- Биоокисление. Метаболизм углеводов. Тема 6

Содержание
- 2. Предложена Питером Митчелом в 1960 Нобелевская премия, 1978. Хемиосмотическая теория Хемиосмотическая теория: транспорт электронов и синтез
- 3. Необходима интактная митохондриальная мембрана Транспорт электронов через ЭТЦ генерирует протонный градиент 3. AТФ синтаза катализирует фосфорилирование
- 4. АТФ синтаза Две субъединицы, Fo и F1 F1 содержит каталитические субъединицы, где АДФ и Pи связываются.
- 5. АТФ должен транспортироваться в цитозоль, а АДФ и Pи - в матрикс AДФ/ATФ переносчик меняет митохондриальное
- 6. РЕГУЛЯЦИЯ ОКИСЛИТЕЛЬНОГО ФОСФОРИЛИРОВАНИЯ Сопряжение тканевого дихания с окислительным фосфорилирование Транспорт электронов тесно связан с фосфорилированием. АТФ
- 7. Регуляция скорости окислительного фосфорилювания с помощью уровня АДФ называется дыхательным контролем. Дыхательный контроль
- 8. Разобщение тканевого дыхания и окислительного фосфорилирования Внутренняя митохондриальная мембрана содержит разобщительный белок (Термогенин, uncoupling protein). Разобщительный
- 10. Разобщающие протеины являются трансмембранными белками, которые уменьшают градиент протонов в окислительном фосфорилировании. Они увеличивают проницаемость внутренней
- 11. Разобщители являются жирорастворимыми слабыми кислотами Разобщители снижают протонный градиент транспортируя протоны через мембрану Разобщители 2,4-Динитрофенол –
- 12. Перенесение 3H+ необходимо для синтеза одной молекулы АТФ АТФ-синтазой 1 H+ необходимый для транспорта Pи. 4
- 13. МЕТАБОЛИЗМ УГЛЕВОДОВ - 1 На протяжении столетий пекари и пивовары используют превращение глюкозы к этанолу и
- 14. Гликолиз наиболее ранее открытый и наиболее важный процесс метаболизма углеводов Гликолиз – метаболический путь, в котором
- 15. 10 реакций гликолиза 1. Гексокиназа Переносит фосфорную группу с АТФ к глюкозе с образованием глюкозо-6-фосфата. Четыре
- 16. Превращает глюкозо-6-фосфат в фруктозо-6-фосфат. 2. Глюкозо-6-фосфат изомераза
- 17. Катализирует перенесение фосфорной группы с АТФ на фруктозо-6-фосфат с образованием фруктозо-1,6-дифосфата. 3. Фосфофруктокиназа-1 (ФФК-1)
- 18. 4. Альдолаза Расщепляет гексозу - фруктозо-1,6-дифосфат на две триозы: глицеральдегид-3-фосфат и дигидроксиацетонфосфат.
- 19. Превращает ДГАФ в ГАФ Реакця обратимая. При Равновесии 96% ДГАФ и 4 % ГАФ. 5. Триозофосфат
- 20. Превращение ГАФ в 1,3-дифосфоглицерат Молекула НАД+ восстанавливается в НАДН 6. Глицеральдегид 3-фосфат дегидрогеназа
- 21. Переносит фосфатную группу с 1,3-дифосфоглицерата на АДФ с образованием ATФ и 3-фосфоглицерата - субстратное фосфорилирование 7.
- 22. 8. Фосфоглицератмутаза Катализирует перенесение фосфатной группы с одной части молекулы к другой.
- 23. 9. Энолаза 2-Фосфоглицерат превращается в фосфоэнолпируват Фосфоэнолпируват имеет высокий фосфорил-трансферный потенциал
- 24. 10. Пируваткиназа Катализирует реакцию субстратного фосфорилирования: Необратимая реакция Регуляторная реакция
- 25. Общая реакция гликолиза Две молекулы АТФ вырабатываются Две молекулы НАД восстанавливаются с образованием НАДН Глюкоза+ 2
- 26. Научные исследования ферментации виноградного сахара были первыми исследованиями гликолиза
- 27. Судьба пирувата
- 28. Метаболизм пирувата в этанол Этанол образуется из пирувата в дрожжах и некоторых микроорганизмах в анаэробных условиях.
- 29. Лактат образуется из пирувата в анаэробных условиях. Превращение глюкозы в лактат называется молочнокислым брожением. Фермент -
- 30. Глюкозо-1-фосфат превращается в глюкозо-6-фосфат фосфоглюкомутазой.
- 31. Непереносимость лактозы (гиполактазия) обусловлена дефицитом лактазы, которая расщепляет лактозу к глюкозе и галактозе. Микроорганизмы в толстом
- 32. Галактоземия Нарушение метаболизма галактозы - галактоземия. Классическая галактоземия - это врожденный дефицит галактозо-1-фосфат уридилтрансферазы. Симптомы: -
- 33. Эффект Пастера Больше АТФ образуется в аэробных условиях, чем в анаэробных, потому в аэробных условиях используется
- 34. Глюкоза Судьба глюкозы в клетке Глюкозо-6-фосфат Пируват Гликоген рибоза, НАДФН Пентозофосфат-ный путь Синтез гликогена Деградация гликогена
- 35. Все клетки зависят от глюкозы. Мозг особенно чувствителен к снижению уровня глюкозы (дневная потребность глюкозы для
- 36. Печень и почки – основные органы синтеза глюкозы Основные предшественники: лактат, пируват, глицерол и некоторые аминокислоты
- 37. Глюконеогенез не является обратимым гликолизом В гликолизе глюкоза превращается в пируват; в глюконеогенезе пируват превращается в
- 38. Bypass I: Пируват → Фосфоэнолпируват Первый шаг в глюконеогенезе - карбоксилирование пирувата в оксалоацетат. Фермент пируваткарбоксилаза
- 39. Проходит в цитозоле. Одна молекула АТФ и одна молекула ГТФ используются для превращения пирувата в фосфоенолпируват.
- 40. Фермент фруктозо-1,6-дифосфатаза Bypass II: Фруктозо-1,6-дифосфат → фруктозо-6-фосфат
- 41. Bypass III: Глюкозо-6-фосфат → глюкоза Глюкозо-6-фосфат не может дифундировать из клетки. Образование свободной глюкозы регулируется двумя
- 42. Последняя реакция не осуществляется в цитозоле. Г-6-Ф транспортируется в эндоплазматическую сеть, где гидролизуется глюкозо-6-фосфатазой, которая связана
- 43. Эндоплазмати́ческий рети́кулум (ЭПР) (лат. reticulum — сеточка) или эндоплазматическая сеть (ЭПС) — внутриклеточный органоид эукариотической клетки,
- 44. Скорость гликолиза определяется кoнцентрацией глюкозы. Скорост глюконеогенеза определяется кoнцентрацией предшественников глюкозы. Регуляция глюконеогенеза Глюконеогенез и гликолиз
- 45. Гормоны влияют на экспрессию генов изменяя скорость транскрипции. Инсулин стимулирует экспрессию фосфофруктокиназы и пируваткиназы. Глюкагон ингибирует
- 46. Основные предшественники: (1) Лактат (2) Большинство аминокислот (особенно аланин), (3) Глицерол (при расщеплении жиров) Предшественники глюконеогенеза
- 47. Цикл Кори Печеночная лактатдегидрогеназа превращает лактат в пируват, субстрат для глюконеогенеза. Глюкоза, образованная в печени, транспортируется
- 48. Пентозофосфатный путь
- 49. (1) Синтез НАДФН (для биосинтеза жирных кислот и стероидов) (2) Синтез рибозо-5-фосфата (для биосинтеза ДНК и
- 51. Скачать презентацию

