- Нейрохимия опиатной наркомании

Содержание
- 2. Патогенез В патогенезе опиатной наркомании значительную роль играют нарушения взаимодействия нейромедиаторных систем . Это касается практически
- 3. Сдвиги в системах нейропередачи имеют непосредственное отношение к формированию феномена пристрастия, абстинентного синдрома, толерантности. В практическом
- 4. Считается, что аддиктивный потенциал наркотических средств реализуется не только на уровне синаптической передачи, но включает также
- 5. Нарушения взаимодействия нейромедиаторных систем могут считаться начальным звеном патогенеза опиатной наркомании. Они же являются мишенью фармакотерапии
- 6. Нейрохимические аспекты опиатной наркомании функционирование опиоидных нейромедиаторных систем, нейрохимические основы толерантности и зависимости, состояние систем вторичных
- 7. Опиоидные рецепторы Опиоиды (эндорфины, энкефалины и динорфины) относятся к числу пептидных нейротрансмиттеров. Важным элементом опиоидной нейромедиаторной
- 8. Первоначальное обозначение классов ОР мю, каппа и дельта в основном определялось названием определенного лиганда. Так, мю-рецепторы
- 9. Классификация опиоидных рецепторов, рекомендуемая Международным Союзом фармакологов (IUPHAR) Примечания: * - номер класса отражает динамику исследований
- 10. Опиоидные рецепторы относятся к семейству метаботропных, т. е. передача информации внутрь нейрона после связывания с агонистом
- 11. Схема опиоидного рецептора Примечания: темными кружками обозначены аминокислоты, подвергающиеся фосфорилированию посредством цАМФ-зависимых протеинкиназ и протеинкиназы С;
- 12. Полипептидная цепь опиоидных рецепторов семь раз пронизывает нейрональную мембрану. Соответственно, трансмембранные участки обозначают ТМ-1 - ТМ-7.
- 13. Однако для каждого рецептора участки рецептирования лигандов различны: в дельта-рецепторах это третья экстрацеллюлярная петля, в мю-рецепторах
- 14. C-терминаль и интрацеллюлярные петли имеют несколько участков для фосфорилирования с помощью протеинкиназ, регулируемых циклическим аденозинмонофосфатом (цАМФ)
- 15. Схема взаимодействия G-белка с опиоидным рецептором (цикл G-белка) Наиболее изученными путями трансдукции интрацеллюлярного сигнала с участием
- 16. G-белок - гетеромерный белок, ассоциированный с мембраной. Он включает альфа-, бета- и гамма-субъединицы. В покое все
- 17. После рецептирования агониста облегчается связывание опиоидного рецептора с G-белком. Далее ГДФ заменяется на гуанозинтрифосфат (ГТФ) в
- 18. альфа-Субъединица, обладающая внутренней ГТФ-азной активностью, гидролизует ГТФ до ГДФ после взаимодействия с эффектором. При этом каталитическая
- 19. Гуаниннуклеотидсвязывающие белки отличаются гетерогенностью, что, в свою очередь, определяется множественностью изоформ входящих в их состав субъединиц.
- 20. Вторым по значимости путем передачи внутриклеточного сигнала при активации опиоидных рецепторов следует считать фосфатидилинозитидный или инозитолфосфатный
- 21. Фосфоинозитиды являются важнейшими компонентами нейрональной мембраны. Стимуляция опиоидных рецепторов приводит к активации фосфолипазы С через гуаниннуклеотидсвязывающий
- 22. Оба вещества считаются вторичными мессенджерами. Диацилглицерол является мощным эндогенным активатором протеинкиназы С. Этот фермент, также как
- 23. Краткая характеристика опиоидных рецепторов дельта-рецепторы Рецепторы типа ОР1 (дельта-рецепторы). Доказано существование минимум двух подтипов ОР1: дельта1-
- 24. каппа-рецепторы Рецепторы типа ОР2 (каппа-рецепторы). Существует не менее трех подтипов каппа-рецепторов: каппа1-, каппа2- и каппа3-рецепторы. Наиболее
- 25. мю-опиоидные рецепторы ОР3-рецепторы - наиболее изученный тип. Как и в случае каппа-рецепторов, подразделение на 2 подтипа
- 26. Упрощенная схема фосфатидилинозитидного цикла
- 27. пептидергическая нейромедиаторная система В пептидергической нейромедиаторной системы, передача осуществляется с участием нейропептида ноцицептина или орфанина FQ.
