- Метаболизм липидов

Содержание
- 2. Липиды Органические вещества, нерастворимые в воде, но растворимые в органических растворителях (эфир, хлороформ, бензен).
- 3. Функции липидов Энергетическая Резервная Структурная Теплоизоляция Механическая защита Регуляторная Витаминная
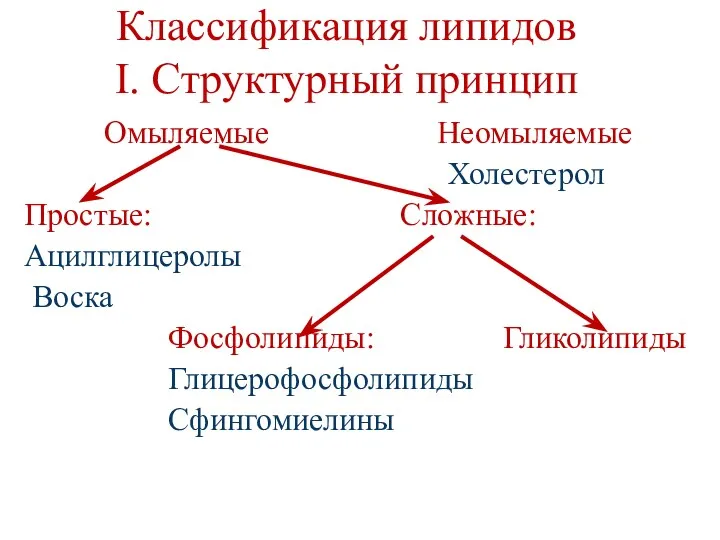
- 4. Классификация липидов I. Cтруктурный принцип Омыляемые Неомыляемые Холестерол Простые: Сложные: Ацилглицеролы Воска Фосфолипиды: Гликолипиды Глицерофосфолипиды Сфингомиелины
- 5. Классификация липидов II. Физико-химический принцип: 1. Нейтральные 2. Полярные (амфифильные) III. Функциональный принцип: Резервные Структурные
- 6. Жирные кислоты Насыщенные Ненасыщенные: Моноеновые Полиеновые
- 7. Ацилглицеролы Глицерол Жирные кислоты
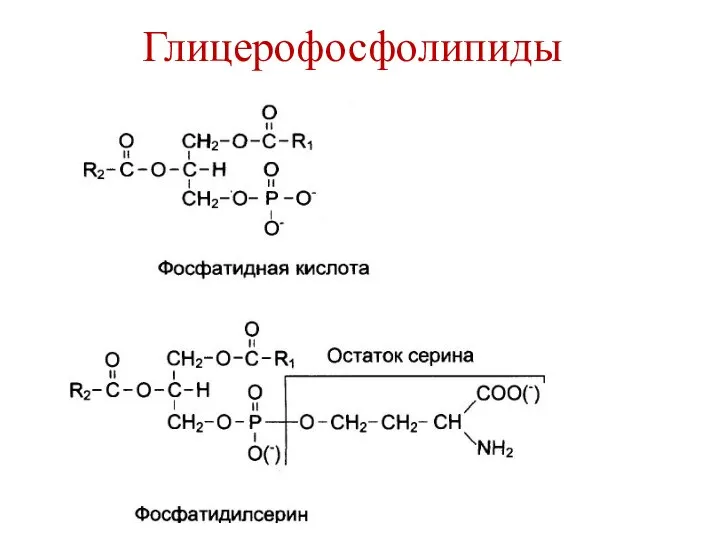
- 8. Глицерофосфолипиды Глицерол Жирные кислоты Фосфорная кислота Аминоспирт
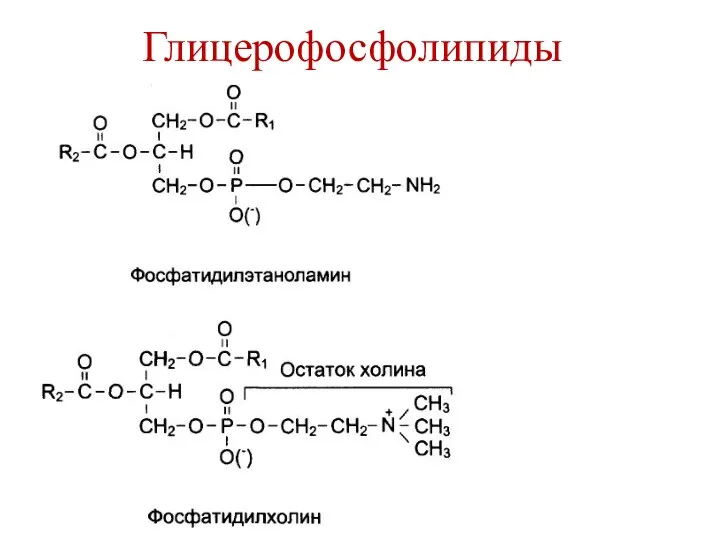
- 9. Глицерофосфолипиды
- 10. Глицерофосфолипиды
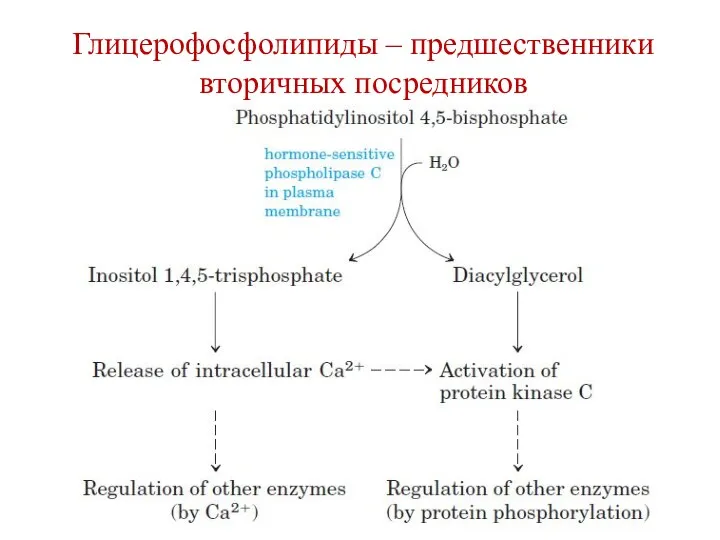
- 11. Глицерофосфолипиды – предшественники вторичных посредников
- 12. Плазмалогены
- 13. Сфингомиелины Сфингозин Жирная кислота Фосфорная кислота Аминоспирт
- 14. Сфингомиелины
- 15. Сфингомиелины
- 16. Гликолипиды Сфингозин Жирная кислота Углеводы Классификация Цереброзиды Сульфатиды Глобозиды Ганглиозиды
- 17. Цереброзиды Сульфатиды
- 18. Ганглиозид GM2
- 19. Группы крови
- 20. Холестерол Эфир холестерола
- 21. Желчные кислоты
- 22. Переваривание липидов. Плазменные липопротеины. Катаболизм липидов
- 23. Основными пищевыми липидами являются Триацилглицеролы Холестерол и холестериды Фосфолипиды Значение пищевых липидов: Суточная норма липидов –
- 24. Переваривание липидов в ЖКТ У детей начинается в ротовой полости под действием лингвальной липазы; продолжается в
- 25. Переваривание липидов в кишечнике (необходимые условия) Щелочная среда – рН = 8 (бикарбонаты панкреатического сока) Липолитические
- 26. Желчные кислоты Синтезируются в печени из холестерола Выделяются с желчью в двенадцатиперстную кишку Функции желчных кислот
- 27. Холановая кислота Желчные кислоты являются производными холановой кислоты Холевая кислота (3,7,12-тригидроксихолановая кислота) НО НО ОН 3
- 28. Таурин Глицин
- 29. Гидролиз триацилгдицеролов Поджелудочная липаза 2Н2О R1COOH R2COOH ТАГ МАГ
- 30. Гидролиз фосфолипидов Поджелудочные фосфолипазы А1, А2, С и Д 4Н2О глицерол 2 ЖК фосфорная кислота азотистое
- 31. Гидролиз фосфолипидов Основным ферментом является фосфолипаза А2, под действием которой из фосфолипидов образуется лизолецитин и ЖК
- 32. Гидролиз холестеридов Поджелудочная холестеролэстераза Холестерол + ЖК Н2О
- 33. Всасывание продуктов гидролиза липидов Глицерол, коротко- и среднецепочечные ЖК – свободная диффузия. МАГ, длинноцепочечные ЖК, холестерол
- 34. Всасывание продуктов гидролиза липидов В энтероцитах мицеллы распадаются. Продукты гидролиза липидов подвергаются ресинтезу. Желчные кислоты всасываются
- 35. Регуляция переваривания липидов Секретин стимулирует выделение печенью и поджелудочной железой водянистого раствора с высоким содержанием бикарбонатов
- 36. Нарушения переваривания и всасывания липидов Стеаторея – появление в фекалиях неперевареных липидов Печеночная (желчнокаменная болезнь, дискинезия
- 37. Ресинтез липидов в энтероцитах Моноацилглицерол + 2 R-CO-S-CoA триацилглицерол + 2 НS-CoA Лизолецитин + R-CO-S-CoA лецитин
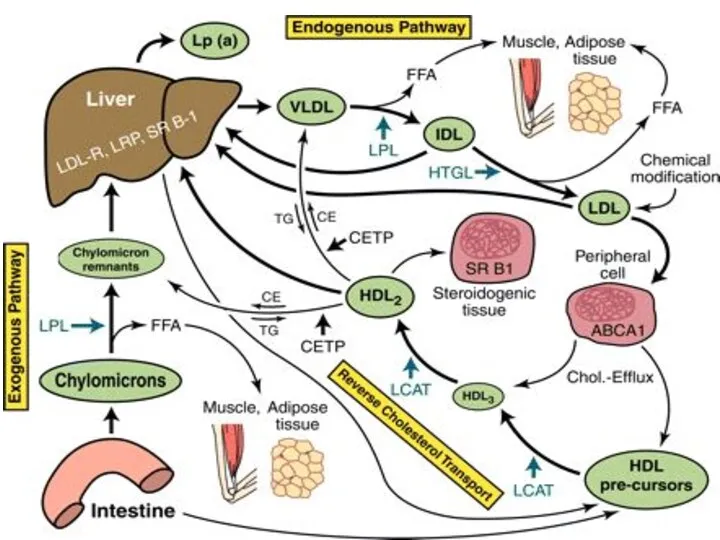
- 38. Плазменные липопротеины липидно-белковые комплексы, транспортные формы липидов. Все липопротеины имеют одинаковую структуру: состоят из гидрофобного ядра
- 39. Гидрофильная оболчка Гидрофобная поверхность Структура липопротеинов
- 40. Липопротеины различаются по содержанию липидов и апопротеинов, что определяет их различную плотность и электрический заряд. Методы
- 41. Разделение липопротеинов ультрацентрифугированием HDL VLDL LDL Хиломикроны (CM); Липопротеины очень низкой плотности - ЛПОНП (VLDL –
- 42. Разделение липопротеинов электрофорезом (PH = 8,6, полиакриламидный гель) Хиломикроны не мигрируют в электрическом поле; пре-β-ЛП (соответствуют
- 43. Обмен хиломикронов образуются в эпителии кишечника; содержат много ТАГ (85%), содержат апопротеин В-48 в качестве главного
- 44. Обмен хиломикронов после синтеза секретируются в лимфу и через грудной проток попадают в кровь; в крови
- 45. Обмен хиломикронов ЛПЛ гидролизует ТАГ из хиломикронов. ЖК поступают в скелетные мышцы, миокард (используются для энергии),
- 47. Обмен ЛПОНП Образуются в печени; Содержат много ТАГ (65%) и апоВ-100 в качестве главного структурного белка.
- 48. Обмен ЛПОНП Катаболизм ЛПНП происходит под действием фермента ЛПЛ (аналогично хиломикронам). после гидролиза ТАГ, ЛПОНП преобразуется
- 50. Обмен ЛПНП Образуются в кровотоке из ЛПОНП. Являются основными переносчиками эндогенного ХС к внепеченочным тканям (транспортирует
- 51. Обмен ЛПНП Регулируемый путь катаболизма – связывание с апо В/Е-рецепторами печени, клеток надпочечников и периферических клеток.
- 53. Обмен ЛПНП Альтернативный путь (нерегулируемый) – характерен для окисленных ЛПНП. Перекисно-модифицированные ЛПНП распознаются и захватываются скэвенджер-рецепторами
- 54. Обмен ЛПВП ЛПВП образуются в печени в виде незрелых дисковидных частиц, состоящих из фосфолипидов и апопротеинов
- 55. Обмен ЛПВП ЛПВП – хорошие акцепторы свободного ХС из периферических тканей. Свободный ХС из клеточных мембран
- 56. Обмен ЛПВП Эфиры ХС перемещаются с поверхности частиц ЛПВП в гидрофобное ядро, освобождая таким образом дополнительную
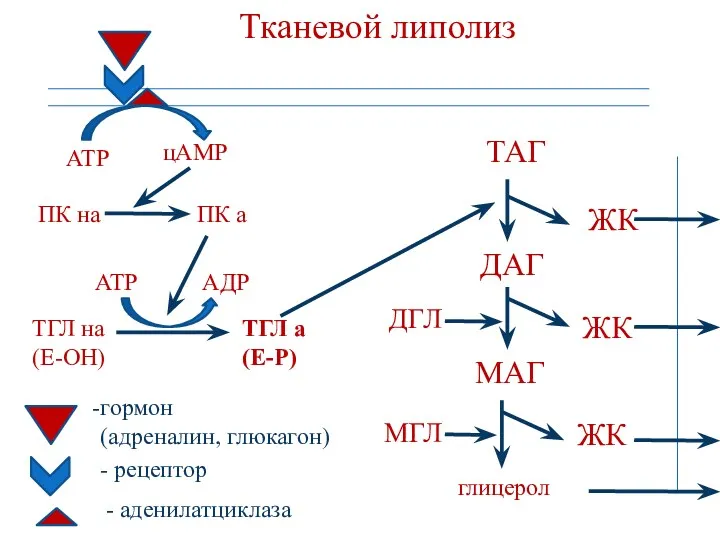
- 59. Мобилизация жиров из жировой ткани (тканевой липолиз) Резервные липиды обеспечивают энергией организм в течение 7-8 недель.
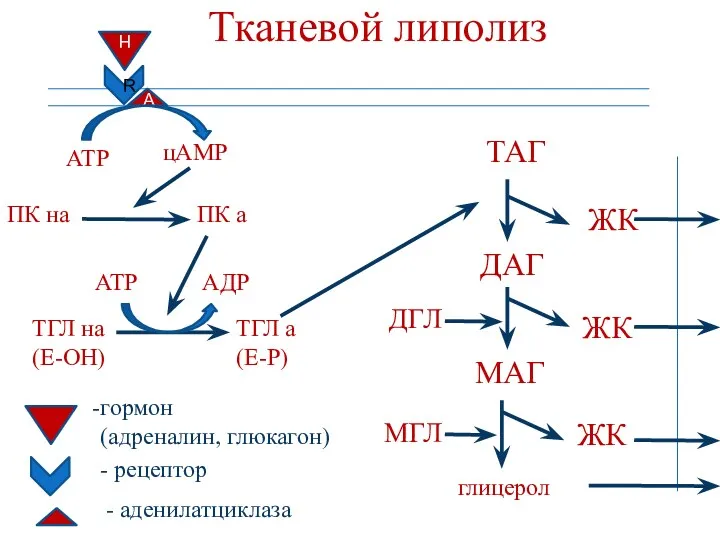
- 60. Тканевой липолиз R H A АТР цАМР ПК на ПК а ТГЛ на (Е-ОН) ТГЛ а
- 61. Регуляция липолиза Адреналин, глюкагон стимулируют липолиз (активируют триглицеридлипазу – фосфорилирование фермента); Глюкокортикоиды (кортизол) стимулируют липолиз (индуцируют
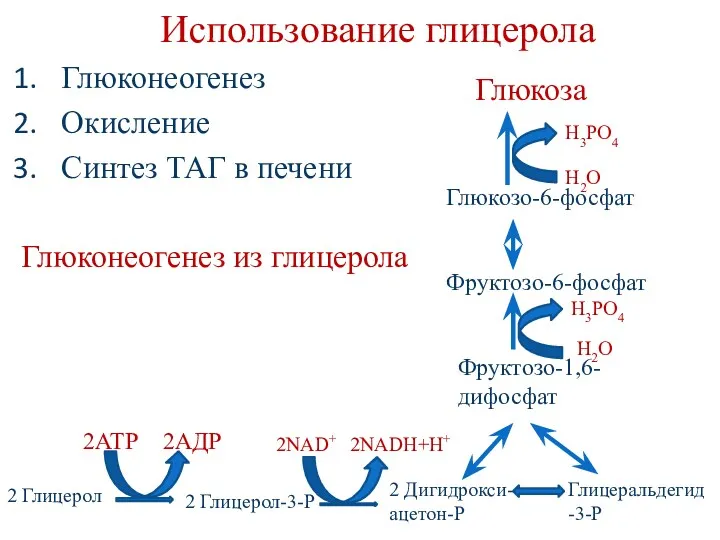
- 62. Использование глицерола Глюконеогенез Окисление Синтез ТАГ в печени Глюконеогенез из глицерола 2 Глицерол 2 Глицерол-3-Р Глицеральдегид
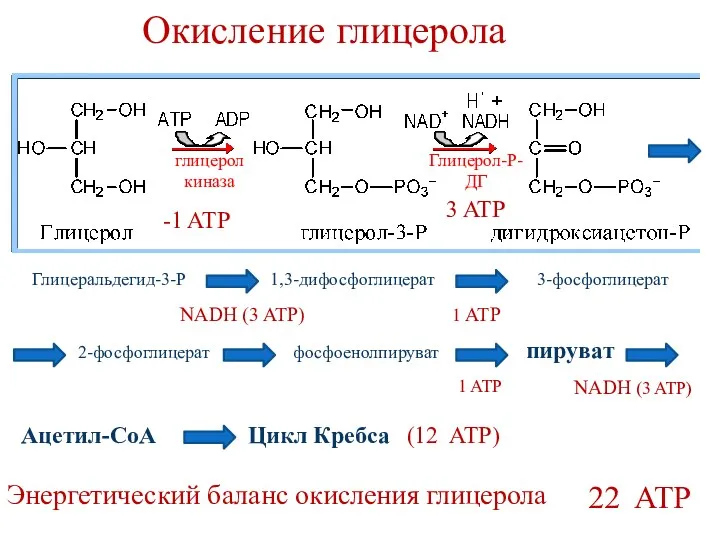
- 63. Окисление глицерола Глицеральдегид-3-Р 1,3-дифосфоглицерат 3-фосфоглицерат 2-фосфоглицерат фосфоенолпируват пируват Ацетил-СоА Цикл Кребса NADH (3 ATP) NADH (3
- 64. Бета-окисление жирных кислот Локализация – скелетные мышцы, миокард, печень, почки Не происходит в нервной ткани, эритроцитах
- 65. Активация жирных кислот R–COOH + HS-CoA ATP AMP + H4P2O7 H3PO4 H3PO4 Жирная кислота Ацил-СоА Ацил-СоА-
- 66. Перенос ацил-СоА в митохондрии
- 67. Первая реакция бета-окисления
- 68. Вторая реакция бета-окисления
- 69. Третья реакция бета-окисления
- 70. Четвертая реакция бета-окисления
- 72. Сn Сn-4 С4 С2 Сn-2 С2 С2 С2 С2 С2 С2 Цикл Кребса СО2 СО2 2Н
- 73. Энергетический баланс бета-окисления N – количество атомов углерода ЖК N/2-1 – количество циклов бета-окисления N/2 –
- 74. Особенности окисления ненасыщенных жирных кислот При окислении ненасыщенных ЖК, после 3-х циклов бета-окисления, образуется Δ3,4-цис-еноил-СоА, а
- 75. Особенности окисления ненасыщенных жирных кислот Δ3,4-цис –> Δ2,3-транс-еноил-КоА-изомераза
- 76. Особенности окисления жирных кислот с нечетным числом атомов углерода В последнем цикле бета-окисления образуется пропионил-СоА, который
- 78. Особенности окисления жирных кислот в пероксисомах Характерно для ЖК с 20-26 атомами углерода. Особенности – первая
- 79. Биосинтез липидов
- 80. Биосинтез жирных кислот Локализация процесса – печень, жировая ткань, лактирующая молочная железа. Происходит в цитоплазме. Субстрат
- 81. Транспорт ацетил-СоА из митохондрий в цитоплазму
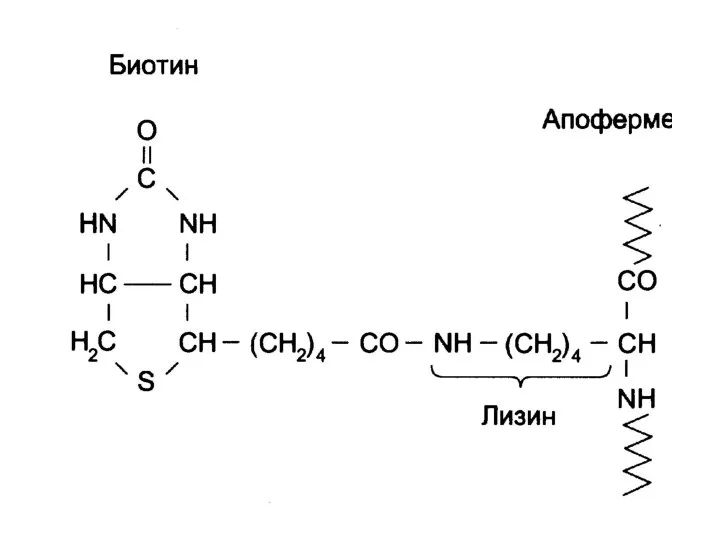
- 82. Синтез малонил-СоА CH3-CO-SCoA + HOOC-CH2-CO-SCoA CO2 ATP ADP+ H3PO4 Ацетил-CoA- карбоксилаза Простетическая группа фермента – биотин
- 84. Собственно-биосинтез жирных кислот Происходит под действием мультиферментного комплекса, синтаза жирных кислот (пальмитат синтаза), состоящего из 7
- 85. Синтаза жирных кислот SН Е-АПБ SH SН Е-АСР SH
- 86. Биосинтез жирных кислот ацетил-СоА-АПБ-трансфераза (ацетилтрансацилаза) Малонил-СоА-АПБ-трансфераза (малонилтрансацилаза) β-кетоацил-АПБ-синтаза
- 87. Биосинтез жирных кислот 4.β-кетоацил-АПБ-редуктаза 5.β-гидроксиацил-АПБ-дегидратаза 6.еноил-СоА-АПБ-редуктаза
- 88. Биосинтез жирных кислот Реакции повторяется 7 раз до синтеза пальмитоил-АПБ. Завершается синтез жирной кислоты отщеплением пальмитиновой
- 89. Суммарная реакция биосинтеза пальмитиновой кислоты СН3–СО–SСoA + 7НООС–СН2–СО–SСoA + 14НАДФН +14Н+ → СН3–(СН2)14–СООН + 7СO2 +
- 90. Регуляция биосинтеза жирных кислот Основной регуляторный фермент – ацетил-СоА карбоксилаза Цитрат – активатор, пальмитоил-СоА – ингибитор
- 91. Элонгация (удлинение) жирных кислот Происходит в ЭР под действием энзиматической системы элонгаза. Удлинение цепи жирной кислоты
- 92. Элонгация жирных кислот
- 93. Синтез ненасыщенных жирных кислот Моноеновые жирные кислоты – пальмитоолеиновая и олеиновая – синтезируются из пальмитиновой и
- 94. Синтез ненасыщенных жирных кислот
- 95. Десатураза млекопитающих может образовать двойные связи только на участке цепи жирной кислоты от 9-го до 1-го
- 96. Синтез арахидоновой кислоты
- 97. Синтез триацилглицеролов Локализация процесса – печень, жировая ткань. Синтез происходит из глицерол-3-фосфата и ацил-СоА (главным образом
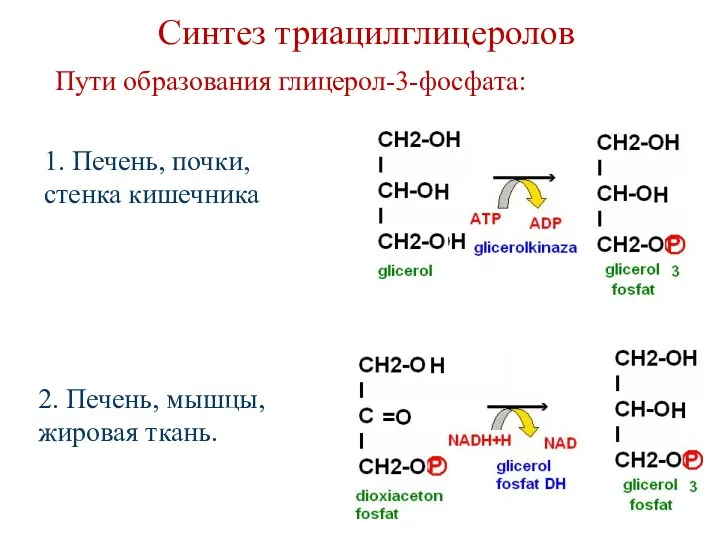
- 98. Синтез триацилглицеролов 1. Печень, почки, стенка кишечника 2. Печень, мышцы, жировая ткань. Пути образования глицерол-3-фосфата:
- 99. Синтез триацилглицеролов
- 100. Синтез фосфолипидов интенсивно происходит в печени, стенке кишечника, семенниках, яичниках, молочной железе. Протекает в эндоплазматической сети.
- 101. I. Синтез фосфолипидов
- 102. I. Синтез фосфолипидов +
- 103. II. Синтез фосфолипидов + + + +
- 104. Синтез сфингомиелинов Пальмитоил-СоА + серин →→→сфингозин Сфингозин + ацил-СоА →церамид Церамид + СДФ-холин → сфингомиелин
- 105. Синтез сфинголипидов
- 106. Синтез гликолипидов Церамид + УДФ-глюкоза → глюкоцереброзид + УДФ Церамид + УДФ-галактоза → галактоцереброзид + УДФ
- 107. Синтез холестерола За сутки синтезируется 1г холестерола. С пищей поступает 0,3-0,5г. Локализация – печень, эпителий кишечника,
- 108. Синтез холестерола
- 109. Синтез холестерола
- 110. Регуляция биосинтеза холестерола Регуляторный фермент – ГМГ-КоА-редуктаза Ингибиторы – холестерол, мевалоновая кислота, желчные кислоты. Гормональная регуляция:
- 111. ≪Кетоновые тела≫: ацетон ацетоуксусная кислота (ацетоацетат) β-оксимасляная кислота (гидроксибутират)
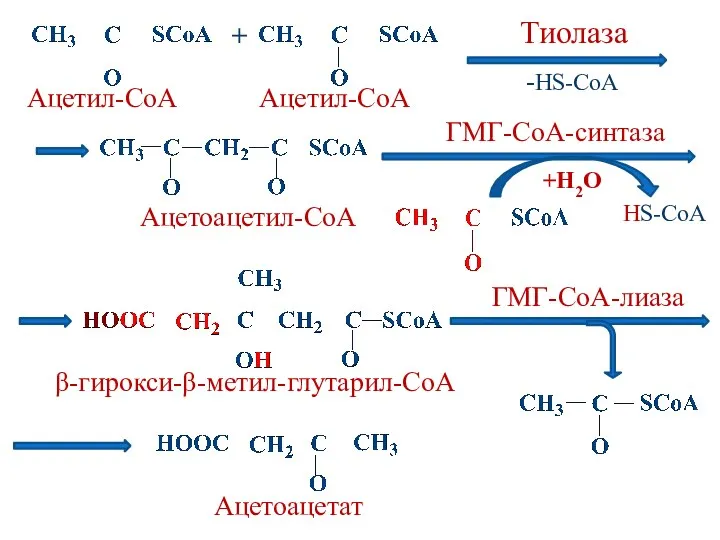
- 112. Синтез ≪кетоновых тел≫ Происходит только в печени. Субстратом является ацетил-СоА.
- 113. + Ацетил-СоА Ацетил-СоА Тиолаза Ацетоацетил-СоА -HS-CoA HS-CoA +H2O β-гирокси-β-метил-глутарил-СоА Ацетоацетат ГМГ-СоА-синтаза ГМГ-СоА-лиаза
- 114. Ацетоацетат Ацетон -CO2 NADH+H+ β-гидроксибутират NAD+
- 115. Использование «кетоновых тел» в тканях используются в качестве ≪топлива≫ сердечной мышцей, корковым слоем почек (в нормальных
- 116. Использование «кетоновых тел» в тканях СН3-СН(ОН)-СН2-СООН СН3-СО-СН2-СООН СН3-СО-СН2-СО -SCoA 2 СН3-СО-SCoA 2 циклa Кребса (СО2, Н2О)
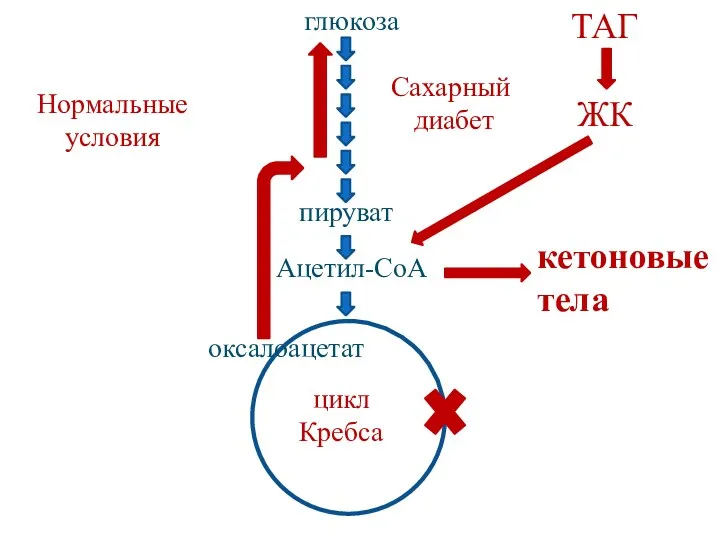