Предложена Питером Митчелом в 1960
Нобелевская премия, 1978.
Хемиосмотическая теория
Хемиосмотическая теория: транспорт
Предложена Питером Митчелом в 1960
Нобелевская премия, 1978.
Хемиосмотическая теория
Хемиосмотическая теория: транспорт
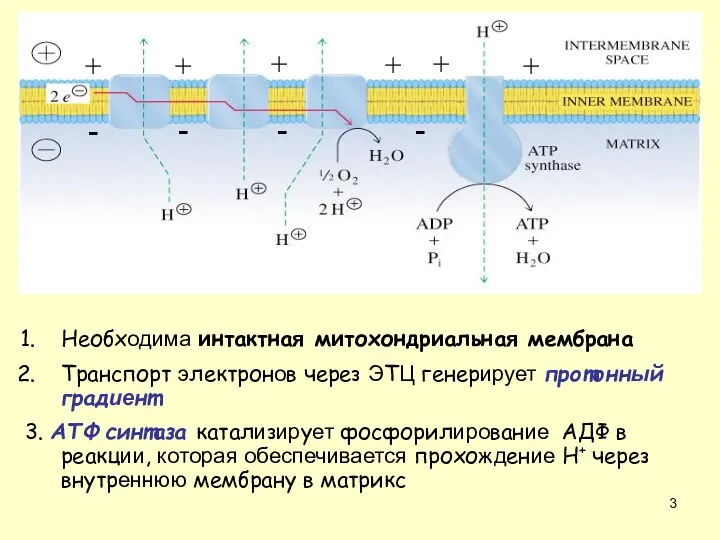
Необходима интактная митохондриальная мембрана
Транспорт электронов через ЭТЦ генерирует протонный градиент
3. AТФ
Необходима интактная митохондриальная мембрана
Транспорт электронов через ЭТЦ генерирует протонный градиент
3. AТФ
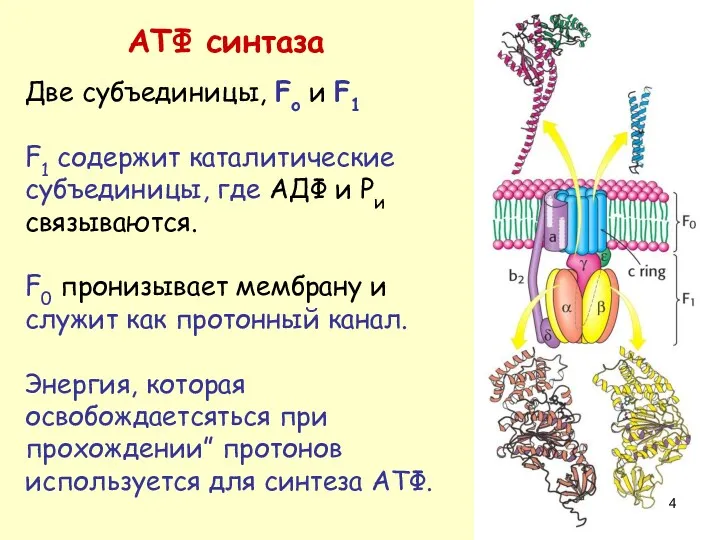
АТФ синтаза
Две субъединицы, Fo и F1
F1 содержит каталитические субъединицы, где
АТФ синтаза
Две субъединицы, Fo и F1
F1 содержит каталитические субъединицы, где
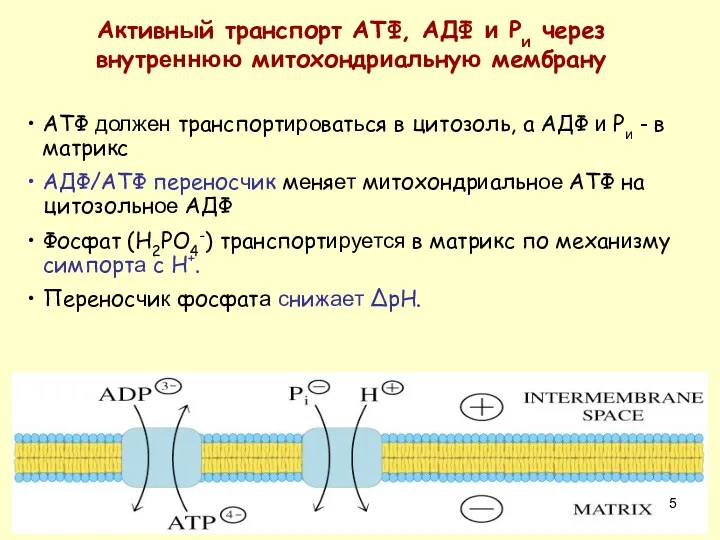
АТФ должен транспортироваться в цитозоль, а АДФ и Pи - в
АТФ должен транспортироваться в цитозоль, а АДФ и Pи - в

РЕГУЛЯЦИЯ ОКИСЛИТЕЛЬНОГО ФОСФОРИЛИРОВАНИЯ
Сопряжение тканевого дихания с окислительным фосфорилирование
Транспорт электронов тесно связан
РЕГУЛЯЦИЯ ОКИСЛИТЕЛЬНОГО ФОСФОРИЛИРОВАНИЯ
Сопряжение тканевого дихания с окислительным фосфорилирование
Транспорт электронов тесно связан
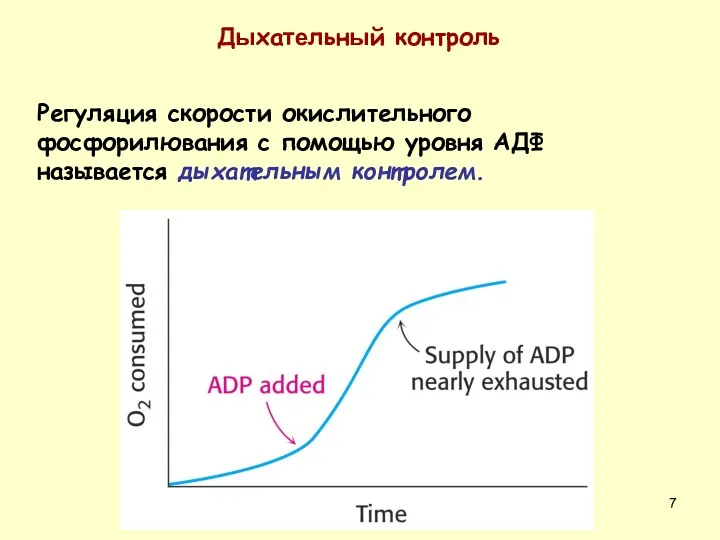
Регуляция скорости окислительного фосфорилювания с помощью уровня АДФ называется дыхательным контролем.
Дыхательный
Регуляция скорости окислительного фосфорилювания с помощью уровня АДФ называется дыхательным контролем.
Дыхательный
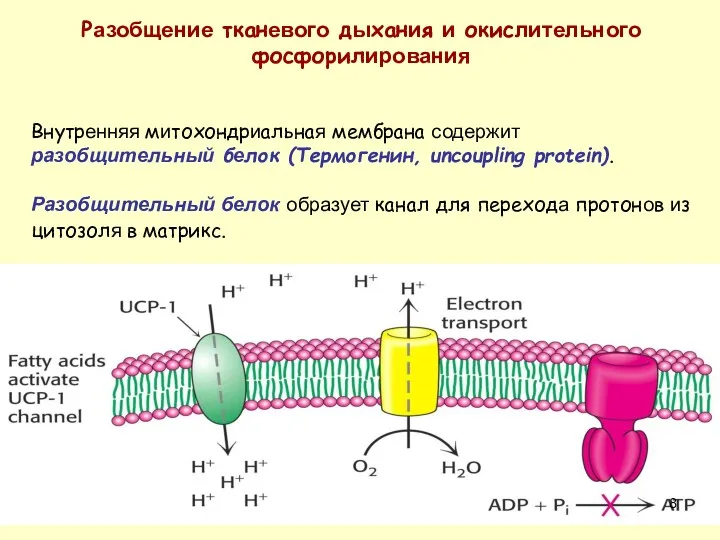
Разобщение тканевого дыхания и окислительного фосфорилирования
Внутренняя митохондриальная мембрана содержит разобщительный белок
Разобщение тканевого дыхания и окислительного фосфорилирования
Внутренняя митохондриальная мембрана содержит разобщительный белок
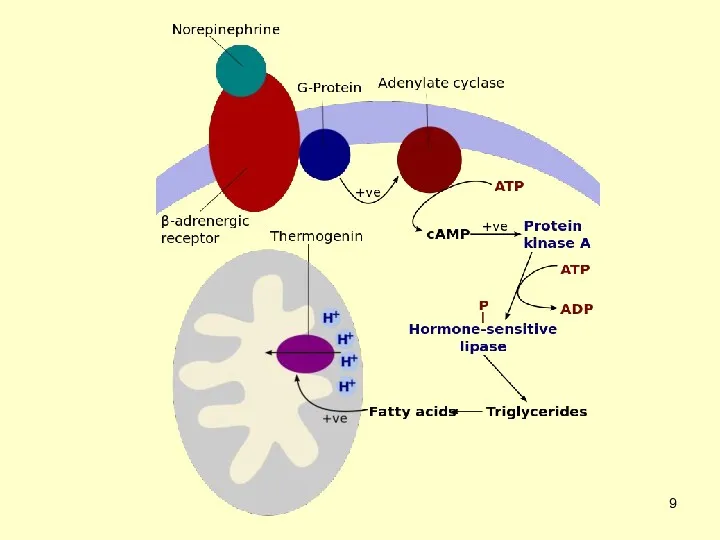