- 28. Эндогенный лиганд ORL-1-рецепторов ноцицептин/орфанин FG состоит из 17 аминокислотных остатков, что соответствует длине лиганда каппа-рецепторов динорфина
- 29. Передача сигнала рецепторами ORL-1 осуществляется с участием сопряженных G-белков посредством модуляции активности аденилатциклазы, тока К+ внутрь
- 30. Как известно, одним из основных компонентов формирования пристрастия является активация церебральной системы вознаграждения (reward system). Центральное
- 31. Схема взаимодействия опиоидных и дофаминергических систем вентральной области покрышки и прилежащего ядра В регуляции функциональной активности
- 32. Системы вторичных мессенджеров при опиатной наркомании
- 33. Нейрохимические проблемы толерантности и абстинентного синдрома Модуляция систем вторичных передатчиков при хронической опиатной интоксикации имеет прямое
- 34. Негативные отношения между опиатами/опиоидами и аденилатциклазной системой первоначально показаны на клеточных культурах. Как оказалось, эти процессы
- 35. Наркотизация крыс морфином сопровождалась достоверным повышением активности аденилатциклазы и цАМФ-зависимой протеинкиназы в прилежащем ядре, голубом пятне,
- 36. Это подтверждается сходными изменениями аденилатциклазной системы при наркотизации животных другим аддиктивным веществом - кокаином. Препараты, не
- 37. Одной из причин дезинтеграции аденилатциклазной системы при развитии толерантности может быть нарушение взаимодействия опиоидных рецепторов с
- 38. Модификация активности определенных G-белков оказывает влияние на функциональное состояние систем вознаграждения. Например, локальное введение коклюшного токсина
- 39. Как известно, система циклического аденозинмонофосфата регулирует биохимические процессы в клетке посредством фосфорилирования белков-мишеней [46]. Можно предположить,
- 40. Дезинтеграция многочисленных биохимических процессов может считаться основой формирования пристрастия, толерантности и абстинентного синдром. Если к этому
- 41. Наиболее длительным нарушением считается дезадаптационная экспрессия определенных участков генома в ответ на поступление в организм агента,
- 42. Традиционно в патогенезе опиатной наркомании важное место отводили изменениям опиоидных рецепторов (десенсистизация, down-regulation), дезинтеграции связей "опиоидные
- 43. Есть сведения о вовлечении в механизмы толерантности к опиатами каскада "N-метил-D-аспартатные рецепторы - оксид азота". Участие
- 44. Одной из структур головного мозга, вовлеченных в развитие толерантности и физической зависимости, считается голубое пятно (locus
- 45. Менее определенно можно говорить о роли NMDA-рецепторов в модуляции систем вознаграждения при опиатной наркомании. Так, в
- 46. Изменения генома при опиатной наркомании Представления о мутагенной активности опиатов начали формироваться в 70-х годах 20-го
- 47. Не менее важным элементом патогенеза опиатной наркомании является способность аддиктивных веществ инициировать экспрессию определенных участков генома.
- 48. Работы последних лет свидетельствуют, что сдвиги экспрессии мРНК для многих белков зависят от ряда факторов: длительности
- 49. Не ясно, носят ли изменения экспрессии генома транзиторный характер и всегда ли они коррелируют с нарушениями
- 50. Если исходить из классических представлений о структурно-метаболических комплексах клетки, можно утверждать, что при опиатной наркомании имеет
- 51. Изменения экспрессии гена, кодирующего синтез ингибитора связывания (DBI, эндогенный лиганд бензодиазепиновых рецепторов), в определенной степени объясняют
- 52. Сложнее экстраполировать данные о сдвигах содержания мРНК в плане формирования феномена пристрастия. Основная проблема - временные
- 53. Нейрохимические исследования, посвященные патогенезу опиатной наркомании, являются бурно прогрессирующей областью современной медицины. Накапливающиеся научные факты расширяют
- 55. Скачать презентацию

