- Эволюционная биология развития животных

Содержание
- 2. РЕГЛАМЕНТ ЗАНЯТИЙ: 12 Семинаров 10 Лекций 2
- 3. Алеев Ю.Г. Экоморфология. – Киев: Наукова думка, 1986. – 424с. Алёшин В.В., Петров Н.Б. Молекулярные свидетельства
- 4. Некоторые источники литературы: Малахов В.В. Новые представления о происхождении Bilateria (опыт применения метода эволюционной тетрады) //
- 5. Расклад по баллам: Исходные данные: 10 лекций и 14 семинаров (реально – 12). Присутствие на лекции
- 6. Животные – царство эвкариотных организмов, представители которого исходно отличаются отсутствием клеточной стенки (благодаря развитию механизма осморегуляции),
- 7. 1758 г. «Systema Naturae» - 10-е издание Земные царства отряды классы Lapideum Vegetabile (~1500) Mammalia Aves
- 8. ФИЛОГЕНЕТИЧЕСКИЕ ОТНОШЕНИЯ И ЧИСЛЕННОСТЬ ЦАРСТВ ЖИВЫХ ОРГАНИЗМОВ Plantae (Растения) ~ 350 тыс. видов Fungi (Грибы) ~100
- 9. Любарский Георгий Юрьевич р. 1959 г. С. 149: … Линней выбрал 4 признака в цветке и
- 10. 10 Система царств Уиттекера Система организмов Ю.В. Чайковского О системе организмов: «…искусственная – создаётся, естественная -
- 11. Хищные колониальные бактерии (Панов, 2001) Хищная нитчатая бактерия диктиобактер. Слева – деление «колонии» надвое. Справа диктиобактер
- 12. Изменения таксономического статуса биосистемы на разных уровнях организации (моно-, мета- и ценометабионт) 12 Уровни моно-, мета-
- 13. ЭТИМОЛОГИЯ НАЗВАНИЯ КУРСА и Соотношение понятий «ЭВОЛЮЦИЯ» и «РАЗВИТИЕ» Развитие: Направленность Необратимость Движущая сила находится в
- 14. Чарльз Дарвин (1809-1882) Чарльз Лайель (1797-1875) Альфред Уоллес (1823-1913) карикатура на Ч. Дарвина Эрнст Геккель (1834-1919)
- 15. Системы классификации (варианты мегасистематики) 14 Структура биоразнообразия как отражение его филогении. Монофилия и иерархия
- 16. НОВЕЙШЕЕ ЭВОЛЮЦИОННОЕ ДРЕВО ЖИВОТНОГО ЦАРСТВА НА ОСНОВЕ ОБЪЕДИНЕННЫХ ДАННЫХ АНАЛИЗА РАЗЛИЧНЫХ ГЕНОВ Protostomia Lophotrochozoa Ecdysozoa Deuterostomia
- 17. Э В О Л Ю Ц И О Н Н Ы Е О Т Н О
- 18. Фриц Мюллер (1821-1897) Эрнст Геккель (1834-1919) 18 Биогенетический закон
- 19. Недостатки существующих подходов в систематике сегодня, как отражение черт доминирующей эволюционной доктрины: засилье молекулярно-генетических методов, используемыми
- 20. В 1917 году этот шотландский зоолог написал книгу «Рост и форма», где представил свой «метод трансформации»
- 21. 21 Номогенез Работы Д’Арси Томпсона
- 22. 22
- 23. Номогенез Берг Лев Семёнович (1876-1950) Любищев Александр Александрович (1890-1972) Мейен Сергей Викторович (1935-1987) Чайковский Юрий Викторович
- 24. Коваленко Е.Е., Попов И.Ю. Новый подход к анализу свойств изменчивости // ЖОБ, 1997. т.58. №1. 24
- 25. Некоторые имена и понятия И.И. Шмальгаузен К. Уоддингтон В.Н. Беклемишев С.В. Мейен М.А. Шишкин Д.Л. Гродницкий
- 26. Ключевой признак биосистем, независимо от их иерархического статуса - - способность к самосохранению (в языке кибернетики
- 27. - система, обладающая свойствами, специфичными для живой природы параорганизменные (вирусы и внеклеточные биомолекулы – экзометаболиты и
- 28. Жизнь – природный феномен, сопутствующий квазистационарным круговоротам вещества в градиентах доступных форм энергии, и создающим для
- 29. Определения Жизни: - питание, рост и одряхление, причиной которых - принцип, имеющий смысл в самом себе
- 30. Системно-динамический подход: Мир состоит из движения элементарная система - это циклическое движение (с возвращением к начальному
- 31. Любой процесс становится возможным благодаря какому-либо градиенту потенциала, и прекращается по его исчерпании Свойства динамических систем
- 32. Гурвич Александр Гаврилович (1874-1954) Книга излагает оригинальные взгляды автора на методологию биологических исследований и пути создания
- 33. Уранов Алексей Александрович (1901-1974) Стр. 31: … участок ассоциации, занимаемый популяцией, можно представить как поле изменчивых
- 34. Уровни организации Биосистем (Голубец, 1982) 34 - избыточность понятийного аппарата, порой приводящая к путанице
- 35. Организм, особь, индивидуум Прокариотный - Эвкариотный Одноклеточный - Многоклеточный Безтканевой - Тканевой Унитарный - Модульный Симбионтный
- 36. Одноклеточное млекопитающее - - Хелацитон От человека произошёл новый вид. Известно даже имя этого человека: Генриетта
- 37. Самостоятельно произошедшие родственники Хелацитона : Рак тасманийских дьяволов (100-процентная смертность) - между животными передаются не вирусы
- 38. Андрей Валентинович Макрушин (род. 1934г.) российский гидробиолог, д.б.н. (г. Борок Ярославской обл.) А. В. Макрушин, В.