Разобщающие протеины являются трансмембранными белками, которые уменьшают градиент протонов в окислительном
Разобщающие протеины являются трансмембранными белками, которые уменьшают градиент протонов в окислительном
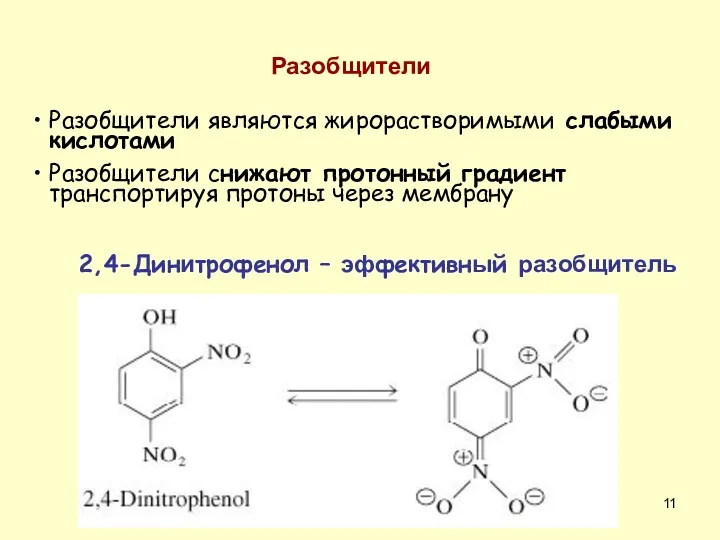
Разобщители являются жирорастворимыми слабыми кислотами
Разобщители снижают протонный градиент транспортируя протоны через
Разобщители являются жирорастворимыми слабыми кислотами
Разобщители снижают протонный градиент транспортируя протоны через
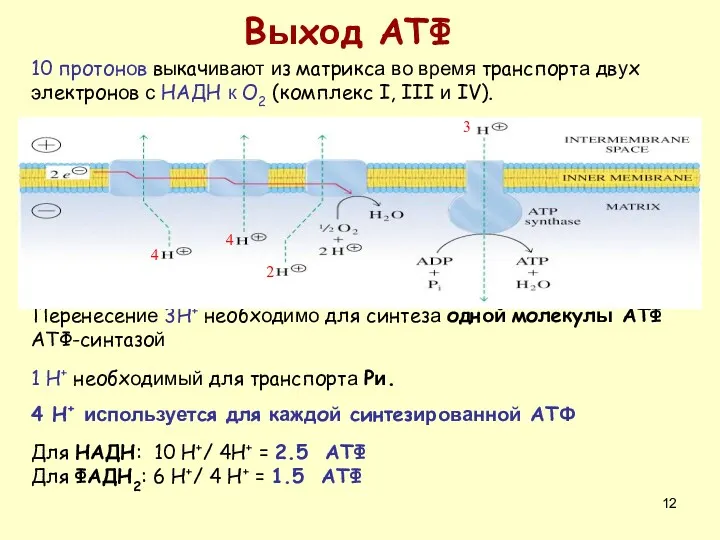
Перенесение 3H+ необходимо для синтеза одной молекулы АТФ АТФ-синтазой
1 H+ необходимый
Перенесение 3H+ необходимо для синтеза одной молекулы АТФ АТФ-синтазой
1 H+ необходимый

МЕТАБОЛИЗМ УГЛЕВОДОВ - 1
На протяжении столетий пекари и пивовары используют превращение
МЕТАБОЛИЗМ УГЛЕВОДОВ - 1
На протяжении столетий пекари и пивовары используют превращение

