЬ В РАСПОЗНАВАНИИ АНТИГЕНА
МОЛЕКУЛЫ И КЛЕТКИ ВРОЖДЕННОГО
ОСНОВНОЕ СВОЙСТВО ВРОЖДЕННОГО ИММУНИТЕТА
НЕСПЕЦИФИЧНОСТЬ В РАСПОЗНАВАНИИ АНТИГЕНА
МОЛЕКУЛЫ И КЛЕТКИ ВРОЖДЕННОГО

Система врожденного иммунитета должна иметь возможность отличать свое от чужого
Фагоциты могут
Система врожденного иммунитета должна иметь возможность отличать свое от чужого
Фагоциты могут

ПАТТЕРН-РАСПОЗНАЮЩИЕ РЕЦЕПТОРЫ
PRR -расположены на клеточной мембране фагоцитов и дендритных клеток
Примерами PAMP
ПАТТЕРН-РАСПОЗНАЮЩИЕ РЕЦЕПТОРЫ
PRR -расположены на клеточной мембране фагоцитов и дендритных клеток
Примерами PAMP

PRR можно разделить на 3 группы
рецепторы передачи сигналов
рецепторы эндоцитоза
PRR можно разделить на 3 группы
рецепторы передачи сигналов
рецепторы эндоцитоза

РЕЦЕПТОРЫ ПЕРЕДАЧИ СИГНАЛОВ
TOLL-ПОДОБНЫЕ РЕЦЕПТОРЫ
(toll-like receptor)
сигнализируют о появлении чужого
имеются на мембранах
РЕЦЕПТОРЫ ПЕРЕДАЧИ СИГНАЛОВ
TOLL-ПОДОБНЫЕ РЕЦЕПТОРЫ
(toll-like receptor)
сигнализируют о появлении чужого
имеются на мембранах

ЭНДОЦИТОЗНЫЕ, МЕМБРАННЫЕ PRR
Экспрессированы на поверхности фагоцитов
Отвечают за поглощение и транспорт
ЭНДОЦИТОЗНЫЕ, МЕМБРАННЫЕ PRR
Экспрессированы на поверхности фагоцитов
Отвечают за поглощение и транспорт

СЕКРЕТИРУЕМЫЕ, РАСТВОРИМЫЕ, PRR
Это опсонины, которые «обволакивают» поверхность антигена, облегчая фагоцитоз:
Компонент С1
СЕКРЕТИРУЕМЫЕ, РАСТВОРИМЫЕ, PRR
Это опсонины, которые «обволакивают» поверхность антигена, облегчая фагоцитоз:
Компонент С1

У человека установлены типы TLR 1 – 11 типов
Все известные
У человека установлены типы TLR 1 – 11 типов
Все известные
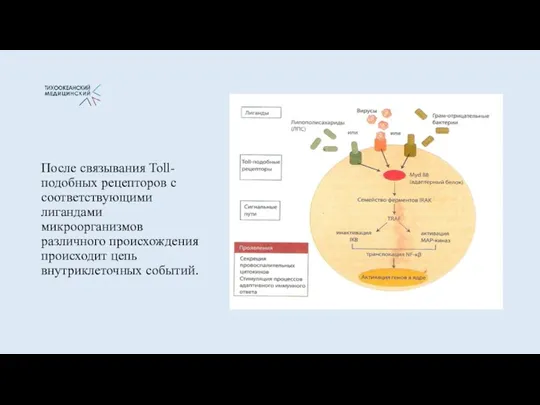
После связывания Toll- подобных рецепторов с соответствующими лигандами микроорганизмов различного происхождения
После связывания Toll- подобных рецепторов с соответствующими лигандами микроорганизмов различного происхождения

Эти структуры взаимодействуют со специализированными паттерн-распознающими (PRR) рецепторами на поверхности клеток
Эти структуры взаимодействуют со специализированными паттерн-распознающими (PRR) рецепторами на поверхности клеток