- 39. Современные представители многих типов многоклеточных - «унитаризированные» потомки некогда колониальных предков. Пример – наутилус, сохранивший следы
- 40. Примеры миелоидной экоморфы: 1 – Раффлезия (цветковое) 2 – Бовиста (гриб) 3 – Саккулина (рак) 4
- 41. 41 Метаболический Репродуктивный Локомоторный Место организма в системе экоморф определяется комбинативно, соотношением его позиций в рефренах
- 42. Биосистема (организм) отходы ресурсы деструкторы Метаболический модуль 42 – основа ценотических связей Потребители: хищники и/или паразиты
- 43. Усложнение других вариантов биосистем Одиночный образ жизни (бесполый) Одиночный образ жизни (половой) Агрегации (внешний фактор) Сообщество
- 44. Введение термина «Симбиогенез» (1909г. ) путал элемент с подсистемой Мережковский Константин Сергеевич (1855-1921) «Множество стало рождаться
- 45. О возможностях для диагностики различения понятий «элемент» и «подсистема» Чем ниже структурно-размерный ранг биосистем, тем выше
- 46. Рост сложности организма Появление эмерджентного cвойства Связи вариантов биосистем и разных уровней их организации популяция →
- 47. - (архе-)Детритная цепь – организмы связаны трофически лишь через детрит - Пастбищная цепь – организмы поедают
- 48. 1 Направления эволюции экосистем Рост сложности пищевой сети (варианты цепей) 2 3 Пастбищная Паразитная (архе-)детритная «Симбиогенетическая»
- 49. - выход, найденный Природой – взаимодействие между симбиосистемами на принципиально ином уровне: приобретение возможности информационного обмена
- 50. Этапы развития экосистемы по оси метаболического модуля Знаками + и – обозначено соответственно преимущественное увеличение или
- 51. Медников Борис Михайлович (1932-2001) С. 89: … сама дискретность гомологичности геномов позвоночных – твёрдо установленный факт,
- 52. Элементы «фоторобота» единой эволюционной теории Теории биологической эволюции Эволюция Биогенного круговорота (БиК) Эволюционные представления из гуманитарных
- 53. Принцип Ле-Шателье – Брауна, как частный случай 2-го закона термодинамики (применительно к Среде) Система состоит из
- 54. Элементы «фоторобота» единой эволюционной теории Теории биологической эволюции Эволюция Биогенного круговорота (БиК) Эволюционные представления из гуманитарных
- 55. Формальные свойства систем: Целостность, суммативность, механизация, централизация, иерархическая организация системы 55 Людвиг фон Берталанфи (1901-1972) Австрийский
- 56. Элементы «фоторобота» единой эволюционной теории Теории биологической эволюции Эволюция Биогенного круговорота (БиК) Эволюционные представления из гуманитарных
- 57. Герберт Спенсер (1820-1903) Английский философ, социолог, психолог, основатель органической школы в социологии, один из родоначальников позитивизма.
- 58. Уровни организации Материи (Кудрин Б.И., 2001) 58
- 59. Элементы «фоторобота» единой эволюционной теории Теории биологической эволюции Эволюция Биогенного круговорота (БиК) Эволюционные представления из гуманитарных
- 60. Принцип Пастера-Пригожина: При возникновении структур более высокого ранга равновероятны две альтернативные зеркальные формы: «левая» и «правая»
- 61. Элементы «фоторобота» единой эволюционной теории Теории биологической эволюции Эволюция Биогенного круговорота (БиК) Эволюционные представления из гуманитарных
- 62. Биогеохимические принципы В.И. Вернадского: Чарльз Дарвин (1809-1882) Вернадский Владимир Иванович (1863-1945) биогенная миграция стремится к максимуму
- 63. Элементы «фоторобота» единой эволюционной теории Теории биологической эволюции Эволюция Биогенного круговорота (БиК) Эволюционные представления из гуманитарных
- 64. Фриц Мюллер (1821-1897) Эрнст Геккель (1834-1919) 64 Биогенетический (системогенетический) закон
- 65. Элементы «фоторобота» единой эволюционной теории Теории биологической эволюции Эволюция Биогенного круговорота (БиК) Эволюционные представления из гуманитарных
- 66. Эпигенетический ландшафт (Уоддингтон 1966, цит. по Р. Рэфф, Т. Коффмен, 1986. рис. 9.1.) модель представляет собой
- 67. Основные постулаты ЭТЭ (Шишкин, 2003): Наследственность — не партнёр естественного отбора, а его продукт, выступающий как
- 68. Мутации дрозофил Мутация aristopedia (конечности на месте усиков) Мутация bithorax (наличие 2-х пар крыльев) Мутации окраски
- 69. Эта форма познания [ онтологическая редукция ] кажется странной … однако распространена гораздо шире, чем обычно
- 70. 70 Палеозой Мезозой Кайнозой Криптозой «ЕСТЕСТВЕННАЯ ИСТОРИЯ»
- 71. Биоэволюция гоминид 71
- 72. Некоторые теории антропогенеза «Научный» креационизм 72
- 74. Скачать презентацию
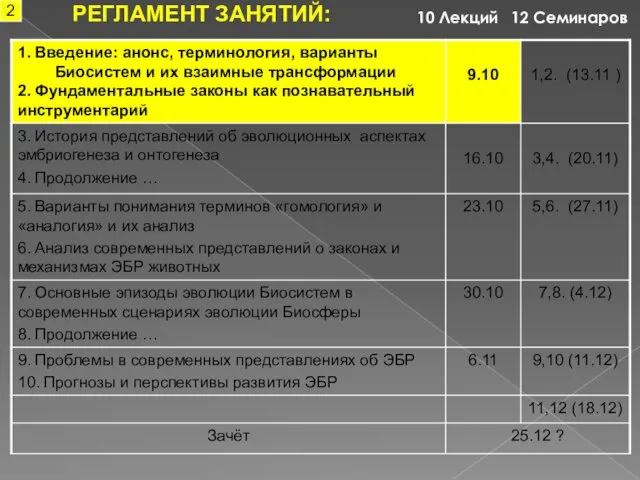
РЕГЛАМЕНТ ЗАНЯТИЙ:
12 Семинаров
10 Лекций
2
РЕГЛАМЕНТ ЗАНЯТИЙ:
12 Семинаров
10 Лекций
2