- 117. Нормальная концентрация «кетоновых тел» (0,03–0,2 ммоль/л). При патологических состояниях (сахарный диабет, голодание) концентрация кетоновых тел в
- 118. Причины кетонемии Увеличение концентрации ацетил-СоА (усиленное расщепление липидов). Уменьшение концентрации оксалоацетата (при отсутствии углеводов или нарушении
- 119. глюкоза пируват Ацетил-СоА оксалоацетат ТАГ ЖК кетоновые тела цикл Кребса Нормальные условия Сахарный диабет
- 120. Эйкозаноиды. Жирорастворимые витамины. Регуляция и патология липидного обмена
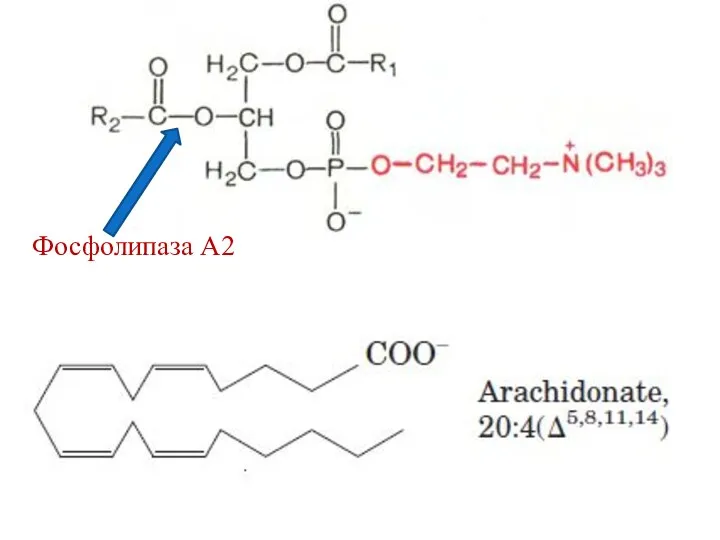
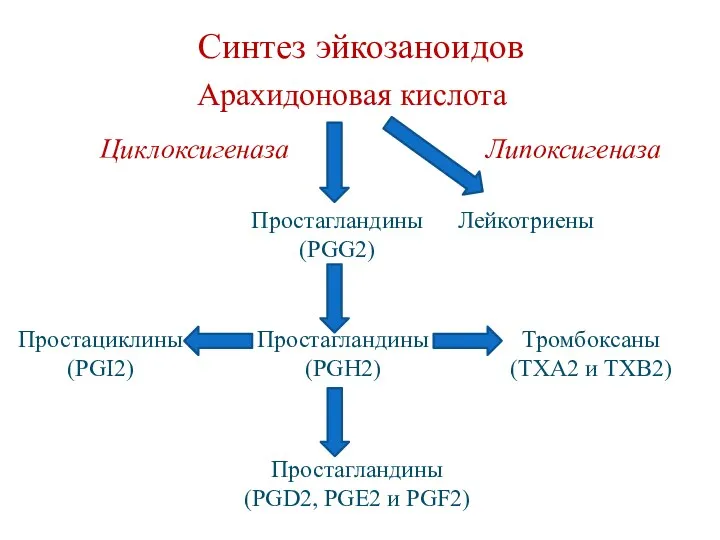
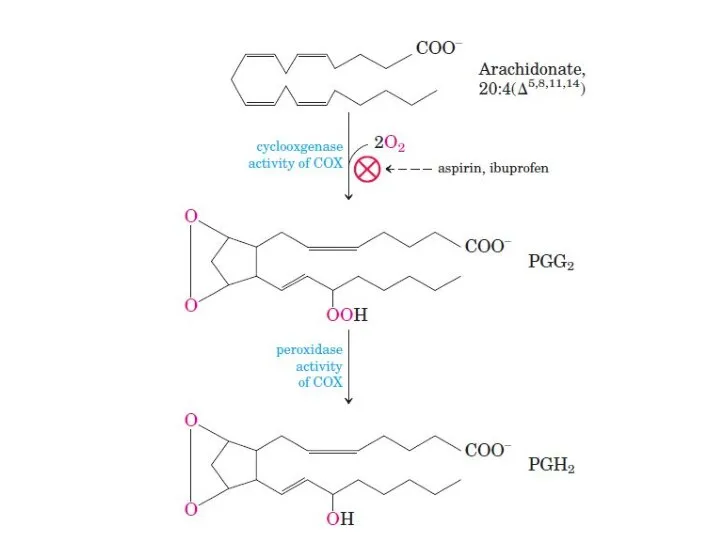
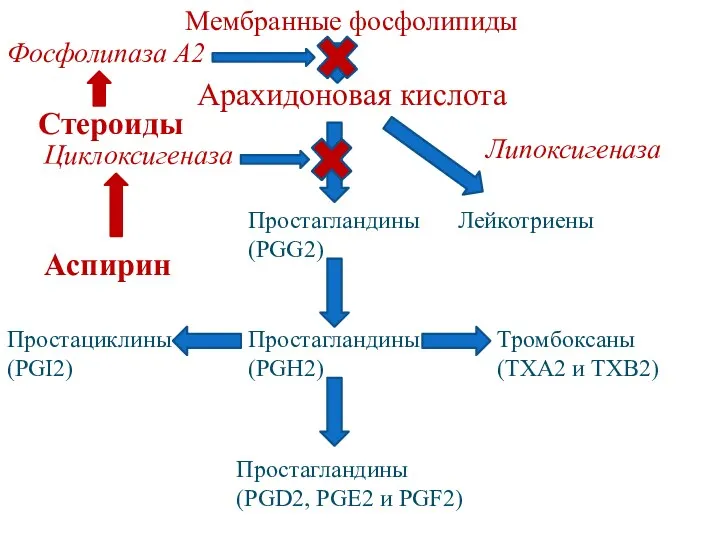
- 121. Эйкозаноиды Биологически активные вещества, аутокринные и паракринные гормоны. К ним относятся: простагландины; простациклины; тромбоксаны; лейкотриены. Синтезируются
- 122. Фосфолипаза А2
- 123. Синтез эйкозаноидов Арахидоновая кислота Липоксигеназа Циклоксигеназа Простагландины (PGG2) Простагландины (PGH2) Тромбоксаны (ТХА2 и ТХВ2) Простациклины (PGI2)
- 125. Механизм действия эйкозаноидов Являются гормонами местного действия. Действуют по аутокринному и/или паракринному механизмам. Действуют на клетки
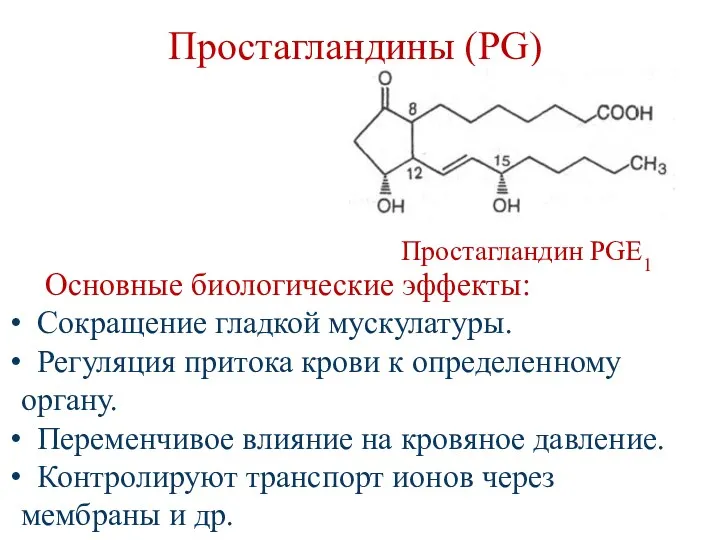
- 126. Простагландины (PG) представляют собой 20-углеродные жирные кислоты, содержащие 5-углеродное кольцо и гидрокси- и/или кетогруппы. Делятся на
- 127. Простагландины (PG) Простагландин PGЕ1 Основные биологические эффекты: Сокращение гладкой мускулатуры. Регуляция притока крови к определенному органу.
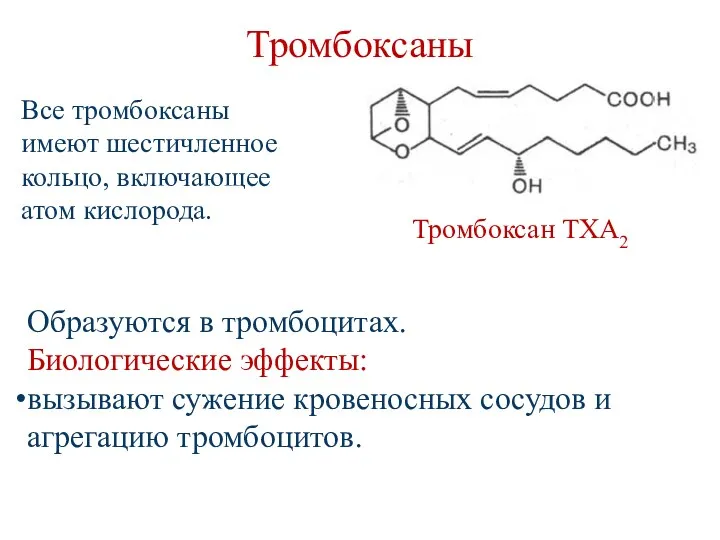
- 128. Тромбоксаны Тромбоксан TXA2 Образуются в тромбоцитах. Биологические эффекты: вызывают сужение кровеносных сосудов и агрегацию тромбоцитов. Все
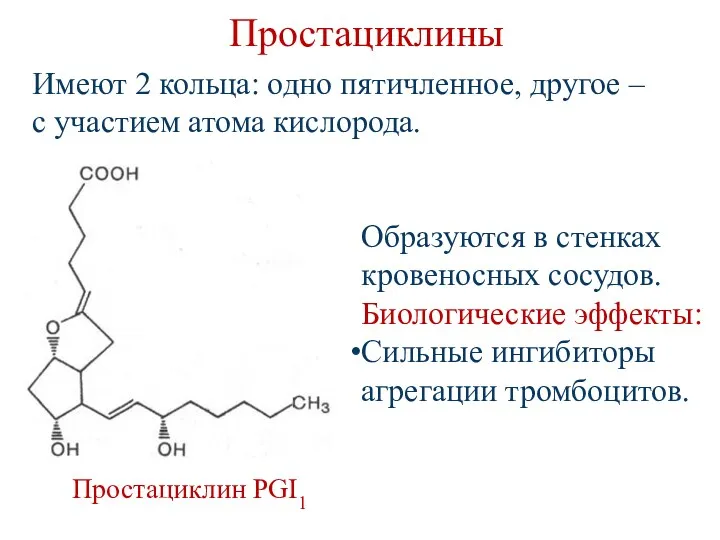
- 129. Простациклины Простациклин PGI1 Образуются в стенках кровеносных сосудов. Биологические эффекты: Сильные ингибиторы агрегации тромбоцитов. Имеют 2
- 130. Роль эйкозаноидов в тромбообразовании Тромбоксаны и простациклины являются антагонистами. Соотношение тромбоксана и простациклина определяет тромбообразование на
- 131. Роль эйкозаноидов в тромбообразовании При контакте тромбоцитов с поврежденной стенкой сосуда активируется фосфолипаза А2, в результате
- 132. !!! При потреблении пищи с преобладанием эйкозапентаеновой кислоты (рыбий жир) в клетках эндотелия синтезируются более сильные
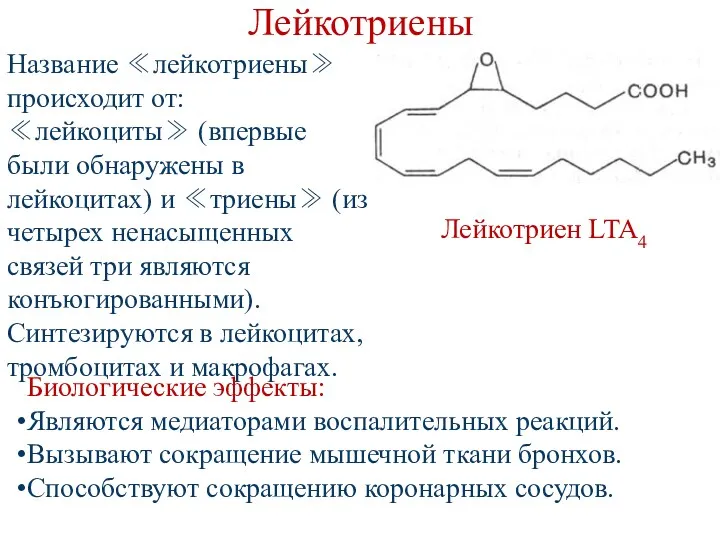
- 133. Лейкотриены Лейкотриен LTA4 Название ≪лейкотриены≫ происходит от: ≪лейкоциты≫ (впервые были обнаружены в лейкоцитах) и ≪триены≫ (из
- 134. Роль эйкозаноидов в воспалении Воспаление – реакция организма на повреждение или инфекцию, направленная на уничтожение инфекционного
- 135. Роль эйкозаноидов в воспалении Многие эйкозаноиды выполняют функцию медиаторов воспаления и действуют на всех этапах воспаления.
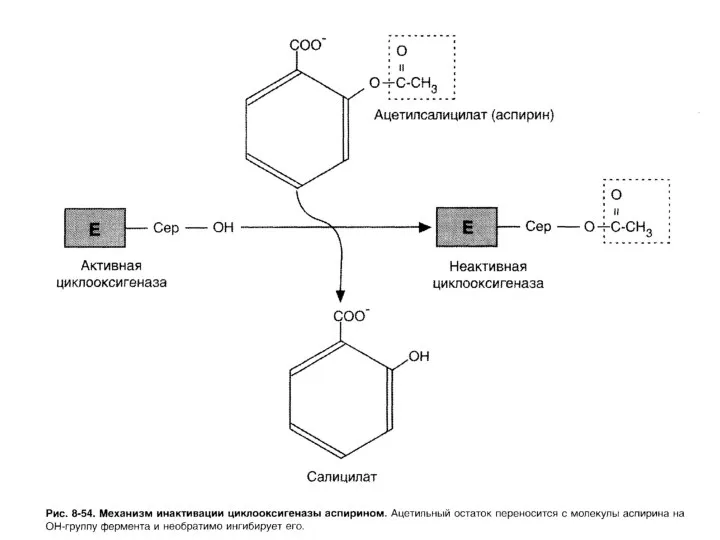
- 136. Лекарственные препараты – ингибиторы синтеза эйкозаноидов Нестероидные противовоспалительные препараты (аспирин, ибупрофен) ингибируют циклоксигеназу, что приводит к
- 137. Арахидоновая кислота Липоксигеназа Циклоксигеназа Простагландины (PGG2) Простагландины (PGH2) Тромбоксаны (ТХА2 и ТХВ2) Простациклины (PGI2) Лейкотриены Простагландины
- 139. Бронхиальная астма Развитие симптомов бронхиальной астмы (бронхоспазм и экссудация слизи в просвет бронхов) обусловлено избыточным синтезом
- 140. Использование эйкозаноидов в качестве лекарств Аналоги PGE1 и PGE2 подавляют секрецию соляной кислоты в желудке, блокируя
- 141. Жирорастворимые витамины Витамин А (антиксерофтальмический); ретинол. Витамин D (антирахитический); кальциферолы. Витамин Е (антистерильный, витамин размножения); токоферолы.
- 142. Витамин А (ретинол; антиксерофтальмический витамин) Источники: Животные продукты – рыбий жир, печень крупного рогатого скота и
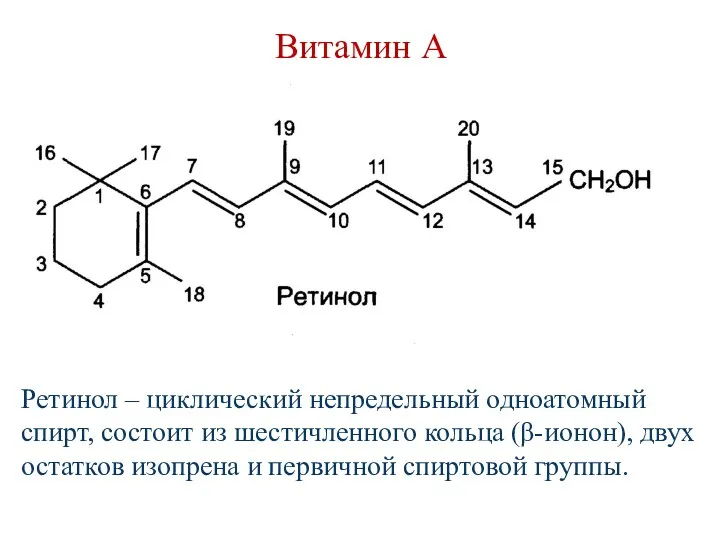
- 143. Витамин А Ретинол – циклический непредельный одноатомный спирт, состоит из шестичленного кольца (β-ионон), двух остатков изопрена
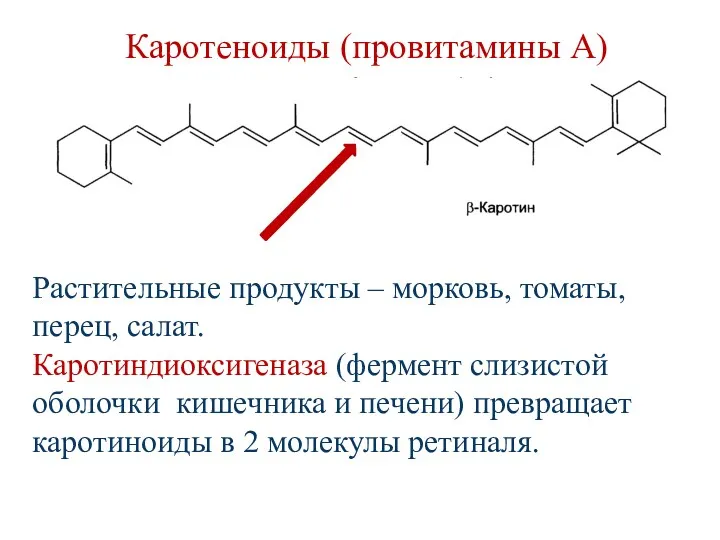
- 144. Каротеноиды (провитамины А) Растительные продукты – морковь, томаты, перец, салат. Каротиндиоксигеназа (фермент слизистой оболочки кишечника и
- 145. Биологические функции витамина А В организме ретинол превращается в ретиналь и ретиноевую кислоту. Ретиноевая кислота является
- 147. Гиповитаминоз А Ксерофтальмия, сухость роговой оболочки глаза (от греч. xeros – сухой, ophthalmos – глаз) вследствие
- 148. Гиповитаминоз А Торможение роста, снижение массы тела, общее истощение организма. Специфические поражения эпителия кожи (пролиферация и
- 149. Гипервитаминоз А Развивается при употреблении печени белого медведя, тюленя, моржа, у детей в результате приема больших
- 150. Витамин D3 (холекальциферол, антирахитический витамин) синтезируется из 7-дегидрохолестерина, содержащегося в коже человека, при солнечном облучении или
- 151. Витамин D2 (эргокальциферол) Эргостерин Витамин D2 (эргокальциферол) Эргостерин содержится в растительных маслах, дрожжах, грибах. УФ
- 152. Витамин D (кальциферол, антирахитический витамин) Распространение в природе: витамин D3 – продукты животного происхождения: сливочно масло,
- 153. Биологическая роль В организме из витамина D образуется 1,25-дигидроксихолекальциферол [1,25(OH)2D3] (кальцитриол) и 24,25-дигидроксихолекальциферол [24,25(ОН)2D3]. Гидроксилирование в
- 154. Биологическая роль Кальцитриол выполняет гормональную роль в регуляции обмена кальция и минерализации костной ткани. 1,25(OH)2D3 участвует
- 155. Гиповитаминоз D (рахит) Основные симптомы рахита у детей: остеомаляция – размягчение костей (О- или Х-образные формы).
- 156. Гипервитаминоз D При ≪ударной≫ терапии рахита и некоторых дерматозов (волчанка). Клинические проявления: Кальцификация тканей легких, почек,
- 157. Витамин Е, токоферолы α-токоферол (от греч. tokos – потомство, phero – несу). α-, β-, γ-, δ-токоферолы
- 158. Витамин Е Распространение в природе: Растительные масла (подсолнечное, хлопковое, соевое, кукурузное). Салат, капуста и семена злаков.
- 159. Биологическая роль витамина Е Активный природный антиоксидант: токоферолы разрушают наиболее реактивные формы кислорода и соответственно предохраняют
- 160. Гиповитаминоз Е Нарушение эмбриогенеза и дегенеративные изменения репродуктивных органов, что приводит к стерильности. У самок в
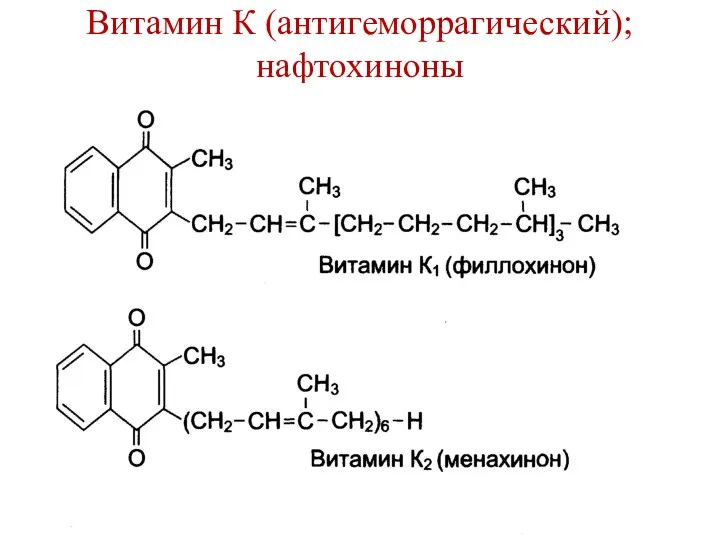
- 161. Витамин К (антигеморрагический); нафтохиноны Производные хинонов с боковыми изопреноидными цепями. Витамин K1 (филлохинон) – растительный. Витамин
- 162. Витамин К (антигеморрагический); нафтохиноны
- 163. Витамин К Распространение в природе: Растительные продукты – капуста, шпинат, тыква, зеленые томаты, арахисовое масло, ягоды
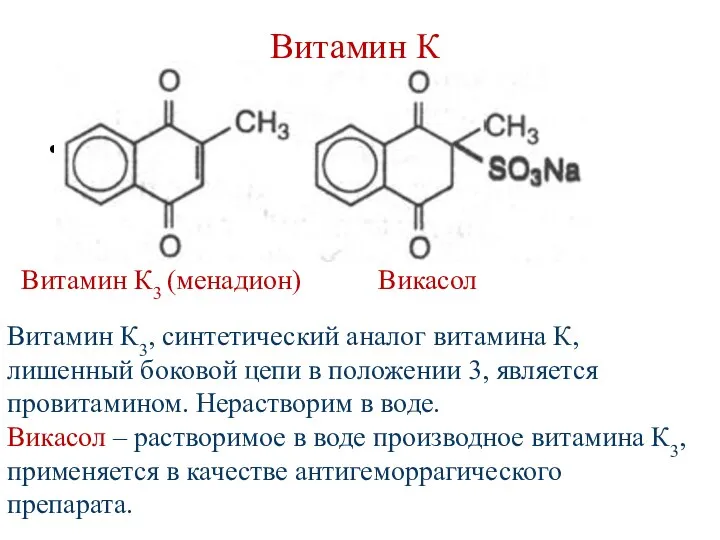
- 164. Витамин К Витамин К3 (менадион) Викасол Витамин К3, синтетический аналог витамина К, лишенный боковой цепи в
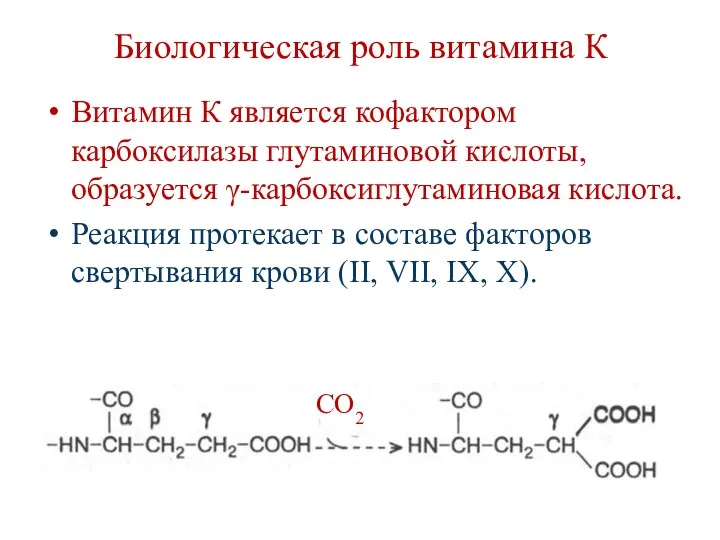
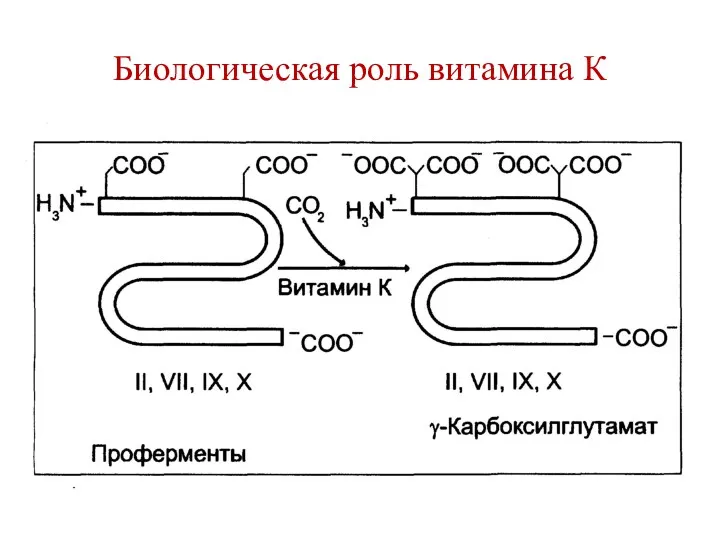
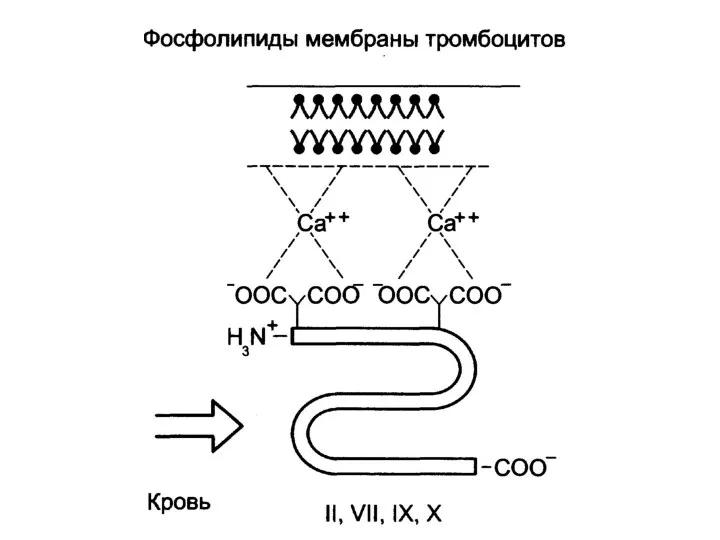
- 165. Биологическая роль витамина К Витамин К является кофактором карбоксилазы глутаминовой кислоты, образуется γ-карбоксиглутаминовая кислота. Реакция протекает
- 166. Биологическая роль витамина К
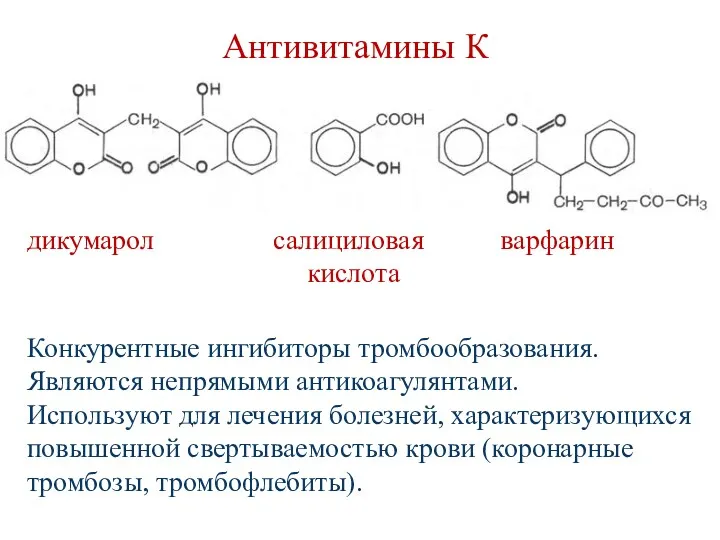
- 168. Антивитамины К дикумарол салициловая варфарин кислота Конкурентные ингибиторы тромбообразования. Являются непрямыми антикоагулянтами. Используют для лечения болезней,
- 169. Гиповитаминоз К Встречается редко (пища богата витамином К и он синтезируется кишечной микрофлорой). Развивается обычно при
- 170. Регуляция липидного обмена Адреналин, норадреналин и глюкагон увеличивают скорость липолиза в жировой ткани; в результате усиливается
- 171. Тканевой липолиз АТР цАМР ПК на ПК а ТГЛ на (Е-ОН) ТГЛ а (Е-Р) АТР АДР
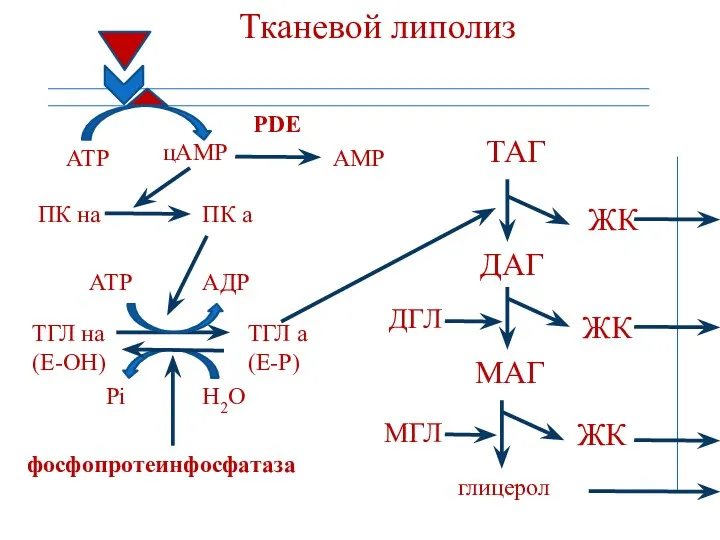
- 172. Регуляция липидного обмена Инсулин оказывает противоположное адреналину и глюкагону действие на липолиз. Ингибирование триглицеридлипазы: Активация фосфопротеинфосфатазы
- 173. Тканевой липолиз АТР цАМР ПК на ПК а ТГЛ на (Е-ОН) ТГЛ а (Е-Р) АТР АДР
- 174. Регуляция липидного обмена Инсулин стимулирует липогенез: Активирует липопротеинлипазу обеспечивает липогенез жирными кислотами. Стимулирует гликолиз (Глюкоза дигидроксиацетонфосфат
- 175. Регуляция липидного обмена Глюкокортикоиды: Стимулируют липолиз в конечностях и липогенез в других частях тела (лицо, туловище).
- 176. Регуляция липидного обмена Соматотропин: Усиливает липолиз в жировой ткани (за счет усиления синтеза аденилатциклазы и триглицеридлипазы).
- 177. Регуляция липидного обмена Тиреоидные гормоны: Повышают чувствительность β-адренорецепторов к действию адреналина и косвенно стимулирует липолиз в
- 178. Концентрация холестерина Нормальный уровень – менее 5,2 ммоль/л Пограничный уровень – 5,2-6,2 ммоль/л Высокий уровень –
- 179. ХОЛЕСТЕРИН ЛПНП Формула Friedwald : ХС ЛПНП, ммоль/л=Общий ХС – ХС ЛПВП – (0,45 хТАГ) ХС
- 180. ХОЛЕСТЕРИН ЛПВП Нормальная концентрация: Мужчины – 0,9-1,4 mmol/L; Женщины – 1,2-1,7 mmol/L.
- 181. Концентрация триглицеридов Нормальная Допустимая 1,6- 2,2 mmol/L Умеренно повышенная 2,2-2,8 mmol/L Высокая >2,8 mmol/L
- 182. Дислипопротеинемии Нарушения образования, транспорта и утилизации ЛП, сопровождаемые изменением плазменного уровня липидов. Делятся на гипер- и
- 183. Первичные гиперлипопортеидемии ГЛП I типа. Семейная гиперхиломикронемия. ГЛП II а типа. Семейная гипер-бета-липопротеинемия. Семейная гиперхолестеролемия. ГЛП
- 184. ГЛП I типа. Семейная гиперхиломикронемия Генетический дефект липопротеидлипазы или апобелка СII. Нарушение катаболизма хиломикронов. Повышение содержания
- 185. ГЛП II типа. Семейная гиперхолестеролемия. Мутациея гена ЛПНП-рецептора или апо В100. Частота гетерозиготных форм – 1
- 186. ГЛП II а типа. Семейная гиперхолестеролемия. Гомозиготные формы (1 на 1 000 000) Уровень ХС ˃15,5
- 187. Ксантомы сухожилий
- 188. Липоидная дуга роговицы
- 189. Первичные гиполипопортеидемии Абеталипопротеинемия Гипобеталипопротеинемия Анальфалипопротеинемия Семейная наследственная недостаточность ЛХАТ
- 190. Aн-альфа-липопротеинемия (болезнь Тангера) Мутация гена транспортного белка ABCA-1 (ATP-binding cassette transporter) – участвует в переносе холестерола
- 191. Дислипидемия при сахарном диабете “липидная триада”: • гипертриглицеридемия; • низкий уровнем ХС ЛПВП; • увеличение количества
- 192. Атеросклероз Хроническое прогрессирующее заболевание крупных и средних эластических и мышечно-эластических артерий, характеризующееся формированием в интиме атером
- 193. Главные факторы риска развития атеросклероза Дислипопротеинемии (как наследственные, так и приобретенные). Гипертензия (особенно у лиц старше
- 194. «Мягкие» факторы риска развития атеросклероза Ожирение (особенно абдоминального типа). Гиподинамия. Хронический стресс. Соревновательно-стрессорный тип жизнедеятельности. Гиперурикемия.
- 195. Атерогенная дислипидемия Увеличение концентрации общего холестерина. Повышение содержания ХС ЛПНП (особенно мелких частиц ЛПНП и химически
- 196. Роль ЛПНП в атерогенезе Перекисно-модифицированные ЛПНП распознаются и захватываются скэвенджер-рецепторами макрофагов (в переводе с англ. –
- 197. Oжирение ИМТ (индекс массы тела) ИМТ = вес(kg)/квадрат роста(m2)
- 198. Факторы риска ожирения Генетические дефекты (лептин, рецептор лептина, β3-адренергический рецептор, IRS-1). Алиментарные факторы (гиперкалорийность, высокое содержание
- 200. Скачать презентацию