ФАГОЦИТОЗ
ЭТО ПОГЛОЩЕНИЕ ГРАНУЛОЦИТАМИ (НЕЙТРОФИЛАМИ, ЭОЗИНОФИЛАМИ, БАЗОФИЛАМИ) И АГРАНУЛОЦИТАМИ (МОНОЦИТАМИ/МАКРОФАГАМИ) ЧАСТИЦ ДИАМЕТРОМ
ФАГОЦИТОЗ
ЭТО ПОГЛОЩЕНИЕ ГРАНУЛОЦИТАМИ (НЕЙТРОФИЛАМИ, ЭОЗИНОФИЛАМИ, БАЗОФИЛАМИ) И АГРАНУЛОЦИТАМИ (МОНОЦИТАМИ/МАКРОФАГАМИ) ЧАСТИЦ ДИАМЕТРОМ
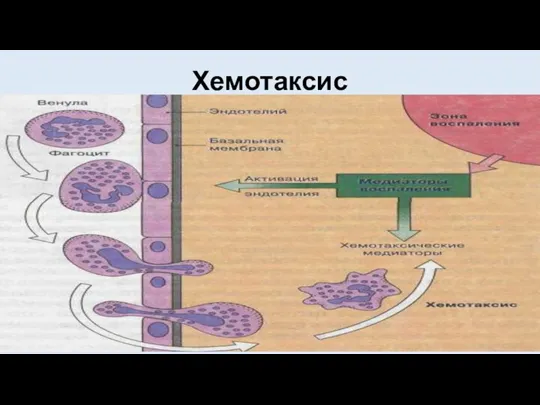
Хемотаксис
Хемотаксис
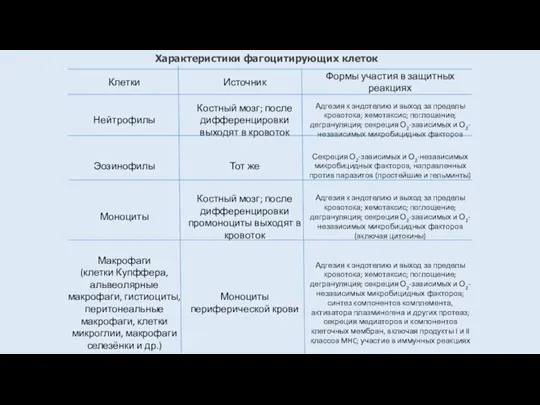
Характеристики фагоцитирующих клеток
Характеристики фагоцитирующих клеток

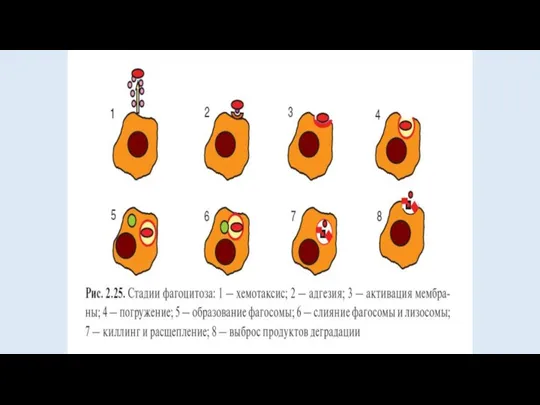

Условно весь процесс принято делить на несколько этапов.
Первым из них
Условно весь процесс принято делить на несколько этапов.
Первым из них

Наиболее типичными хемоаттрактантами собственного происхождения являются:
медиаторы воспаления (лейкотриен В4, гистамин и
Наиболее типичными хемоаттрактантами собственного происхождения являются:
медиаторы воспаления (лейкотриен В4, гистамин и

Для этих веществ на поверхности фагоцитирующих клеток имеются специфические рецепторы, присоединение
Для этих веществ на поверхности фагоцитирующих клеток имеются специфические рецепторы, присоединение