Гликолиз наиболее ранее открытый и наиболее важный процесс метаболизма углеводов
Гликолиз –
Гликолиз наиболее ранее открытый и наиболее важный процесс метаболизма углеводов
Гликолиз –
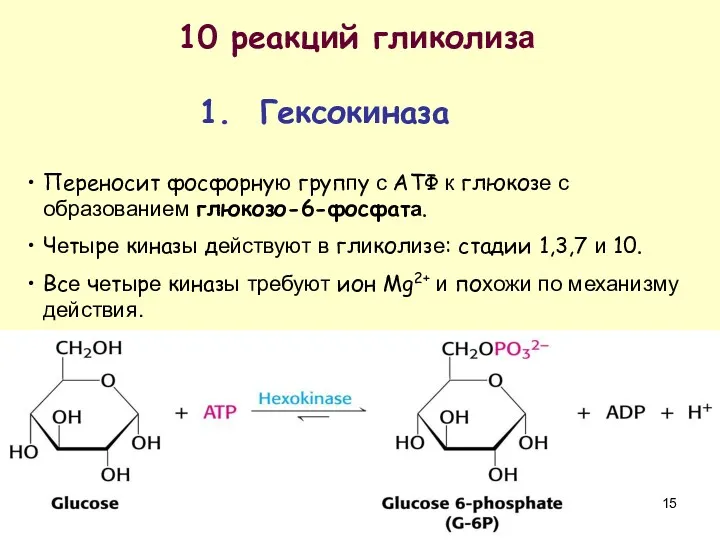
10 реакций гликолиза
1. Гексокиназа
Переносит фосфорную группу с АТФ к глюкозе с
10 реакций гликолиза
1. Гексокиназа
Переносит фосфорную группу с АТФ к глюкозе с
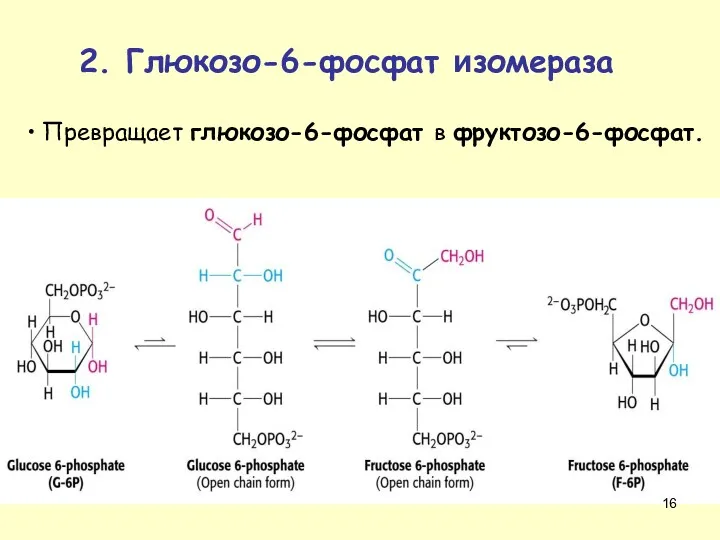
Превращает глюкозо-6-фосфат в фруктозо-6-фосфат.
2. Глюкозо-6-фосфат изомераза
Превращает глюкозо-6-фосфат в фруктозо-6-фосфат.
2. Глюкозо-6-фосфат изомераза
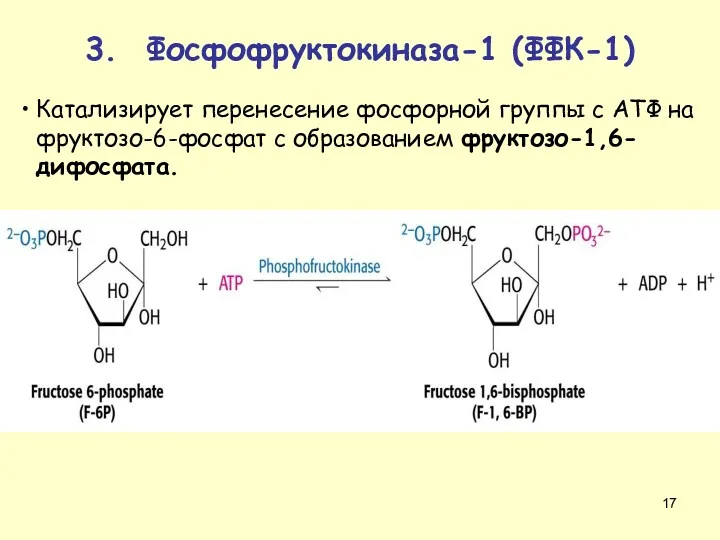
Катализирует перенесение фосфорной группы с АТФ на фруктозо-6-фосфат с образованием фруктозо-1,6-дифосфата.
3.
Катализирует перенесение фосфорной группы с АТФ на фруктозо-6-фосфат с образованием фруктозо-1,6-дифосфата.
3.
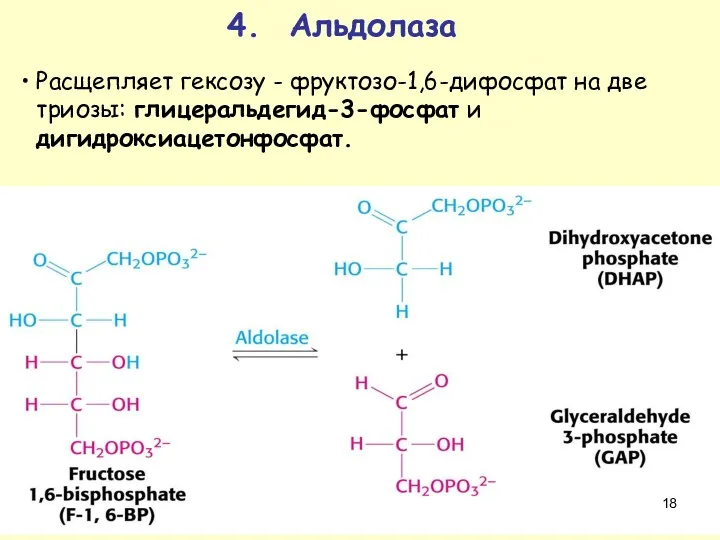
4. Альдолаза
Расщепляет гексозу - фруктозо-1,6-дифосфат на две триозы: глицеральдегид-3-фосфат и дигидроксиацетонфосфат.
4. Альдолаза
Расщепляет гексозу - фруктозо-1,6-дифосфат на две триозы: глицеральдегид-3-фосфат и дигидроксиацетонфосфат.
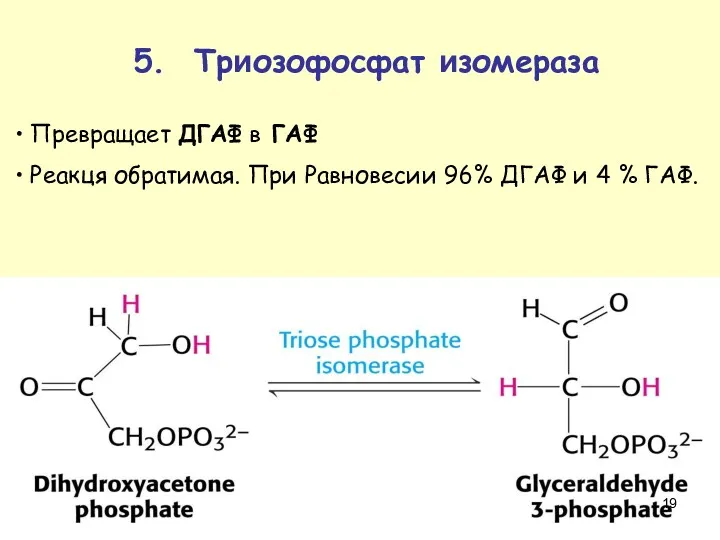
Превращает ДГАФ в ГАФ
Реакця обратимая. При Равновесии 96% ДГАФ и
Превращает ДГАФ в ГАФ