Липиды
Органические вещества, нерастворимые в воде, но растворимые в органических растворителях
Липиды
Органические вещества, нерастворимые в воде, но растворимые в органических растворителях

Функции липидов
Энергетическая
Резервная
Структурная
Теплоизоляция
Механическая защита
Регуляторная
Витаминная
Функции липидов
Энергетическая
Резервная
Структурная
Теплоизоляция
Механическая защита
Регуляторная
Витаминная
Классификация липидов
I. Cтруктурный принцип
Омыляемые Неомыляемые
Холестерол
Простые: Сложные:
Ацилглицеролы
Классификация липидов
I. Cтруктурный принцип
Омыляемые Неомыляемые
Холестерол
Простые: Сложные:
Ацилглицеролы

Классификация липидов
II. Физико-химический принцип:
1. Нейтральные
2. Полярные (амфифильные)
III. Функциональный принцип:
Резервные
Структурные
Классификация липидов
II. Физико-химический принцип:
1. Нейтральные
2. Полярные (амфифильные)
III. Функциональный принцип:
Резервные
Структурные

Жирные кислоты
Насыщенные
Ненасыщенные:
Моноеновые
Полиеновые
Жирные кислоты
Насыщенные
Ненасыщенные:
Моноеновые
Полиеновые

Ацилглицеролы
Глицерол
Жирные кислоты
Ацилглицеролы
Глицерол
Жирные кислоты

Глицерофосфолипиды
Глицерол
Жирные кислоты
Фосфорная кислота
Аминоспирт
Глицерофосфолипиды
Глицерол
Жирные кислоты
Фосфорная кислота
Аминоспирт
Глицерофосфолипиды
Глицерофосфолипиды
Глицерофосфолипиды
Глицерофосфолипиды
Глицерофосфолипиды – предшественники вторичных посредников
Глицерофосфолипиды – предшественники вторичных посредников
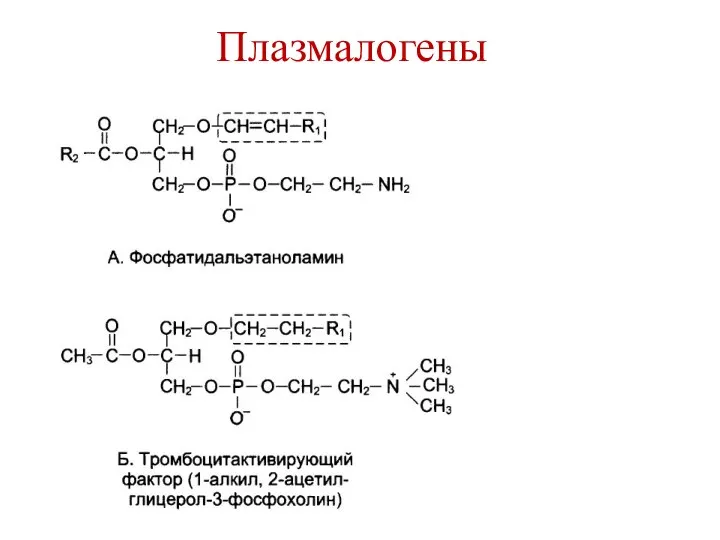
Плазмалогены
Плазмалогены

