- Регуляция и функции репродуктивной системы

Содержание
- 2. Репродуктивная система (РС) РС выполняет множество функций, наиболее важной из которых является продолжение биологического рода. В
- 3. Основой регуляции функции РС является принцип отрицательной обратной связи между различными уровнями, т.е.при снижении концентрации периферических
- 6. РС состоит из 5 уровней: Экстрагипоталамического Гипоталамуса Гипофиза Яичников Органов-мишеней Как и все системы в организме,
- 7. Основная роль в регуляции функции РС принадлежит гипоталамо-гипофизарной системе, которая координирует все эндокринные системы в организме.
- 8. ДА, НА и серотонину принадлежит ведущая роль в контроле гипоталамической секреции гонадостропного рилизинг-гормона (ГнРГ). ДА поддерживает
- 9. В последние годы установлено, что в структурах мозга синтезируются стероиды, в частности ДЭА, ДЭА-С и их
- 10. Гипоталамус Высший вегетативный центр, гибрид нервной и эндокринной систем, координирующий функции всех внутренних органов и систем,
- 11. Секреция ГнРГ генетически запрограммирована, происходит в определенном пульсирующем режиме, примерно 1 раз в час. Этот ритм
- 12. Гипофиз Гипофиз – место синтеза и выделения всех тропных гормонов, непосредственной регулирующих функцию периферических эндокринных желез.
- 13. Биологическая роль ФСГ Рост фолликулов в яичниках Пролиферация клеток гранулезы в фоллликулах Синтез ароматаз, метаболирующих андрогены
- 14. Биологическая роль ЛГ: Синтез андрогенов в клетках тека фолликулов Синтез эстрадиола в доминантном фолликуле Способствует овуляции
- 15. Пролактин (ПРЛ) синтезируется клетками аденогипофиза (лактотрофами), период его полураспада составляет 50-60 мин. По химическим и биологическим
- 16. Таким образом, синтез гонадотропинов контролируется гипоталамическим ГнРГ и периферическими овариальными стероидами по механизму обратной связи.
- 17. Яичники Яичники являются местом синтеза половых стероидов: эстрогенов, андрогенов и прогестерона в процессе роста и созревания
- 19. Факторы роста (ФР) ФР – биологически активные вещества, стимулирующие или ингибирующие дифференцировку клеток, передающих гормональный сигнал.
- 20. Наиболее важную роль в физиологии РС играют следующие ФР: Инсулиноподобные факторы роста (ИФР) Эпидермальный фактор роста
- 21. Ингибины – белковые вещества, образуются в клетках гранулезы фолликула, участвуют в регуляции секреции ФСГ, тормозя ее,
- 22. Активины обнаружены в гранулезных клетках фолликула и гонадотрофах гипофиза. Активины … стимулируют синтез ФСГ, пролиферацию клеток
- 23. Инсулиноподобные факторы роста I и II (ИФР-I и ИФР-II) синтезируются в клетках гранулы и других тканях,
- 24. СЭФР играет важную роль в ангиогенезе растущих фолликулов, а в доминантном фолликуле создает повышенные концентрации ФСГ.
- 25. . Эпидермальный вактор роста (ЭФР) – обнаружен в клетках гранулезы, строме эндометрия, молочных железах и др.
- 26. Трансформирующий фактор роста ТФР-α и ТФР-β) стимулируют клеточную пролиферацию, оказывают митогенный и онкогенный эффекты. Экспрессия этих
- 27. Фолликулогенез в яичниках В яичнике женщине репродуктивного возраста фолликулы находятся на различных стадиях зрелости. Фолликулогенез начинается
- 28. Примордиальные фолликулы характеризуются одним слоем плоских прегранулезных клеток, небольшим ооцитом, клетки тека отсутствуют. Первичные преантральные фолликулы
