- Органы кроветворения (гемопоэза). Лекция №11

Содержание
- 2. статистика заболеваний кроветворных органов: их опухолевые поражения входят в число 5 самых распространенных опухолей, а у
- 3. Первичные иммунодефицитные состояния относятся к группе тяжелых генетически детерминированных заболеваний, вызванных нарушением одного или нескольких иммунных
- 4. Неспецифические факторы защиты Противоинфекционную защиту осуществляют: 1) кожа и слизистые оболочки; 2) лимфатические узлы; 3) лизоцим
- 5. Недели гестации КР. КОСТНЫЙ МОЗГ Осевой скелет конечности СЕЛЕЗЕНКА ПЕЧЕНЬ Желточный мешок Постнатальный период МЕДУЛЛЯРНЫЙ ГЕПАТОЛИЕНАЛЬНЫЙ
- 6. VI МОРФОФУНКЦИОНАЛЬНЫЕ ПРИЗНАКИ КЛЕТОК IV КЛАССА Миелоидный отдел Лимфоидный отдел
- 7. NB! По мере созревания кроветворной системы происходит специализация органов: Печень, в которой впервые появляются очаги лекоцито-
- 8. Поэтому понятие об органах кроветворения шире, чем об органах иммуногенеза, которые составляют специализированную часть кроветворной системы.
- 9. Лимфоидная ткань - специализированная ткань, обеспечивающая концентрацию антигенов; контакт клеток с антигенами; транспорт гуморальных веществ.
- 10. Строение лимфоидной ткани: рыхлая лимфоидная ткань (представлена ретикулярными волокнами, ретикулярными клетками и фиксированными макрофагами); плотная лимфоидная
- 11. Типы лимфоидной ткани: Диффузная – инфильтрация кожи и слизистых Неинкапсулированная – скопление лимфоидной ткани в виде
- 12. 1.Раннее и интенсивное формирование в эмбриогенезе и ранняя инволюция. 2.Формирование из мезенхимы (NB!тимус – ретикулоэпителиальное происхождение).
- 13. Классификация
- 14. Особенности центральных органов иммунной системы Расположены в зонах организма, защищенных от внешних воздействий (костный мозг –
- 15. Особенности периферических органов иммунной системы Расположены на путях возможного внедрения в организм чужеродных антигенов Последовательное усложнение
- 16. основные элементы органов системы иммуногенеза: строма, паренхима, сосудистые и нервные образования. NB! особенность кроветворных органов заключается
- 17. Морфологические особенности Организации паренхимы красного костного мозга NB! имеется четкая островковая организация гемопоэтических клеток в связи