Клетка из округлой становится треугольной, обращенной в сторону движения. В части
Клетка из округлой становится треугольной, обращенной в сторону движения. В части

Связывание фагоцита и чужеродного объекта.
Стадия прилипания включает опсонизацию, распознавание и прикрепление
Связывание фагоцита и чужеродного объекта.
Стадия прилипания включает опсонизацию, распознавание и прикрепление
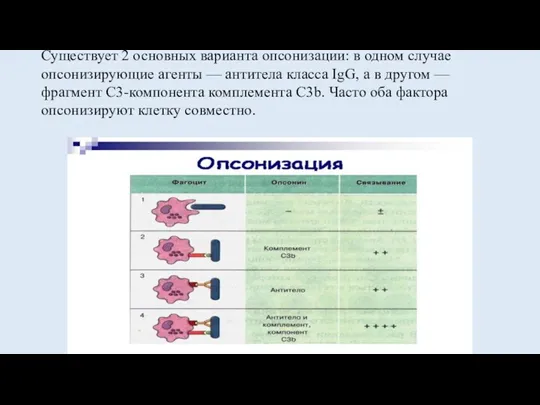
Существует 2 основных варианта опсонизации: в одном случае опсонизирующие агенты —
Существует 2 основных варианта опсонизации: в одном случае опсонизирующие агенты —

РЕЦЕПТОРЫ ДЛЯ РАСПОЗНАВАНИЯ ОПСОНИНОВ (FC- И C3-РЕЦЕПТОРЫ)
Распознавание патогенов и других клеток,
РЕЦЕПТОРЫ ДЛЯ РАСПОЗНАВАНИЯ ОПСОНИНОВ (FC- И C3-РЕЦЕПТОРЫ)
Распознавание патогенов и других клеток,

С3-рецепторы играют важную
роль в контроле активации комплемента, ингибируя связывание С3 с
С3-рецепторы играют важную
роль в контроле активации комплемента, ингибируя связывание С3 с

СТАДИЯ АКТИВАЦИИ МЕМБРАНЫ
На начальных стадиях фагоцитоза основные события происходят на обращенном
СТАДИЯ АКТИВАЦИИ МЕМБРАНЫ
На начальных стадиях фагоцитоза основные события происходят на обращенном

Следующий этап активации направлен на образование продуктов, участвующих в полимеризации актина
Следующий этап активации направлен на образование продуктов, участвующих в полимеризации актина

СТАДИИ ПОГРУЖЕНИЯ И ОБРАЗОВАНИЯ ФАГОСОМЫ.
Погружение частицы обусловлено сокращением нитей актина, сконцентрированных
СТАДИИ ПОГРУЖЕНИЯ И ОБРАЗОВАНИЯ ФАГОСОМЫ.
Погружение частицы обусловлено сокращением нитей актина, сконцентрированных

Сразу после образования фагосома не несет бактерицидных веществ и ферментов, способных
Сразу после образования фагосома не несет бактерицидных веществ и ферментов, способных

СЛИЯНИЕ ФАГОСОМЫ И ЛИЗОСОМЫ
Решающий вклад в созревание фагосом и обретение ими
СЛИЯНИЕ ФАГОСОМЫ И ЛИЗОСОМЫ
Решающий вклад в созревание фагосом и обретение ими

Стадия киллинга
обеспечивается наличием в фагоцитах бактерицидных факторов, которые выделяются
Стадия киллинга
обеспечивается наличием в фагоцитах бактерицидных факторов, которые выделяются

Кислородзависимые механизмы связаны с образованием активных форм кислорода, обеспечивающих микробицидное действие
К
Кислородзависимые механизмы связаны с образованием активных форм кислорода, обеспечивающих микробицидное действие
К

Образующиеся активные радикалы обусловливают повреждение и деструкцию белков и липидов мембран,
Образующиеся активные радикалы обусловливают повреждение и деструкцию белков и липидов мембран,