Алеев Ю.Г. Экоморфология. – Киев: Наукова думка, 1986. – 424с.
Алёшин В.В.,
Алеев Ю.Г. Экоморфология. – Киев: Наукова думка, 1986. – 424с.
Алёшин В.В.,

Некоторые источники литературы:
Малахов В.В. Новые представления о происхождении Bilateria (опыт применения
Некоторые источники литературы:
Малахов В.В. Новые представления о происхождении Bilateria (опыт применения

Расклад по баллам:
Исходные данные: 10 лекций и 14 семинаров (реально –
Расклад по баллам:
Исходные данные: 10 лекций и 14 семинаров (реально –

Животные – царство эвкариотных организмов, представители которого исходно отличаются отсутствием клеточной
Животные – царство эвкариотных организмов, представители которого исходно отличаются отсутствием клеточной
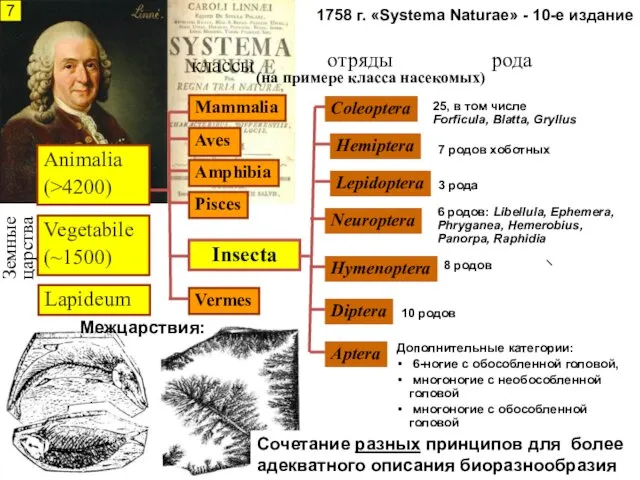
1758 г. «Systema Naturae» - 10-е издание
Земные царства
отряды
классы
Lapideum
Vegetabile
(~1500)
Mammalia
Aves
Amphibia
Pisces
Vermes
Coleoptera
Lepidoptera
Hemiptera
Neuroptera
Hymenoptera
Diptera
Aptera
рода
25, в
1758 г. «Systema Naturae» - 10-е издание
Земные царства
отряды
классы
Lapideum
Vegetabile
(~1500)
Mammalia
Aves
Amphibia
Pisces
Vermes
Coleoptera
Lepidoptera
Hemiptera
Neuroptera
Hymenoptera
Diptera
Aptera
рода
25, в
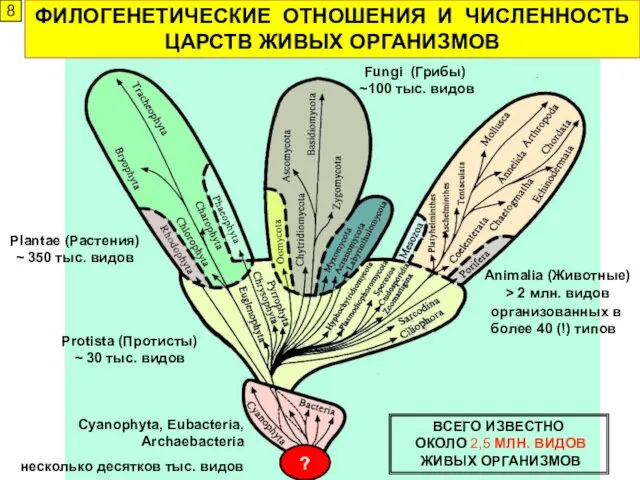
ФИЛОГЕНЕТИЧЕСКИЕ ОТНОШЕНИЯ И ЧИСЛЕННОСТЬ ЦАРСТВ ЖИВЫХ ОРГАНИЗМОВ
Plantae (Растения)
~ 350 тыс. видов
Fungi
ФИЛОГЕНЕТИЧЕСКИЕ ОТНОШЕНИЯ И ЧИСЛЕННОСТЬ ЦАРСТВ ЖИВЫХ ОРГАНИЗМОВ
Plantae (Растения)
~ 350 тыс. видов
Fungi

Любарский Георгий Юрьевич
р. 1959 г.
С. 149: … Линней выбрал
Любарский Георгий Юрьевич
р. 1959 г.
С. 149: … Линней выбрал

10
Система царств
Уиттекера
Система организмов
Ю.В. Чайковского
О системе организмов:
«…искусственная – создаётся,
естественная -
10
Система царств
Уиттекера
Система организмов
Ю.В. Чайковского
О системе организмов:
«…искусственная – создаётся,
естественная -
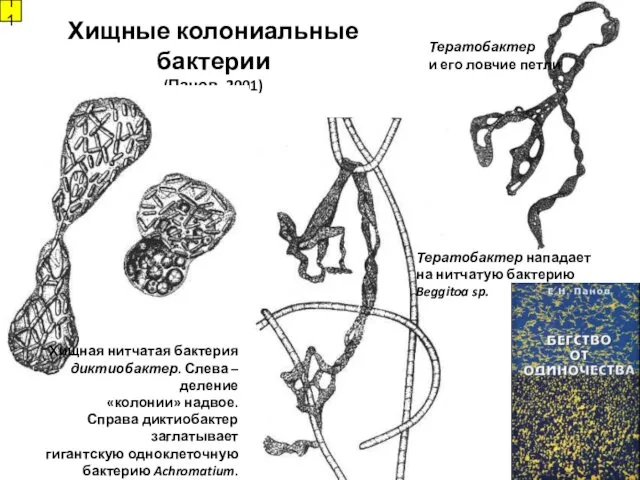
Хищные колониальные бактерии
(Панов, 2001)
Хищная нитчатая бактерия
диктиобактер. Слева – деление
«колонии»
Хищные колониальные бактерии
(Панов, 2001)
Хищная нитчатая бактерия
диктиобактер. Слева – деление
«колонии»

Изменения таксономического статуса биосистемы на разных уровнях организации (моно-, мета- и
Изменения таксономического статуса биосистемы на разных уровнях организации (моно-, мета- и

