- Радиационно-индуцированные аберрации хромосом

Содержание
- 2. Одним из наиболее чувствительных радиобиологических способов определения изменений, происходящих в организме при его облучении, является цитогенетический
- 3. 1. ОСНОВНЫЕ ТИПЫ ХРОМОСОМНЫХ АБЕРРАЦИЙ Все хромосомные аберрации, возникающие в соматических клетках человека и регистрируемые на
- 4. Отнесение той или иной аберрации к хромосомному или хроматидному типу зависит от того, на каком уровне
- 5. Обмены классифицируются по нескольким принципам: ⎯ в зависимости от того, произошел ли обмен внутри одной хромосомы
- 6. 1. 1. Аберрации хромосомного типа Среди аберраций хромосомного типа выделяют простые (парные фрагменты) и обмены. Обмены
- 7. Рис. 1.1 Происхождение парных и изохроматидных фрагментов и отличие парных фрагментов от изохроматидных пробелов. Парные фрагменты
- 8. Внутрихромосомные обмены Внутрихромосомные обмены могут быть как внутриплечевыми, так и межплечевыми. К внутрихромосомным внутриплечевым обменам относятся
- 9. Точечные ацентрические кольца трудно отдифференцировать от парных фрагментов небольших размеров. Их следует относить к ацентрическим кольцам
- 10. Перицентрические и парацентрические инверсии. Перицентрические инверсии относятся к межплечевым обменам и представляют собой результат инверсии (переворота
- 11. Межхромосомные обмены Межхромосомные обмены встречаются довольно часто, в особенности при исследовании действия излучения. В обмен могут
- 12. Полицентрические хромосомы. Это межхромосомные обмены, включающие две и более центромеры. Все они являются ассиметричными обменами. Чаще
- 13. 1.2. Аберрации хроматидного типа Конфигурация аберраций хроматидного типа крайне разнообразна. Среди аберраций хроматидного типа также выделяют
- 14. Внутрихромосомные внутриплечевые обмены Они могут возникать чаще всего в результате двух повреждений в одной и той
- 15. Два повреждения в одном и том же плече хроматиды приводят к появлению дистального и интерстициального сегментов.
- 16. Межхроматидные обмены. Принято называть их обменами со слиянием сестринских хроматид (рис. 1.9.). Рис. 1.9. Обмены со
- 17. Внутрихромосомные межплечевые обмены Конфигурация внутрихромосомных межплечевых обменов хроматидного типа часто бывает сложной и их идентификация порой
- 18. Межхромосомные обмены. Такие обмены хроматидного типа встречаются более часто, чем другие обмены из этой группы, при
- 19. Рис. 1.11. Межхромосомные хроматидо-хроматидные обмены. Развернутое обозначение этих обменов, применяемое не часто и то лишь в
- 20. Хроматидо-изохроматидные обмены. В обменах этого типа принимают участие две хромосомы, в одной из которых произошел хроматидный
- 21. Основные правила при анализе и учете аберраций хромосом: ∙В первом митозе дицентрические и кольцевые хромосомы почти
- 22. ПРОТОКОЛ ЦИТОГЕНЕТИЧЕСКОГО АНАЛИЗА Рис. 1.13. Образец бланка-протокола цитогенетического анализа
- 23. Литература 1. Бектон К., Эванс Г. Методы анализа хромосомных аберраций у человека. М.: Медицина, 1975. 64
- 24. Таблица 2.1. Частота спонтанных аберраций хромосом у лиц разного возраста и пола 2. СПОНТАННЫЙ УРОВЕНЬ АБЕРРАЦИЙ
- 25. Все проанализированные культуры были разделены на 2 группы: зафиксированные на 50−60−м часу культивирования (первый митоз) и
- 26. Таблица 2. 3. Типы спонтанных хромосомных аберраций Среди обнаруженных аберраций более 90% составили ацентрические фрагменты (одиночные
- 27. Распределение исследованных культур по частоте клеток с хромосомными аберрациями оказалось следующим (табл. 2.4): из 527 изученных
- 28. Таблица 2.4. Распределение культур по частоте клеток с хромосомными аберрациями Индивиды, со средней частотой встречаемости аберраций
- 29. Литература 1. Бочков Н. П. Учет хромосомных аберраций в соматических клетках / Кн.: Основы цитогенетики человека.
- 30. 3. МОЛЕКУЛЯРНЫЕ МЕХАНИЗМЫ ОБРАЗОВАНИЯ РАДИАЦИОННО-ИНДУЦИРОВАННЫХ АБЕРРАЦИЙ ХРОМОСОМ Радиация высоких энергий способна разрушить любую связь в органических
- 31. Фрагментация ДНК. В настоящее время доказано существование высоко интенсивных радиационно индуцированных процессов деградации молекул ДНК до
- 32. Разрывы водородных связей двойной спирали ДНК. Вторичные реакции − это разрывы водородных связей, конфигурационные изменения надмолекулярных
- 33. Потеря ДНК пуриновых оснований (депуринирование). Одним из частых изменений ДНК при облучении является повреждение оснований (потери
- 34. 3.1. Механизмы рекомбинации ДНК Существует три вида рекомбинации: общая (гомологическая), сайт-специфическая и случайная (негомологичная). Общая рекомбинация.
- 35. Сайт−специфическая рекомбинация. Рекомбинация называется сайт−специфическая, если сайты разрыва и воссоединения в двух рекомбинирующих молекул или двух
- 36. 3.2. Радиационно-индуцированные аберрации хромосом Аберрации хромосом, индуцируемые ионизирующим излучением в соматических клетках человека, подразделяются на 2
- 37. Аберрации хроматидного типа возникают при облучении клеток в G 2, S и позднем G1 − периодах
- 38. Одноударные аберрации линейно зависят от дозы, а для более сложных аберраций наблюдается нелинейная зависимость. Выход одноударных
- 39. Следующим важным параметром в биологической дозиметрии является линейная передача энергии (ЛПЭ), поскольку одиночный трек плотно ионизирующей
- 41. Скачать презентацию