Сфингомиелины
Сфингозин
Жирная кислота
Фосфорная кислота
Аминоспирт
Сфингомиелины
Сфингозин
Жирная кислота
Фосфорная кислота
Аминоспирт
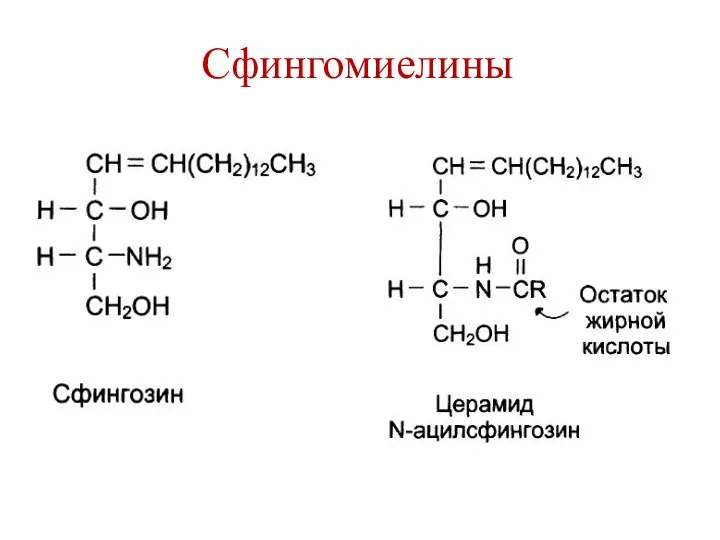
Сфингомиелины
Сфингомиелины
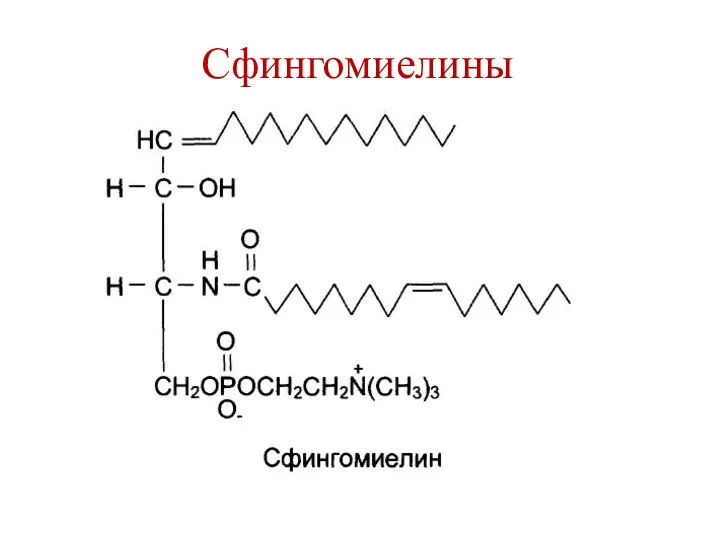
Сфингомиелины
Сфингомиелины

Гликолипиды
Сфингозин
Жирная кислота
Углеводы
Классификация
Цереброзиды
Сульфатиды
Глобозиды
Ганглиозиды
Гликолипиды
Сфингозин
Жирная кислота
Углеводы
Классификация
Цереброзиды
Сульфатиды
Глобозиды
Ганглиозиды
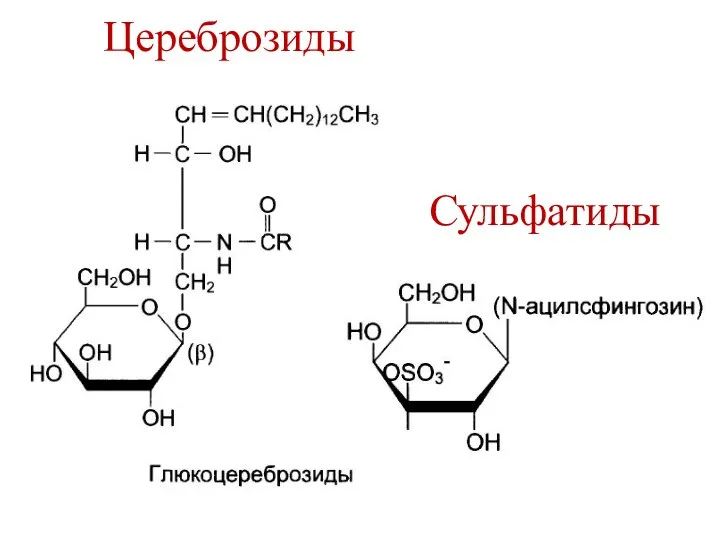
Цереброзиды
Сульфатиды
Цереброзиды
Сульфатиды
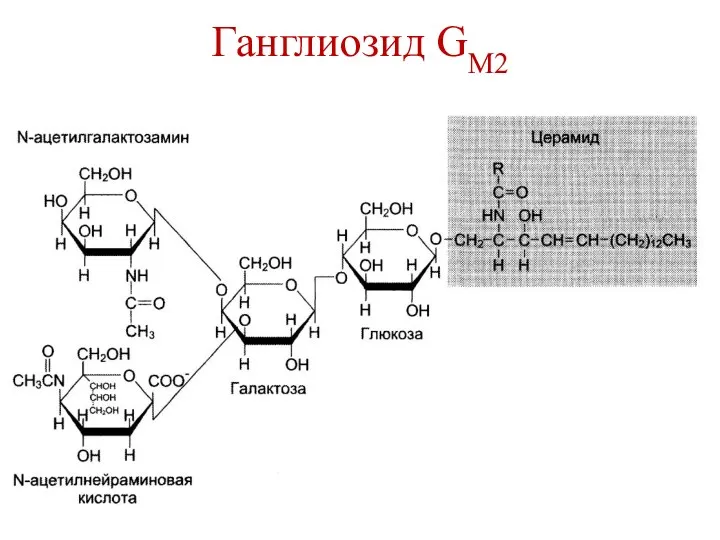
Ганглиозид GM2
Ганглиозид GM2
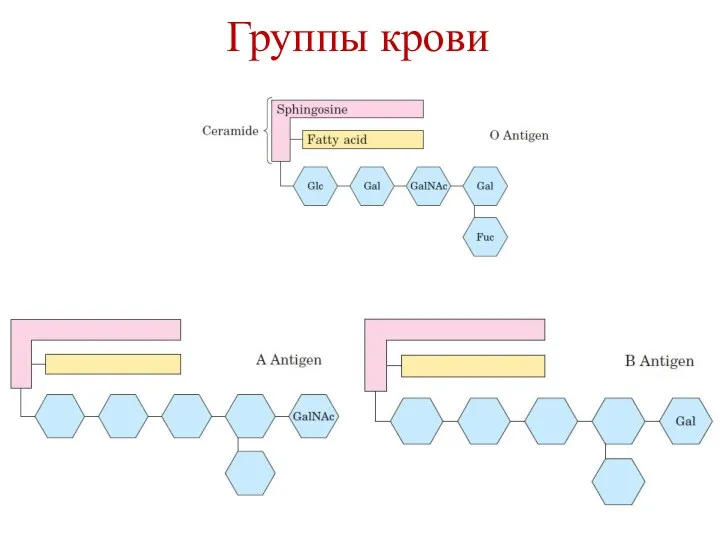
Группы крови
Группы крови
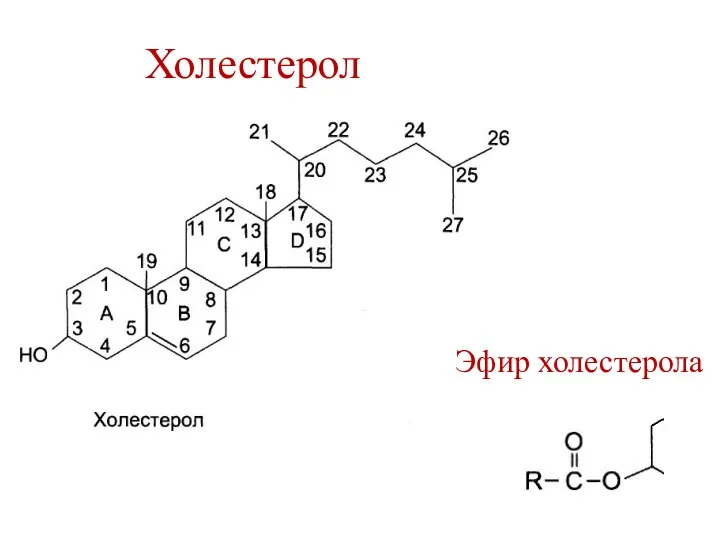
Холестерол
Эфир холестерола
Холестерол
Эфир холестерола
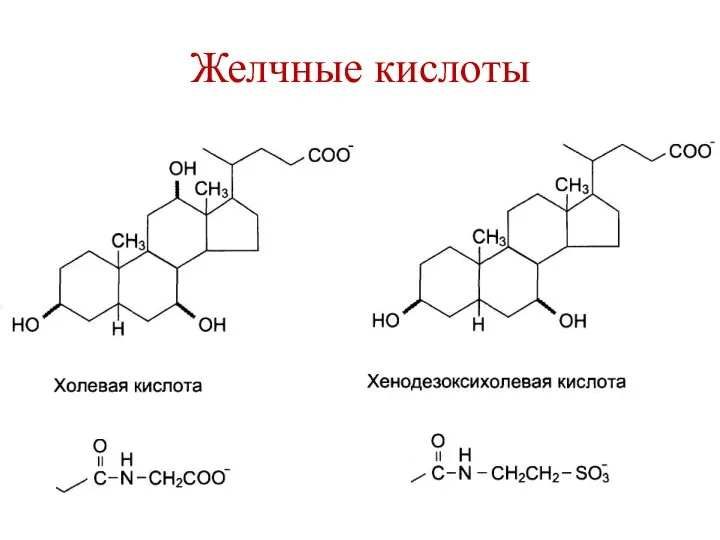
Желчные кислоты
Желчные кислоты

Переваривание липидов.
Плазменные липопротеины.
Катаболизм липидов
Переваривание липидов.
Плазменные липопротеины.
Катаболизм липидов

Основными пищевыми липидами являются
Триацилглицеролы
Холестерол и холестериды
Фосфолипиды
Значение пищевых липидов:
Суточная норма липидов
Основными пищевыми липидами являются
Триацилглицеролы
Холестерол и холестериды
Фосфолипиды
Значение пищевых липидов:
Суточная норма липидов

Переваривание липидов в ЖКТ
У детей начинается в ротовой полости под действием
Переваривание липидов в ЖКТ
У детей начинается в ротовой полости под действием

Переваривание липидов в кишечнике
(необходимые условия)
Щелочная среда – рН = 8
Переваривание липидов в кишечнике
(необходимые условия)
Щелочная среда – рН = 8

Желчные кислоты
Синтезируются в печени из холестерола
Выделяются с желчью в двенадцатиперстную кишку
Функции
Желчные кислоты
Синтезируются в печени из холестерола
Выделяются с желчью в двенадцатиперстную кишку
Функции
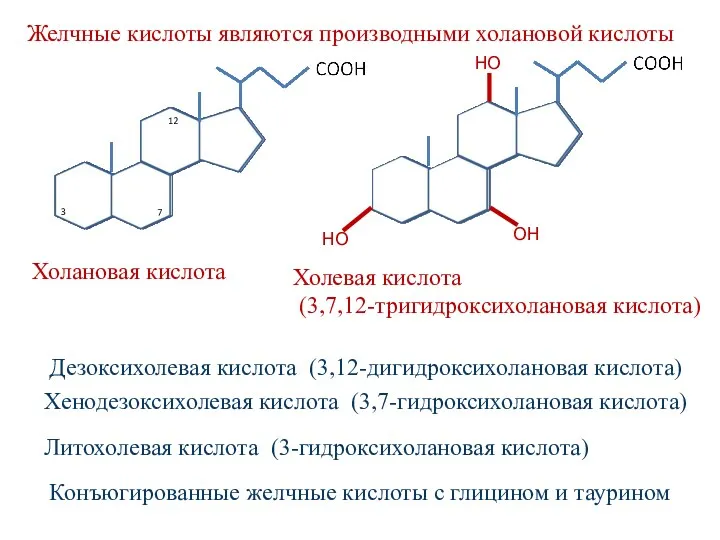
Холановая кислота
Желчные кислоты являются производными холановой кислоты
Холевая кислота
(3,7,12-тригидроксихолановая кислота)
НО
НО
ОН
3
7
12
Дезоксихолевая кислота
Холановая кислота
Желчные кислоты являются производными холановой кислоты
Холевая кислота
(3,7,12-тригидроксихолановая кислота)
НО
НО
ОН
3
7
12
Дезоксихолевая кислота
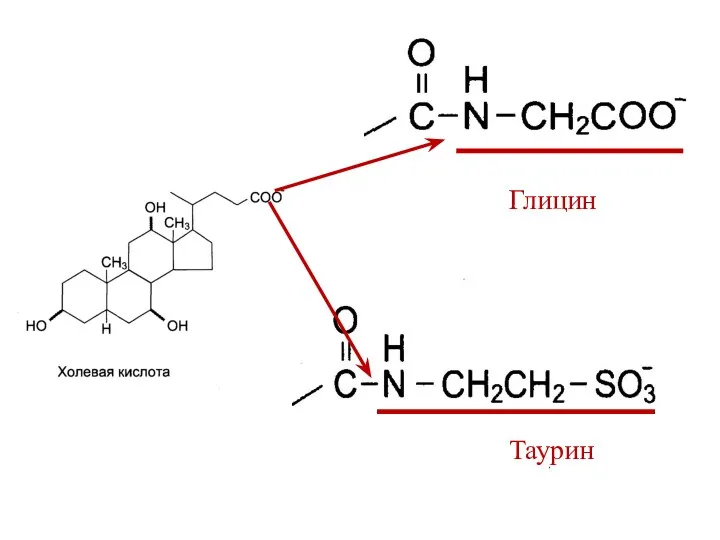
Таурин
Глицин
Таурин
Глицин
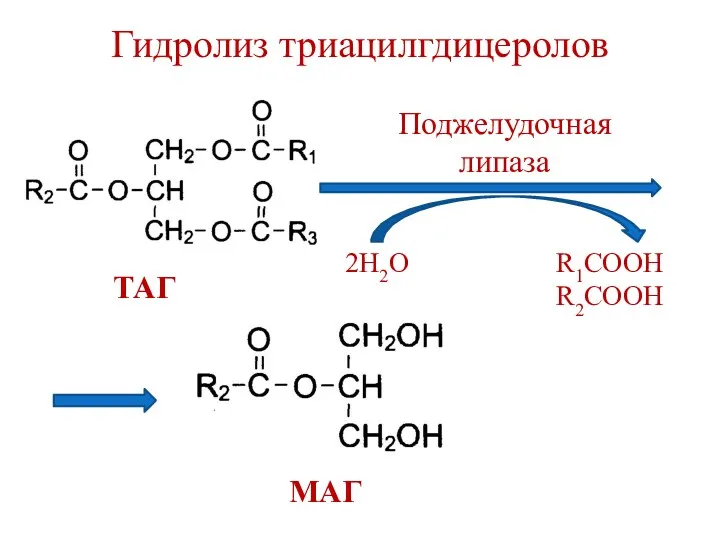
Гидролиз триацилгдицеролов
Поджелудочная
липаза
2Н2О
R1COOH
R2COOH
ТАГ
МАГ
Гидролиз триацилгдицеролов
Поджелудочная
липаза
2Н2О
R1COOH
R2COOH
ТАГ
МАГ
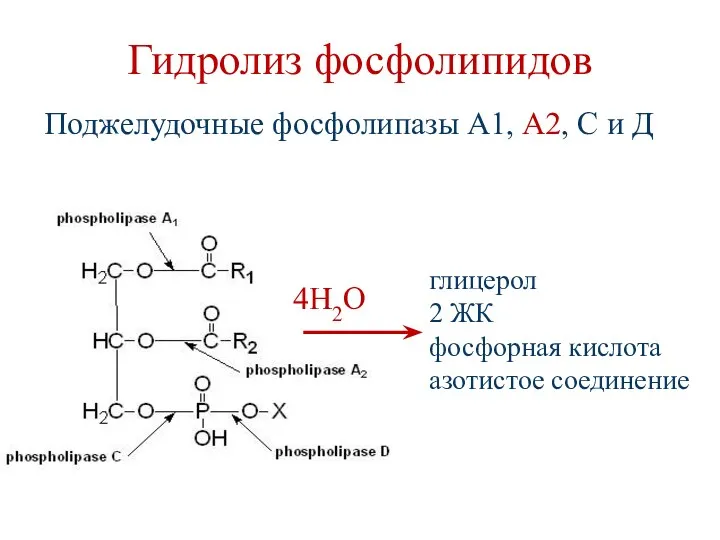
Гидролиз фосфолипидов
Поджелудочные фосфолипазы А1, А2, С и Д
4Н2О
глицерол
2 ЖК
фосфорная кислота
азотистое
Гидролиз фосфолипидов
Поджелудочные фосфолипазы А1, А2, С и Д
4Н2О
глицерол
2 ЖК
фосфорная кислота
азотистое

Гидролиз фосфолипидов
Основным ферментом является фосфолипаза А2, под действием которой из фосфолипидов
Гидролиз фосфолипидов
Основным ферментом является фосфолипаза А2, под действием которой из фосфолипидов
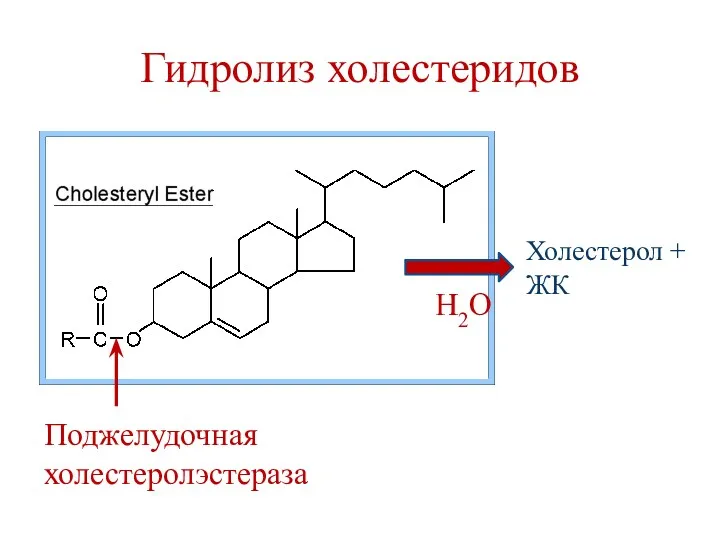
Гидролиз холестеридов
Поджелудочная
холестеролэстераза
Холестерол + ЖК
Н2О
Гидролиз холестеридов
Поджелудочная
холестеролэстераза
Холестерол + ЖК
Н2О

Всасывание продуктов гидролиза липидов
Глицерол, коротко- и среднецепочечные ЖК – свободная диффузия.
МАГ,
Всасывание продуктов гидролиза липидов
Глицерол, коротко- и среднецепочечные ЖК – свободная диффузия.
МАГ,

Всасывание продуктов гидролиза липидов
В энтероцитах мицеллы распадаются.
Продукты гидролиза липидов подвергаются
Всасывание продуктов гидролиза липидов
В энтероцитах мицеллы распадаются.
Продукты гидролиза липидов подвергаются

Регуляция переваривания липидов
Секретин
стимулирует выделение печенью и поджелудочной железой водянистого раствора
Регуляция переваривания липидов
Секретин
стимулирует выделение печенью и поджелудочной железой водянистого раствора

Нарушения переваривания и всасывания липидов
Стеаторея – появление в фекалиях
Нарушения переваривания и всасывания липидов
Стеаторея – появление в фекалиях

Ресинтез липидов в энтероцитах
Моноацилглицерол + 2 R-CO-S-CoA триацилглицерол + 2 НS-CoA
Ресинтез липидов в энтероцитах
Моноацилглицерол + 2 R-CO-S-CoA триацилглицерол + 2 НS-CoA

Плазменные липопротеины
липидно-белковые комплексы, транспортные формы липидов.
Все липопротеины имеют
Плазменные липопротеины
липидно-белковые комплексы, транспортные формы липидов.
Все липопротеины имеют
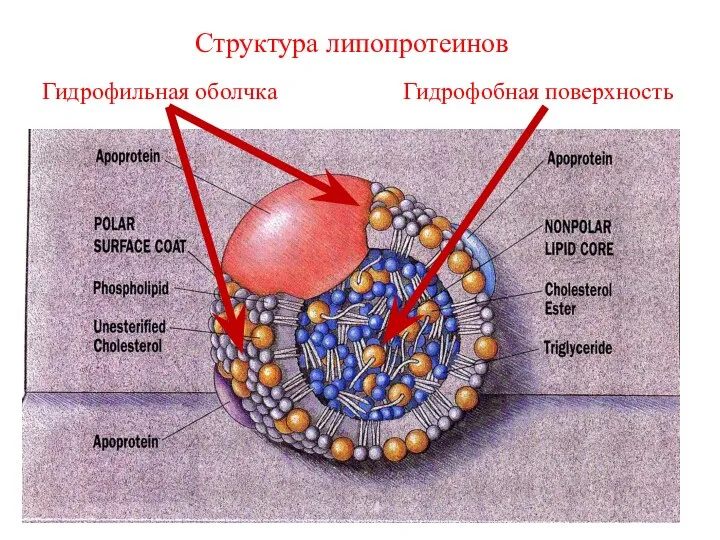
Гидрофильная оболчка Гидрофобная поверхность
Структура липопротеинов
Гидрофильная оболчка Гидрофобная поверхность
Структура липопротеинов

Липопротеины различаются по содержанию липидов и апопротеинов, что определяет
Липопротеины различаются по содержанию липидов и апопротеинов, что определяет
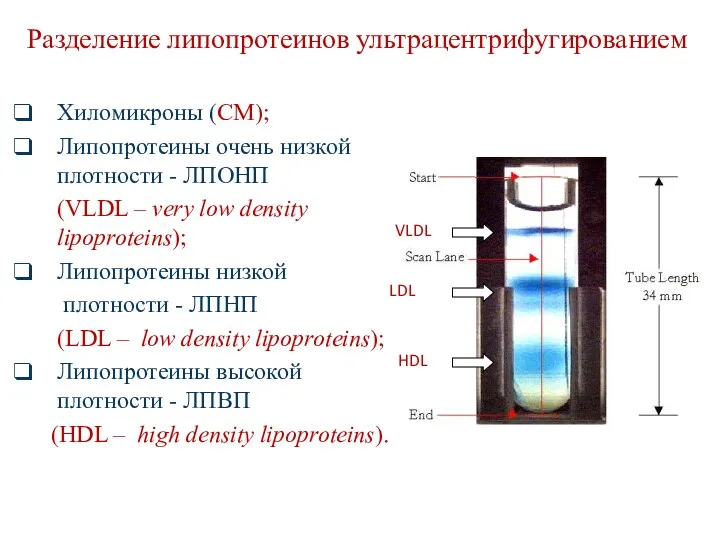
Разделение липопротеинов ультрацентрифугированием
HDL
VLDL
LDL
Хиломикроны (CM);
Липопротеины очень низкой плотности - ЛПОНП
(VLDL
Разделение липопротеинов ультрацентрифугированием
HDL
VLDL
LDL
Хиломикроны (CM);
Липопротеины очень низкой плотности - ЛПОНП
(VLDL
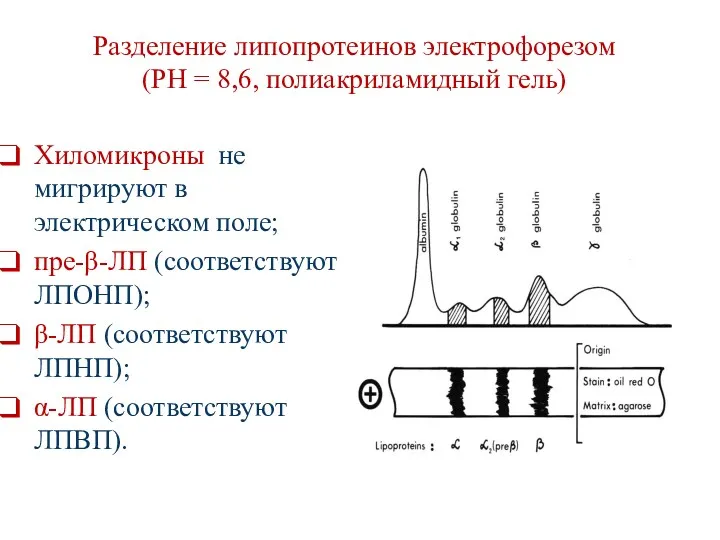
Разделение липопротеинов электрофорезом
(PH = 8,6, полиакриламидный гель)
Хиломикроны не мигрируют в электрическом
Разделение липопротеинов электрофорезом
(PH = 8,6, полиакриламидный гель)
Хиломикроны не мигрируют в электрическом