- 20. Роль костного мозга в кроветворении может быть суммирована следующим: это единственный в норме орган миелопоэза; 2)
- 21. Вилочковая железа
- 22. Вилочковая железа Стволовая гемопоэтическая клетка в тимусе дифференцируется под влиянием тимического микроокружения в Т-лимфоцит. Цель дифференцировки:
- 23. ГЛАВНЫЙ КОМПЛЕКC ГИCТОCОВМЕCТИМОCТИ Синтез главных молекул гистосовместимости (ГКГС) контролирует комплекс генов ГКГС. Гены ГКГС расположены в
- 24. СТРУКТУРА Каждый антиген классов I и II содержит нековалентно связанные полипептидные цепи a и b. Цепи
- 25. Cтволовая гемопоэтическая клетка попадает в корковую зону тимуса и превращается в ранний предшественник Т-лимфоцита. Фенотип этой
- 26. Суть позитивной и негативной селекции: Позитивная селекция. Т-лимфоциты, ТcR которых обладает способностью распознавать молекулы ГКГС стромальных
- 27. Далее, здесь же в корковой зоне тимуса, под влиянием тимического микроокружения, гормонов тимуса и, особенно, ИЛ-7
- 28. В последующем одна из молекул CD4 или CD8 утрачивается и клетки становятся зрелыми. Клетки сохранившие CD4
- 29. Вилочковая железа Фотография из диссертации О.М. Шепелевой. Научные руководители Профессор И.И. Бобынцев, профессор А.В. Иванов
- 30. Вилочковая железа Фотография из диссертации О.М. Шепелевой. Научные руководители Профессор И.И. Бобынцев, профессор А.В. Иванов
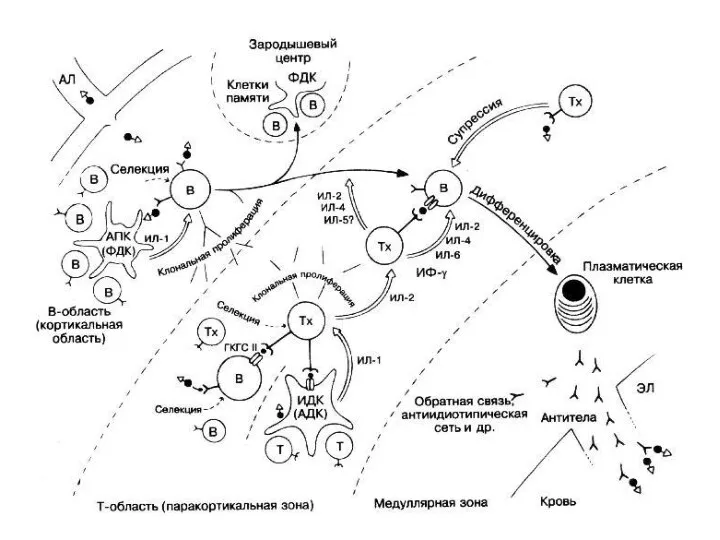
- 31. Антигеннезависимая дифференцировка В-лимфоцитов 1. Происходит под контролем локальных клеточных и гуморальных сигналов от микроокружения пре-В-лимфоцитов и
- 32. Антигензависимая дифференцировка В-лимфоцитов Начинается с момента контакта этих клеток с Аг. В результате происходит активация В-лимфоцитов,
- 33. Процессы деления и специализации В-клетки осуществляются не только под влиянием Аг, но и при обязательном участии
- 35. статистика заболеваний кроветворных органов: их опухолевые поражения входят в число 5 самых распространенных опухолей, а у
- 37. Скачать презентацию
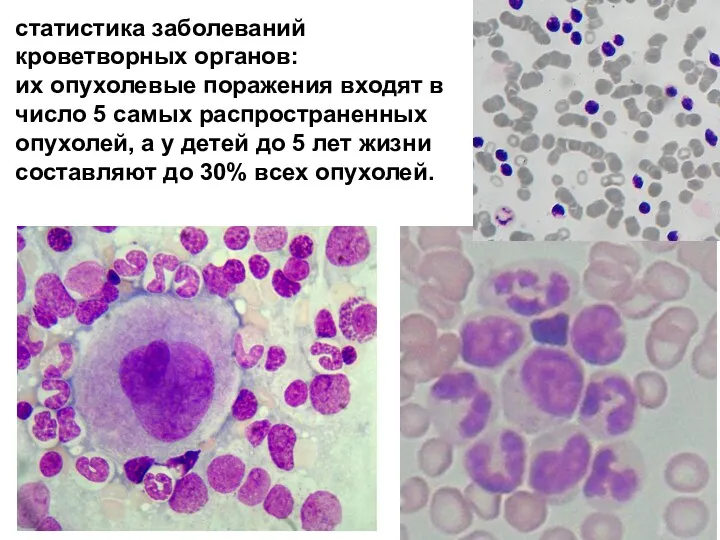
статистика заболеваний кроветворных органов:
их опухолевые поражения входят в число 5
статистика заболеваний кроветворных органов:
их опухолевые поражения входят в число 5