- 29. Стадии роста фолликулов От примордиальных до преполостных фолликулов негормональнозависимый рост. Он продолжается до образования фолликулов диаметром
- 30. Рост фолликула от покоящегося примордиального до преовуляторного носит последовательный и непрерывный характер; до конца неизвестно сколько
- 31. Гонадотропин-зависимый рост овариальных фолликулов начинается с увеличения уровня ФСГ в конце предыдущего МЦ.
- 32. Гонадотропин-зависимый рост овариальных фолликулов начинается с увеличения уровня ФСГ в конце предыдущего МЦ. Повышение синтеза и
- 33. Особое значение имеет базальный уровень ФСГ на 2-3 д.м.ц. Этот показатель отражает минимальный уровень ФСГ, необходимый
- 34. В ранней фолликулярной фаз до 5-го дня цикла рост фолликулов зависит от ФСГ, их размеры составляют
- 35. Биологическая роль ИФР в созревании фолликула заключается в стимуляции клеток гранулезы к пролиферации и дифференцировке, повышению
- 36. МАХ значения уровень ФСГ достигает к 5 д.м.ц, после чего снижается, до повышения одновременно с ЛГ,
- 37. Девиация – (с англ. Deviation – отклонение) способность роста в условиях снижения уровня ФСГ. В быстром
- 38. Овуляция Процесс овуляции происходит при достижении максимального уровня эстрадиола в преовуляторном фолликуле ,который по положительной обратной
- 39. После овуляции клетки гранулезы подвергаются дальнейшей лютеинизации с образованием ЖТ, секретирующего прогестерон под влиянием ЛГ. Структурное
- 40. Основным регулятором синтеза стероидов в ЖТ является ЛГ. Снижение активности ЖТ м.б. связан с уменьшением количества
- 41. Биосинтез стероидов в яичниках – сложный процесс. Он происходит в соответствии с двухклеточной теорией, предложенной Flack
- 42. Первые этапы синтеза стероидов – до прегненолона. Энзимные системы в стероидогенезе, идентичны во всех трех железах.
- 43. Первые этапы синтеза овариальных стероидов детерминированы гонадтропинами. Под влиянием ЛГ, рецепторы которого находятся на мембране клеток
- 46. Максимальные концентрации прогестерона синтезируются после лютеинизации клеток гранулезы при функционировании ЖТ. Синтез половых стероидов происходит также
- 47. Важная роль в гормональном балансе отводится половым стероид-связывающим глобулинам (ПССГ), синтез которых происходит в печени, под
- 48. Биологическое действие эстрогенов I На репродуктивные органы: Пролиферация и гиперплазия эндо- и миометрия, эрителия влагалища, шейки
- 49. Биологическое действие эстрогенов II На экстрагенитальную систему: Пролиферативные процессы слизистой уретры, мочевого пузыря; Развитие костно-мышечной системы;
- 50. Биологическое действие прогестерона Секреторная трансформация эндометрия; Миорелаксирующий эффект; Пролиферация альвеолярного эпителия; Антиминералокортикоидное (диуретическое) действие; Антиэстрогенное действие.
- 51. Органы-мишени
- 52. Органы-мишени К органам и тканям-мишеням стероидов относятся гипоталамус, гипофиз, половые органы, молочные железы. К непродуктивным органам-мишеням
- 53. В гормонально-чувствительной клетке стероид связывается специфическим белком-рецептором, обладающим высокой степенью родства к этому гормону и подобным
- 54. Таким образом, функциональное состояние репродуктивной системы определяется обратной афферентацией ее подсистем. В ней выделяют: длинную петлю
- 57. В регуляции функции РС основным является цирхоральная секреция и выделение ГнРГ гипоталамусом и регуляция синтеза и
- 64. Скачать презентацию