Обмен хиломикронов
образуются в эпителии кишечника;
содержат много ТАГ (85%), содержат апопротеин В-48
Обмен хиломикронов
образуются в эпителии кишечника;
содержат много ТАГ (85%), содержат апопротеин В-48

Обмен хиломикронов
после синтеза секретируются в лимфу и через грудной проток попадают
Обмен хиломикронов
после синтеза секретируются в лимфу и через грудной проток попадают

Обмен хиломикронов
ЛПЛ гидролизует ТАГ из хиломикронов.
ЖК поступают в скелетные мышцы,
Обмен хиломикронов
ЛПЛ гидролизует ТАГ из хиломикронов.
ЖК поступают в скелетные мышцы,
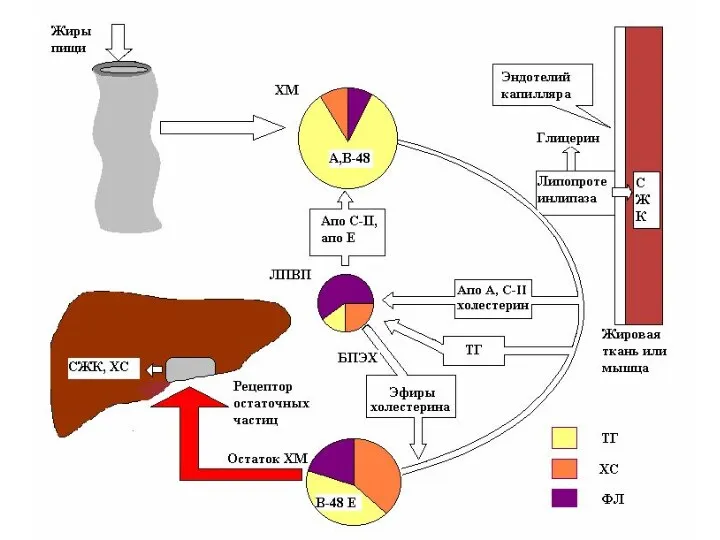

Обмен ЛПОНП
Образуются в печени;
Содержат много ТАГ (65%) и апоВ-100 в качестве
Обмен ЛПОНП
Образуются в печени;
Содержат много ТАГ (65%) и апоВ-100 в качестве

Обмен ЛПОНП
Катаболизм ЛПНП происходит под действием фермента ЛПЛ (аналогично хиломикронам).
после гидролиза
Обмен ЛПОНП
Катаболизм ЛПНП происходит под действием фермента ЛПЛ (аналогично хиломикронам).
после гидролиза
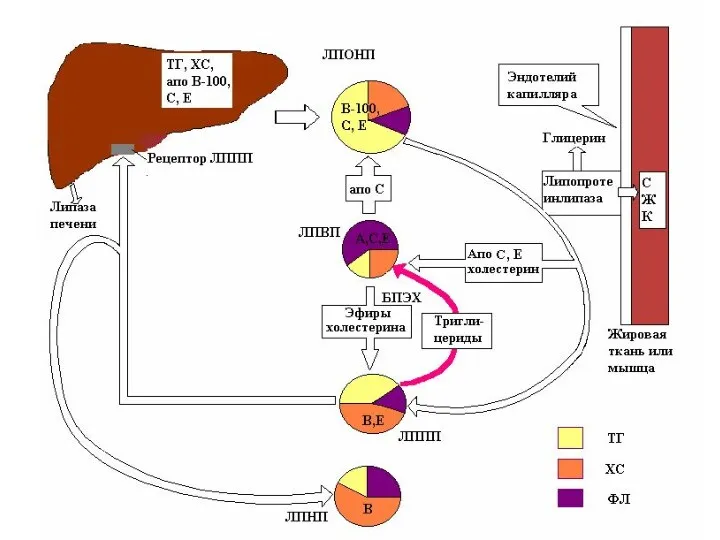

Обмен ЛПНП
Образуются в кровотоке из ЛПОНП.
Являются основными переносчиками эндогенного ХС к
Обмен ЛПНП
Образуются в кровотоке из ЛПОНП.
Являются основными переносчиками эндогенного ХС к

Обмен ЛПНП
Регулируемый путь катаболизма – связывание с апо В/Е-рецепторами печени, клеток
Обмен ЛПНП
Регулируемый путь катаболизма – связывание с апо В/Е-рецепторами печени, клеток
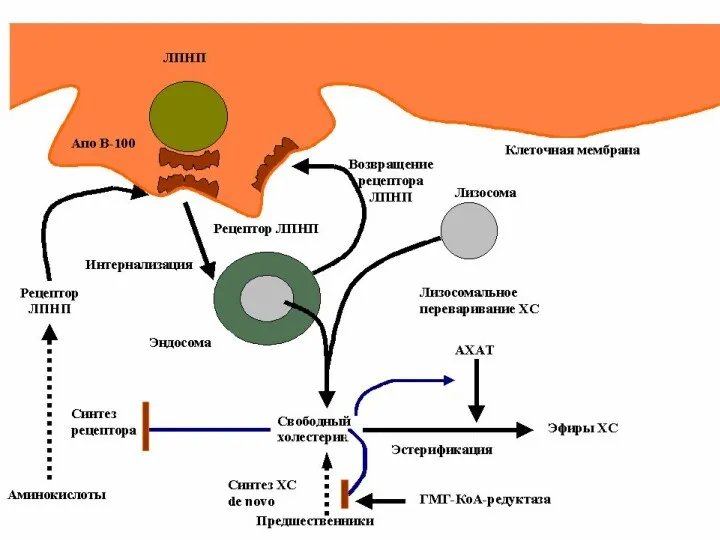

Обмен ЛПНП
Альтернативный путь (нерегулируемый) – характерен для окисленных ЛПНП.
Перекисно-модифицированные ЛПНП
Обмен ЛПНП
Альтернативный путь (нерегулируемый) – характерен для окисленных ЛПНП.
Перекисно-модифицированные ЛПНП

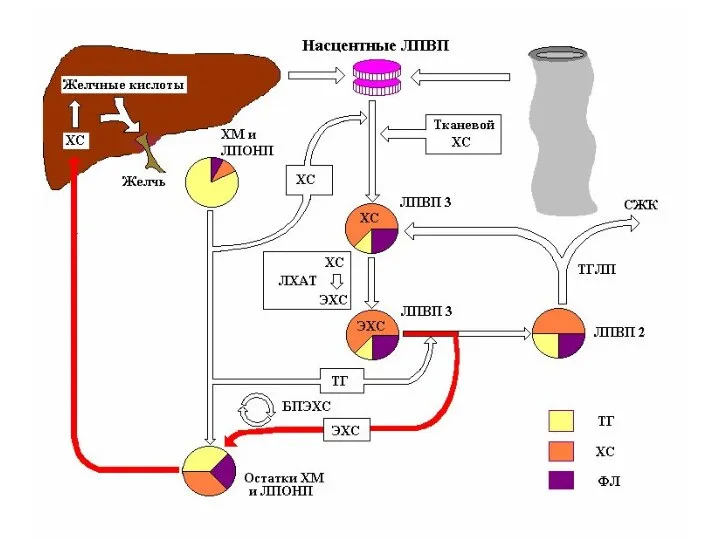
Обмен ЛПВП
ЛПВП образуются в печени в виде незрелых дисковидных частиц, состоящих
Обмен ЛПВП
ЛПВП образуются в печени в виде незрелых дисковидных частиц, состоящих

Обмен ЛПВП
ЛПВП – хорошие акцепторы свободного ХС из периферических тканей.
Свободный
Обмен ЛПВП
ЛПВП – хорошие акцепторы свободного ХС из периферических тканей.
Свободный

Обмен ЛПВП
Эфиры ХС перемещаются с поверхности частиц ЛПВП в гидрофобное ядро,
Обмен ЛПВП
Эфиры ХС перемещаются с поверхности частиц ЛПВП в гидрофобное ядро,

Мобилизация жиров из жировой ткани (тканевой липолиз)
Резервные липиды обеспечивают энергией организм
Мобилизация жиров из жировой ткани (тканевой липолиз)
Резервные липиды обеспечивают энергией организм
Тканевой липолиз
R
H
A
АТР
цАМР
ПК на
ПК а
ТГЛ на
(Е-ОН)
ТГЛ а
(Е-Р)
АТР
АДР
ТАГ
МАГ
ДАГ
глицерол
ЖК
ЖК
ЖК
гормон
(адреналин, глюкагон)
- рецептор
- аденилатциклаза
ДГЛ
МГЛ
Тканевой липолиз
R
H
A
АТР
цАМР
ПК на
ПК а
ТГЛ на
(Е-ОН)
ТГЛ а
(Е-Р)
АТР
АДР
ТАГ
МАГ
ДАГ
глицерол
ЖК
ЖК
ЖК
гормон
(адреналин, глюкагон)
- рецептор
- аденилатциклаза
ДГЛ
МГЛ

Регуляция липолиза
Адреналин, глюкагон
стимулируют липолиз (активируют триглицеридлипазу – фосфорилирование фермента);
Глюкокортикоиды (кортизол)
стимулируют липолиз
Регуляция липолиза
Адреналин, глюкагон
стимулируют липолиз (активируют триглицеридлипазу – фосфорилирование фермента);
Глюкокортикоиды (кортизол)
стимулируют липолиз
Использование глицерола
Глюконеогенез
Окисление
Синтез ТАГ в печени
Глюконеогенез из глицерола
2 Глицерол
2 Глицерол-3-Р
Глицеральдегид
-3-Р
2 Дигидрокси-
ацетон-Р
Фруктозо-1,6-дифосфат
Фруктозо-6-фосфат
Глюкозо-6-фосфат
Глюкоза
2АТР
2АДР
2NAD+
2NADH+H+
H2O
H2O
H3PO4
H3PO4
Использование глицерола
Глюконеогенез
Окисление
Синтез ТАГ в печени
Глюконеогенез из глицерола
2 Глицерол
2 Глицерол-3-Р
Глицеральдегид
-3-Р
2 Дигидрокси-
ацетон-Р
Фруктозо-1,6-дифосфат
Фруктозо-6-фосфат
Глюкозо-6-фосфат
Глюкоза
2АТР
2АДР
2NAD+
2NADH+H+
H2O
H2O
H3PO4
H3PO4
Окисление глицерола
Глицеральдегид-3-Р
1,3-дифосфоглицерат
3-фосфоглицерат
2-фосфоглицерат
фосфоенолпируват
пируват
Ацетил-СоА
Цикл Кребса
NADH (3 ATP)
NADH (3 ATP)
22 ATP
1 ATP
3 ATP
1 ATP
-1
Окисление глицерола
Глицеральдегид-3-Р
1,3-дифосфоглицерат
3-фосфоглицерат
2-фосфоглицерат
фосфоенолпируват
пируват
Ацетил-СоА
Цикл Кребса
NADH (3 ATP)
NADH (3 ATP)
22 ATP
1 ATP
3 ATP
1 ATP
-1

Бета-окисление жирных кислот
Локализация – скелетные мышцы, миокард, печень, почки
Не происходит в
Бета-окисление жирных кислот
Локализация – скелетные мышцы, миокард, печень, почки
Не происходит в
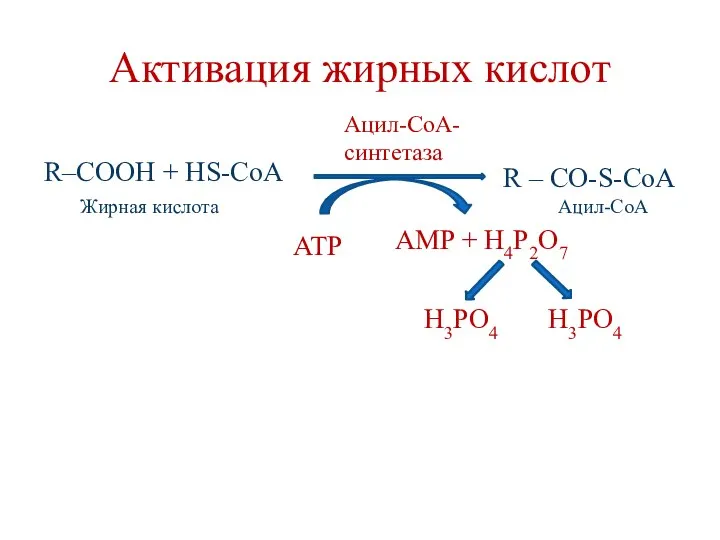
Активация жирных кислот
R–COOH + HS-CoA
ATP
AMP + H4P2O7
H3PO4
H3PO4
Жирная кислота
Ацил-СоА
Ацил-СоА-
синтетаза
R – CO-S-CoA
Активация жирных кислот
R–COOH + HS-CoA
ATP
AMP + H4P2O7
H3PO4
H3PO4
Жирная кислота
Ацил-СоА
Ацил-СоА-
синтетаза
R – CO-S-CoA
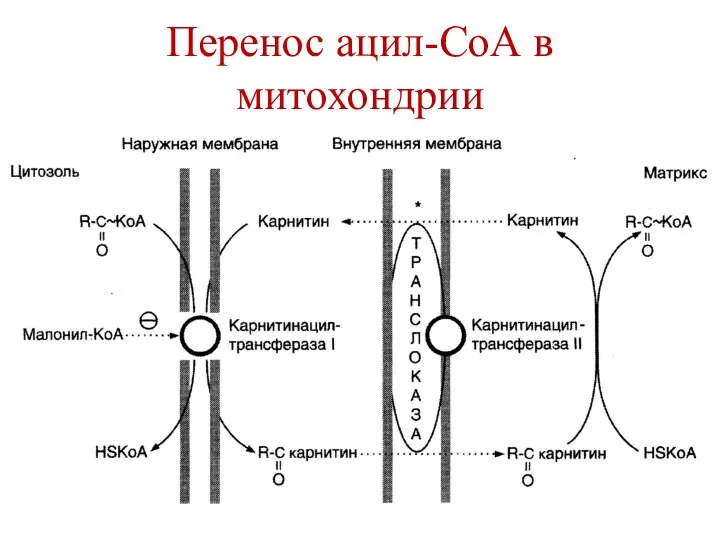
Перенос ацил-СоА в митохондрии
Перенос ацил-СоА в митохондрии
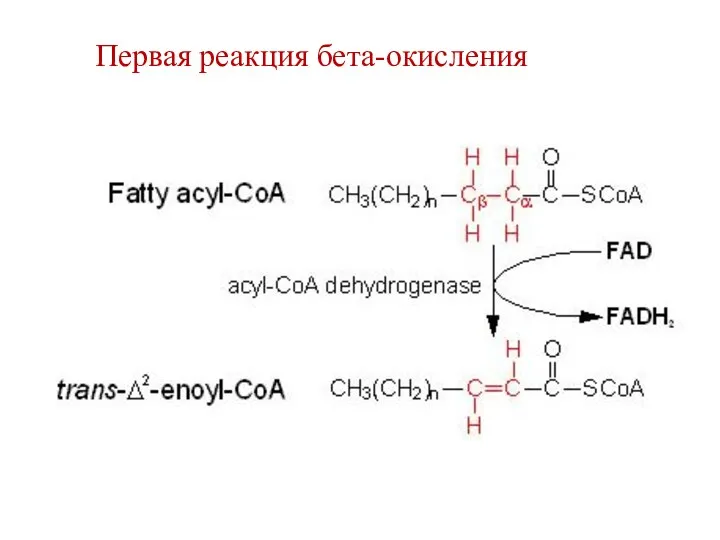
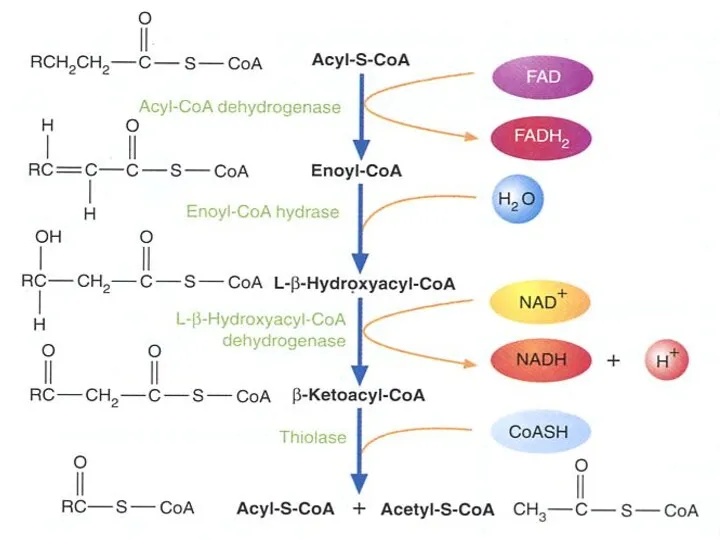
Первая реакция бета-окисления
Первая реакция бета-окисления
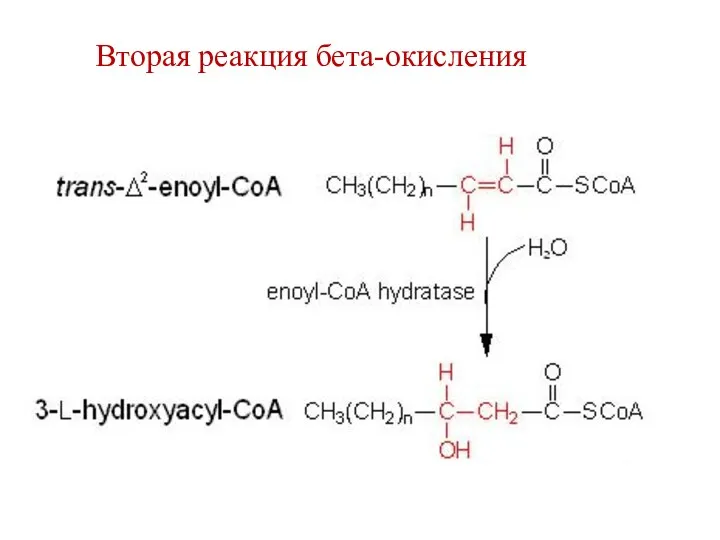
Вторая реакция бета-окисления
Вторая реакция бета-окисления
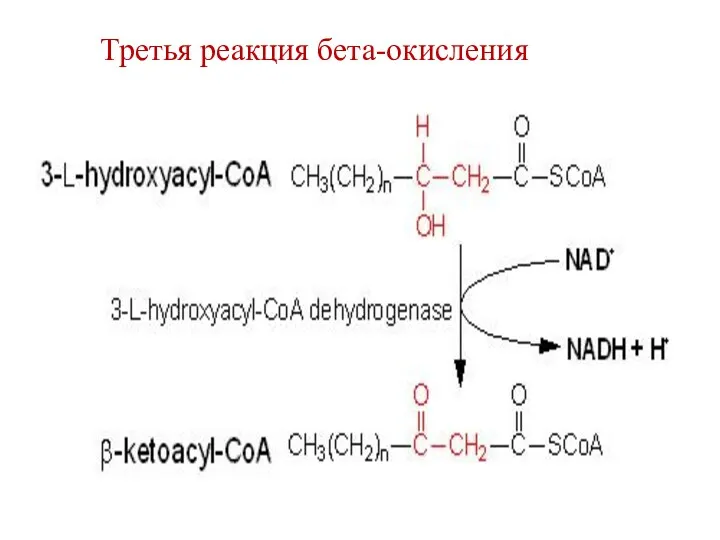
Третья реакция бета-окисления
Третья реакция бета-окисления
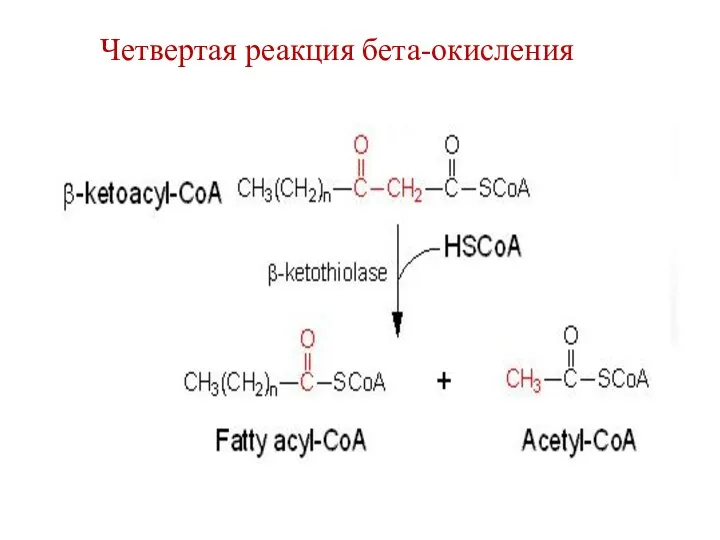
Четвертая реакция бета-окисления
Четвертая реакция бета-окисления
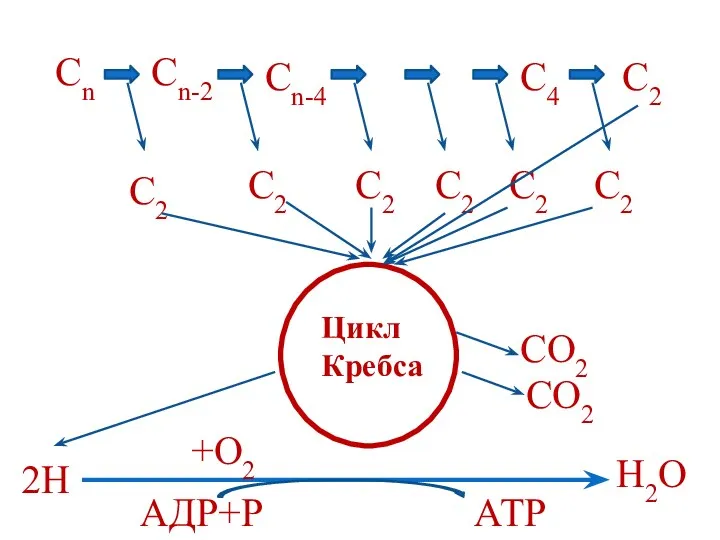
Сn
Сn-4
С4
С2
Сn-2
С2
С2
С2
С2
С2
С2
Цикл Кребса
СО2
СО2
2Н
+О2
Н2О
АТР
АДР+Р
Сn
Сn-4
С4
С2
Сn-2
С2
С2
С2
С2
С2
С2
Цикл Кребса
СО2
СО2
2Н
+О2
Н2О
АТР
АДР+Р