Патогенез
В патогенезе опиатной наркомании значительную роль играют нарушения взаимодействия нейромедиаторных
Патогенез
В патогенезе опиатной наркомании значительную роль играют нарушения взаимодействия нейромедиаторных

Сдвиги в системах нейропередачи имеют непосредственное отношение к формированию феномена
Сдвиги в системах нейропередачи имеют непосредственное отношение к формированию феномена

Считается, что аддиктивный потенциал наркотических средств реализуется не только на
Считается, что аддиктивный потенциал наркотических средств реализуется не только на

Нарушения взаимодействия нейромедиаторных систем могут считаться начальным звеном патогенеза опиатной
Нарушения взаимодействия нейромедиаторных систем могут считаться начальным звеном патогенеза опиатной

Нейрохимические аспекты опиатной наркомании
функционирование опиоидных нейромедиаторных систем,
нейрохимические основы толерантности
Нейрохимические аспекты опиатной наркомании
функционирование опиоидных нейромедиаторных систем,
нейрохимические основы толерантности

Опиоидные рецепторы
Опиоиды (эндорфины, энкефалины и динорфины) относятся к числу
Опиоидные рецепторы
Опиоиды (эндорфины, энкефалины и динорфины) относятся к числу

Первоначальное обозначение классов ОР мю, каппа и дельта в основном
Первоначальное обозначение классов ОР мю, каппа и дельта в основном

Классификация опиоидных рецепторов, рекомендуемая Международным Союзом фармакологов (IUPHAR)
Примечания: * -
Классификация опиоидных рецепторов, рекомендуемая Международным Союзом фармакологов (IUPHAR)
Примечания: * -

Опиоидные рецепторы относятся к семейству метаботропных, т. е. передача информации
Опиоидные рецепторы относятся к семейству метаботропных, т. е. передача информации

Схема опиоидного рецептора
Примечания: темными кружками обозначены аминокислоты, подвергающиеся фосфорилированию посредством
Схема опиоидного рецептора
Примечания: темными кружками обозначены аминокислоты, подвергающиеся фосфорилированию посредством

Полипептидная цепь опиоидных рецепторов семь раз пронизывает нейрональную мембрану. Соответственно,
Полипептидная цепь опиоидных рецепторов семь раз пронизывает нейрональную мембрану. Соответственно,

Однако для каждого рецептора участки рецептирования лигандов различны: в дельта-рецепторах
Однако для каждого рецептора участки рецептирования лигандов различны: в дельта-рецепторах

C-терминаль и интрацеллюлярные петли имеют несколько участков для фосфорилирования с
C-терминаль и интрацеллюлярные петли имеют несколько участков для фосфорилирования с

Схема взаимодействия G-белка с опиоидным рецептором (цикл G-белка)
Наиболее изученными путями трансдукции
Схема взаимодействия G-белка с опиоидным рецептором (цикл G-белка)
Наиболее изученными путями трансдукции

G-белок - гетеромерный белок, ассоциированный с мембраной. Он включает альфа-,
G-белок - гетеромерный белок, ассоциированный с мембраной. Он включает альфа-,

После рецептирования агониста облегчается связывание опиоидного рецептора с G-белком. Далее
После рецептирования агониста облегчается связывание опиоидного рецептора с G-белком. Далее

альфа-Субъединица, обладающая внутренней ГТФ-азной активностью, гидролизует ГТФ до ГДФ после
альфа-Субъединица, обладающая внутренней ГТФ-азной активностью, гидролизует ГТФ до ГДФ после

Гуаниннуклеотидсвязывающие белки отличаются гетерогенностью, что, в свою очередь, определяется множественностью
Гуаниннуклеотидсвязывающие белки отличаются гетерогенностью, что, в свою очередь, определяется множественностью

Вторым по значимости путем передачи внутриклеточного сигнала при активации опиоидных
Вторым по значимости путем передачи внутриклеточного сигнала при активации опиоидных

Фосфоинозитиды являются важнейшими компонентами нейрональной мембраны. Стимуляция опиоидных рецепторов приводит
Фосфоинозитиды являются важнейшими компонентами нейрональной мембраны. Стимуляция опиоидных рецепторов приводит

Оба вещества считаются вторичными мессенджерами. Диацилглицерол является мощным эндогенным активатором
Оба вещества считаются вторичными мессенджерами. Диацилглицерол является мощным эндогенным активатором

Краткая характеристика опиоидных рецепторов
дельта-рецепторы
Рецепторы типа ОР1 (дельта-рецепторы). Доказано существование
Краткая характеристика опиоидных рецепторов
дельта-рецепторы
Рецепторы типа ОР1 (дельта-рецепторы). Доказано существование

каппа-рецепторы
Рецепторы типа ОР2 (каппа-рецепторы). Существует не менее трех подтипов каппа-рецепторов:
каппа-рецепторы
Рецепторы типа ОР2 (каппа-рецепторы). Существует не менее трех подтипов каппа-рецепторов:

мю-опиоидные рецепторы
ОР3-рецепторы - наиболее изученный тип. Как и в
мю-опиоидные рецепторы
ОР3-рецепторы - наиболее изученный тип. Как и в
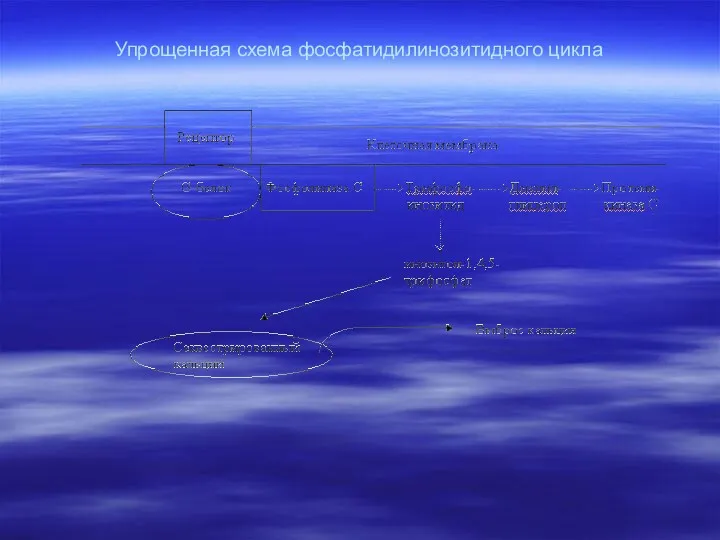
Упрощенная схема фосфатидилинозитидного цикла
Упрощенная схема фосфатидилинозитидного цикла

пептидергическая нейромедиаторная система
В пептидергической нейромедиаторной системы, передача осуществляется с участием
пептидергическая нейромедиаторная система
В пептидергической нейромедиаторной системы, передача осуществляется с участием

Эндогенный лиганд ORL-1-рецепторов ноцицептин/орфанин FG состоит из 17 аминокислотных остатков,
Эндогенный лиганд ORL-1-рецепторов ноцицептин/орфанин FG состоит из 17 аминокислотных остатков,

Передача сигнала рецепторами ORL-1 осуществляется с участием сопряженных G-белков
Передача сигнала рецепторами ORL-1 осуществляется с участием сопряженных G-белков

Как известно, одним из основных компонентов формирования пристрастия является активация
Как известно, одним из основных компонентов формирования пристрастия является активация

Схема взаимодействия опиоидных и дофаминергических систем вентральной области покрышки и прилежащего
Схема взаимодействия опиоидных и дофаминергических систем вентральной области покрышки и прилежащего

Системы вторичных мессенджеров при опиатной наркомании
Системы вторичных мессенджеров при опиатной наркомании

Нейрохимические проблемы толерантности и абстинентного синдрома
Модуляция систем вторичных передатчиков при
Нейрохимические проблемы толерантности и абстинентного синдрома
Модуляция систем вторичных передатчиков при

Негативные отношения между опиатами/опиоидами и аденилатциклазной системой первоначально показаны на
Негативные отношения между опиатами/опиоидами и аденилатциклазной системой первоначально показаны на

Наркотизация крыс морфином сопровождалась достоверным повышением активности аденилатциклазы и цАМФ-зависимой
Наркотизация крыс морфином сопровождалась достоверным повышением активности аденилатциклазы и цАМФ-зависимой

Это подтверждается сходными изменениями аденилатциклазной системы при наркотизации животных другим
Это подтверждается сходными изменениями аденилатциклазной системы при наркотизации животных другим

Одной из причин дезинтеграции аденилатциклазной системы при развитии толерантности может
Одной из причин дезинтеграции аденилатциклазной системы при развитии толерантности может

Модификация активности определенных G-белков оказывает влияние на функциональное состояние систем
Модификация активности определенных G-белков оказывает влияние на функциональное состояние систем

Как известно, система циклического аденозинмонофосфата регулирует биохимические процессы в клетке
Как известно, система циклического аденозинмонофосфата регулирует биохимические процессы в клетке

Дезинтеграция многочисленных биохимических процессов может считаться основой формирования пристрастия, толерантности
Дезинтеграция многочисленных биохимических процессов может считаться основой формирования пристрастия, толерантности

Наиболее длительным нарушением считается дезадаптационная экспрессия определенных участков генома в
Наиболее длительным нарушением считается дезадаптационная экспрессия определенных участков генома в

Традиционно в патогенезе опиатной наркомании важное место отводили изменениям опиоидных
Традиционно в патогенезе опиатной наркомании важное место отводили изменениям опиоидных

Есть сведения о вовлечении в механизмы толерантности к опиатами каскада
Есть сведения о вовлечении в механизмы толерантности к опиатами каскада

Одной из структур головного мозга, вовлеченных в развитие толерантности и
Одной из структур головного мозга, вовлеченных в развитие толерантности и

Менее определенно можно говорить о роли NMDA-рецепторов в модуляции систем вознаграждения
Менее определенно можно говорить о роли NMDA-рецепторов в модуляции систем вознаграждения