Репродуктивная система (РС)
РС выполняет множество функций, наиболее важной из которых является
Репродуктивная система (РС)
РС выполняет множество функций, наиболее важной из которых является

Основой регуляции функции РС является принцип отрицательной обратной связи между различными
Основой регуляции функции РС является принцип отрицательной обратной связи между различными
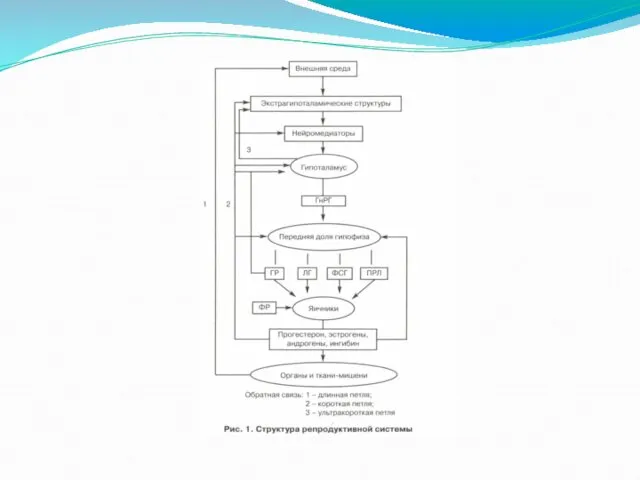


РС состоит из 5 уровней:
Экстрагипоталамического
Гипоталамуса
Гипофиза
Яичников
Органов-мишеней
Как и все системы в организме, РС
РС состоит из 5 уровней:
Экстрагипоталамического
Гипоталамуса
Гипофиза
Яичников
Органов-мишеней
Как и все системы в организме, РС

Основная роль в регуляции функции РС принадлежит гипоталамо-гипофизарной системе, которая координирует
Основная роль в регуляции функции РС принадлежит гипоталамо-гипофизарной системе, которая координирует

ДА, НА и серотонину принадлежит ведущая роль в контроле гипоталамической секреции
ДА, НА и серотонину принадлежит ведущая роль в контроле гипоталамической секреции

В последние годы установлено, что в структурах мозга синтезируются стероиды, в
В последние годы установлено, что в структурах мозга синтезируются стероиды, в

Гипоталамус
Высший вегетативный центр, гибрид нервной и эндокринной систем, координирующий функции всех
Гипоталамус
Высший вегетативный центр, гибрид нервной и эндокринной систем, координирующий функции всех

Секреция ГнРГ генетически запрограммирована, происходит в определенном пульсирующем режиме, примерно 1
Секреция ГнРГ генетически запрограммирована, происходит в определенном пульсирующем режиме, примерно 1

Гипофиз
Гипофиз – место синтеза и выделения всех тропных гормонов, непосредственной регулирующих
Гипофиз
Гипофиз – место синтеза и выделения всех тропных гормонов, непосредственной регулирующих

Биологическая роль ФСГ
Рост фолликулов в яичниках
Пролиферация клеток гранулезы в фоллликулах
Синтез ароматаз,
Биологическая роль ФСГ
Рост фолликулов в яичниках
Пролиферация клеток гранулезы в фоллликулах
Синтез ароматаз,

Биологическая роль ЛГ:
Синтез андрогенов в клетках тека фолликулов
Синтез эстрадиола в доминантном
Биологическая роль ЛГ:
Синтез андрогенов в клетках тека фолликулов
Синтез эстрадиола в доминантном

Пролактин (ПРЛ) синтезируется клетками аденогипофиза (лактотрофами), период его полураспада составляет 50-60
Пролактин (ПРЛ) синтезируется клетками аденогипофиза (лактотрофами), период его полураспада составляет 50-60

Таким образом, синтез гонадотропинов контролируется гипоталамическим ГнРГ и периферическими овариальными стероидами
Таким образом, синтез гонадотропинов контролируется гипоталамическим ГнРГ и периферическими овариальными стероидами