Энергетический баланс бета-окисления
N – количество атомов углерода ЖК
N/2-1 – количество циклов
Энергетический баланс бета-окисления
N – количество атомов углерода ЖК
N/2-1 – количество циклов

Особенности окисления
ненасыщенных жирных кислот
При окислении ненасыщенных ЖК, после 3-х циклов
Особенности окисления
ненасыщенных жирных кислот
При окислении ненасыщенных ЖК, после 3-х циклов
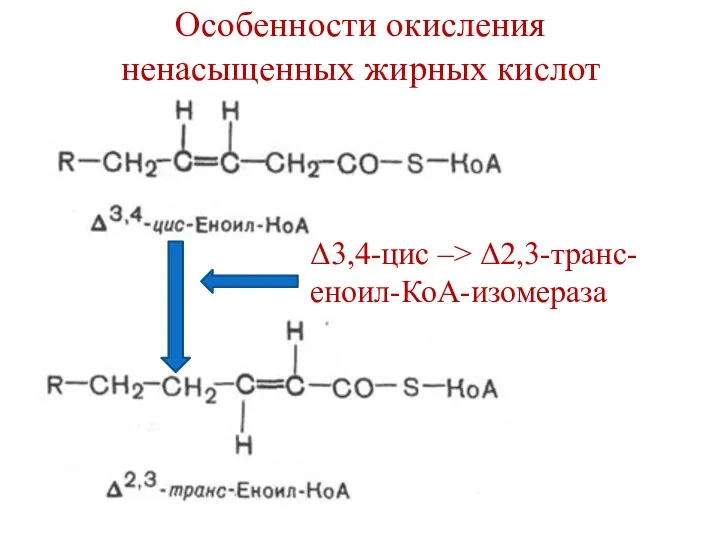
Особенности окисления
ненасыщенных жирных кислот
Δ3,4-цис –> Δ2,3-транс-еноил-КоА-изомераза
Особенности окисления
ненасыщенных жирных кислот
Δ3,4-цис –> Δ2,3-транс-еноил-КоА-изомераза

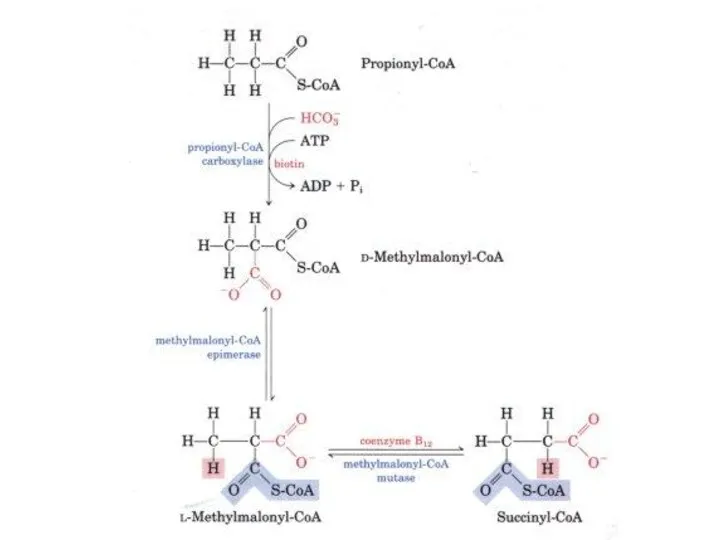
Особенности окисления жирных кислот с нечетным числом атомов углерода
В последнем цикле
Особенности окисления жирных кислот с нечетным числом атомов углерода
В последнем цикле

Особенности окисления жирных кислот в пероксисомах
Характерно для ЖК с 20-26 атомами
Особенности окисления жирных кислот в пероксисомах
Характерно для ЖК с 20-26 атомами

Биосинтез липидов
Биосинтез липидов

Биосинтез жирных кислот
Локализация процесса – печень, жировая ткань, лактирующая молочная
Биосинтез жирных кислот
Локализация процесса – печень, жировая ткань, лактирующая молочная
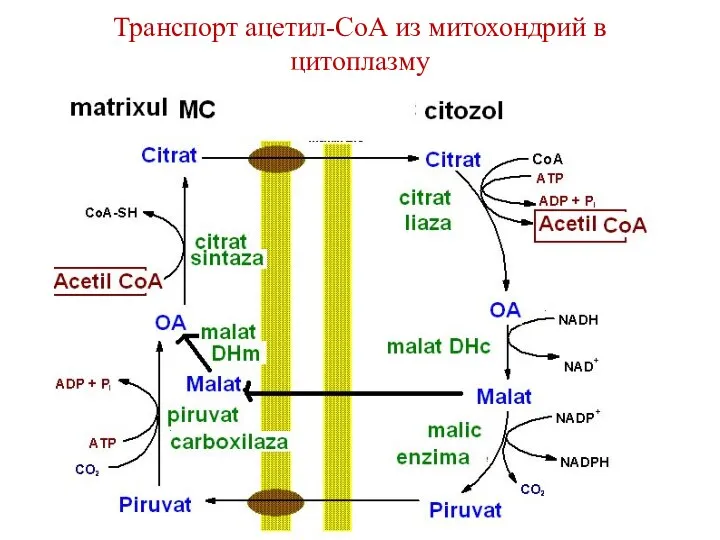
Транспорт ацетил-СоА из митохондрий в цитоплазму
Транспорт ацетил-СоА из митохондрий в цитоплазму
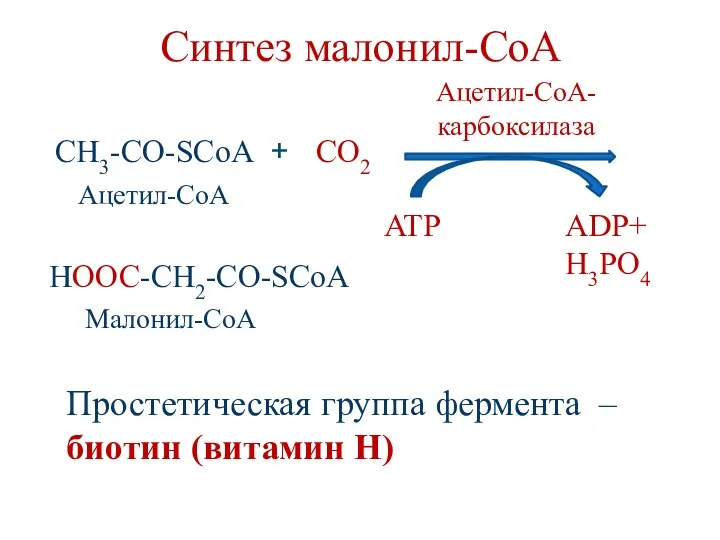
Синтез малонил-СоА
CH3-CO-SCoA
+
HOOC-CH2-CO-SCoA
CO2
ATP
ADP+
H3PO4
Ацетил-CoA-
карбоксилаза
Простетическая группа фермента –
биотин (витамин Н)
Малонил-CoA
Ацетил-CoA
Синтез малонил-СоА
CH3-CO-SCoA
+
HOOC-CH2-CO-SCoA
CO2
ATP
ADP+
H3PO4
Ацетил-CoA-
карбоксилаза
Простетическая группа фермента –
биотин (витамин Н)
Малонил-CoA
Ацетил-CoA

Собственно-биосинтез жирных кислот
Происходит под действием мультиферментного комплекса, синтаза жирных кислот (пальмитат
Собственно-биосинтез жирных кислот
Происходит под действием мультиферментного комплекса, синтаза жирных кислот (пальмитат
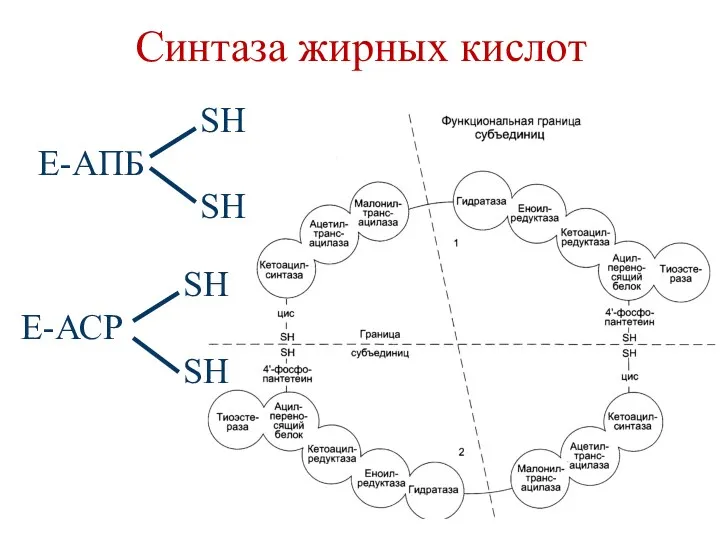
Синтаза жирных кислот
SН
Е-АПБ
SH
SН
Е-АСР
SH
Синтаза жирных кислот
SН
Е-АПБ
SH
SН
Е-АСР
SH
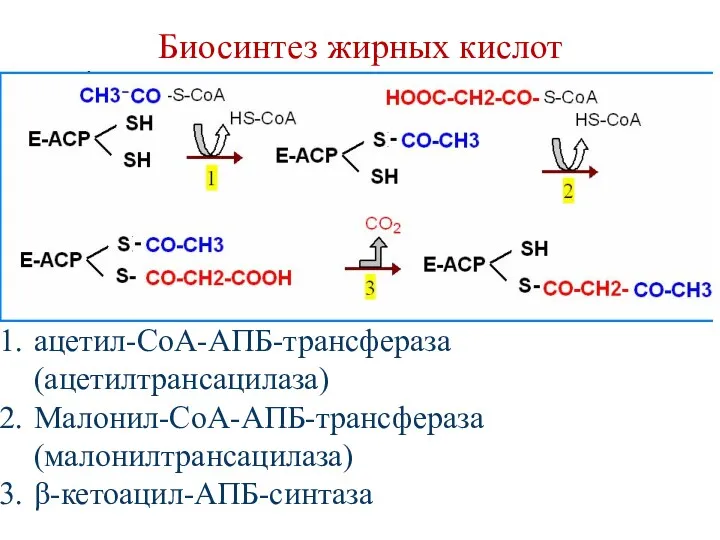
Биосинтез жирных кислот
ацетил-СоА-АПБ-трансфераза (ацетилтрансацилаза)
Малонил-СоА-АПБ-трансфераза (малонилтрансацилаза)
β-кетоацил-АПБ-синтаза
Биосинтез жирных кислот
ацетил-СоА-АПБ-трансфераза (ацетилтрансацилаза)
Малонил-СоА-АПБ-трансфераза (малонилтрансацилаза)
β-кетоацил-АПБ-синтаза
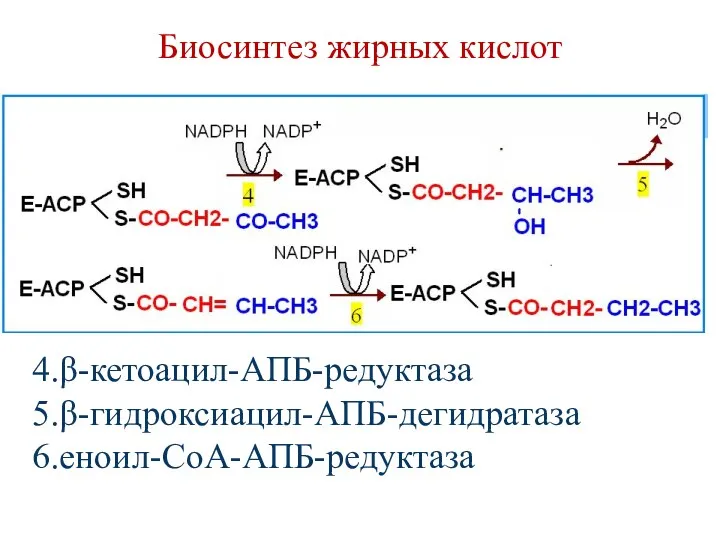
Биосинтез жирных кислот
4.β-кетоацил-АПБ-редуктаза
5.β-гидроксиацил-АПБ-дегидратаза
6.еноил-СоА-АПБ-редуктаза
Биосинтез жирных кислот
4.β-кетоацил-АПБ-редуктаза
5.β-гидроксиацил-АПБ-дегидратаза
6.еноил-СоА-АПБ-редуктаза
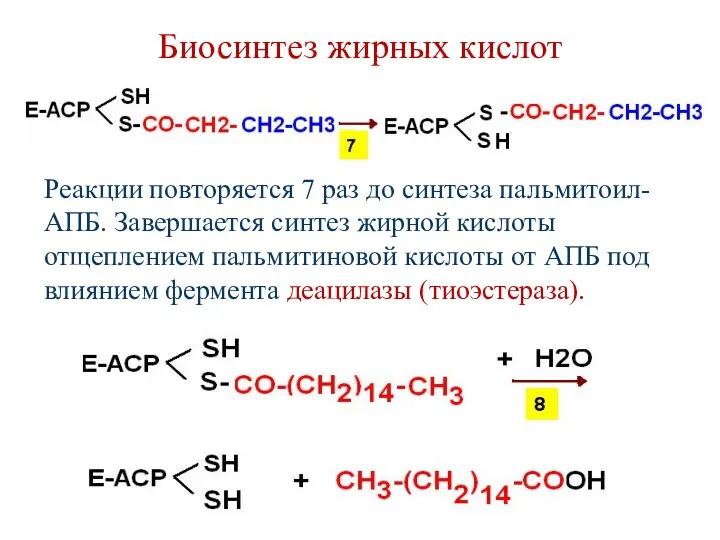
Биосинтез жирных кислот
Реакции повторяется 7 раз до синтеза пальмитоил-АПБ. Завершается синтез
Биосинтез жирных кислот
Реакции повторяется 7 раз до синтеза пальмитоил-АПБ. Завершается синтез

Суммарная реакция
биосинтеза пальмитиновой кислоты
СН3–СО–SСoA + 7НООС–СН2–СО–SСoA + 14НАДФН
Суммарная реакция
биосинтеза пальмитиновой кислоты
СН3–СО–SСoA + 7НООС–СН2–СО–SСoA + 14НАДФН

Регуляция биосинтеза жирных кислот
Основной регуляторный фермент –
ацетил-СоА карбоксилаза
Цитрат
Регуляция биосинтеза жирных кислот
Основной регуляторный фермент –
ацетил-СоА карбоксилаза
Цитрат

Элонгация (удлинение) жирных кислот
Происходит в ЭР под действием энзиматической системы элонгаза.
Элонгация (удлинение) жирных кислот
Происходит в ЭР под действием энзиматической системы элонгаза.
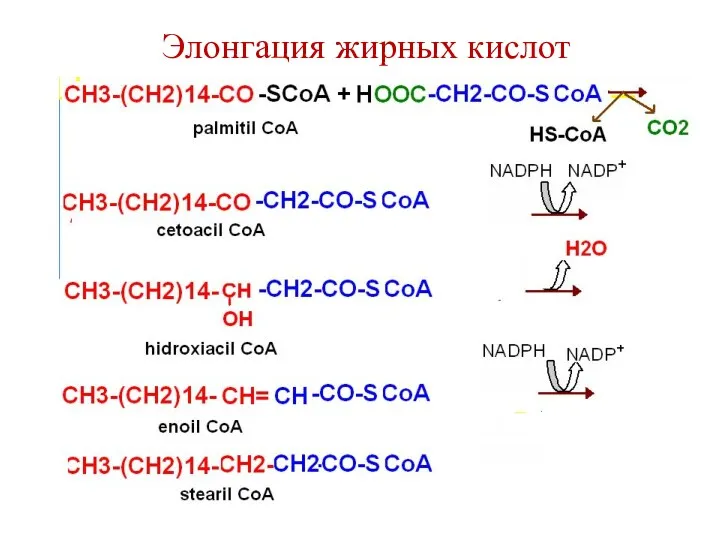
Элонгация жирных кислот
Элонгация жирных кислот

Синтез ненасыщенных жирных кислот
Моноеновые жирные кислоты – пальмитоолеиновая и олеиновая –
Синтез ненасыщенных жирных кислот
Моноеновые жирные кислоты – пальмитоолеиновая и олеиновая –
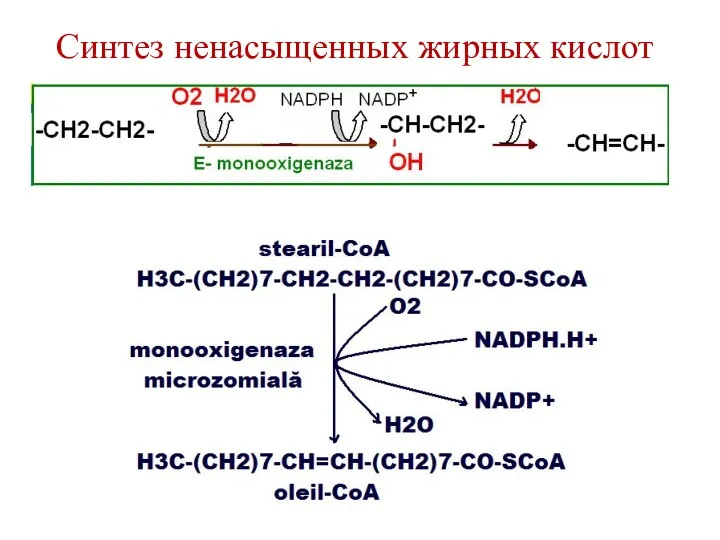
Синтез ненасыщенных жирных кислот
Синтез ненасыщенных жирных кислот

Десатураза млекопитающих может образовать двойные связи только на участке цепи жирной
Десатураза млекопитающих может образовать двойные связи только на участке цепи жирной
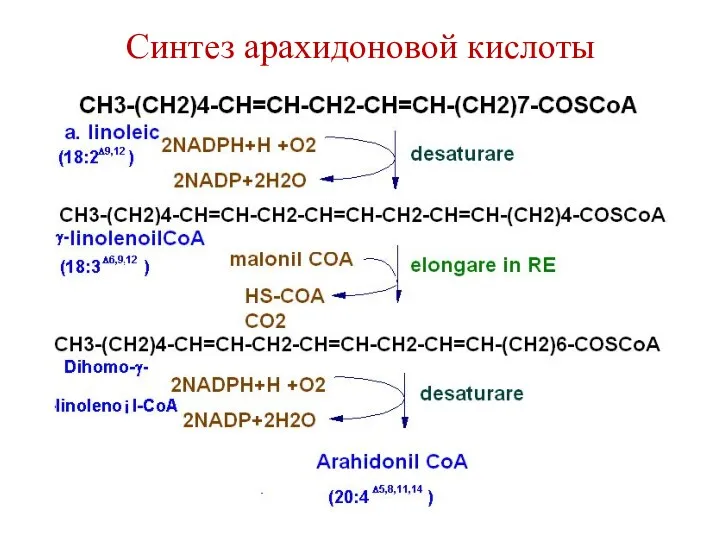
Синтез арахидоновой кислоты
Синтез арахидоновой кислоты

Синтез триацилглицеролов
Локализация процесса – печень, жировая ткань.
Синтез происходит из глицерол-3-фосфата и
Синтез триацилглицеролов
Локализация процесса – печень, жировая ткань.
Синтез происходит из глицерол-3-фосфата и
Синтез триацилглицеролов
1. Печень, почки, стенка кишечника
2. Печень, мышцы, жировая ткань.
Пути
Синтез триацилглицеролов
1. Печень, почки, стенка кишечника
2. Печень, мышцы, жировая ткань.
Пути
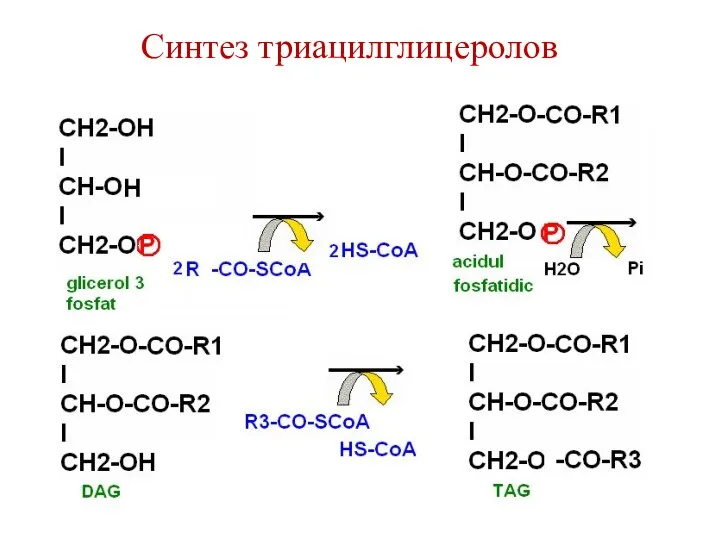
Синтез триацилглицеролов
Синтез триацилглицеролов