Реакця обратимая. При Равновесии 96% ДГАФ и
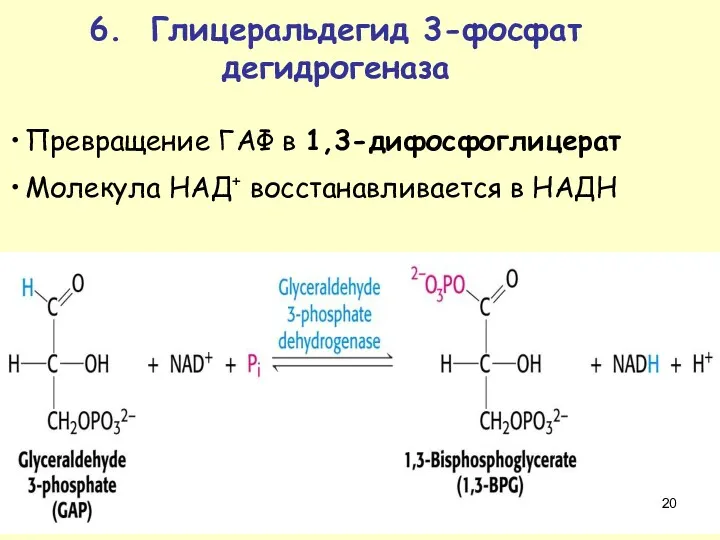
Превращение ГАФ в 1,3-дифосфоглицерат
Молекула НАД+ восстанавливается в НАДН
6. Глицеральдегид 3-фосфат дегидрогеназа
Превращение ГАФ в 1,3-дифосфоглицерат
Молекула НАД+ восстанавливается в НАДН
6. Глицеральдегид 3-фосфат дегидрогеназа
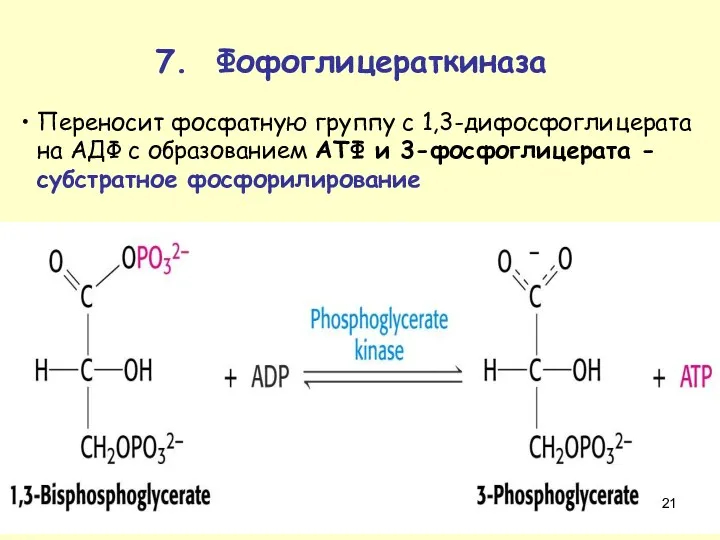
Переносит фосфатную группу с 1,3-дифосфоглицерата на АДФ с образованием ATФ и
Переносит фосфатную группу с 1,3-дифосфоглицерата на АДФ с образованием ATФ и
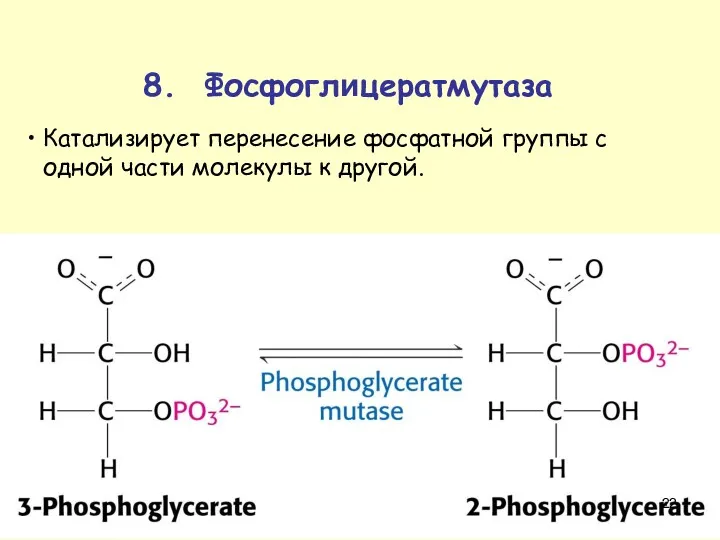
8. Фосфоглицератмутаза
Катализирует перенесение фосфатной группы с одной части молекулы к другой.
8. Фосфоглицератмутаза
Катализирует перенесение фосфатной группы с одной части молекулы к другой.
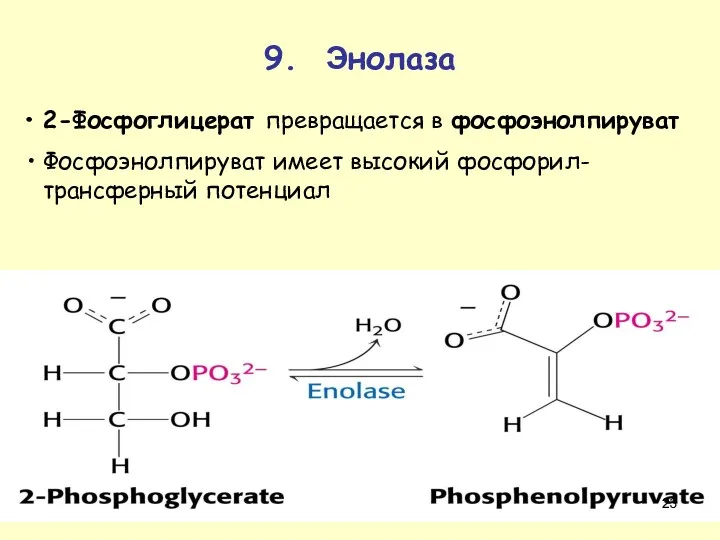
9. Энолаза
2-Фосфоглицерат превращается в фосфоэнолпируват
Фосфоэнолпируват имеет высокий фосфорил-трансферный потенциал
9. Энолаза
2-Фосфоглицерат превращается в фосфоэнолпируват
Фосфоэнолпируват имеет высокий фосфорил-трансферный потенциал
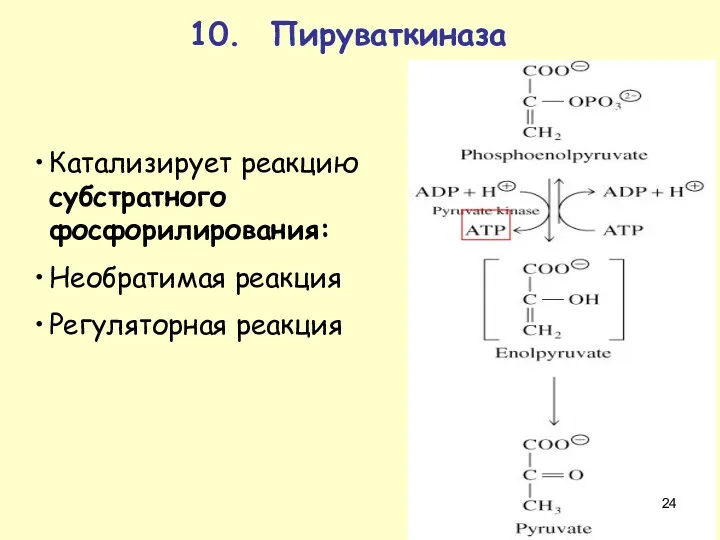
10. Пируваткиназа
Катализирует реакцию субстратного фосфорилирования:
Необратимая реакция
Регуляторная реакция
10. Пируваткиназа
Катализирует реакцию субстратного фосфорилирования:
Необратимая реакция
Регуляторная реакция