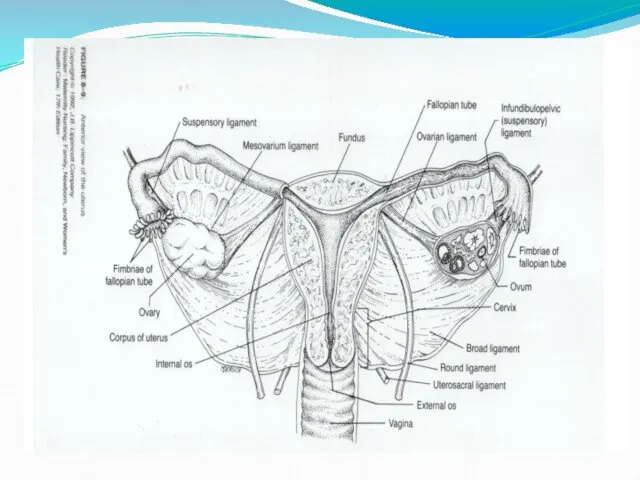
Яичники
Яичники являются местом синтеза половых стероидов: эстрогенов, андрогенов и прогестерона в
Яичники
Яичники являются местом синтеза половых стероидов: эстрогенов, андрогенов и прогестерона в
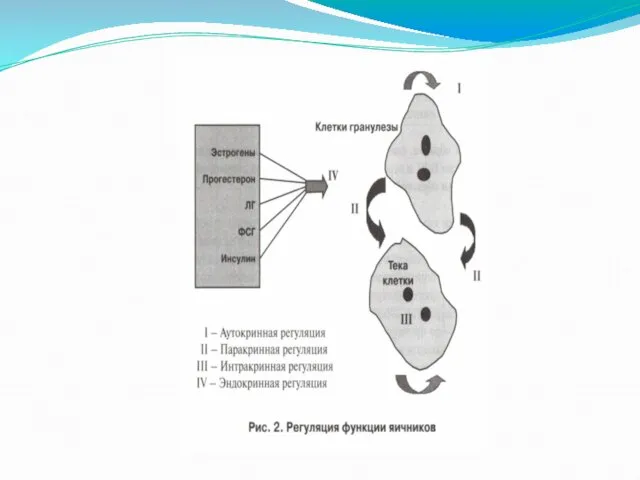

Факторы роста (ФР)
ФР – биологически активные вещества, стимулирующие или ингибирующие дифференцировку
Факторы роста (ФР)
ФР – биологически активные вещества, стимулирующие или ингибирующие дифференцировку

Наиболее важную роль в физиологии РС играют следующие ФР:
Инсулиноподобные факторы роста
Наиболее важную роль в физиологии РС играют следующие ФР:
Инсулиноподобные факторы роста

Ингибины – белковые вещества, образуются в клетках гранулезы фолликула, участвуют в
Ингибины – белковые вещества, образуются в клетках гранулезы фолликула, участвуют в

Активины обнаружены в гранулезных клетках фолликула и гонадотрофах гипофиза.
Активины …
стимулируют синтез
Активины обнаружены в гранулезных клетках фолликула и гонадотрофах гипофиза.
Активины …
стимулируют синтез

Инсулиноподобные факторы роста I и II (ИФР-I и ИФР-II) синтезируются в
Инсулиноподобные факторы роста I и II (ИФР-I и ИФР-II) синтезируются в

СЭФР играет важную роль в ангиогенезе растущих фолликулов, а в доминантном

.
Эпидермальный вактор роста (ЭФР) – обнаружен в клетках гранулезы, строме эндометрия,
.
Эпидермальный вактор роста (ЭФР) – обнаружен в клетках гранулезы, строме эндометрия,

Трансформирующий фактор роста ТФР-α и ТФР-β) стимулируют клеточную пролиферацию, оказывают митогенный
Трансформирующий фактор роста ТФР-α и ТФР-β) стимулируют клеточную пролиферацию, оказывают митогенный

Фолликулогенез в яичниках
В яичнике женщине репродуктивного возраста фолликулы находятся на различных
Фолликулогенез в яичниках
В яичнике женщине репродуктивного возраста фолликулы находятся на различных

Примордиальные фолликулы характеризуются одним слоем плоских прегранулезных клеток, небольшим ооцитом, клетки
Примордиальные фолликулы характеризуются одним слоем плоских прегранулезных клеток, небольшим ооцитом, клетки