ВЫБРОС ФАГОЦИТАМИ ПРОДУКТОВ ДЕГРАДАЦИИ (ДЕГРАНУЛЯЦИЯ)
Заключительная фаза фагоцитоза — выброс содержимого фаголизосом
ВЫБРОС ФАГОЦИТАМИ ПРОДУКТОВ ДЕГРАДАЦИИ (ДЕГРАНУЛЯЦИЯ)
Заключительная фаза фагоцитоза — выброс содержимого фаголизосом

При незавершенном фагоцитозе чужеродные клетки, поглощенные фагоцитами, остаются жизнеспособными. Некоторые микроорганизмы,
При незавершенном фагоцитозе чужеродные клетки, поглощенные фагоцитами, остаются жизнеспособными. Некоторые микроорганизмы,
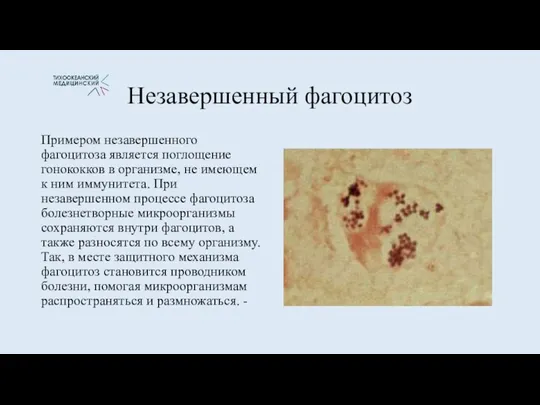
Незавершенный фагоцитоз
Примером незавершенного фагоцитоза является поглощение гонококков в организме, не имеющем
Незавершенный фагоцитоз
Примером незавершенного фагоцитоза является поглощение гонококков в организме, не имеющем

Многие факультативные и облигатные внутриклеточные паразиты не только сохраняют жизнеспособность внутри
Многие факультативные и облигатные внутриклеточные паразиты не только сохраняют жизнеспособность внутри

Механизмы незавершенного фагоцитоза
Блокада фагосомо–лизосомального слияния. Этот феномен обнаружен у вирусов
Механизмы незавершенного фагоцитоза
Блокада фагосомо–лизосомального слияния. Этот феномен обнаружен у вирусов
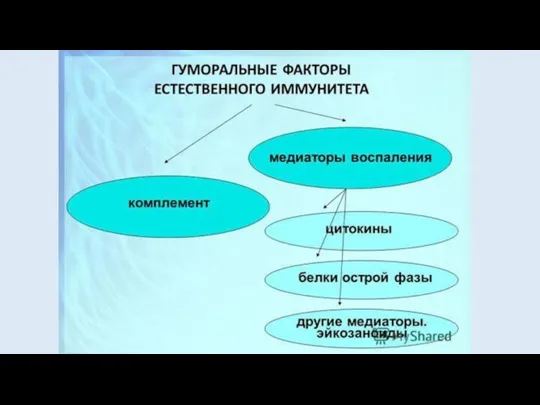

СИСТЕМА КОМПЛЕМЕНТА
СИСТЕМА СЫВОРОТОЧНЫХ БЕЛКОВ (БОЛЕЕ 20 КОМПОНЕНТОВ), КОТОРЫЕ НАХОДЯТСЯ В КРОВИ
СИСТЕМА КОМПЛЕМЕНТА
СИСТЕМА СЫВОРОТОЧНЫХ БЕЛКОВ (БОЛЕЕ 20 КОМПОНЕНТОВ), КОТОРЫЕ НАХОДЯТСЯ В КРОВИ
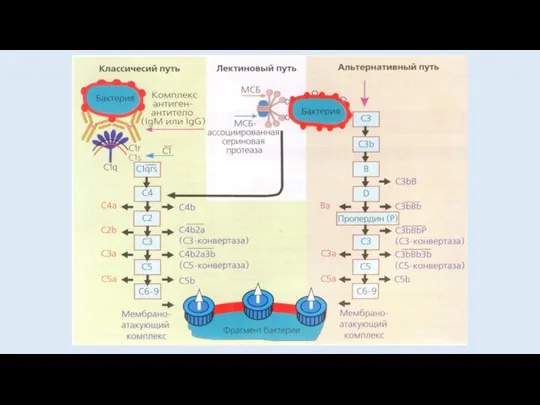