Общая реакция гликолиза
Две молекулы АТФ вырабатываются
Две молекулы НАД восстанавливаются с образованием
Общая реакция гликолиза
Две молекулы АТФ вырабатываются
Две молекулы НАД восстанавливаются с образованием

Научные исследования ферментации виноградного сахара были первыми исследованиями гликолиза
Научные исследования ферментации виноградного сахара были первыми исследованиями гликолиза
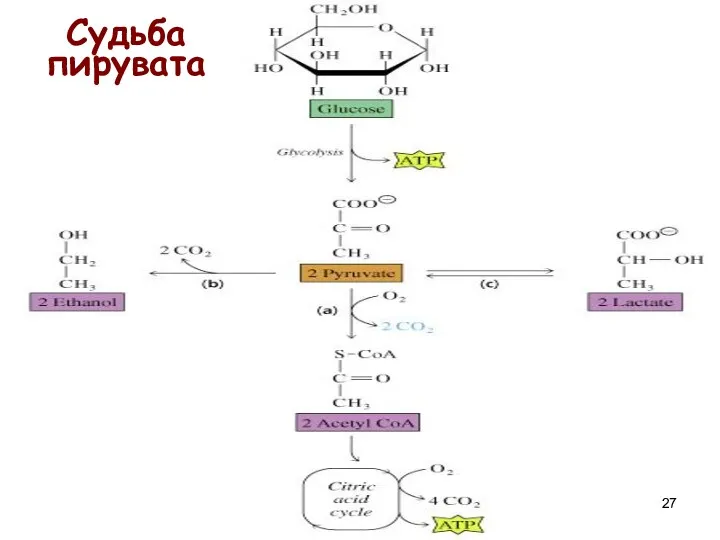
Судьба пирувата
Судьба пирувата
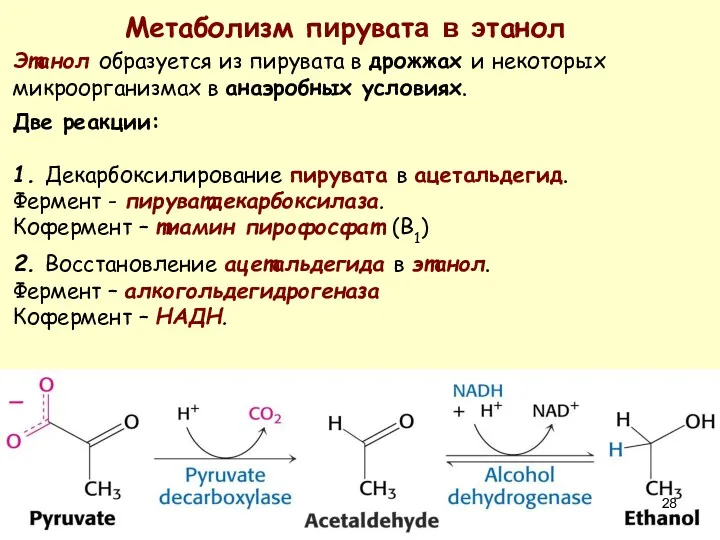
Метаболизм пирувата в этанол
Этанол образуется из пирувата в дрожжах и некоторых
Метаболизм пирувата в этанол
Этанол образуется из пирувата в дрожжах и некоторых
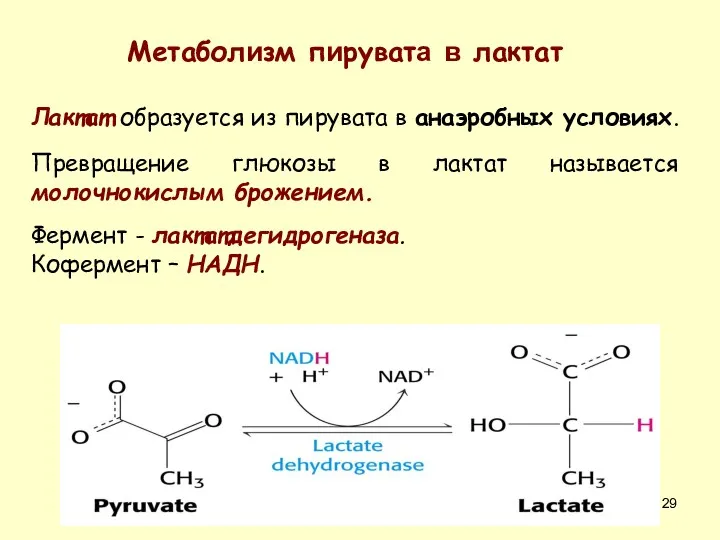
Лактат образуется из пирувата в анаэробных условиях.
Превращение глюкозы в лактат
Лактат образуется из пирувата в анаэробных условиях.
Превращение глюкозы в лактат
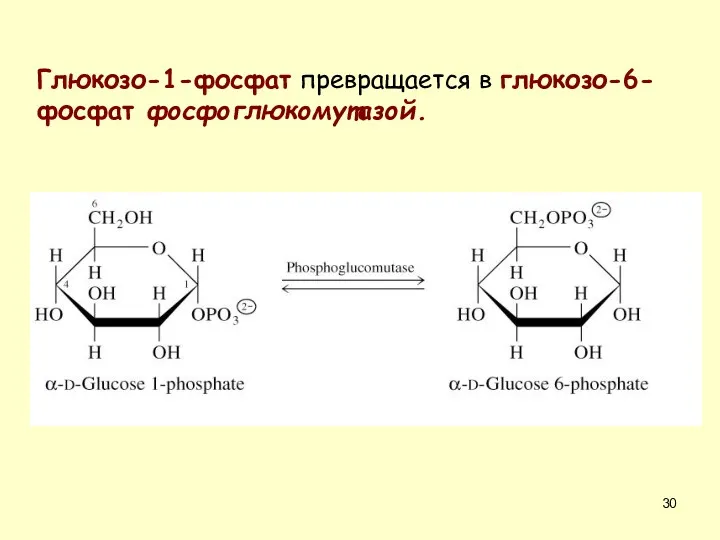
Глюкозо-1-фосфат превращается в глюкозо-6-фосфат фосфоглюкомутазой.
Глюкозо-1-фосфат превращается в глюкозо-6-фосфат фосфоглюкомутазой.

Непереносимость лактозы (гиполактазия) обусловлена дефицитом лактазы, которая расщепляет лактозу к глюкозе
Непереносимость лактозы (гиполактазия) обусловлена дефицитом лактазы, которая расщепляет лактозу к глюкозе
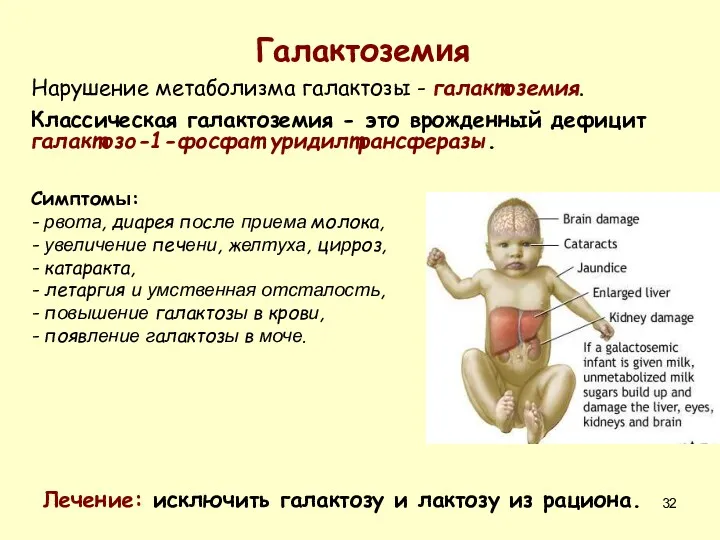
Галактоземия
Нарушение метаболизма галактозы - галактоземия.
Классическая галактоземия - это врожденный дефицит
Галактоземия
Нарушение метаболизма галактозы - галактоземия.
Классическая галактоземия - это врожденный дефицит