Стадии роста фолликулов
От примордиальных до преполостных фолликулов негормональнозависимый рост.
Он продолжается до
Стадии роста фолликулов
От примордиальных до преполостных фолликулов негормональнозависимый рост.
Он продолжается до

Рост фолликула от покоящегося примордиального до преовуляторного носит последовательный и непрерывный
Рост фолликула от покоящегося примордиального до преовуляторного носит последовательный и непрерывный
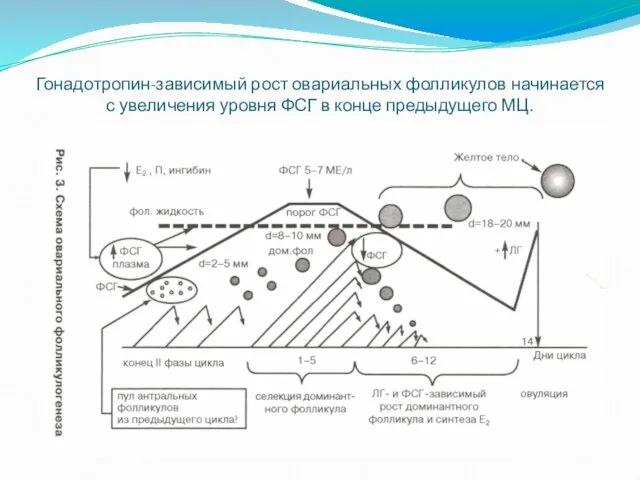
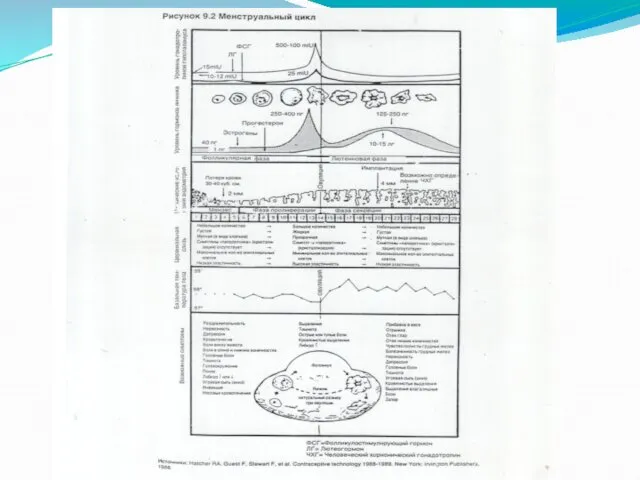
Гонадотропин-зависимый рост овариальных фолликулов начинается с увеличения уровня ФСГ в конце
Гонадотропин-зависимый рост овариальных фолликулов начинается с увеличения уровня ФСГ в конце

Гонадотропин-зависимый рост овариальных фолликулов начинается с увеличения уровня ФСГ в конце
Гонадотропин-зависимый рост овариальных фолликулов начинается с увеличения уровня ФСГ в конце

Особое значение имеет базальный уровень ФСГ на 2-3 д.м.ц. Этот показатель
Особое значение имеет базальный уровень ФСГ на 2-3 д.м.ц. Этот показатель

В ранней фолликулярной фаз до 5-го дня цикла рост фолликулов зависит
В ранней фолликулярной фаз до 5-го дня цикла рост фолликулов зависит

Биологическая роль ИФР в созревании фолликула заключается в стимуляции клеток гранулезы
Биологическая роль ИФР в созревании фолликула заключается в стимуляции клеток гранулезы

МАХ значения уровень ФСГ достигает к 5 д.м.ц, после чего снижается,
МАХ значения уровень ФСГ достигает к 5 д.м.ц, после чего снижается,

Девиация – (с англ. Deviation – отклонение) способность роста в условиях
Девиация – (с англ. Deviation – отклонение) способность роста в условиях