КЛАССИЧЕСКИЙ ПУТЬ АКТИВАЦИИ КОМПЛЕМЕНТА
АНТИТЕЛОЗАВИСИМЫЙ
НАЧИНАЕТСЯ ПОСЛЕ ПРИСОЕДИНЕНИЯ С1 К КОМПЛЕКСУ АНТИГЕН-АНТИТЕЛО
С5-КОНВЕРТАЗА СИНТЕЗИРУЕТСЯ
КЛАССИЧЕСКИЙ ПУТЬ АКТИВАЦИИ КОМПЛЕМЕНТА
АНТИТЕЛОЗАВИСИМЫЙ
НАЧИНАЕТСЯ ПОСЛЕ ПРИСОЕДИНЕНИЯ С1 К КОМПЛЕКСУ АНТИГЕН-АНТИТЕЛО
С5-КОНВЕРТАЗА СИНТЕЗИРУЕТСЯ

АЛЬТЕРНАТИВНЫЙ ПУТЬ АКТИВАЦИИ КОМПЛЕМЕНТА
АКТИВАТОРЫ – КОМПОНЕНТЫ МИКРОБОВ, АГРЕГАТЫ РАЗЛИЧНЫХ
АЛЬТЕРНАТИВНЫЙ ПУТЬ АКТИВАЦИИ КОМПЛЕМЕНТА
АКТИВАТОРЫ – КОМПОНЕНТЫ МИКРОБОВ, АГРЕГАТЫ РАЗЛИЧНЫХ

ЛЕКТИНОВЫЙ ПУТЬ АКТИВАЦИИ КОМПЛЕМЕНТА
ИНИЦИИРУЕТСЯ МАННОЗОСВЯЗЫВАЮЩИМ БЕЛКОМ – ЛЕКТИНОМ КРОВИ, СТРУКТУРНЫМ
ЛЕКТИНОВЫЙ ПУТЬ АКТИВАЦИИ КОМПЛЕМЕНТА
ИНИЦИИРУЕТСЯ МАННОЗОСВЯЗЫВАЮЩИМ БЕЛКОМ – ЛЕКТИНОМ КРОВИ, СТРУКТУРНЫМ
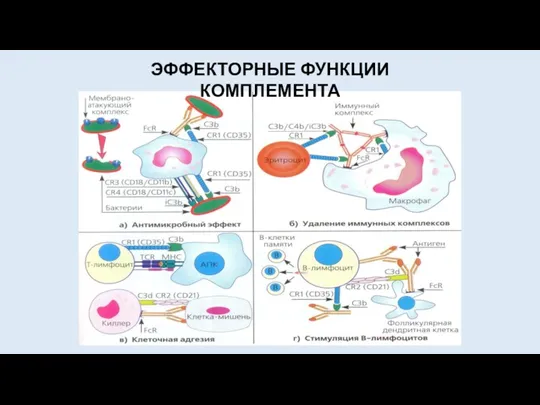
ЭФФЕКТОРНЫЕ ФУНКЦИИ КОМПЛЕМЕНТА
ЭФФЕКТОРНЫЕ ФУНКЦИИ КОМПЛЕМЕНТА

Антимикробные пептиды
Лизоцим.
Содержится в сыворотке, крови, слюне, слезах и других тканевых
Антимикробные пептиды
Лизоцим.
Содержится в сыворотке, крови, слюне, слезах и других тканевых