Первичные иммунодефицитные состояния относятся к группе тяжелых генетически детерминированных заболеваний, вызванных
Первичные иммунодефицитные состояния относятся к группе тяжелых генетически детерминированных заболеваний, вызванных

Неспецифические факторы защиты
Противоинфекционную защиту осуществляют:
1) кожа и слизистые оболочки;
2) лимфатические узлы;
3) лизоцим и другие
Неспецифические факторы защиты
Противоинфекционную защиту осуществляют:
1) кожа и слизистые оболочки;
2) лимфатические узлы;
3) лизоцим и другие
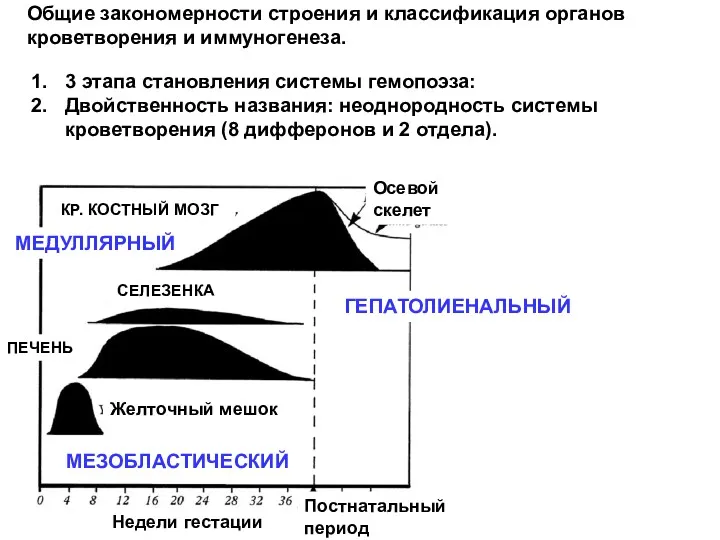
Недели гестации
КР. КОСТНЫЙ МОЗГ
Осевой скелет
конечности
СЕЛЕЗЕНКА
ПЕЧЕНЬ
Желточный мешок
Постнатальный
период
МЕДУЛЛЯРНЫЙ
ГЕПАТОЛИЕНАЛЬНЫЙ
МЕЗОБЛАСТИЧЕСКИЙ
Общие закономерности строения и классификация
Недели гестации
КР. КОСТНЫЙ МОЗГ
Осевой скелет
конечности
СЕЛЕЗЕНКА
ПЕЧЕНЬ
Желточный мешок
Постнатальный
период
МЕДУЛЛЯРНЫЙ
ГЕПАТОЛИЕНАЛЬНЫЙ
МЕЗОБЛАСТИЧЕСКИЙ
Общие закономерности строения и классификация
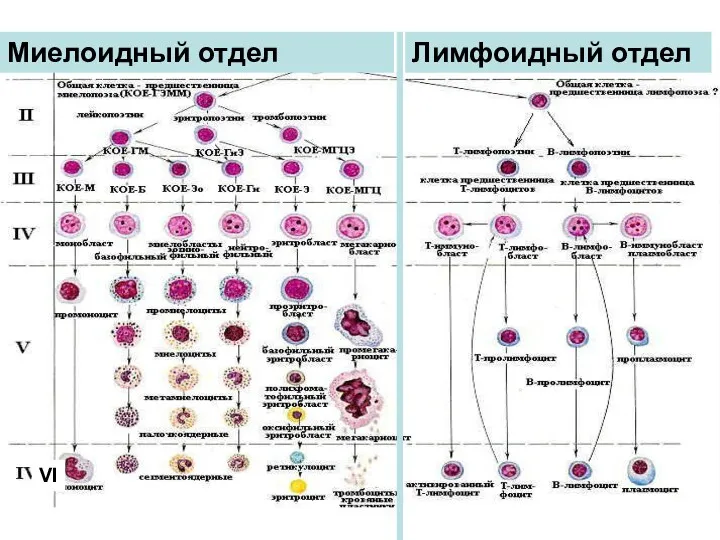
VI
МОРФОФУНКЦИОНАЛЬНЫЕ ПРИЗНАКИ КЛЕТОК IV КЛАССА
Миелоидный отдел
Лимфоидный отдел
VI
МОРФОФУНКЦИОНАЛЬНЫЕ ПРИЗНАКИ КЛЕТОК IV КЛАССА
Миелоидный отдел
Лимфоидный отдел