ЭТИМОЛОГИЯ НАЗВАНИЯ КУРСА и Соотношение понятий
«ЭВОЛЮЦИЯ» и «РАЗВИТИЕ»
Развитие:
Направленность
Необратимость
Движущая сила
ЭТИМОЛОГИЯ НАЗВАНИЯ КУРСА и Соотношение понятий
«ЭВОЛЮЦИЯ» и «РАЗВИТИЕ»
Развитие:
Направленность
Необратимость
Движущая сила
Чарльз Дарвин
(1809-1882)
Чарльз Лайель
(1797-1875)
Альфред Уоллес
(1823-1913)
карикатура
на Ч. Дарвина
Эрнст Геккель
(1834-1919)
Идея
эволюции
питекантропы
Геккеля
Жан Батист
Ламарк
Чарльз Дарвин
(1809-1882)
Чарльз Лайель
(1797-1875)
Альфред Уоллес
(1823-1913)
карикатура
на Ч. Дарвина
Эрнст Геккель
(1834-1919)
Идея
эволюции
питекантропы
Геккеля
Жан Батист
Ламарк
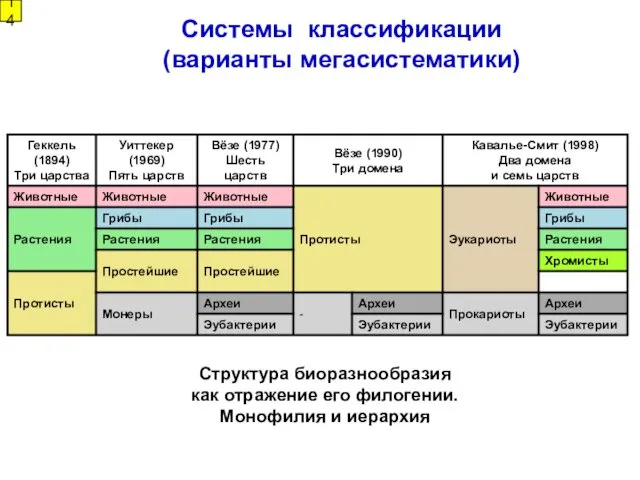
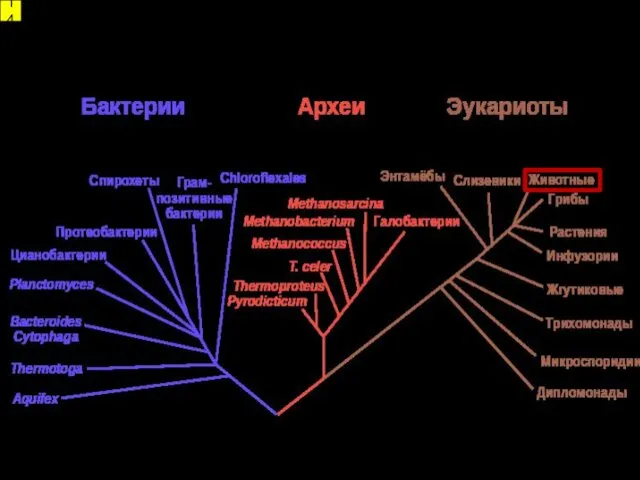
Системы классификации (варианты мегасистематики)
14
Структура биоразнообразия как отражение его филогении. Монофилия и
Системы классификации (варианты мегасистематики)
14
Структура биоразнообразия как отражение его филогении. Монофилия и
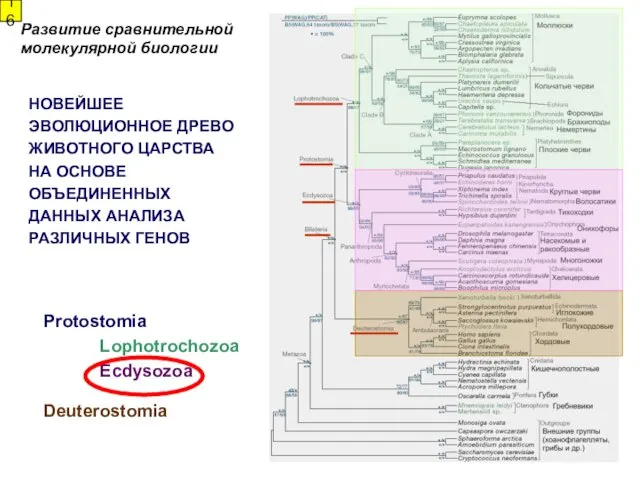
НОВЕЙШЕЕ
ЭВОЛЮЦИОННОЕ ДРЕВО
ЖИВОТНОГО ЦАРСТВА
НА ОСНОВЕ
ОБЪЕДИНЕННЫХ
ДАННЫХ АНАЛИЗА
РАЗЛИЧНЫХ ГЕНОВ
Protostomia
Lophotrochozoa
Ecdysozoa
Deuterostomia
Развитие сравнительной
молекулярной биологии
НОВЕЙШЕЕ
ЭВОЛЮЦИОННОЕ ДРЕВО
ЖИВОТНОГО ЦАРСТВА
НА ОСНОВЕ
ОБЪЕДИНЕННЫХ
ДАННЫХ АНАЛИЗА
РАЗЛИЧНЫХ ГЕНОВ
Protostomia
Lophotrochozoa
Ecdysozoa
Deuterostomia
Развитие сравнительной
молекулярной биологии
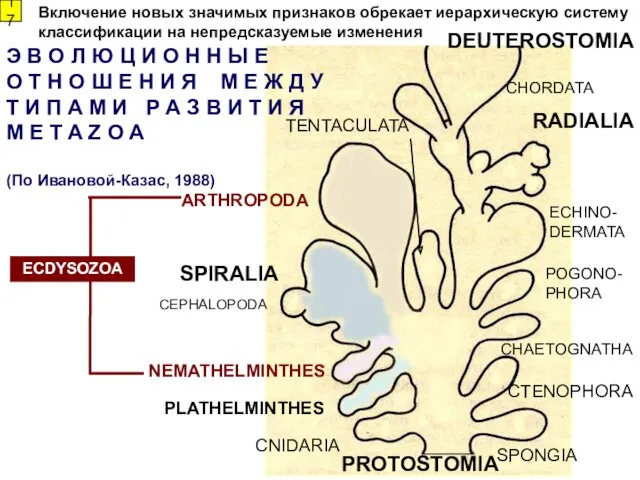
Э В О Л Ю Ц И О Н Н Ы
Э В О Л Ю Ц И О Н Н Ы
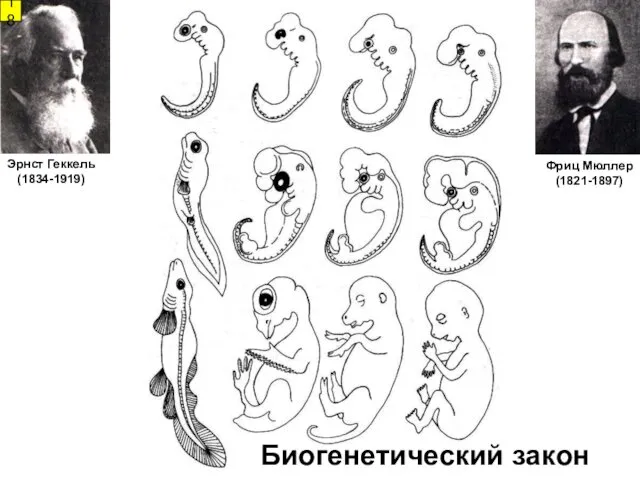
Фриц Мюллер
(1821-1897)
Эрнст Геккель
(1834-1919)
18
Биогенетический закон
Фриц Мюллер
(1821-1897)
Эрнст Геккель
(1834-1919)
18
Биогенетический закон

Недостатки существующих подходов в систематике сегодня, как отражение черт доминирующей
Недостатки существующих подходов в систематике сегодня, как отражение черт доминирующей
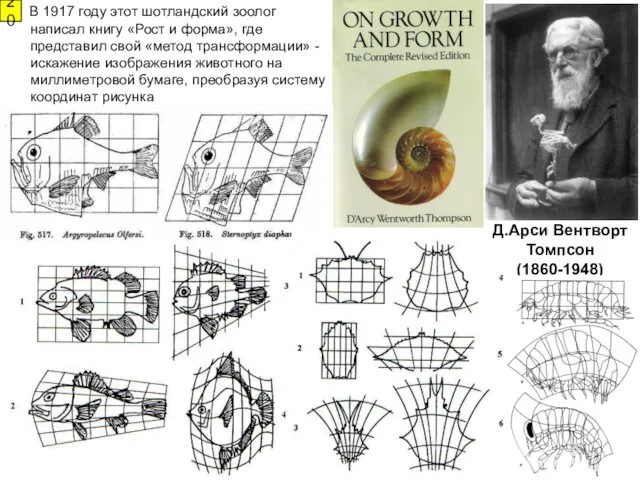
В 1917 году этот шотландский зоолог написал книгу «Рост и
В 1917 году этот шотландский зоолог написал книгу «Рост и
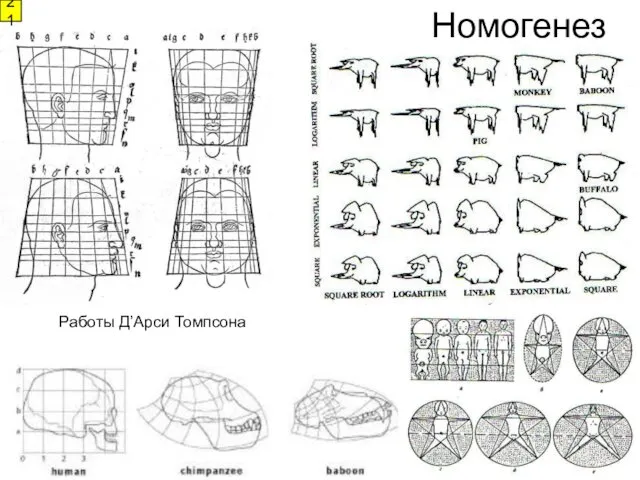
21
Номогенез
Работы Д’Арси Томпсона
21
Номогенез
Работы Д’Арси Томпсона
22
22