Одним из наиболее чувствительных радиобиологических способов определения изменений, происходящих в организме
Одним из наиболее чувствительных радиобиологических способов определения изменений, происходящих в организме
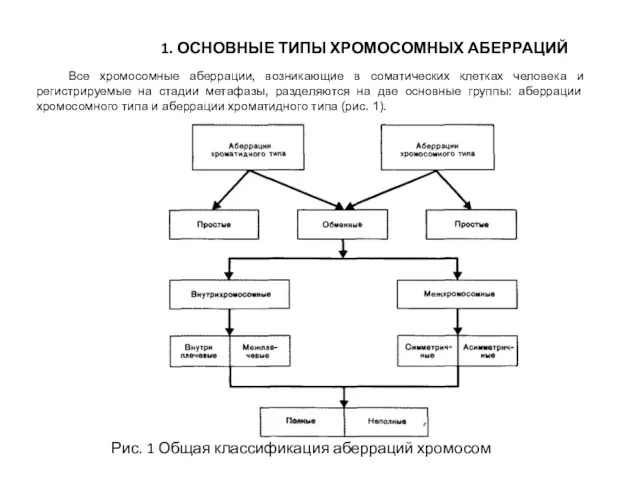
1. ОСНОВНЫЕ ТИПЫ ХРОМОСОМНЫХ АБЕРРАЦИЙ
Все хромосомные аберрации, возникающие в соматических клетках
1. ОСНОВНЫЕ ТИПЫ ХРОМОСОМНЫХ АБЕРРАЦИЙ
Все хромосомные аберрации, возникающие в соматических клетках

Отнесение той или иной аберрации к хромосомному или хроматидному типу зависит
Отнесение той или иной аберрации к хромосомному или хроматидному типу зависит

Обмены классифицируются по нескольким принципам:
⎯ в зависимости от того, произошел ли
Обмены классифицируются по нескольким принципам:
⎯ в зависимости от того, произошел ли