NB! По мере созревания кроветворной системы происходит специализация органов:
Печень, в
NB! По мере созревания кроветворной системы происходит специализация органов:
Печень, в

Поэтому понятие об органах кроветворения шире, чем об органах иммуногенеза, которые
Поэтому понятие об органах кроветворения шире, чем об органах иммуногенеза, которые

Лимфоидная ткань -
специализированная ткань, обеспечивающая
концентрацию антигенов;
контакт клеток с антигенами;
транспорт
Лимфоидная ткань -
специализированная ткань, обеспечивающая
концентрацию антигенов;
контакт клеток с антигенами;
транспорт

Строение лимфоидной ткани:
рыхлая лимфоидная ткань (представлена ретикулярными волокнами, ретикулярными клетками
Строение лимфоидной ткани:
рыхлая лимфоидная ткань (представлена ретикулярными волокнами, ретикулярными клетками

Типы лимфоидной ткани:
Диффузная – инфильтрация кожи и слизистых
Неинкапсулированная – скопление лимфоидной
Типы лимфоидной ткани:
Диффузная – инфильтрация кожи и слизистых
Неинкапсулированная – скопление лимфоидной

1.Раннее и интенсивное формирование в эмбриогенезе и ранняя инволюция.
2.Формирование из мезенхимы
1.Раннее и интенсивное формирование в эмбриогенезе и ранняя инволюция.
2.Формирование из мезенхимы
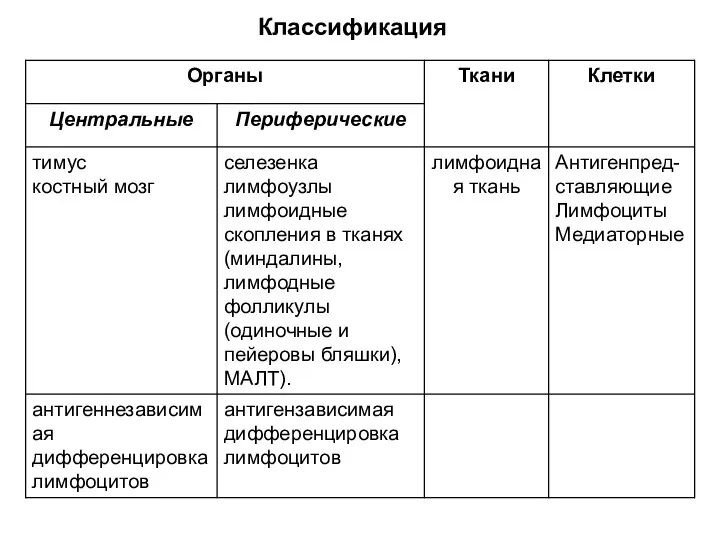
Классификация
Классификация

Особенности центральных органов иммунной системы
Расположены в зонах организма, защищенных от внешних
Особенности центральных органов иммунной системы
Расположены в зонах организма, защищенных от внешних

Особенности периферических органов иммунной системы
Расположены на путях возможного внедрения в организм
Особенности периферических органов иммунной системы
Расположены на путях возможного внедрения в организм

основные элементы органов системы иммуногенеза: строма, паренхима, сосудистые и нервные образования.
основные элементы органов системы иммуногенеза: строма, паренхима, сосудистые и нервные образования.