Номогенез
Берг Лев Семёнович
(1876-1950)
Любищев
Александр Александрович
(1890-1972)
Мейен
Сергей Викторович
(1935-1987)
Чайковский
Юрий
Номогенез
Берг Лев Семёнович
(1876-1950)
Любищев
Александр Александрович
(1890-1972)
Мейен
Сергей Викторович
(1935-1987)
Чайковский
Юрий

Коваленко Е.Е., Попов И.Ю. Новый подход к анализу свойств изменчивости
//
Коваленко Е.Е., Попов И.Ю. Новый подход к анализу свойств изменчивости
//

Некоторые имена и понятия
И.И. Шмальгаузен
К. Уоддингтон
В.Н. Беклемишев
С.В. Мейен
М.А. Шишкин
Д.Л. Гродницкий
А.П. Расницын
и
Некоторые имена и понятия
И.И. Шмальгаузен
К. Уоддингтон
В.Н. Беклемишев
С.В. Мейен
М.А. Шишкин
Д.Л. Гродницкий
А.П. Расницын
и

Ключевой признак биосистем, независимо от их иерархического статуса - - способность
Ключевой признак биосистем, независимо от их иерархического статуса - - способность

- система, обладающая свойствами, специфичными для живой природы
параорганизменные (вирусы и внеклеточные
- система, обладающая свойствами, специфичными для живой природы
параорганизменные (вирусы и внеклеточные

Жизнь – природный феномен, сопутствующий квазистационарным круговоротам вещества в градиентах
Жизнь – природный феномен, сопутствующий квазистационарным круговоротам вещества в градиентах

Определения Жизни:
- питание, рост и одряхление, причиной которых - принцип,
Определения Жизни:
- питание, рост и одряхление, причиной которых - принцип,
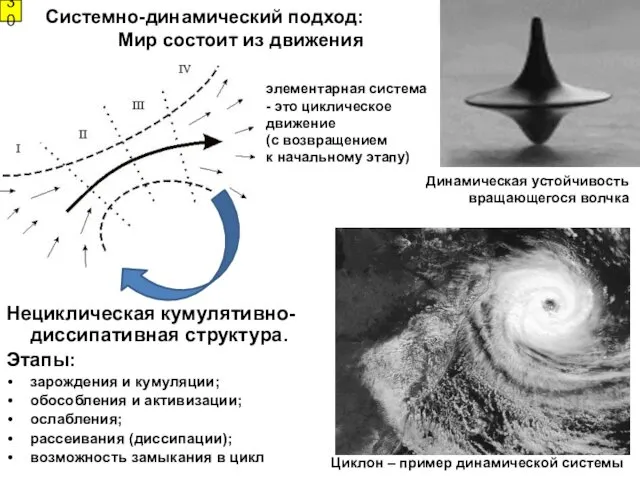
Системно-динамический подход: Мир состоит из движения
элементарная система - это циклическое движение
Системно-динамический подход: Мир состоит из движения
элементарная система - это циклическое движение
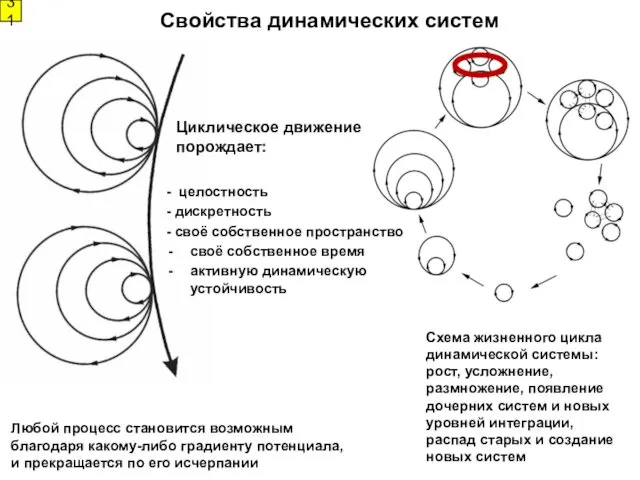
Любой процесс становится возможным благодаря какому-либо градиенту потенциала, и прекращается по
Любой процесс становится возможным благодаря какому-либо градиенту потенциала, и прекращается по

Гурвич Александр Гаврилович
(1874-1954)
Книга излагает оригинальные взгляды автора на методологию биологических исследований
Гурвич Александр Гаврилович
(1874-1954)
Книга излагает оригинальные взгляды автора на методологию биологических исследований

Уранов
Алексей Александрович
(1901-1974)
Стр. 31:
… участок ассоциации, занимаемый популяцией, можно представить
Уранов
Алексей Александрович
(1901-1974)
Стр. 31:
… участок ассоциации, занимаемый популяцией, можно представить
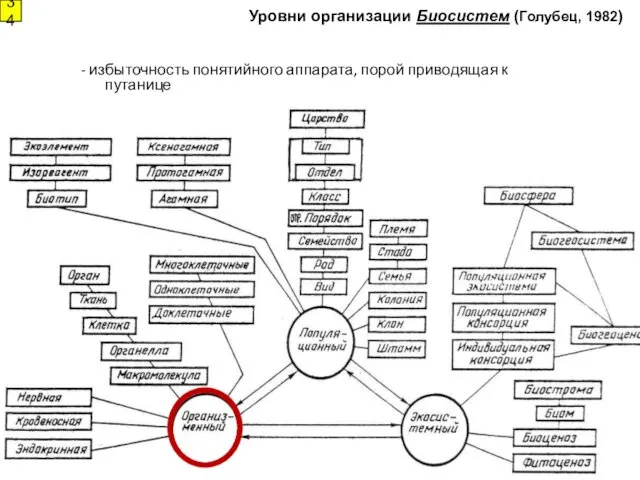
Уровни организации Биосистем (Голубец, 1982)
34
- избыточность понятийного аппарата, порой приводящая к
Уровни организации Биосистем (Голубец, 1982)
34
- избыточность понятийного аппарата, порой приводящая к

Организм, особь, индивидуум
Прокариотный - Эвкариотный
Одноклеточный - Многоклеточный
Безтканевой - Тканевой
Унитарный - Модульный
Симбионтный
и
Организм, особь, индивидуум
Прокариотный - Эвкариотный
Одноклеточный - Многоклеточный
Безтканевой - Тканевой
Унитарный - Модульный
Симбионтный
и