Синтез фосфолипидов
интенсивно происходит в печени, стенке кишечника, семенниках, яичниках, молочной железе.
Синтез фосфолипидов
интенсивно происходит в печени, стенке кишечника, семенниках, яичниках, молочной железе.
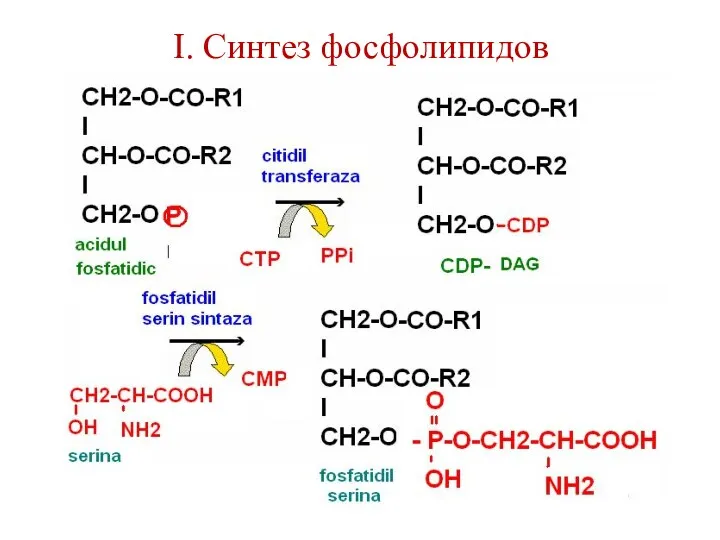
I. Синтез фосфолипидов
I. Синтез фосфолипидов
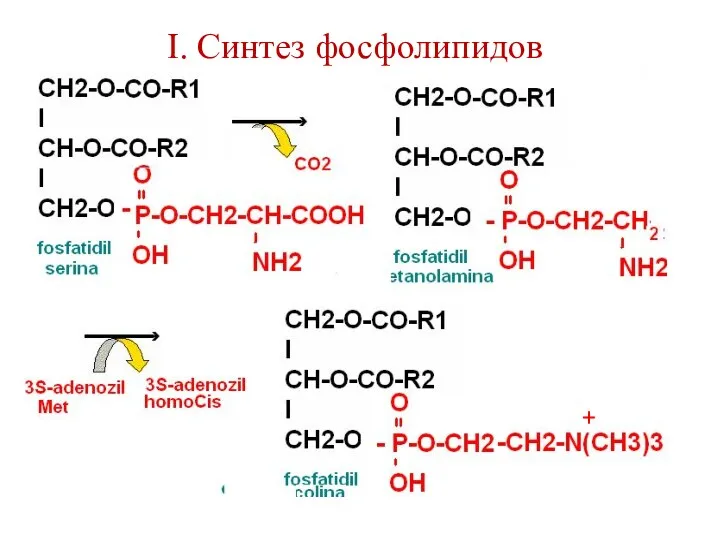
I. Синтез фосфолипидов
+
I. Синтез фосфолипидов
+
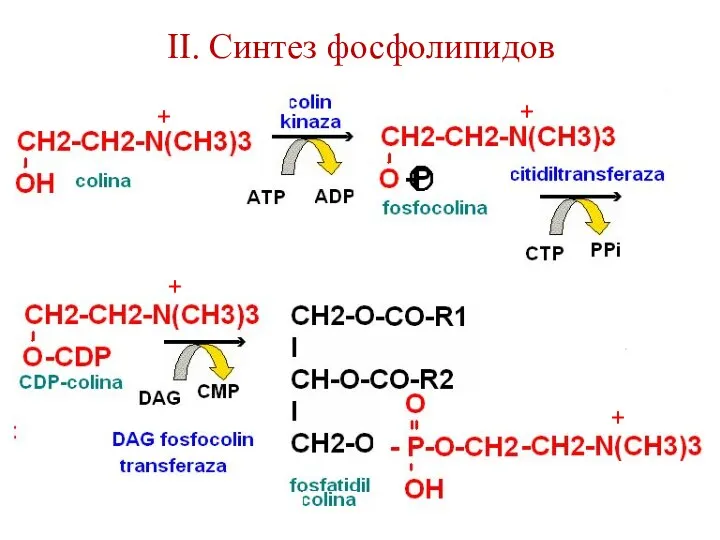
II. Синтез фосфолипидов
+
+
+
+
II. Синтез фосфолипидов
+
+
+
+

Синтез сфингомиелинов
Пальмитоил-СоА + серин →→→сфингозин
Сфингозин + ацил-СоА →церамид
Церамид + СДФ-холин
Синтез сфингомиелинов
Пальмитоил-СоА + серин →→→сфингозин
Сфингозин + ацил-СоА →церамид
Церамид + СДФ-холин
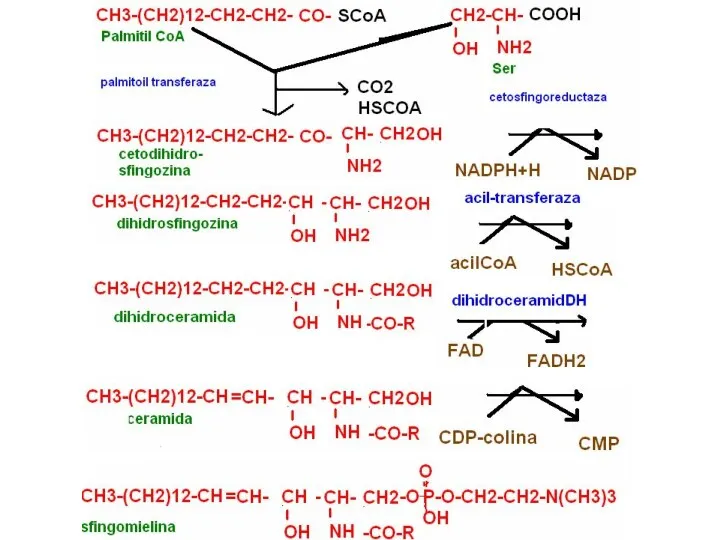
Синтез сфинголипидов
Синтез сфинголипидов

Синтез гликолипидов
Церамид + УДФ-глюкоза → глюкоцереброзид + УДФ
Церамид + УДФ-галактоза
Синтез гликолипидов
Церамид + УДФ-глюкоза → глюкоцереброзид + УДФ
Церамид + УДФ-галактоза

Синтез холестерола
За сутки синтезируется 1г холестерола.
С пищей поступает 0,3-0,5г.
Синтез холестерола
За сутки синтезируется 1г холестерола.
С пищей поступает 0,3-0,5г.
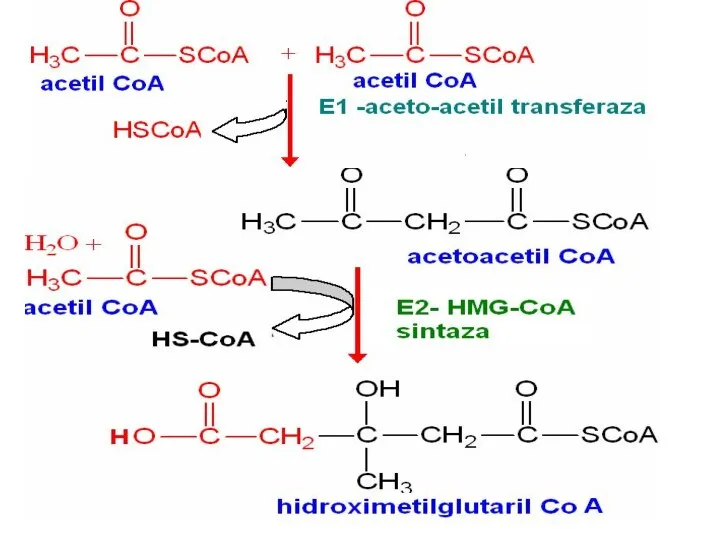
Синтез холестерола
Синтез холестерола
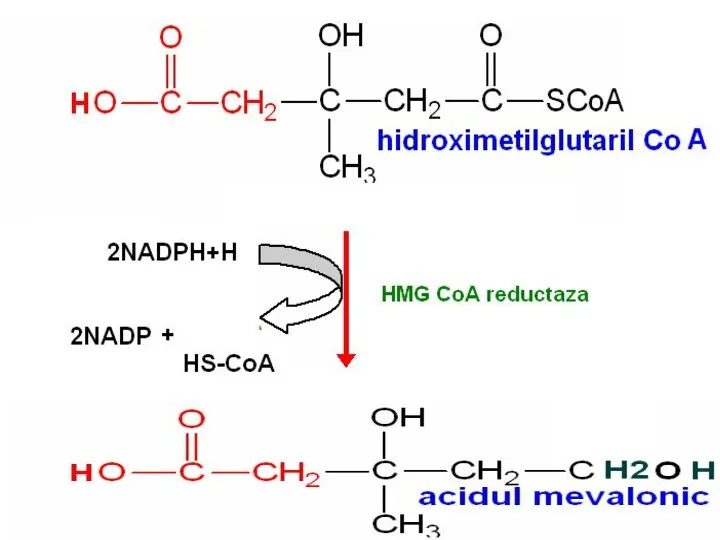
Синтез холестерола
Синтез холестерола

Регуляция биосинтеза холестерола
Регуляторный фермент –
ГМГ-КоА-редуктаза
Ингибиторы – холестерол, мевалоновая кислота,
Регуляция биосинтеза холестерола
Регуляторный фермент –
ГМГ-КоА-редуктаза
Ингибиторы – холестерол, мевалоновая кислота,
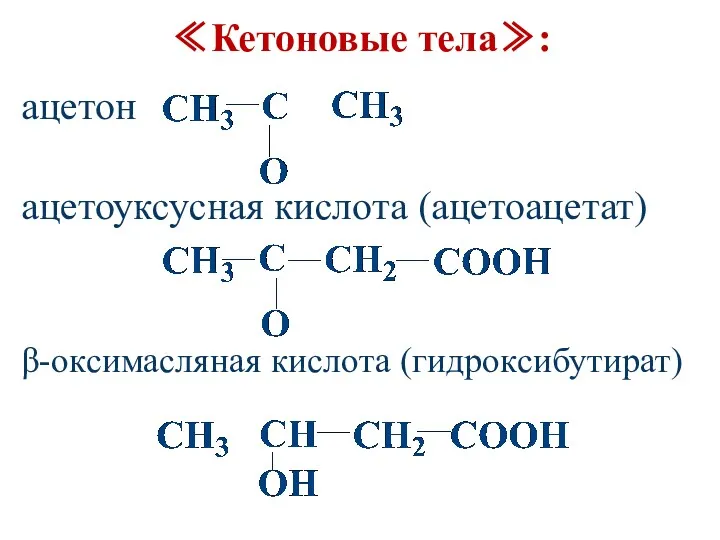
≪Кетоновые тела≫:
ацетон
ацетоуксусная кислота (ацетоацетат)
β-оксимасляная кислота (гидроксибутират)
≪Кетоновые тела≫:
ацетон
ацетоуксусная кислота (ацетоацетат)
β-оксимасляная кислота (гидроксибутират)

Синтез ≪кетоновых тел≫
Происходит только в печени.
Субстратом является ацетил-СоА.
Синтез ≪кетоновых тел≫
Происходит только в печени.
Субстратом является ацетил-СоА.
+
Ацетил-СоА
Ацетил-СоА
Тиолаза
Ацетоацетил-СоА
-HS-CoA
HS-CoA
+H2O
β-гирокси-β-метил-глутарил-СоА
Ацетоацетат
ГМГ-СоА-синтаза
ГМГ-СоА-лиаза
+
Ацетил-СоА
Ацетил-СоА
Тиолаза
Ацетоацетил-СоА
-HS-CoA
HS-CoA
+H2O
β-гирокси-β-метил-глутарил-СоА
Ацетоацетат
ГМГ-СоА-синтаза
ГМГ-СоА-лиаза
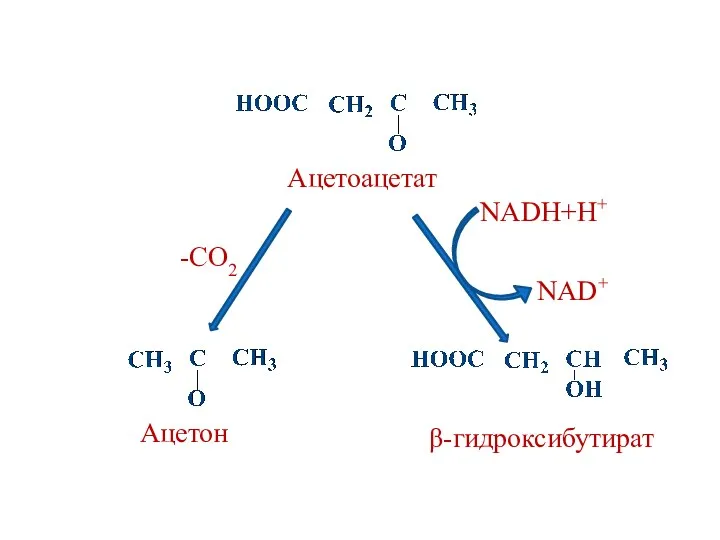
Ацетоацетат
Ацетон
-CO2
NADH+H+
β-гидроксибутират
NAD+
Ацетоацетат
Ацетон
-CO2
NADH+H+
β-гидроксибутират
NAD+

Использование «кетоновых тел» в тканях
используются в качестве ≪топлива≫ сердечной мышцей, корковым
Использование «кетоновых тел» в тканях
используются в качестве ≪топлива≫ сердечной мышцей, корковым
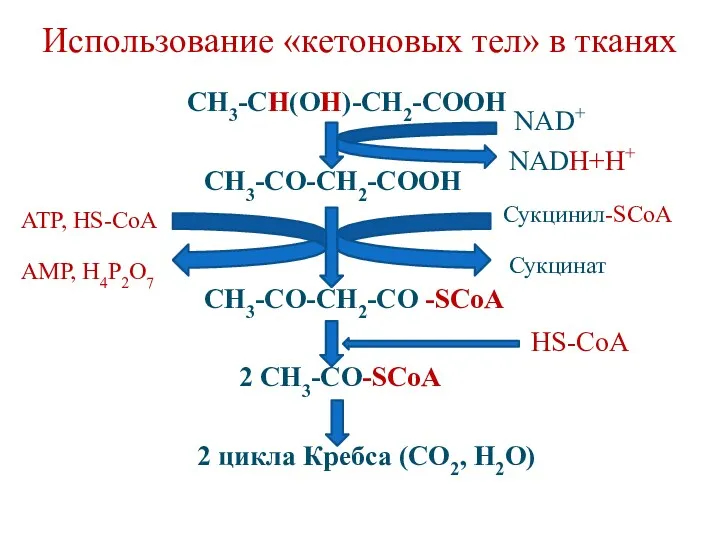
Использование «кетоновых тел» в тканях
СН3-СН(ОН)-СН2-СООН
СН3-СО-СН2-СООН
СН3-СО-СН2-СО -SCoA
2 СН3-СО-SCoA
Использование «кетоновых тел» в тканях
СН3-СН(ОН)-СН2-СООН
СН3-СО-СН2-СООН
СН3-СО-СН2-СО -SCoA
2 СН3-СО-SCoA

Нормальная концентрация «кетоновых тел» (0,03–0,2 ммоль/л).
При патологических состояниях (сахарный диабет, голодание)
Нормальная концентрация «кетоновых тел» (0,03–0,2 ммоль/л).
При патологических состояниях (сахарный диабет, голодание)

Причины кетонемии
Увеличение концентрации ацетил-СоА (усиленное расщепление липидов).
Уменьшение концентрации оксалоацетата (при отсутствии
Причины кетонемии
Увеличение концентрации ацетил-СоА (усиленное расщепление липидов).
Уменьшение концентрации оксалоацетата (при отсутствии
глюкоза
пируват
Ацетил-СоА
оксалоацетат
ТАГ
ЖК
кетоновые тела
цикл
Кребса
Нормальные
условия
Сахарный
диабет
глюкоза
пируват
Ацетил-СоА
оксалоацетат
ТАГ
ЖК
кетоновые тела
цикл
Кребса
Нормальные
условия
Сахарный
диабет

Эйкозаноиды.
Жирорастворимые витамины. Регуляция и патология липидного обмена
Эйкозаноиды.
Жирорастворимые витамины. Регуляция и патология липидного обмена

Эйкозаноиды
Биологически активные вещества, аутокринные и паракринные гормоны. К ним относятся:
простагландины;
простациклины;
тромбоксаны;
лейкотриены.
Эйкозаноиды
Биологически активные вещества, аутокринные и паракринные гормоны. К ним относятся:
простагландины;
простациклины;
тромбоксаны;
лейкотриены.
Фосфолипаза А2
Фосфолипаза А2
Синтез эйкозаноидов
Арахидоновая кислота
Липоксигеназа
Циклоксигеназа
Простагландины
(PGG2)
Простагландины
(PGH2)
Тромбоксаны
(ТХА2 и ТХВ2)
Простациклины
(PGI2)
Лейкотриены
Простагландины
(PGD2, PGE2 и PGF2)
Синтез эйкозаноидов
Арахидоновая кислота
Липоксигеназа
Циклоксигеназа
Простагландины
(PGG2)
Простагландины
(PGH2)
Тромбоксаны
(ТХА2 и ТХВ2)
Простациклины
(PGI2)
Лейкотриены
Простагландины
(PGD2, PGE2 и PGF2)

Механизм действия эйкозаноидов
Являются гормонами местного действия.
Действуют по аутокринному и/или паракринному
Механизм действия эйкозаноидов
Являются гормонами местного действия.
Действуют по аутокринному и/или паракринному

Простагландины (PG)
представляют собой 20-углеродные жирные кислоты, содержащие 5-углеродное кольцо и
Простагландины (PG)
представляют собой 20-углеродные жирные кислоты, содержащие 5-углеродное кольцо и
Простагландины (PG)
Простагландин PGЕ1
Основные биологические эффекты:
Сокращение гладкой мускулатуры.
Простагландины (PG)
Простагландин PGЕ1
Основные биологические эффекты:
Сокращение гладкой мускулатуры.
Тромбоксаны
Тромбоксан TXA2
Образуются в тромбоцитах.
Биологические эффекты:
вызывают сужение кровеносных сосудов и агрегацию тромбоцитов.
Все
Тромбоксаны
Тромбоксан TXA2
Образуются в тромбоцитах.
Биологические эффекты:
вызывают сужение кровеносных сосудов и агрегацию тромбоцитов.
Все
Простациклины
Простациклин PGI1
Образуются в стенках кровеносных сосудов.
Биологические эффекты:
Сильные ингибиторы агрегации тромбоцитов.
Имеют
Простациклины
Простациклин PGI1
Образуются в стенках кровеносных сосудов.
Биологические эффекты:
Сильные ингибиторы агрегации тромбоцитов.
Имеют

Роль эйкозаноидов в тромбообразовании
Тромбоксаны и простациклины являются антагонистами.
Соотношение тромбоксана и простациклина
Роль эйкозаноидов в тромбообразовании
Тромбоксаны и простациклины являются антагонистами.
Соотношение тромбоксана и простациклина

Роль эйкозаноидов в тромбообразовании
При контакте тромбоцитов с поврежденной стенкой сосуда активируется
Роль эйкозаноидов в тромбообразовании
При контакте тромбоцитов с поврежденной стенкой сосуда активируется

!!! При потреблении пищи с преобладанием эйкозапентаеновой кислоты (рыбий жир) в
!!! При потреблении пищи с преобладанием эйкозапентаеновой кислоты (рыбий жир) в
Лейкотриены
Лейкотриен LTA4
Название ≪лейкотриены≫ происходит от: ≪лейкоциты≫ (впервые были обнаружены в лейкоцитах)
Лейкотриены
Лейкотриен LTA4
Название ≪лейкотриены≫ происходит от: ≪лейкоциты≫ (впервые были обнаружены в лейкоцитах)

Роль эйкозаноидов в воспалении
Воспаление – реакция организма на повреждение или инфекцию,
Роль эйкозаноидов в воспалении
Воспаление – реакция организма на повреждение или инфекцию,

Роль эйкозаноидов в воспалении
Многие эйкозаноиды выполняют функцию
медиаторов воспаления и действуют
Роль эйкозаноидов в воспалении
Многие эйкозаноиды выполняют функцию
медиаторов воспаления и действуют

Лекарственные препараты –
ингибиторы синтеза эйкозаноидов
Нестероидные противовоспалительные препараты (аспирин, ибупрофен)
Лекарственные препараты –
ингибиторы синтеза эйкозаноидов
Нестероидные противовоспалительные препараты (аспирин, ибупрофен)
Арахидоновая кислота
Липоксигеназа
Циклоксигеназа
Простагландины
(PGG2)
Простагландины
(PGH2)
Тромбоксаны
(ТХА2 и ТХВ2)
Простациклины
(PGI2)
Лейкотриены
Простагландины
(PGD2, PGE2 и PGF2)
Мембранные фосфолипиды
Фосфолипаза
Арахидоновая кислота
Липоксигеназа
Циклоксигеназа
Простагландины
(PGG2)
Простагландины
(PGH2)
Тромбоксаны
(ТХА2 и ТХВ2)
Простациклины
(PGI2)
Лейкотриены
Простагландины
(PGD2, PGE2 и PGF2)
Мембранные фосфолипиды
Фосфолипаза

Бронхиальная астма
Развитие симптомов бронхиальной астмы (бронхоспазм и экссудация слизи в
Бронхиальная астма
Развитие симптомов бронхиальной астмы (бронхоспазм и экссудация слизи в

Использование эйкозаноидов
в качестве лекарств
Аналоги PGE1 и PGE2 подавляют секрецию соляной
Использование эйкозаноидов
в качестве лекарств
Аналоги PGE1 и PGE2 подавляют секрецию соляной

Жирорастворимые витамины
Витамин А (антиксерофтальмический); ретинол.
Витамин D (антирахитический); кальциферолы.
Витамин Е (антистерильный, витамин
Жирорастворимые витамины
Витамин А (антиксерофтальмический); ретинол.
Витамин D (антирахитический); кальциферолы.
Витамин Е (антистерильный, витамин

Витамин А (ретинол; антиксерофтальмический витамин)
Источники:
Животные продукты – рыбий жир, печень
Витамин А (ретинол; антиксерофтальмический витамин)
Источники:
Животные продукты – рыбий жир, печень
Витамин А
Ретинол – циклический непредельный одноатомный спирт, состоит из шестичленного кольца
Витамин А
Ретинол – циклический непредельный одноатомный спирт, состоит из шестичленного кольца
Каротеноиды (провитамины А)
Растительные продукты – морковь, томаты, перец, салат.
Каротиндиоксигеназа (фермент слизистой
Каротеноиды (провитамины А)
Растительные продукты – морковь, томаты, перец, салат.
Каротиндиоксигеназа (фермент слизистой

Биологические функции витамина А
В организме ретинол превращается в ретиналь и ретиноевую
Биологические функции витамина А
В организме ретинол превращается в ретиналь и ретиноевую
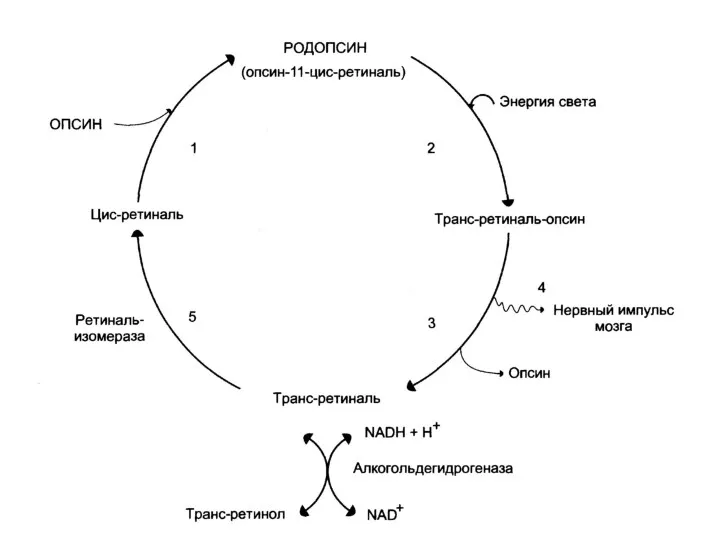

Гиповитаминоз А
Ксерофтальмия, сухость роговой оболочки глаза (от греч. xeros – сухой,
Гиповитаминоз А
Ксерофтальмия, сухость роговой оболочки глаза (от греч. xeros – сухой,

Гиповитаминоз А
Торможение роста, снижение массы тела, общее истощение организма.
Специфические поражения эпителия
Гиповитаминоз А
Торможение роста, снижение массы тела, общее истощение организма.
Специфические поражения эпителия