Морфологические особенности
Организации паренхимы красного костного мозга
NB! имеется четкая островковая организация
Морфологические особенности
Организации паренхимы красного костного мозга
NB! имеется четкая островковая организация
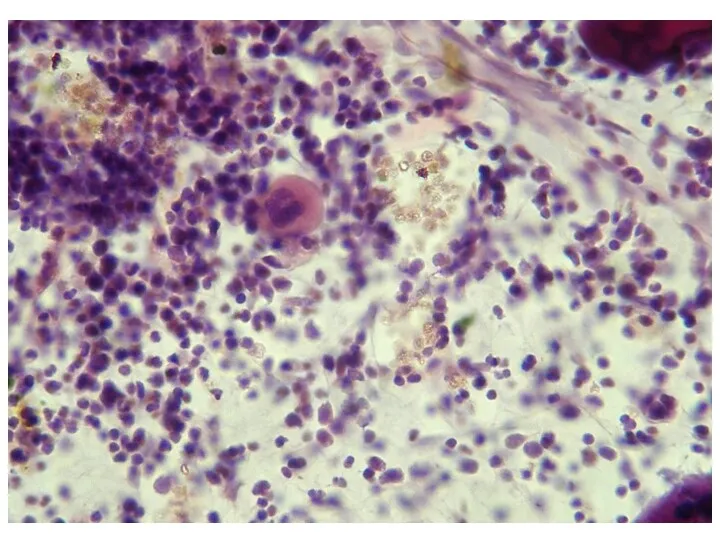
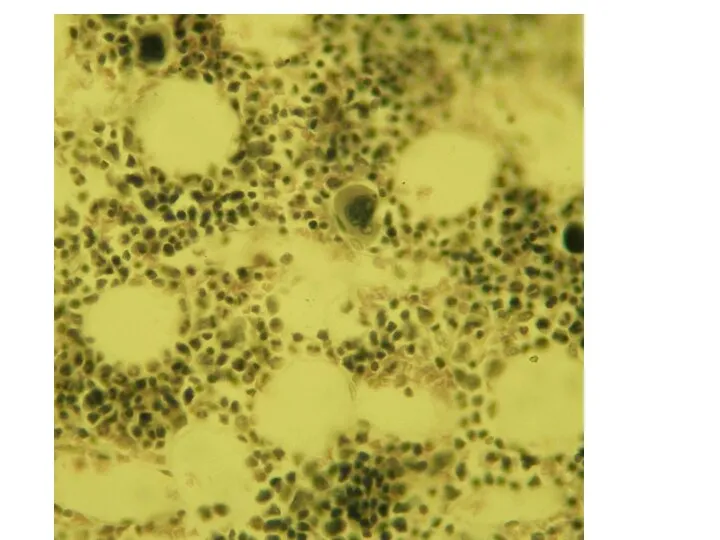

Роль костного мозга в кроветворении может быть суммирована следующим:
это единственный в
Роль костного мозга в кроветворении может быть суммирована следующим:
это единственный в
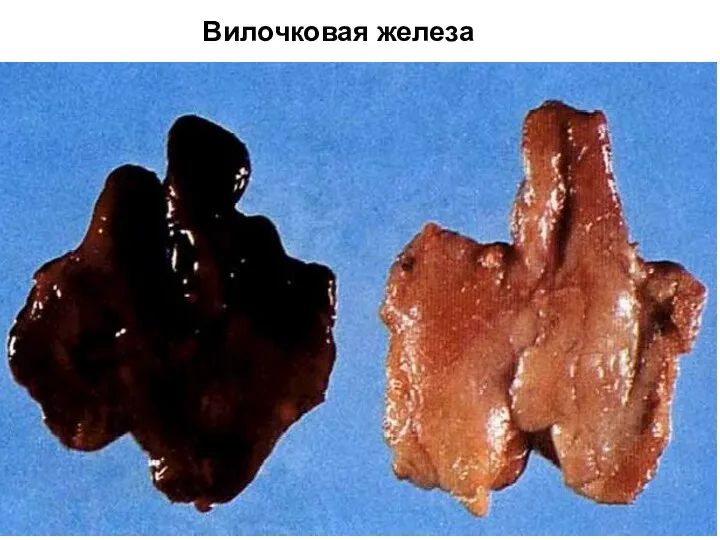
Вилочковая железа
Вилочковая железа