Одноклеточное млекопитающее - - Хелацитон
От человека произошёл новый вид. Известно
Одноклеточное млекопитающее - - Хелацитон
От человека произошёл новый вид. Известно
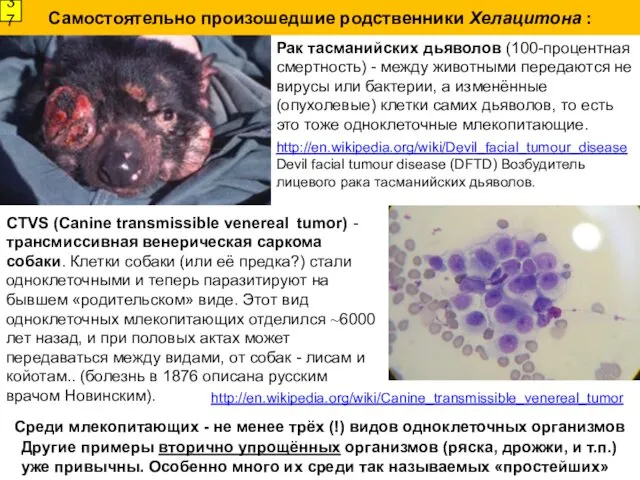
Самостоятельно произошедшие родственники Хелацитона :
Рак тасманийских дьяволов (100-процентная смертность) - между
Самостоятельно произошедшие родственники Хелацитона :
Рак тасманийских дьяволов (100-процентная смертность) - между

Андрей Валентинович Макрушин
(род. 1934г.)
российский гидробиолог,
д.б.н.
(г. Борок Ярославской обл.)
А. В. Макрушин, В. В.
Андрей Валентинович Макрушин
(род. 1934г.)
российский гидробиолог,
д.б.н.
(г. Борок Ярославской обл.)
А. В. Макрушин, В. В.

Современные представители многих типов многоклеточных - «унитаризированные» потомки некогда колониальных
Современные представители многих типов многоклеточных - «унитаризированные» потомки некогда колониальных

Примеры миелоидной
экоморфы:
1 – Раффлезия (цветковое)
2 – Бовиста (гриб)
3 – Саккулина
Примеры миелоидной
экоморфы:
1 – Раффлезия (цветковое)
2 – Бовиста (гриб)
3 – Саккулина
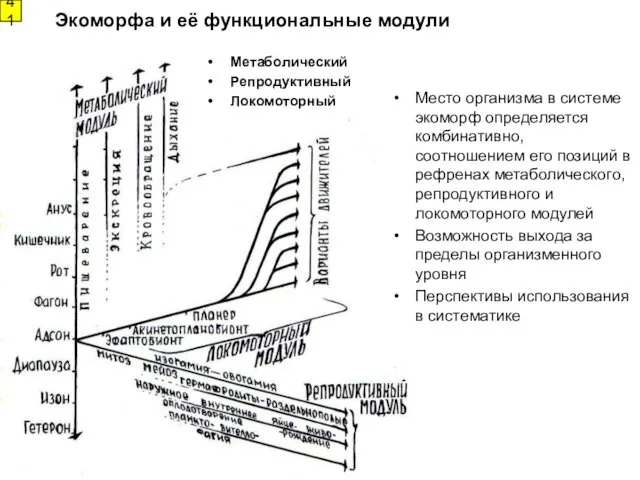
41
Метаболический
Репродуктивный
Локомоторный
Место организма в системе экоморф определяется комбинативно, соотношением его
41
Метаболический
Репродуктивный
Локомоторный
Место организма в системе экоморф определяется комбинативно, соотношением его
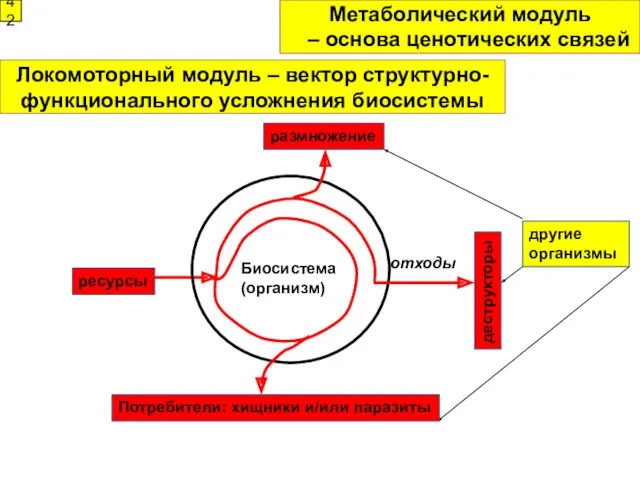
Биосистема
(организм)
отходы
ресурсы
деструкторы
Метаболический модуль
42
– основа ценотических связей
Потребители: хищники и/или паразиты
размножение
другие организмы
Локомоторный модуль
Биосистема
(организм)
отходы
ресурсы
деструкторы
Метаболический модуль
42
– основа ценотических связей
Потребители: хищники и/или паразиты
размножение
другие организмы
Локомоторный модуль

Усложнение других вариантов биосистем
Одиночный образ жизни (бесполый)
Одиночный образ жизни (половой)
Агрегации (внешний
Усложнение других вариантов биосистем
Одиночный образ жизни (бесполый)
Одиночный образ жизни (половой)
Агрегации (внешний

Введение термина
«Симбиогенез» (1909г. )
путал элемент
с подсистемой
Мережковский
Константин Сергеевич
(1855-1921)
«Множество стало
Введение термина
«Симбиогенез» (1909г. )
путал элемент
с подсистемой
Мережковский
Константин Сергеевич
(1855-1921)
«Множество стало