1. 1. Аберрации хромосомного типа
Среди аберраций хромосомного типа выделяют простые (парные
1. 1. Аберрации хромосомного типа
Среди аберраций хромосомного типа выделяют простые (парные

Рис. 1.1 Происхождение парных и изохроматидных фрагментов и отличие парных фрагментов
Рис. 1.1 Происхождение парных и изохроматидных фрагментов и отличие парных фрагментов

Внутрихромосомные обмены
Внутрихромосомные обмены могут быть как внутриплечевыми, так и межплечевыми.
К
Внутрихромосомные обмены
Внутрихромосомные обмены могут быть как внутриплечевыми, так и межплечевыми.
К

Точечные ацентрические кольца трудно отдифференцировать от парных фрагментов небольших размеров. Их
Точечные ацентрические кольца трудно отдифференцировать от парных фрагментов небольших размеров. Их

Перицентрические и парацентрические инверсии.
Перицентрические инверсии относятся к межплечевым обменам и
Перицентрические и парацентрические инверсии.
Перицентрические инверсии относятся к межплечевым обменам и

Межхромосомные обмены
Межхромосомные обмены встречаются довольно часто, в особенности при исследовании действия
Межхромосомные обмены
Межхромосомные обмены встречаются довольно часто, в особенности при исследовании действия

Полицентрические хромосомы. Это межхромосомные обмены, включающие две и более центромеры. Все
Полицентрические хромосомы. Это межхромосомные обмены, включающие две и более центромеры. Все

1.2. Аберрации хроматидного типа
Конфигурация аберраций хроматидного типа крайне разнообразна. Среди аберраций
1.2. Аберрации хроматидного типа
Конфигурация аберраций хроматидного типа крайне разнообразна. Среди аберраций
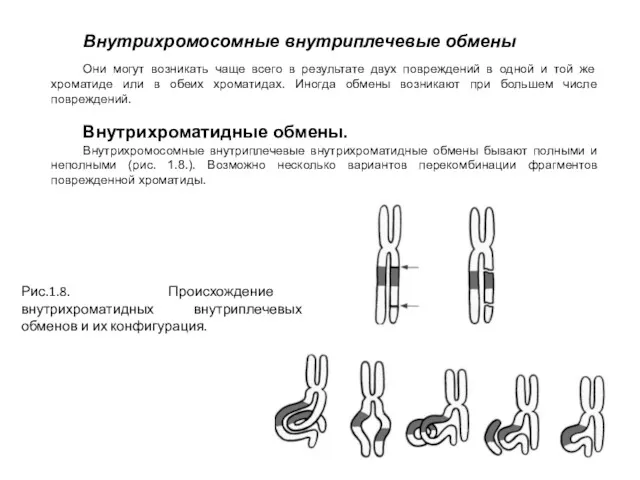
Внутрихромосомные внутриплечевые обмены
Они могут возникать чаще всего в результате двух повреждений
Внутрихромосомные внутриплечевые обмены
Они могут возникать чаще всего в результате двух повреждений

Два повреждения в одном и том же плече хроматиды приводят к
Два повреждения в одном и том же плече хроматиды приводят к

Межхроматидные обмены.
Принято называть их обменами со слиянием сестринских хроматид (рис.
Межхроматидные обмены.
Принято называть их обменами со слиянием сестринских хроматид (рис.