Вилочковая железа
Стволовая гемопоэтическая клетка в тимусе дифференцируется под влиянием тимического микроокружения
Вилочковая железа
Стволовая гемопоэтическая клетка в тимусе дифференцируется под влиянием тимического микроокружения

ГЛАВНЫЙ КОМПЛЕКC ГИCТОCОВМЕCТИМОCТИ
Синтез главных молекул гистосовместимости (ГКГС) контролирует комплекс генов ГКГС. Гены ГКГС
ГЛАВНЫЙ КОМПЛЕКC ГИCТОCОВМЕCТИМОCТИ
Синтез главных молекул гистосовместимости (ГКГС) контролирует комплекс генов ГКГС. Гены ГКГС

СТРУКТУРА
Каждый антиген классов I и II содержит нековалентно связанные полипептидные цепи a и b. Цепи на
СТРУКТУРА
Каждый антиген классов I и II содержит нековалентно связанные полипептидные цепи a и b. Цепи на

Cтволовая гемопоэтическая клетка попадает в корковую зону тимуса и превращается в
Cтволовая гемопоэтическая клетка попадает в корковую зону тимуса и превращается в

Суть позитивной и негативной селекции:
Позитивная селекция. Т-лимфоциты, ТcR которых обладает способностью
Суть позитивной и негативной селекции:
Позитивная селекция. Т-лимфоциты, ТcR которых обладает способностью

Далее, здесь же в корковой зоне тимуса, под влиянием тимического микроокружения,
Далее, здесь же в корковой зоне тимуса, под влиянием тимического микроокружения,

В последующем одна из молекул CD4 или CD8 утрачивается и клетки
В последующем одна из молекул CD4 или CD8 утрачивается и клетки
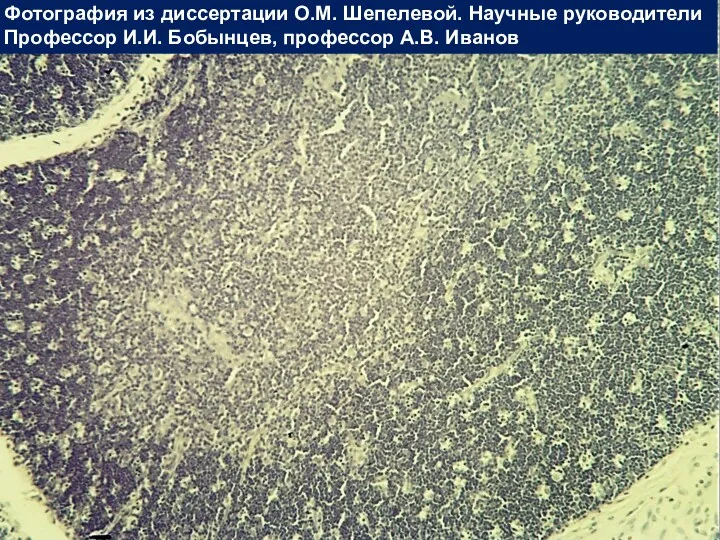
Вилочковая железа
Фотография из диссертации О.М. Шепелевой. Научные руководители
Профессор И.И. Бобынцев, профессор
Вилочковая железа
Фотография из диссертации О.М. Шепелевой. Научные руководители
Профессор И.И. Бобынцев, профессор
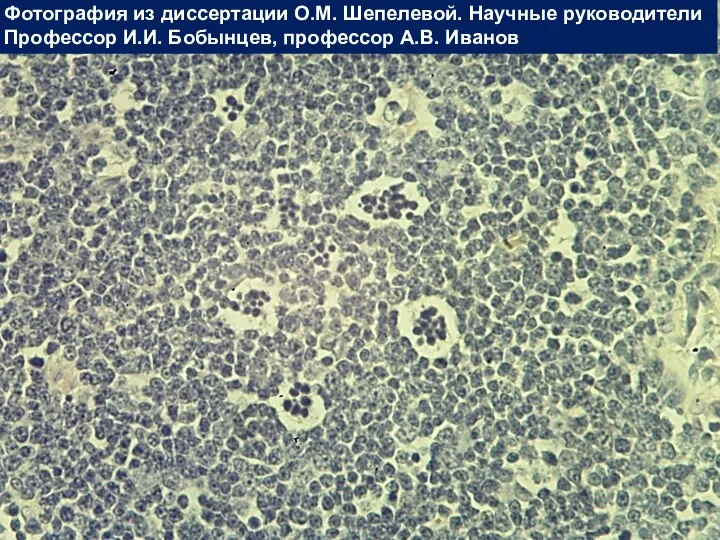
Вилочковая железа
Фотография из диссертации О.М. Шепелевой. Научные руководители
Профессор И.И. Бобынцев, профессор
Вилочковая железа
Фотография из диссертации О.М. Шепелевой. Научные руководители
Профессор И.И. Бобынцев, профессор