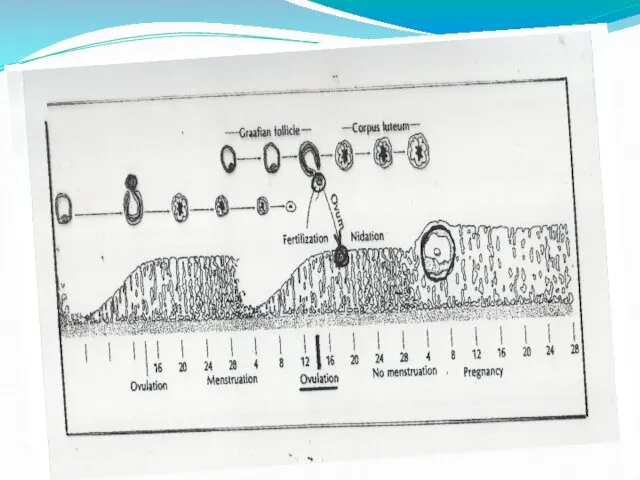
Овуляция
Процесс овуляции происходит при достижении максимального уровня эстрадиола в преовуляторном фолликуле
Овуляция
Процесс овуляции происходит при достижении максимального уровня эстрадиола в преовуляторном фолликуле

После овуляции клетки гранулезы подвергаются дальнейшей лютеинизации с образованием ЖТ, секретирующего
После овуляции клетки гранулезы подвергаются дальнейшей лютеинизации с образованием ЖТ, секретирующего

Основным регулятором синтеза стероидов в ЖТ является ЛГ.
Снижение активности ЖТ
Основным регулятором синтеза стероидов в ЖТ является ЛГ.
Снижение активности ЖТ

Биосинтез стероидов в яичниках – сложный процесс.
Он происходит в соответствии с
Биосинтез стероидов в яичниках – сложный процесс.
Он происходит в соответствии с

Первые этапы синтеза стероидов – до прегненолона.
Энзимные системы в стероидогенезе, идентичны
Первые этапы синтеза стероидов – до прегненолона.
Энзимные системы в стероидогенезе, идентичны

Первые этапы синтеза овариальных стероидов детерминированы гонадтропинами.
Под влиянием ЛГ, рецепторы
Первые этапы синтеза овариальных стероидов детерминированы гонадтропинами.
Под влиянием ЛГ, рецепторы
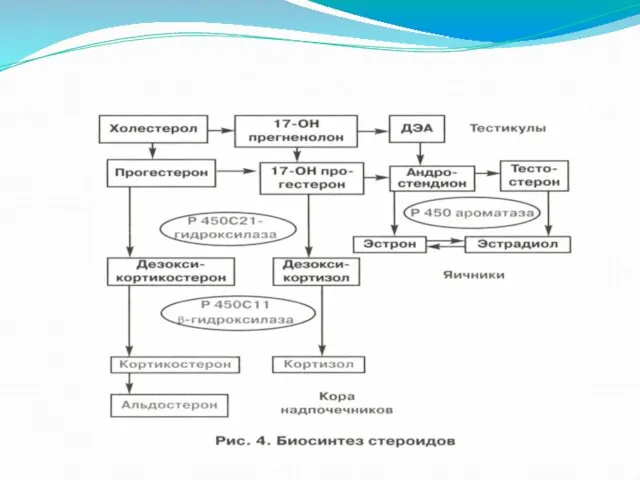


Максимальные концентрации прогестерона синтезируются после лютеинизации клеток гранулезы при функционировании ЖТ.
Максимальные концентрации прогестерона синтезируются после лютеинизации клеток гранулезы при функционировании ЖТ.