Гипервитаминоз А
Развивается при употреблении печени белого медведя, тюленя, моржа,
у
Гипервитаминоз А
Развивается при употреблении печени белого медведя, тюленя, моржа,
у
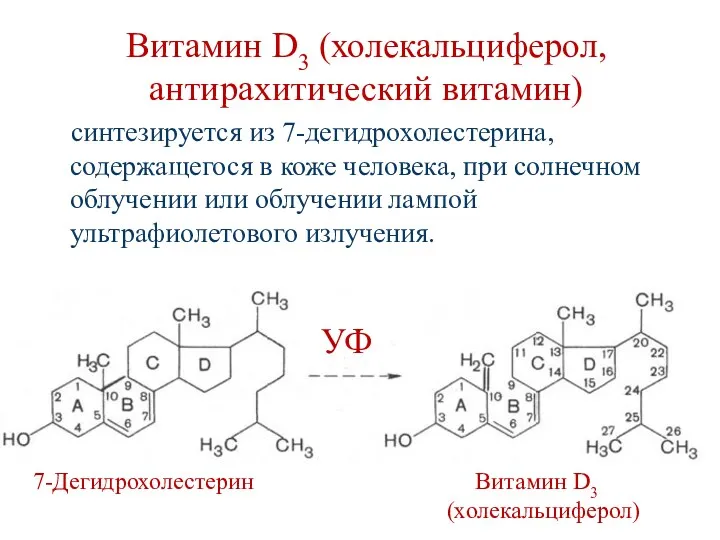
Витамин D3 (холекальциферол, антирахитический витамин)
синтезируется из 7-дегидрохолестерина, содержащегося в коже
Витамин D3 (холекальциферол, антирахитический витамин)
синтезируется из 7-дегидрохолестерина, содержащегося в коже
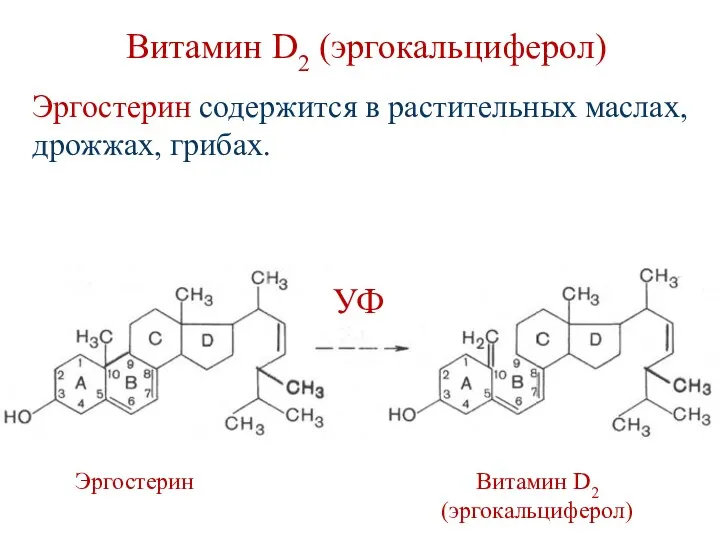
Витамин D2 (эргокальциферол)
Эргостерин Витамин D2
(эргокальциферол)
Эргостерин содержится в растительных
Витамин D2 (эргокальциферол)
Эргостерин Витамин D2
(эргокальциферол)
Эргостерин содержится в растительных

Витамин D (кальциферол, антирахитический витамин)
Распространение в природе:
витамин D3 – продукты
Витамин D (кальциферол, антирахитический витамин)
Распространение в природе:
витамин D3 – продукты
![Биологическая роль В организме из витамина D образуется 1,25-дигидроксихолекальциферол [1,25(OH)2D3]](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/389398/slide-152.jpg)
Биологическая роль
В организме из витамина D образуется 1,25-дигидроксихолекальциферол [1,25(OH)2D3] (кальцитриол) и
Биологическая роль
В организме из витамина D образуется 1,25-дигидроксихолекальциферол [1,25(OH)2D3] (кальцитриол) и

Биологическая роль
Кальцитриол выполняет гормональную роль в регуляции обмена кальция и минерализации
Биологическая роль
Кальцитриол выполняет гормональную роль в регуляции обмена кальция и минерализации

Гиповитаминоз D (рахит)
Основные симптомы рахита у детей:
остеомаляция – размягчение
Гиповитаминоз D (рахит)
Основные симптомы рахита у детей:
остеомаляция – размягчение

Гипервитаминоз D
При ≪ударной≫ терапии рахита и некоторых дерматозов (волчанка).
Гипервитаминоз D
При ≪ударной≫ терапии рахита и некоторых дерматозов (волчанка).
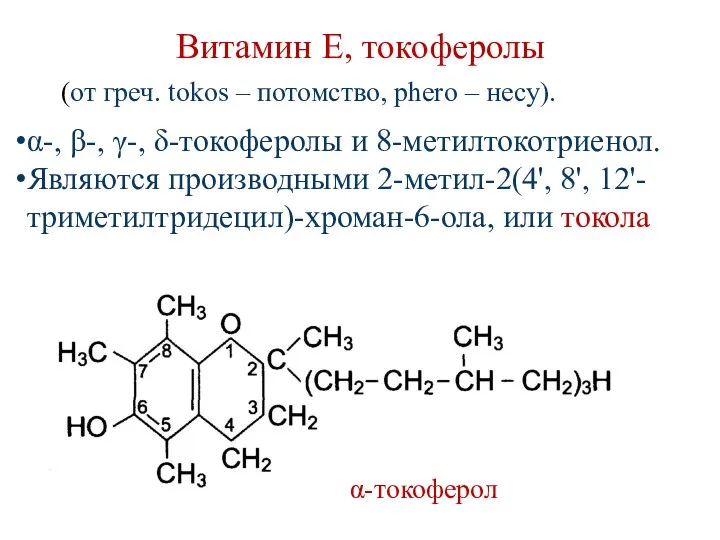
Витамин Е, токоферолы
α-токоферол
(от греч. tokos – потомство, phero – несу).
α-, β-,
Витамин Е, токоферолы
α-токоферол
(от греч. tokos – потомство, phero – несу).
α-, β-,

Витамин Е
Распространение в природе:
Растительные масла (подсолнечное, хлопковое, соевое, кукурузное).
Салат,
Витамин Е
Распространение в природе:
Растительные масла (подсолнечное, хлопковое, соевое, кукурузное).
Салат,

Биологическая роль витамина Е
Активный природный антиоксидант: токоферолы разрушают наиболее реактивные формы
Биологическая роль витамина Е
Активный природный антиоксидант: токоферолы разрушают наиболее реактивные формы

Гиповитаминоз Е
Нарушение эмбриогенеза и дегенеративные изменения репродуктивных органов, что приводит к
Гиповитаминоз Е
Нарушение эмбриогенеза и дегенеративные изменения репродуктивных органов, что приводит к

Витамин К (антигеморрагический); нафтохиноны
Производные хинонов с боковыми изопреноидными цепями.
Витамин K1
Витамин К (антигеморрагический); нафтохиноны
Производные хинонов с боковыми изопреноидными цепями.
Витамин K1
Витамин К (антигеморрагический); нафтохиноны
Витамин К (антигеморрагический); нафтохиноны

Витамин К
Распространение в природе:
Растительные продукты – капуста, шпинат, тыква, зеленые
Витамин К
Распространение в природе:
Растительные продукты – капуста, шпинат, тыква, зеленые
Витамин К
Витамин К3 (менадион) Викасол
Витамин К3, синтетический аналог витамина К,
Витамин К
Витамин К3 (менадион) Викасол
Витамин К3, синтетический аналог витамина К,
Биологическая роль витамина К
Витамин К является кофактором карбоксилазы глутаминовой кислоты, образуется
Биологическая роль витамина К
Витамин К является кофактором карбоксилазы глутаминовой кислоты, образуется
Биологическая роль витамина К
Биологическая роль витамина К
Антивитамины К
дикумарол салициловая варфарин
кислота
Конкурентные ингибиторы тромбообразования.
Являются непрямыми антикоагулянтами.
Антивитамины К
дикумарол салициловая варфарин
кислота
Конкурентные ингибиторы тромбообразования.
Являются непрямыми антикоагулянтами.

Гиповитаминоз К
Встречается редко (пища богата витамином К и он синтезируется кишечной
Гиповитаминоз К
Встречается редко (пища богата витамином К и он синтезируется кишечной

Регуляция липидного обмена
Адреналин, норадреналин и глюкагон увеличивают скорость липолиза в жировой
Регуляция липидного обмена
Адреналин, норадреналин и глюкагон увеличивают скорость липолиза в жировой
Тканевой липолиз
АТР
цАМР
ПК на
ПК а
ТГЛ на
(Е-ОН)
ТГЛ а
(Е-Р)
АТР
АДР
ТАГ
МАГ
ДАГ
глицерол
ЖК
ЖК
ЖК
гормон
(адреналин, глюкагон)
- рецептор
- аденилатциклаза
ДГЛ
МГЛ
Тканевой липолиз
АТР
цАМР
ПК на
ПК а
ТГЛ на
(Е-ОН)
ТГЛ а
(Е-Р)
АТР
АДР
ТАГ
МАГ
ДАГ
глицерол
ЖК
ЖК
ЖК
гормон
(адреналин, глюкагон)
- рецептор
- аденилатциклаза
ДГЛ
МГЛ

Регуляция липидного обмена
Инсулин оказывает противоположное адреналину и глюкагону действие на липолиз.
Регуляция липидного обмена
Инсулин оказывает противоположное адреналину и глюкагону действие на липолиз.
Тканевой липолиз
АТР
цАМР
ПК на
ПК а
ТГЛ на
(Е-ОН)
ТГЛ а
(Е-Р)
АТР
АДР
ТАГ
МАГ
ДАГ
глицерол
ЖК
ЖК
ЖК
ДГЛ
МГЛ
АМР
PDE
H2O
Рi
фосфопротеинфосфатаза
Тканевой липолиз
АТР
цАМР
ПК на
ПК а
ТГЛ на
(Е-ОН)
ТГЛ а
(Е-Р)
АТР
АДР
ТАГ
МАГ
ДАГ
глицерол
ЖК
ЖК
ЖК
ДГЛ
МГЛ
АМР
PDE
H2O
Рi
фосфопротеинфосфатаза

Регуляция липидного обмена
Инсулин стимулирует липогенез:
Активирует липопротеинлипазу обеспечивает липогенез жирными кислотами.
Стимулирует
Регуляция липидного обмена
Инсулин стимулирует липогенез:
Активирует липопротеинлипазу обеспечивает липогенез жирными кислотами.
Стимулирует

Регуляция липидного обмена
Глюкокортикоиды:
Стимулируют липолиз в конечностях и липогенез в других
Регуляция липидного обмена
Глюкокортикоиды:
Стимулируют липолиз в конечностях и липогенез в других

Регуляция липидного обмена
Соматотропин:
Усиливает липолиз в жировой ткани (за счет усиления
Регуляция липидного обмена
Соматотропин:
Усиливает липолиз в жировой ткани (за счет усиления

Регуляция липидного обмена
Тиреоидные гормоны:
Повышают чувствительность β-адренорецепторов к действию адреналина и
Регуляция липидного обмена
Тиреоидные гормоны:
Повышают чувствительность β-адренорецепторов к действию адреналина и

Концентрация холестерина
Нормальный уровень –
менее 5,2 ммоль/л
Пограничный уровень –
Концентрация холестерина
Нормальный уровень –
менее 5,2 ммоль/л
Пограничный уровень –

ХОЛЕСТЕРИН ЛПНП
Формула Friedwald :
ХС ЛПНП, ммоль/л=Общий ХС – ХС ЛПВП
ХОЛЕСТЕРИН ЛПНП
Формула Friedwald :
ХС ЛПНП, ммоль/л=Общий ХС – ХС ЛПВП

ХОЛЕСТЕРИН ЛПВП
Нормальная концентрация:
Мужчины –
0,9-1,4 mmol/L;
Женщины –
1,2-1,7 mmol/L.
ХОЛЕСТЕРИН ЛПВП
Нормальная концентрация:
Мужчины –
0,9-1,4 mmol/L;
Женщины –
1,2-1,7 mmol/L.
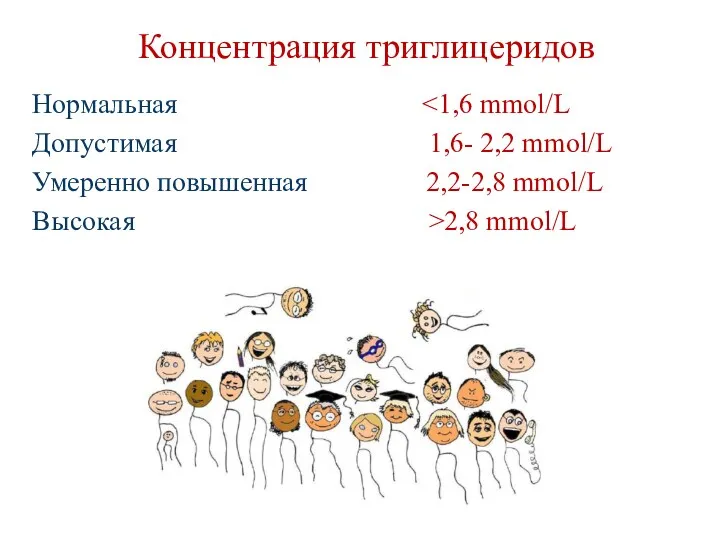
Концентрация триглицеридов
Нормальная <1,6 mmol/L
Допустимая 1,6- 2,2 mmol/L
Умеренно повышенная 2,2-2,8 mmol/L
Высокая >2,8
Концентрация триглицеридов
Нормальная <1,6 mmol/L
Допустимая 1,6- 2,2 mmol/L
Умеренно повышенная 2,2-2,8 mmol/L
Высокая >2,8

Дислипопротеинемии
Нарушения образования, транспорта и
утилизации ЛП, сопровождаемые изменением плазменного уровня
Дислипопротеинемии
Нарушения образования, транспорта и
утилизации ЛП, сопровождаемые изменением плазменного уровня

Первичные гиперлипопортеидемии
ГЛП I типа. Семейная гиперхиломикронемия.
ГЛП II а типа. Семейная гипер-бета-липопротеинемия.
Первичные гиперлипопортеидемии
ГЛП I типа. Семейная гиперхиломикронемия.
ГЛП II а типа. Семейная гипер-бета-липопротеинемия.

ГЛП I типа.
Семейная гиперхиломикронемия
Генетический дефект липопротеидлипазы или апобелка СII.
Нарушение катаболизма
ГЛП I типа.
Семейная гиперхиломикронемия
Генетический дефект липопротеидлипазы или апобелка СII.
Нарушение катаболизма

ГЛП II типа.
Семейная гиперхолестеролемия.
Мутациея гена ЛПНП-рецептора или апо В100.
Частота
ГЛП II типа.
Семейная гиперхолестеролемия.
Мутациея гена ЛПНП-рецептора или апо В100.
Частота

ГЛП II а типа.
Семейная гиперхолестеролемия.
Гомозиготные формы (1 на 1 000
ГЛП II а типа.
Семейная гиперхолестеролемия.
Гомозиготные формы (1 на 1 000
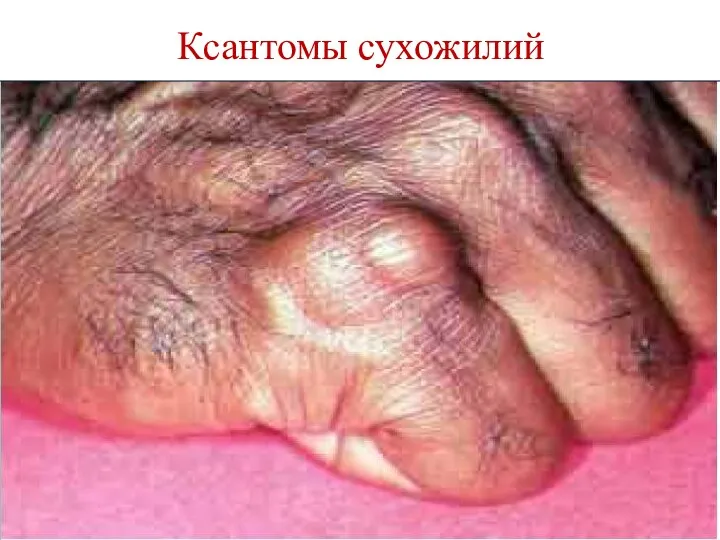
Ксантомы сухожилий
Ксантомы сухожилий
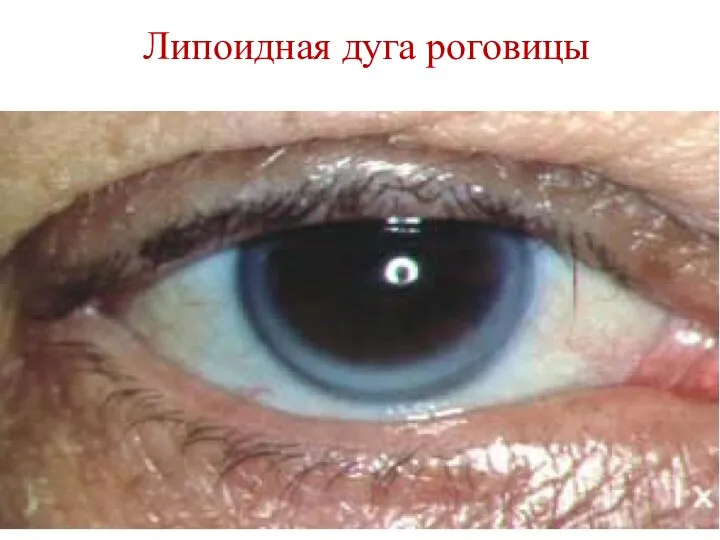
Липоидная дуга роговицы
Липоидная дуга роговицы

Первичные гиполипопортеидемии
Абеталипопротеинемия
Гипобеталипопротеинемия
Анальфалипопротеинемия
Семейная наследственная недостаточность ЛХАТ
Первичные гиполипопортеидемии
Абеталипопротеинемия
Гипобеталипопротеинемия
Анальфалипопротеинемия
Семейная наследственная недостаточность ЛХАТ

Aн-альфа-липопротеинемия
(болезнь Тангера)
Мутация гена транспортного белка ABCA-1 (ATP-binding cassette transporter) –
Aн-альфа-липопротеинемия
(болезнь Тангера)
Мутация гена транспортного белка ABCA-1 (ATP-binding cassette transporter) –

Дислипидемия при сахарном диабете
“липидная триада”:
• гипертриглицеридемия;
• низкий
Дислипидемия при сахарном диабете
“липидная триада”:
• гипертриглицеридемия;
• низкий

Атеросклероз
Хроническое прогрессирующее заболевание крупных и средних эластических и мышечно-эластических артерий, характеризующееся
Атеросклероз
Хроническое прогрессирующее заболевание крупных и средних эластических и мышечно-эластических артерий, характеризующееся

Главные факторы
риска развития атеросклероза
Дислипопротеинемии
(как наследственные, так и
Главные факторы
риска развития атеросклероза
Дислипопротеинемии
(как наследственные, так и

«Мягкие» факторы
риска развития атеросклероза
Ожирение (особенно абдоминального типа).
Гиподинамия.
Хронический
«Мягкие» факторы
риска развития атеросклероза
Ожирение (особенно абдоминального типа).
Гиподинамия.
Хронический

Атерогенная дислипидемия
Увеличение концентрации общего холестерина.
Повышение содержания ХС ЛПНП (особенно мелких частиц
Атерогенная дислипидемия
Увеличение концентрации общего холестерина.
Повышение содержания ХС ЛПНП (особенно мелких частиц

Роль ЛПНП в атерогенезе
Перекисно-модифицированные ЛПНП распознаются и захватываются скэвенджер-рецепторами макрофагов (в
Роль ЛПНП в атерогенезе
Перекисно-модифицированные ЛПНП распознаются и захватываются скэвенджер-рецепторами макрофагов (в
Oжирение
ИМТ (индекс массы тела)
ИМТ = вес(kg)/квадрат роста(m2)
Oжирение
ИМТ (индекс массы тела)
ИМТ = вес(kg)/квадрат роста(m2)

Факторы риска ожирения
Генетические дефекты (лептин, рецептор лептина, β3-адренергический рецептор,
IRS-1).
Алиментарные
Факторы риска ожирения
Генетические дефекты (лептин, рецептор лептина, β3-адренергический рецептор,
IRS-1).
Алиментарные
 Общие правила оказания первой доврачебной помощи. Алгоритм оказания первой помощи. Юридические и моральные аспекты
Общие правила оказания первой доврачебной помощи. Алгоритм оказания первой помощи. Юридические и моральные аспекты Клинические формы вторичного туберкулеза
Клинические формы вторичного туберкулеза Рентгеноконтрастные исследования и препараты
Рентгеноконтрастные исследования и препараты Анатомо-физиологические особенности эндокринной системы у детей
Анатомо-физиологические особенности эндокринной системы у детей Мочекаменная болезнь. Гидронефроз
Мочекаменная болезнь. Гидронефроз Вич и Спид
Вич и Спид Інфузійна терапія
Інфузійна терапія Градация доказательств и уровни рекомендаций
Градация доказательств и уровни рекомендаций Особенности сестринского ухода за инфекционными больными. Сестринский процесс. Сестринский диагноз
Особенности сестринского ухода за инфекционными больными. Сестринский процесс. Сестринский диагноз Анализ затрат на лекарственные средства с помощью ABC/VEV методологии
Анализ затрат на лекарственные средства с помощью ABC/VEV методологии Моногибридті будандастыру. Гибридологиялық зерттеу әдісі
Моногибридті будандастыру. Гибридологиялық зерттеу әдісі Опухоли. Онкология
Опухоли. Онкология Анонимные Наркоманы г. Йошкар-Ола
Анонимные Наркоманы г. Йошкар-Ола Энтеробиоз: определение
Энтеробиоз: определение Ожирение. Степени ожирения
Ожирение. Степени ожирения Генетика человека. Генные болезни
Генетика человека. Генные болезни Острые воспалительные заболевания матки и придатков как причина развития клиники острого живота в гинекологии
Острые воспалительные заболевания матки и придатков как причина развития клиники острого живота в гинекологии Лекарственная токсикология
Лекарственная токсикология Орталық және шеткі жүйке жүйесінің клиникалық физиологиясы бен биохимиясы
Орталық және шеткі жүйке жүйесінің клиникалық физиологиясы бен биохимиясы История развития психогенетики в мировой науке
История развития психогенетики в мировой науке Ас қорыту жүйесіне жалпы шолу
Ас қорыту жүйесіне жалпы шолу Микробиологическая диагностика брюшного тифа, паратифов и других сальмонеллезных инфекций. Пищевые отравления и их диагностика
Микробиологическая диагностика брюшного тифа, паратифов и других сальмонеллезных инфекций. Пищевые отравления и их диагностика Лабораторная диагностика заболеваний, вызываемых извитыми формами бактерий. Спирохетозы (сифилис, лептоспироз, возвратные тифы)
Лабораторная диагностика заболеваний, вызываемых извитыми формами бактерий. Спирохетозы (сифилис, лептоспироз, возвратные тифы) Жарақаттар
Жарақаттар Диагностическая информативность онкомаркеров в гинекологии
Диагностическая информативность онкомаркеров в гинекологии Специфическая (антидотная) фармакотерапия острых отравлений
Специфическая (антидотная) фармакотерапия острых отравлений Эректильная дисфункция (ЭД)
Эректильная дисфункция (ЭД) Лекарственные препараты по химии
Лекарственные препараты по химии