Внутрихромосомные межплечевые обмены
Конфигурация внутрихромосомных межплечевых обменов хроматидного типа часто бывает сложной
Внутрихромосомные межплечевые обмены
Конфигурация внутрихромосомных межплечевых обменов хроматидного типа часто бывает сложной

Межхромосомные обмены. Такие обмены хроматидного типа встречаются более часто, чем другие
Межхромосомные обмены. Такие обмены хроматидного типа встречаются более часто, чем другие

Рис. 1.11. Межхромосомные хроматидо-хроматидные обмены.
Развернутое обозначение этих обменов, применяемое не часто
Рис. 1.11. Межхромосомные хроматидо-хроматидные обмены.
Развернутое обозначение этих обменов, применяемое не часто

Хроматидо-изохроматидные обмены. В обменах этого типа принимают участие две хромосомы, в
Хроматидо-изохроматидные обмены. В обменах этого типа принимают участие две хромосомы, в

Основные правила при анализе и учете аберраций хромосом:
∙В первом митозе дицентрические
Основные правила при анализе и учете аберраций хромосом:
∙В первом митозе дицентрические

ПРОТОКОЛ ЦИТОГЕНЕТИЧЕСКОГО АНАЛИЗА
Рис. 1.13. Образец бланка-протокола цитогенетического анализа
ПРОТОКОЛ ЦИТОГЕНЕТИЧЕСКОГО АНАЛИЗА
Рис. 1.13. Образец бланка-протокола цитогенетического анализа

Литература
1. Бектон К., Эванс Г. Методы анализа хромосомных аберраций у человека.
Литература
1. Бектон К., Эванс Г. Методы анализа хромосомных аберраций у человека.
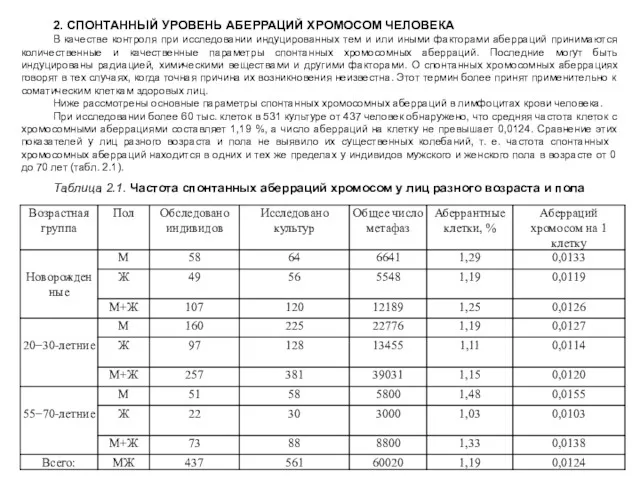
Таблица 2.1. Частота спонтанных аберраций хромосом у лиц разного возраста и
Таблица 2.1. Частота спонтанных аберраций хромосом у лиц разного возраста и
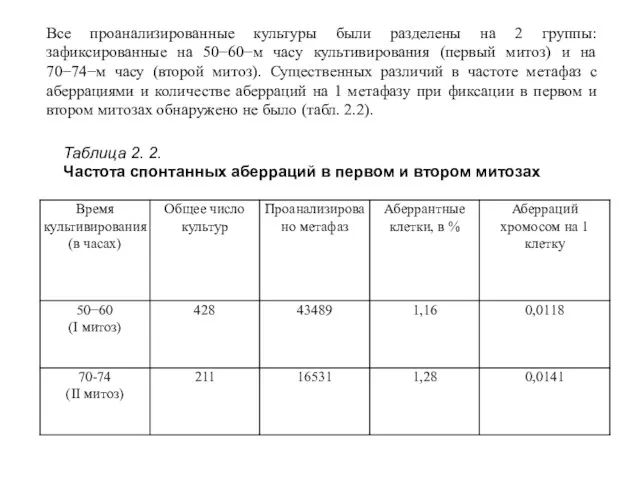
Все проанализированные культуры были разделены на 2 группы: зафиксированные на 50−60−м
Все проанализированные культуры были разделены на 2 группы: зафиксированные на 50−60−м
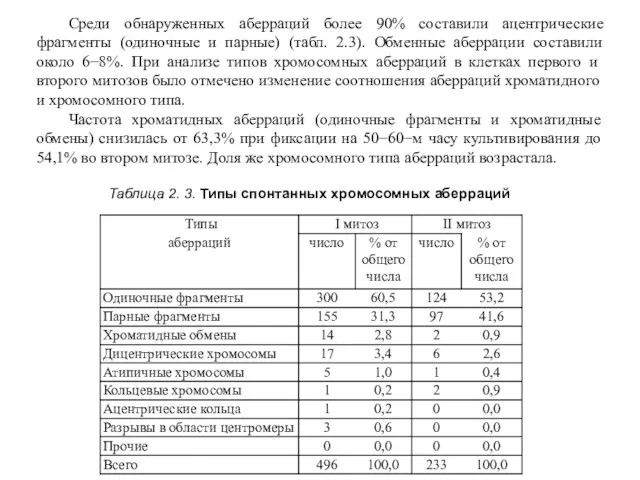
Таблица 2. 3. Типы спонтанных хромосомных аберраций
Среди обнаруженных аберраций более 90%
Таблица 2. 3. Типы спонтанных хромосомных аберраций
Среди обнаруженных аберраций более 90%