Антигеннезависимая дифференцировка В-лимфоцитов
1. Происходит под контролем локальных клеточных и гуморальных
Антигеннезависимая дифференцировка В-лимфоцитов
1. Происходит под контролем локальных клеточных и гуморальных

Антигензависимая дифференцировка В-лимфоцитов
Начинается с момента контакта этих клеток с Аг.
Антигензависимая дифференцировка В-лимфоцитов
Начинается с момента контакта этих клеток с Аг.

Процессы деления и специализации В-клетки осуществляются не только под влиянием Аг,
Процессы деления и специализации В-клетки осуществляются не только под влиянием Аг,
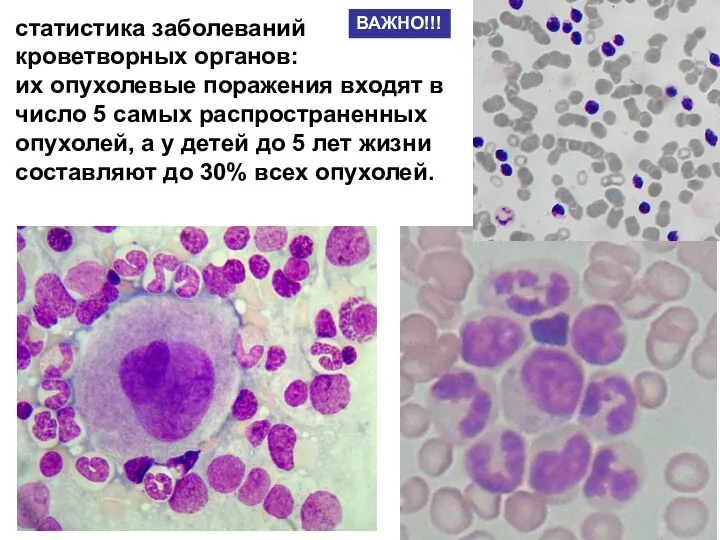
статистика заболеваний кроветворных органов:
их опухолевые поражения входят в число 5
статистика заболеваний кроветворных органов:
их опухолевые поражения входят в число 5
 Врожденный и приобретенный иммунитет. Клеточные и гуморальные механизмы
Врожденный и приобретенный иммунитет. Клеточные и гуморальные механизмы Спортивна медицина
Спортивна медицина Трихинеллёз. Возбудитель Трихинеллёза
Трихинеллёз. Возбудитель Трихинеллёза Инфекция. Классификация инфекций и инвазий человека
Инфекция. Классификация инфекций и инвазий человека Болезни почек
Болезни почек Эндокриндік жүйенің бұзылыстары
Эндокриндік жүйенің бұзылыстары Геморрагический инсульт (спонтанное субарахноидальное кровоизлияние – САК) у детей
Геморрагический инсульт (спонтанное субарахноидальное кровоизлияние – САК) у детей Диагностика рака щитовидной железы
Диагностика рака щитовидной железы Инструментальные и функциональные методы исследования сердца
Инструментальные и функциональные методы исследования сердца Стоматологическая служба в системе охраны материнства и детства
Стоматологическая служба в системе охраны материнства и детства Мочеполовая система
Мочеполовая система General course of syphilis. Primary syphilis secondary syphslis