О возможностях для диагностики различения понятий «элемент» и «подсистема»
Чем ниже структурно-размерный
О возможностях для диагностики различения понятий «элемент» и «подсистема»
Чем ниже структурно-размерный
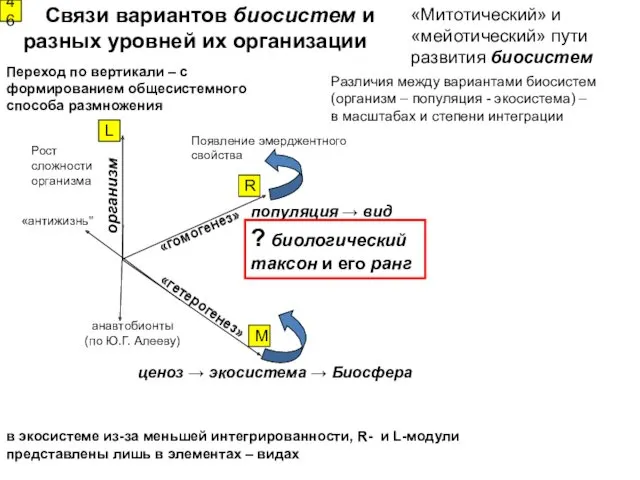
Рост сложности
организма
Появление эмерджентного
cвойства
Связи вариантов биосистем и разных уровней
Рост сложности
организма
Появление эмерджентного
cвойства
Связи вариантов биосистем и разных уровней
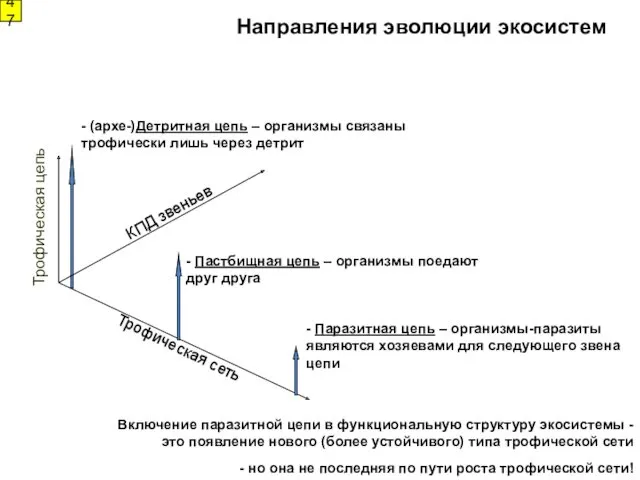
- (архе-)Детритная цепь – организмы связаны
трофически лишь через детрит
-
- (архе-)Детритная цепь – организмы связаны
трофически лишь через детрит
-
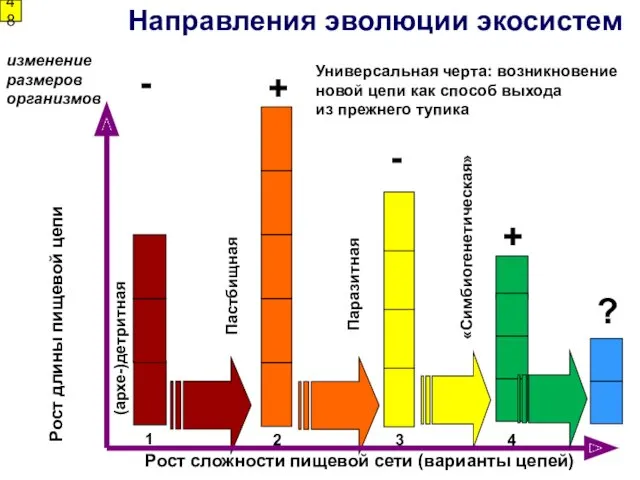
1
Направления эволюции экосистем
Рост сложности пищевой сети (варианты цепей)
2
3
Пастбищная
Паразитная
(архе-)детритная
«Симбиогенетическая»
4
Рост длины пищевой
1
Направления эволюции экосистем
Рост сложности пищевой сети (варианты цепей)
2
3
Пастбищная
Паразитная
(архе-)детритная
«Симбиогенетическая»
4
Рост длины пищевой
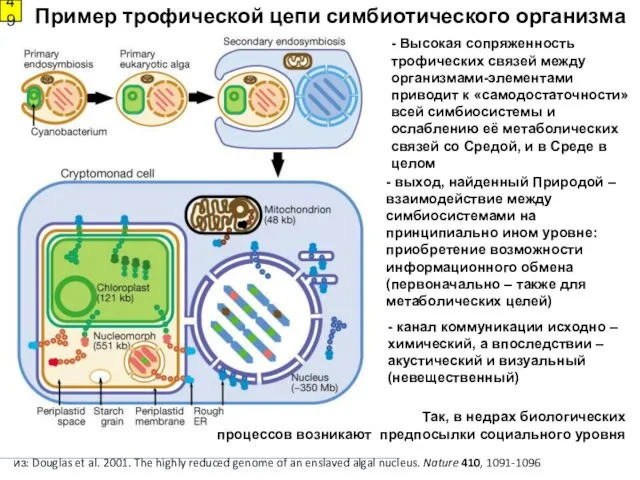
- выход, найденный Природой – взаимодействие между симбиосистемами на принципиально ином
- выход, найденный Природой – взаимодействие между симбиосистемами на принципиально ином
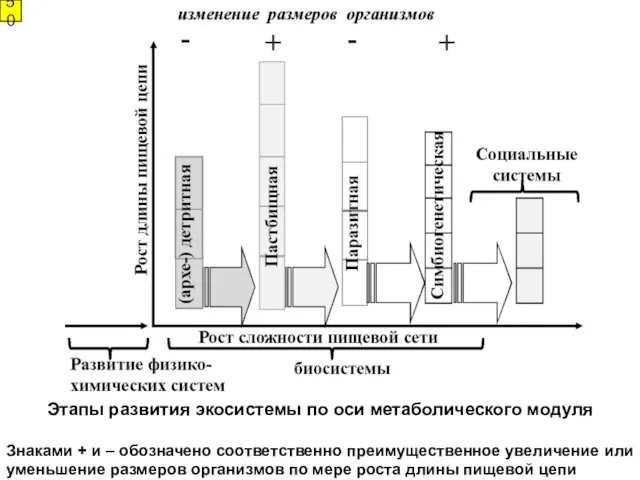
Этапы развития экосистемы по оси метаболического модуля
Знаками + и – обозначено
Этапы развития экосистемы по оси метаболического модуля
Знаками + и – обозначено
Медников
Борис Михайлович
(1932-2001)
С. 89: … сама дискретность гомологичности геномов позвоночных –
Медников
Борис Михайлович
(1932-2001)
С. 89: … сама дискретность гомологичности геномов позвоночных –

Элементы «фоторобота» единой эволюционной теории
Теории биологической эволюции
Эволюция Биогенного круговорота (БиК)
Эволюционные представления
Элементы «фоторобота» единой эволюционной теории
Теории биологической эволюции
Эволюция Биогенного круговорота (БиК)
Эволюционные представления

Принцип Ле-Шателье – Брауна, как частный случай 2-го закона термодинамики
Принцип Ле-Шателье – Брауна, как частный случай 2-го закона термодинамики

Элементы «фоторобота» единой эволюционной теории
Теории биологической эволюции
Эволюция Биогенного круговорота (БиК)
Эволюционные представления
Элементы «фоторобота» единой эволюционной теории
Теории биологической эволюции
Эволюция Биогенного круговорота (БиК)
Эволюционные представления

Формальные свойства систем:
Целостность, суммативность, механизация, централизация, иерархическая организация системы
55
Людвиг фон
Формальные свойства систем:
Целостность, суммативность, механизация, централизация, иерархическая организация системы
55
Людвиг фон

Элементы «фоторобота» единой эволюционной теории
Теории биологической эволюции
Эволюция Биогенного круговорота (БиК)
Эволюционные представления
Элементы «фоторобота» единой эволюционной теории
Теории биологической эволюции
Эволюция Биогенного круговорота (БиК)
Эволюционные представления

Герберт Спенсер
(1820-1903)
Английский философ, социолог, психолог, основатель органической школы в социологии, один
Герберт Спенсер
(1820-1903)
Английский философ, социолог, психолог, основатель органической школы в социологии, один
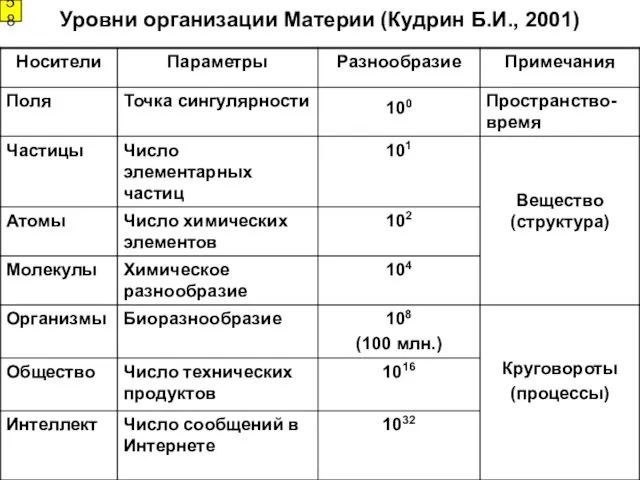
Уровни организации Материи (Кудрин Б.И., 2001)
58
Уровни организации Материи (Кудрин Б.И., 2001)
58