Распределение исследованных культур по частоте клеток с хромосомными аберрациями оказалось следующим
Распределение исследованных культур по частоте клеток с хромосомными аберрациями оказалось следующим
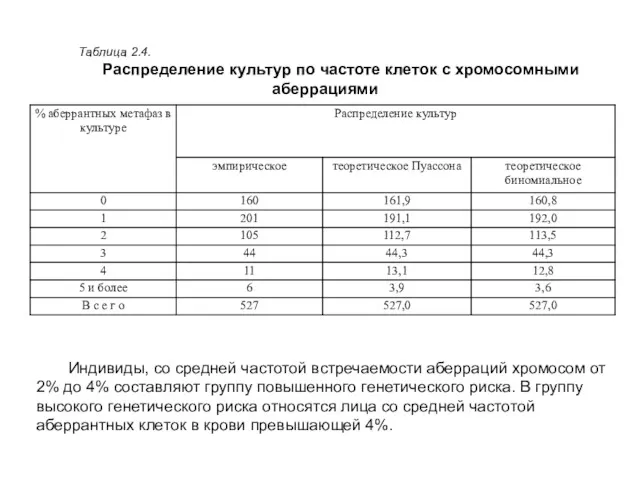
Таблица 2.4.
Распределение культур по частоте клеток с хромосомными аберрациями
Индивиды, со средней
Таблица 2.4.
Распределение культур по частоте клеток с хромосомными аберрациями
Индивиды, со средней

Литература
1. Бочков Н. П. Учет хромосомных аберраций в соматических клетках /
Литература
1. Бочков Н. П. Учет хромосомных аберраций в соматических клетках /

3. МОЛЕКУЛЯРНЫЕ МЕХАНИЗМЫ ОБРАЗОВАНИЯ РАДИАЦИОННО-ИНДУЦИРОВАННЫХ АБЕРРАЦИЙ ХРОМОСОМ
Радиация высоких энергий способна разрушить
3. МОЛЕКУЛЯРНЫЕ МЕХАНИЗМЫ ОБРАЗОВАНИЯ РАДИАЦИОННО-ИНДУЦИРОВАННЫХ АБЕРРАЦИЙ ХРОМОСОМ
Радиация высоких энергий способна разрушить

Фрагментация ДНК. В настоящее время доказано существование высоко интенсивных радиационно индуцированных
Фрагментация ДНК. В настоящее время доказано существование высоко интенсивных радиационно индуцированных

Разрывы водородных связей двойной спирали ДНК. Вторичные реакции − это разрывы водородных связей,
Разрывы водородных связей двойной спирали ДНК. Вторичные реакции − это разрывы водородных связей,

Потеря ДНК пуриновых оснований (депуринирование).
Одним из частых изменений ДНК при
Потеря ДНК пуриновых оснований (депуринирование).
Одним из частых изменений ДНК при

3.1. Механизмы рекомбинации ДНК
Существует три вида рекомбинации: общая (гомологическая), сайт-специфическая и
3.1. Механизмы рекомбинации ДНК
Существует три вида рекомбинации: общая (гомологическая), сайт-специфическая и