Важная роль в гормональном балансе отводится половым стероид-связывающим глобулинам (ПССГ), синтез
Важная роль в гормональном балансе отводится половым стероид-связывающим глобулинам (ПССГ), синтез

Биологическое действие эстрогенов I
На репродуктивные органы:
Пролиферация и гиперплазия эндо- и миометрия,
Биологическое действие эстрогенов I
На репродуктивные органы:
Пролиферация и гиперплазия эндо- и миометрия,

Биологическое действие эстрогенов II
На экстрагенитальную систему:
Пролиферативные процессы слизистой уретры, мочевого пузыря;
Развитие
Биологическое действие эстрогенов II
На экстрагенитальную систему:
Пролиферативные процессы слизистой уретры, мочевого пузыря;
Развитие

Биологическое действие прогестерона
Секреторная трансформация эндометрия;
Миорелаксирующий эффект;
Пролиферация альвеолярного эпителия;
Антиминералокортикоидное (диуретическое) действие;
Антиэстрогенное действие.
Биологическое действие прогестерона
Секреторная трансформация эндометрия;
Миорелаксирующий эффект;
Пролиферация альвеолярного эпителия;
Антиминералокортикоидное (диуретическое) действие;
Антиэстрогенное действие.
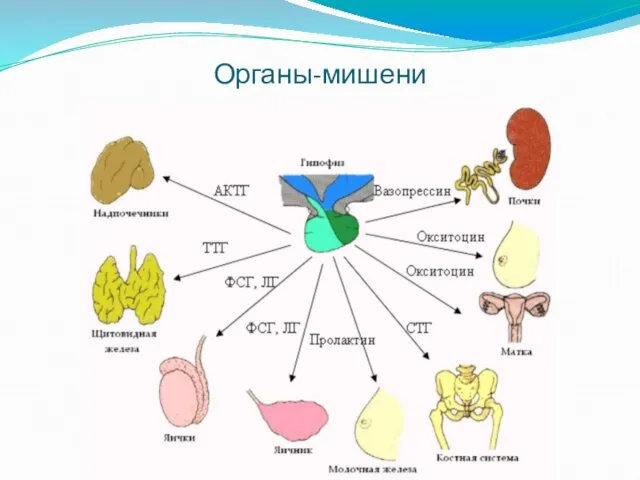
Органы-мишени
Органы-мишени

Органы-мишени
К органам и тканям-мишеням стероидов относятся гипоталамус, гипофиз, половые органы, молочные
Органы-мишени
К органам и тканям-мишеням стероидов относятся гипоталамус, гипофиз, половые органы, молочные

В гормонально-чувствительной клетке стероид связывается специфическим белком-рецептором, обладающим высокой степенью родства
В гормонально-чувствительной клетке стероид связывается специфическим белком-рецептором, обладающим высокой степенью родства

Таким образом, функциональное состояние репродуктивной системы определяется обратной афферентацией ее подсистем.
В
Таким образом, функциональное состояние репродуктивной системы определяется обратной афферентацией ее подсистем.
В

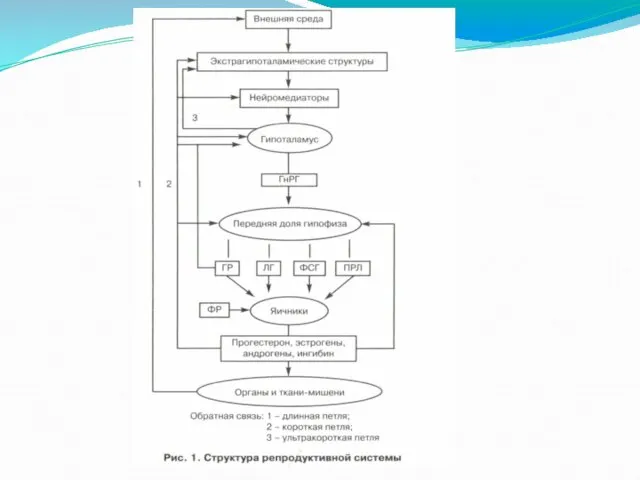

В регуляции функции РС основным является цирхоральная секреция и выделение ГнРГ
В регуляции функции РС основным является цирхоральная секреция и выделение ГнРГ