Антимикробные пептиды
Пропердин
Пропердин или фактор Р-белок, содержащийся в сыворотке крови. Система
Антимикробные пептиды
Пропердин
Пропердин или фактор Р-белок, содержащийся в сыворотке крови. Система

Антимикробные пептиды
Цитокины
Под термином “цитокины” объединяются так называемые ростовые факторы, которые
Антимикробные пептиды
Цитокины
Под термином “цитокины” объединяются так называемые ростовые факторы, которые

Белки острой фазы
Пентраксины:
- СРБ
- Сывороточный амилоид Р
Транспортные /связывающие белки
-
Белки острой фазы
Пентраксины:
- СРБ
- Сывороточный амилоид Р
Транспортные /связывающие белки
-

 Эпидемии и пандемии в Средневековой Европе
Эпидемии и пандемии в Средневековой Европе Мероприятия по снижению риска заражения и распространения новой коронавирусной инфекции 2019-nCoV
Мероприятия по снижению риска заражения и распространения новой коронавирусной инфекции 2019-nCoV Миогимнастика как метод профилактики и лечения деформаций зубочелюстной системы ребенка
Миогимнастика как метод профилактики и лечения деформаций зубочелюстной системы ребенка Клинический случай ХНН
Клинический случай ХНН Средства, действующие на ЦНС
Средства, действующие на ЦНС Виды повязок. Правила наложения бинтовых повязок
Виды повязок. Правила наложения бинтовых повязок Дефект межпредсердной перегородки (ДМПП)
Дефект межпредсердной перегородки (ДМПП) Травма головы, семинар. Оказание помощи пострадавшим с ЧМТ на догоспитальном этапе
Травма головы, семинар. Оказание помощи пострадавшим с ЧМТ на догоспитальном этапе Острые вялые параличи
Острые вялые параличи Теоретичні основи фізичної терапії та ерготерапії
Теоретичні основи фізичної терапії та ерготерапії Первая помощь при вывихах и переломах костей
Первая помощь при вывихах и переломах костей Фармакоекономічний аналіз використання лікарських засобів на прикладі Лікарняна каса Житомирської області
Фармакоекономічний аналіз використання лікарських засобів на прикладі Лікарняна каса Житомирської області Гормональная контрацепция. Взвешенный выбор
Гормональная контрацепция. Взвешенный выбор Эхокардиография, ультразвуковая кардиография
Эхокардиография, ультразвуковая кардиография Питание ребёнка первого года жизни
Питание ребёнка первого года жизни Эпилепсия. Классификация эпилепсии. Диагностические критерии. Неотложная помощь. Принципы лечения
Эпилепсия. Классификация эпилепсии. Диагностические критерии. Неотложная помощь. Принципы лечения Медичне страхування
Медичне страхування Неврологические осложнения остеохондроза позвоночника
Неврологические осложнения остеохондроза позвоночника Денсаулықпен байланысты өмір сүрудің сапасы
Денсаулықпен байланысты өмір сүрудің сапасы Задачи, организационная структура и органы управления Всероссийской службы медицины катастроф
Задачи, организационная структура и органы управления Всероссийской службы медицины катастроф Профилактика и лечение болезни Альцгеймера
Профилактика и лечение болезни Альцгеймера Дизентерия
Дизентерия Осторожно! Клещи
Осторожно! Клещи Современная система контроля и обеспечения качества лекарственных средств в Украине
Современная система контроля и обеспечения качества лекарственных средств в Украине Негізгі психопатологиялық синдромдар
Негізгі психопатологиялық синдромдар Болезни органов брюшной полости. Болезни органов ротовой полости
Болезни органов брюшной полости. Болезни органов ротовой полости Эндокринные заболевания и беременность
Эндокринные заболевания и беременность Рахит. Этиопатогенез рахита
Рахит. Этиопатогенез рахита