Сайт−специфическая рекомбинация. Рекомбинация называется сайт−специфическая, если сайты разрыва и воссоединения в
Сайт−специфическая рекомбинация. Рекомбинация называется сайт−специфическая, если сайты разрыва и воссоединения в

3.2. Радиационно-индуцированные аберрации хромосом
Аберрации хромосом, индуцируемые ионизирующим излучением в соматических клетках
3.2. Радиационно-индуцированные аберрации хромосом
Аберрации хромосом, индуцируемые ионизирующим излучением в соматических клетках

Аберрации хроматидного типа возникают при облучении клеток в G 2, S
Аберрации хроматидного типа возникают при облучении клеток в G 2, S

Одноударные аберрации линейно зависят от дозы, а для более сложных аберраций
Одноударные аберрации линейно зависят от дозы, а для более сложных аберраций

Следующим важным параметром в биологической дозиметрии является линейная передача энергии (ЛПЭ),
Следующим важным параметром в биологической дозиметрии является линейная передача энергии (ЛПЭ),
 Аномалии положения зубов в вертикальной плоскости. Этиопатогенез, клиника, лечение
Аномалии положения зубов в вертикальной плоскости. Этиопатогенез, клиника, лечение Жұлын қантамырлық аурулары
Жұлын қантамырлық аурулары Жасөспірім шақтағы аборт жасатудың медико-әлеуметтік өзекті проблема ретінде анализдеу тақырыбындағы дипломдық жұмыс
Жасөспірім шақтағы аборт жасатудың медико-әлеуметтік өзекті проблема ретінде анализдеу тақырыбындағы дипломдық жұмыс Иммунный ответ организма. Связывание антител с антигеном. (Лекция 6)
Иммунный ответ организма. Связывание антител с антигеном. (Лекция 6) свертывание
свертывание Всероссийская акция Будь здоров
Всероссийская акция Будь здоров Отработка. Особенности анатомического строения зубов боковой группы : премоляров , моляров верхней , нижней челюстей
Отработка. Особенности анатомического строения зубов боковой группы : премоляров , моляров верхней , нижней челюстей Патология пищеварительной системы
Патология пищеварительной системы Острый панкреатит
Острый панкреатит Бронхиальная астма
Бронхиальная астма Шов сухожилия
Шов сухожилия Дыхательная гимнастика Стрельниковой для детей
Дыхательная гимнастика Стрельниковой для детей Ожирение. Целлюлит
Ожирение. Целлюлит Снотворные препараты
Снотворные препараты Оригинальные препараты и дженерики
Оригинальные препараты и дженерики Синдром наличия жидкости и газа в плевральной полости. Плевриты
Синдром наличия жидкости и газа в плевральной полости. Плевриты Лечебно-эвакуационное обеспечение населения в чрезвычайных ситуациях
Лечебно-эвакуационное обеспечение населения в чрезвычайных ситуациях Гнойные заболевания костей, кисти, стопы и суставов
Гнойные заболевания костей, кисти, стопы и суставов Ранние половые связи и их последствия
Ранние половые связи и их последствия Организмнің биоритміне байланысты дәрілік заттардың көрсететін әсерлері
Организмнің биоритміне байланысты дәрілік заттардың көрсететін әсерлері Рациональное использование лекарственных средств, применяемых в кардиологии. Тактика применения гипотензивных средств
Рациональное использование лекарственных средств, применяемых в кардиологии. Тактика применения гипотензивных средств Характеристика уровней речевого развития при ОНР
Характеристика уровней речевого развития при ОНР Корь
Корь Под знаком красного креста
Под знаком красного креста Бюгельный протез
Бюгельный протез Заболевание щитовидной железы
Заболевание щитовидной железы Халық денсаулығы және денсаулық сақтау жүйесі туралы
Халық денсаулығы және денсаулық сақтау жүйесі туралы Вибрионы – возбудители холеры (Vibrio cholerae)
Вибрионы – возбудители холеры (Vibrio cholerae)