 Ранний послеоперационный период
Ранний послеоперационный период Соціальна медицина та організація охорони здоров’я як наука. Предмет, методи, значення для практики охорони здоров’я
Соціальна медицина та організація охорони здоров’я як наука. Предмет, методи, значення для практики охорони здоров’я Физиологические механизмы регуляции в организме
Физиологические механизмы регуляции в организме Герпетическая инфекция
Герпетическая инфекция Аналық бездің қатерлі ісігі
Аналық бездің қатерлі ісігі Обследование альвеолярного отростка верхней и нижней челюсти
Обследование альвеолярного отростка верхней и нижней челюсти Одонтогенные новообразования челюстей у детей (амелобластома, одонтома, цементома)
Одонтогенные новообразования челюстей у детей (амелобластома, одонтома, цементома) Малярия, токсоплазма, лямблии
Малярия, токсоплазма, лямблии Балалар жақ сүйегінің периоститі
Балалар жақ сүйегінің периоститі Патологическая стираемость твердых тканей зубов
Патологическая стираемость твердых тканей зубов Балаларда туа пайда болған жүрек ақаулары
Балаларда туа пайда болған жүрек ақаулары Скажи наркотикам нет
Скажи наркотикам нет Оптимизация обучения студентов медицинского университета
Оптимизация обучения студентов медицинского университета Жасқа байланысты,асқынбаған тіс жегісінде,жүйелі аурулар мен зат алмасу ауруларында ұлпадағы өзгерістер
Жасқа байланысты,асқынбаған тіс жегісінде,жүйелі аурулар мен зат алмасу ауруларында ұлпадағы өзгерістер Предварительные и периодические медицинские осмотры работников
Предварительные и периодические медицинские осмотры работников Применения Ботулинотоксин-А при лечении невралгии тройничного нерва
Применения Ботулинотоксин-А при лечении невралгии тройничного нерва Әйел жыныс мүшелерінің қабыну аурулары
Әйел жыныс мүшелерінің қабыну аурулары Метаболизм липидов
Метаболизм липидов Рак поджелудочной железы
Рак поджелудочной железы Учение о диагнозе
Учение о диагнозе Введение в сексологию
Введение в сексологию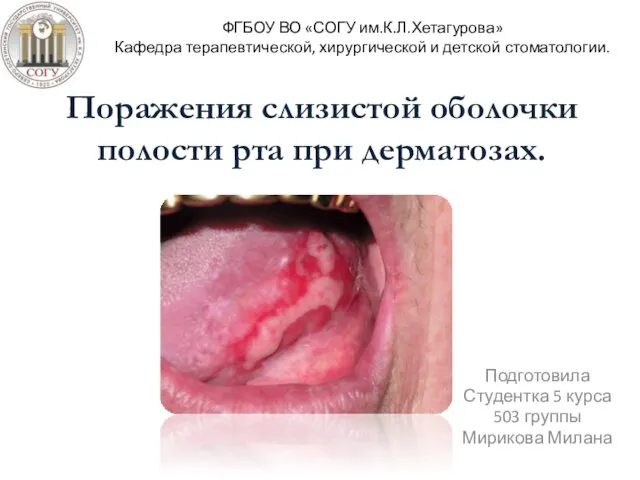 Поражения слизистой оболочки полости рта при дерматозах
Поражения слизистой оболочки полости рта при дерматозах Биохимия слюны
Биохимия слюны Профилактика наркомании и формирование установок на ведение здорового образа жизни среди молодежи
Профилактика наркомании и формирование установок на ведение здорового образа жизни среди молодежи Актуальные вопросы проведения химиопрофилактики перинатальной передачи ВИЧ от матери ребенку в Алтайском крае
Актуальные вопросы проведения химиопрофилактики перинатальной передачи ВИЧ от матери ребенку в Алтайском крае Диагностика раннего рака желудка
Диагностика раннего рака желудка Modal Verbs Lobular Pneumonia
Modal Verbs Lobular Pneumonia Аутоиммунный гепатит. Определение. Классификация
Аутоиммунный гепатит. Определение. Классификация