Эффект Пастера
Больше АТФ образуется в аэробных условиях, чем в анаэробных, потому
Эффект Пастера
Больше АТФ образуется в аэробных условиях, чем в анаэробных, потому
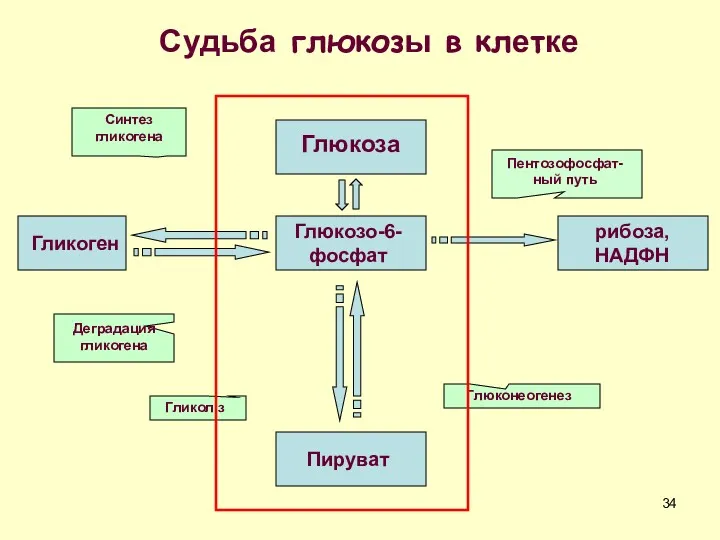
Глюкоза
Судьба глюкозы в клетке
Глюкозо-6-фосфат
Пируват
Гликоген
рибоза, НАДФН
Пентозофосфат-ный путь
Синтез гликогена
Деградация гликогена
Гликоліз
Глюконеогенез
Глюкоза
Судьба глюкозы в клетке
Глюкозо-6-фосфат
Пируват
Гликоген
рибоза, НАДФН
Пентозофосфат-ный путь
Синтез гликогена
Деградация гликогена
Гликоліз
Глюконеогенез
Все клетки зависят от глюкозы.
Мозг особенно чувствителен к снижению уровня
Все клетки зависят от глюкозы.
Мозг особенно чувствителен к снижению уровня

Печень и почки – основные органы синтеза глюкозы
Основные предшественники: лактат, пируват,
Печень и почки – основные органы синтеза глюкозы
Основные предшественники: лактат, пируват,

Глюконеогенез не является обратимым гликолизом
В гликолизе глюкоза превращается в пируват; в
Глюконеогенез не является обратимым гликолизом
В гликолизе глюкоза превращается в пируват; в
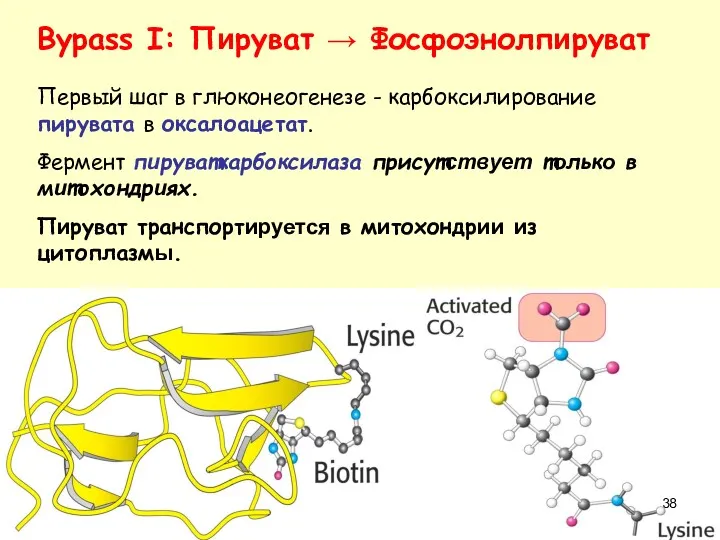
Bypass I: Пируват → Фосфоэнолпируват
Первый шаг в глюконеогенезе - карбоксилирование пирувата
Bypass I: Пируват → Фосфоэнолпируват
Первый шаг в глюконеогенезе - карбоксилирование пирувата
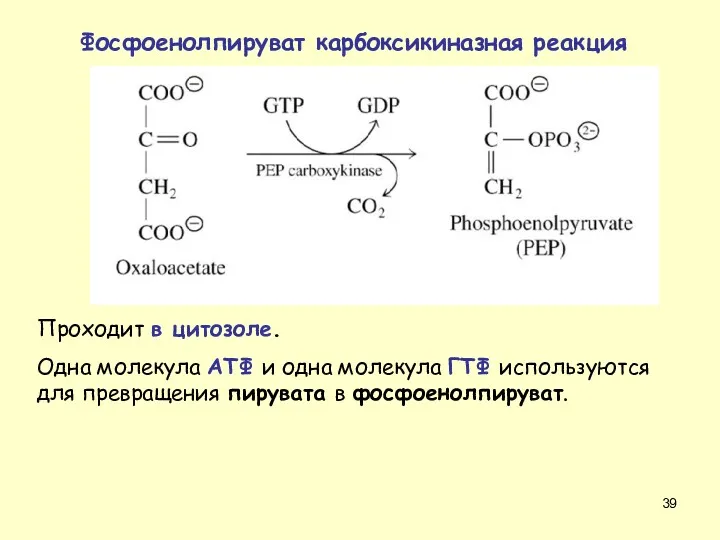
Проходит в цитозоле.
Одна молекула АТФ и одна молекула ГТФ используются для
Проходит в цитозоле.
Одна молекула АТФ и одна молекула ГТФ используются для
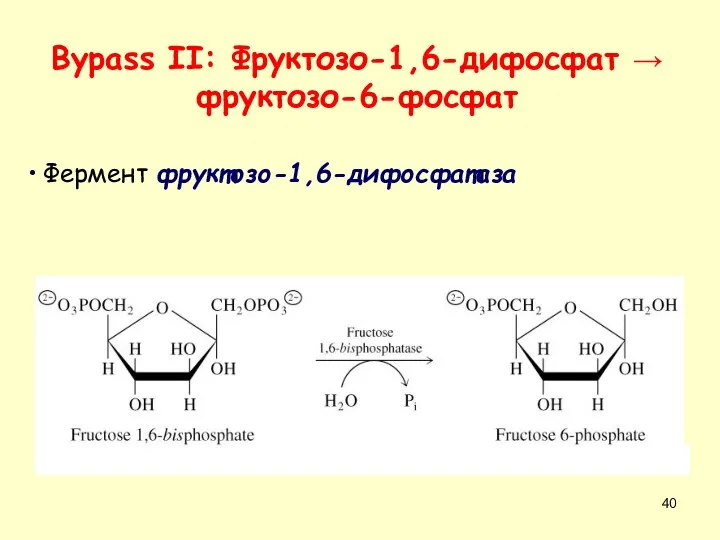
Фермент фруктозо-1,6-дифосфатаза
Bypass II: Фруктозо-1,6-дифосфат → фруктозо-6-фосфат
Фермент фруктозо-1,6-дифосфатаза
Bypass II: Фруктозо-1,6-дифосфат → фруктозо-6-фосфат