General course of syphilis. Primary syphilis secondary syphslis Острый одонтогенный, гематогенный, хронический остеомиелит
Острый одонтогенный, гематогенный, хронический остеомиелит Противовоспалительные лекарственные средства (ПВЛС)
Противовоспалительные лекарственные средства (ПВЛС) Оборотные средства аптечной организации. (Тема 21)
Оборотные средства аптечной организации. (Тема 21) Первая помощь при растяжении связок, вывихах суставов, переломах костей
Первая помощь при растяжении связок, вывихах суставов, переломах костей Общая семиотика экстрапирамидных нарушений
Общая семиотика экстрапирамидных нарушений Остеопороз и его связь со стоматологическими заболеваниями
Остеопороз и его связь со стоматологическими заболеваниями Өлім жақындағанда адамның бойында не болады
Өлім жақындағанда адамның бойында не болады Гомеостаз зуба после прорезывания. Резистентность эмали зуба к кариозному поражению. Влияние на процессы формирования
Гомеостаз зуба после прорезывания. Резистентность эмали зуба к кариозному поражению. Влияние на процессы формирования Сердечная недостаточность у детей
Сердечная недостаточность у детей Нравственная и правовая ответственность врача перед пациентом
Нравственная и правовая ответственность врача перед пациентом Сальмонеллез. Основные факторы патогенности
Сальмонеллез. Основные факторы патогенности Бронхит. Классификация по течению болезни
Бронхит. Классификация по течению болезни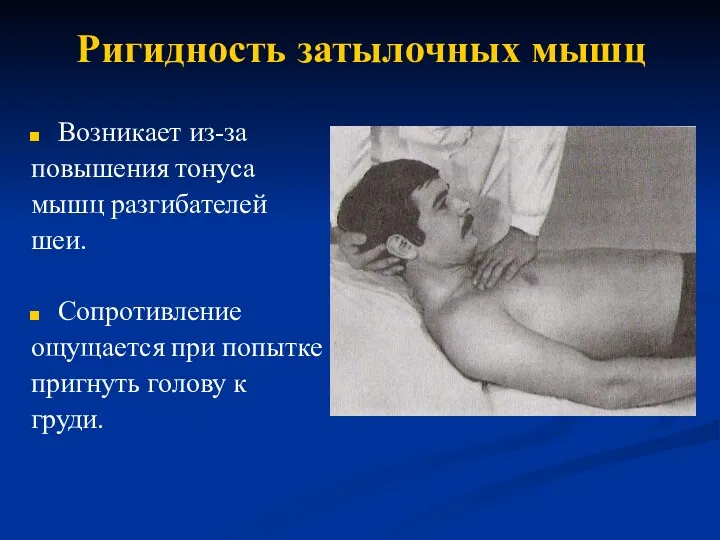 Ригидность затылочных мышц
Ригидность затылочных мышц Ғылыми жұмыстағы әдістер мен материалдар
Ғылыми жұмыстағы әдістер мен материалдар Склера. Заболевания склеры
Склера. Заболевания склеры Артериальная гипертензия у беременных в терапии и гинекологии: два взгляда – одно решение
Артериальная гипертензия у беременных в терапии и гинекологии: два взгляда – одно решение