Изменения генома при опиатной наркомании
Представления о мутагенной активности опиатов
Изменения генома при опиатной наркомании
Представления о мутагенной активности опиатов

Не менее важным элементом патогенеза опиатной наркомании является способность аддиктивных
Не менее важным элементом патогенеза опиатной наркомании является способность аддиктивных

Работы последних лет свидетельствуют, что сдвиги экспрессии мРНК для многих
Работы последних лет свидетельствуют, что сдвиги экспрессии мРНК для многих

Не ясно, носят ли изменения экспрессии генома транзиторный характер и
Не ясно, носят ли изменения экспрессии генома транзиторный характер и

Если исходить из классических представлений о структурно-метаболических комплексах клетки, можно
Если исходить из классических представлений о структурно-метаболических комплексах клетки, можно

Изменения экспрессии гена, кодирующего синтез ингибитора связывания (DBI, эндогенный лиганд бензодиазепиновых
Изменения экспрессии гена, кодирующего синтез ингибитора связывания (DBI, эндогенный лиганд бензодиазепиновых

Сложнее экстраполировать данные о сдвигах содержания мРНК в плане формирования
Сложнее экстраполировать данные о сдвигах содержания мРНК в плане формирования

Нейрохимические исследования, посвященные патогенезу опиатной наркомании, являются бурно прогрессирующей областью
Нейрохимические исследования, посвященные патогенезу опиатной наркомании, являются бурно прогрессирующей областью
 Патологиялық анатомия
Патологиялық анатомия Оба қоздырғышы
Оба қоздырғышы Microscopic colitis
Microscopic colitis Сосудистые анастомозы РУС
Сосудистые анастомозы РУС Нарушение половых функций. Климакс
Нарушение половых функций. Климакс Мeningeal a syndrome in clinic of infectious diseases
Мeningeal a syndrome in clinic of infectious diseases Медсестринський процес – історичний нарис. Професійні організації медичних сестер
Медсестринський процес – історичний нарис. Професійні організації медичних сестер Оксигенотерапия. Виды оксигенотерапии
Оксигенотерапия. Виды оксигенотерапии Потребность в нормальном дыхании
Потребность в нормальном дыхании Cardiovascular system
Cardiovascular system Первая помощь детям при травмах и несчастных случаях. Общие советы для детей и взрослых
Первая помощь детям при травмах и несчастных случаях. Общие советы для детей и взрослых Медикаментозды остеопороз: алдын алудағы және түзетуге негізгі ұстанымдар
Медикаментозды остеопороз: алдын алудағы және түзетуге негізгі ұстанымдар Первая помощь при отморожении, общем охлаждении организма
Первая помощь при отморожении, общем охлаждении организма Обследование зубных рядов, зубов и периодонта в детском возрасте
Обследование зубных рядов, зубов и периодонта в детском возрасте Терапиялық стоматология клиникасындағы ауру сезімі және жансыздандыру
Терапиялық стоматология клиникасындағы ауру сезімі және жансыздандыру Общие вопросы лучевой диагностики. Ультразвуковое исследование
Общие вопросы лучевой диагностики. Ультразвуковое исследование Концепция обеспечения качества лекарственных средств
Концепция обеспечения качества лекарственных средств Клиническая симптоматология рака и абсцесса легких. Синдром полости в легком. (Тема 21)
Клиническая симптоматология рака и абсцесса легких. Синдром полости в легком. (Тема 21) Острая почечная и острая печеночная недостаточность
Острая почечная и острая печеночная недостаточность Ранний детский аутизм
Ранний детский аутизм Технические регламенты и национальные стандарты оказания оздоровительных услуг гостиничными организациями. (Лекция 6.1)
Технические регламенты и национальные стандарты оказания оздоровительных услуг гостиничными организациями. (Лекция 6.1) Уход за больными хирургического профиля
Уход за больными хирургического профиля Требования к организации и проведению профилактической дератизации и дезинсекции
Требования к организации и проведению профилактической дератизации и дезинсекции Периферические венозные катетеры
Периферические венозные катетеры Ортаңғы және ішкі құлақ аурулары. Саңыраулық және мылқаулық орта кұлактың жедел және созылмалы кабынуы мастоидит
Ортаңғы және ішкі құлақ аурулары. Саңыраулық және мылқаулық орта кұлактың жедел және созылмалы кабынуы мастоидит Нейропротезирование. История нейропротезирования
Нейропротезирование. История нейропротезирования Синдромы при заболеваниях дыхательной системы
Синдромы при заболеваниях дыхательной системы Первая медицинская помощь
Первая медицинская помощь