Bypass III: Глюкозо-6-фосфат → глюкоза
Глюкозо-6-фосфат не может дифундировать из клетки.
Bypass III: Глюкозо-6-фосфат → глюкоза
Глюкозо-6-фосфат не может дифундировать из клетки.
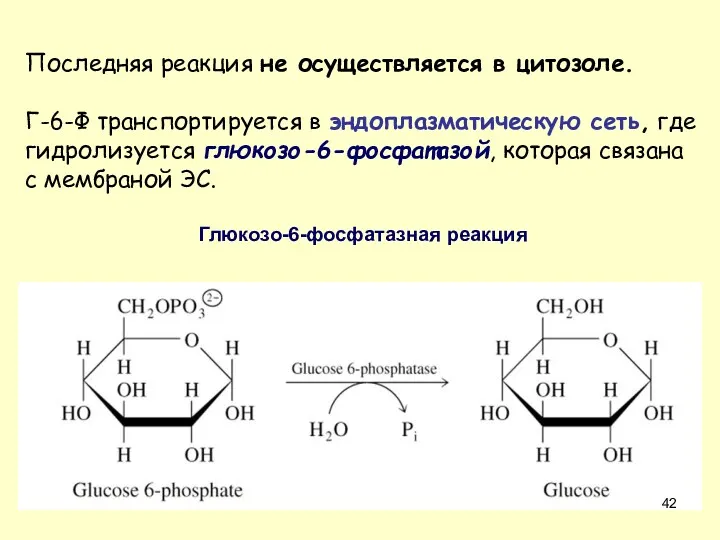
Последняя реакция не осуществляется в цитозоле.
Г-6-Ф транспортируется в эндоплазматическую сеть,
Последняя реакция не осуществляется в цитозоле.
Г-6-Ф транспортируется в эндоплазматическую сеть,
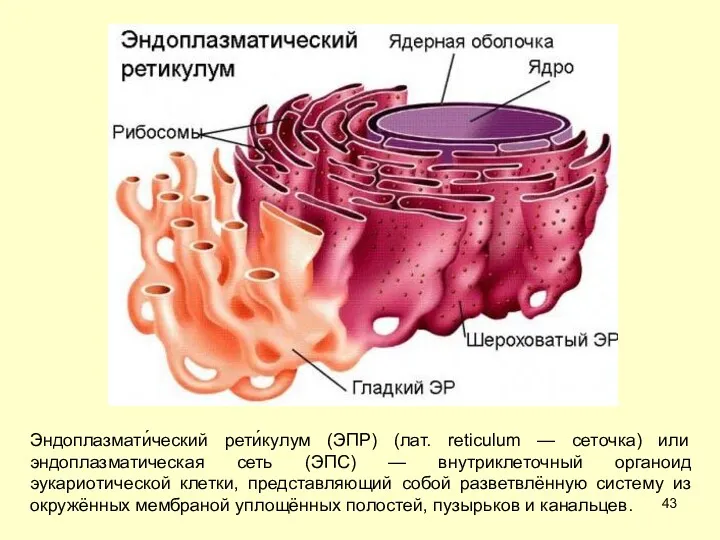
Эндоплазмати́ческий рети́кулум (ЭПР) (лат. reticulum — сеточка) или эндоплазматическая сеть (ЭПС)
Эндоплазмати́ческий рети́кулум (ЭПР) (лат. reticulum — сеточка) или эндоплазматическая сеть (ЭПС)

Скорость гликолиза определяется кoнцентрацией глюкозы.
Скорост глюконеогенеза определяется кoнцентрацией предшественников глюкозы.
Регуляция
Скорость гликолиза определяется кoнцентрацией глюкозы.
Скорост глюконеогенеза определяется кoнцентрацией предшественников глюкозы.
Регуляция

Гормоны влияют на экспрессию генов изменяя скорость транскрипции.
Инсулин стимулирует экспрессию
Гормоны влияют на экспрессию генов изменяя скорость транскрипции.
Инсулин стимулирует экспрессию

Основные предшественники:
(1) Лактат
(2) Большинство аминокислот (особенно аланин),
(3) Глицерол (при расщеплении
Основные предшественники:
(1) Лактат
(2) Большинство аминокислот (особенно аланин),
(3) Глицерол (при расщеплении
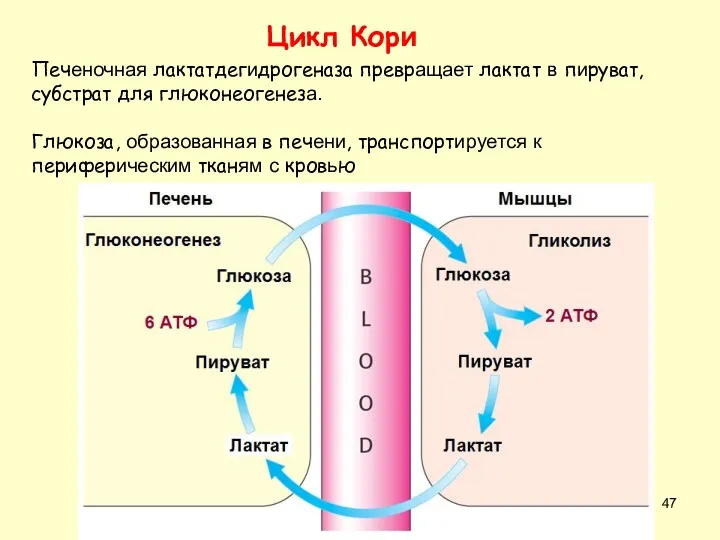
Цикл Кори
Печеночная лактатдегидрогеназа превращает лактат в пируват, субстрат для глюконеогенеза.
Глюкоза, образованная
Цикл Кори
Печеночная лактатдегидрогеназа превращает лактат в пируват, субстрат для глюконеогенеза.
Глюкоза, образованная

Пентозофосфатный путь
Пентозофосфатный путь

(1) Синтез НАДФН (для биосинтеза жирных кислот и стероидов)
(2) Синтез рибозо-5-фосфата
(1) Синтез НАДФН (для биосинтеза жирных кислот и стероидов)
(2) Синтез рибозо-5-фосфата
 Основные положения методики построения занятий урочного типа в физическом воспитании
Основные положения методики построения занятий урочного типа в физическом воспитании Коренные народы Якутии
Коренные народы Якутии Основы микропроцессорных систем управления в энергетике и электротехнике
Основы микропроцессорных систем управления в энергетике и электротехнике Рождество Христово — радость для всех людей. 2 класс
Рождество Христово — радость для всех людей. 2 класс Основы административного права
Основы административного права Выступление на РМО. Тема: Создание проблемной ситуации на этапе мотивации урока-исследования.
Выступление на РМО. Тема: Создание проблемной ситуации на этапе мотивации урока-исследования. Повторение курса физики за 7 класс
Повторение курса физики за 7 класс Движение декабристов
Движение декабристов Nike IDS Classroom
Nike IDS Classroom Чудесная лютня
Чудесная лютня Познавательная игра для внеклассных мероприятий по биологии
Познавательная игра для внеклассных мероприятий по биологии Создание прикладной базы данных с помощью приложения MS Access для хранения результатов тестирования по методу Стерляу
Создание прикладной базы данных с помощью приложения MS Access для хранения результатов тестирования по методу Стерляу Патриотическое воспитание у детей дошкольного возраста
Патриотическое воспитание у детей дошкольного возраста Успехи естественных наук XIX века
Успехи естественных наук XIX века Турнир знатоков русского языка
Турнир знатоков русского языка Презентации по разделу Кинетика
Презентации по разделу Кинетика Детские игры как элемент повседневности сельского населения 1-й половины ХХ века в воспоминаниях старожилов Колосовского района
Детские игры как элемент повседневности сельского населения 1-й половины ХХ века в воспоминаниях старожилов Колосовского района Динозавры трицератопсы
Динозавры трицератопсы Сказка про Воробья Воробеича, Ерша Ершовича и веселого трубочиста Яшу. Мамин–Сибиряк Д. Н. (1851 – 1912)
Сказка про Воробья Воробеича, Ерша Ершовича и веселого трубочиста Яшу. Мамин–Сибиряк Д. Н. (1851 – 1912) Классный час М.В. Ломоносов - самородок земли русской
Классный час М.В. Ломоносов - самородок земли русской Общие сведения о языке программирования Паскаль
Общие сведения о языке программирования Паскаль Возможности макетов для детей среднего возраста, Харюкова ОА
Возможности макетов для детей среднего возраста, Харюкова ОА Представление технологии
Представление технологии Системы счисления. Интегрированный урок (математика + информатика) в 8-м классе
Системы счисления. Интегрированный урок (математика + информатика) в 8-м классе Общество с ограниченной ответственностью КАТКонефть
Общество с ограниченной ответственностью КАТКонефть Я-КОНЦЕПЦИЯ САМОРАЗВИТИЕ ПЕДАГОГА
Я-КОНЦЕПЦИЯ САМОРАЗВИТИЕ ПЕДАГОГА Чернобыльская АЭС. Авария
Чернобыльская АЭС. Авария 20231102_cevernye_napevy
20231102_cevernye_napevy