Элементы «фоторобота» единой эволюционной теории
Теории биологической эволюции
Эволюция Биогенного круговорота (БиК)
Эволюционные представления
Элементы «фоторобота» единой эволюционной теории
Теории биологической эволюции
Эволюция Биогенного круговорота (БиК)
Эволюционные представления
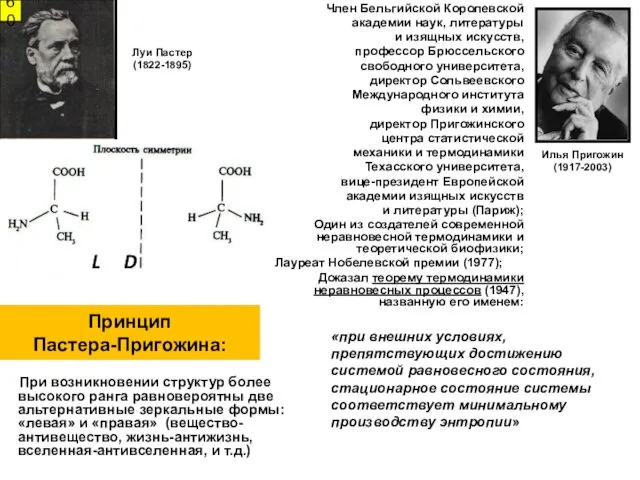
Принцип
Пастера-Пригожина:
При возникновении структур более высокого ранга равновероятны две альтернативные зеркальные
Принцип
Пастера-Пригожина:
При возникновении структур более высокого ранга равновероятны две альтернативные зеркальные

Элементы «фоторобота» единой эволюционной теории
Теории биологической эволюции
Эволюция Биогенного круговорота (БиК)
Эволюционные представления
Элементы «фоторобота» единой эволюционной теории
Теории биологической эволюции
Эволюция Биогенного круговорота (БиК)
Эволюционные представления

Биогеохимические принципы
В.И. Вернадского:
Чарльз Дарвин
(1809-1882)
Вернадский
Владимир Иванович
(1863-1945)
биогенная миграция стремится
Биогеохимические принципы
В.И. Вернадского:
Чарльз Дарвин
(1809-1882)
Вернадский
Владимир Иванович
(1863-1945)
биогенная миграция стремится

Элементы «фоторобота» единой эволюционной теории
Теории биологической эволюции
Эволюция Биогенного круговорота (БиК)
Эволюционные представления
Элементы «фоторобота» единой эволюционной теории
Теории биологической эволюции
Эволюция Биогенного круговорота (БиК)
Эволюционные представления
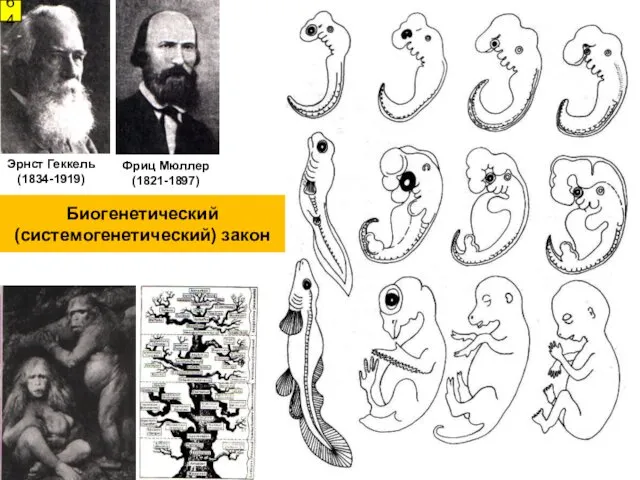
Фриц Мюллер
(1821-1897)
Эрнст Геккель
(1834-1919)
64
Биогенетический (системогенетический) закон
Фриц Мюллер
(1821-1897)
Эрнст Геккель
(1834-1919)
64
Биогенетический (системогенетический) закон

Элементы «фоторобота» единой эволюционной теории
Теории биологической эволюции
Эволюция Биогенного круговорота (БиК)
Эволюционные представления
Элементы «фоторобота» единой эволюционной теории
Теории биологической эволюции
Эволюция Биогенного круговорота (БиК)
Эволюционные представления
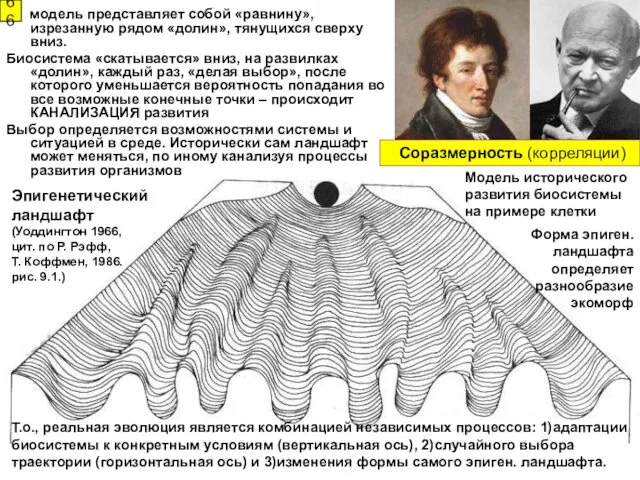
Эпигенетический
ландшафт
(Уоддингтон 1966,
цит. по Р. Рэфф,
Т. Коффмен, 1986.
Эпигенетический ландшафт (Уоддингтон 1966, цит. по Р. Рэфф, Т. Коффмен, 1986.

Основные постулаты ЭТЭ (Шишкин, 2003):
Наследственность — не партнёр естественного отбора, а
Основные постулаты ЭТЭ (Шишкин, 2003):
Наследственность — не партнёр естественного отбора, а

Мутации дрозофил
Мутация aristopedia (конечности на месте усиков)
Мутация bithorax (наличие 2-х пар
Мутации дрозофил
Мутация aristopedia (конечности на месте усиков)
Мутация bithorax (наличие 2-х пар
![Эта форма познания [ онтологическая редукция ] кажется странной …](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/43425/slide-68.jpg)
Эта форма познания [ онтологическая редукция ] кажется странной … однако
Эта форма познания [ онтологическая редукция ] кажется странной … однако

70
Палеозой
Мезозой
Кайнозой
Криптозой
«ЕСТЕСТВЕННАЯ ИСТОРИЯ»
70
Палеозой
Мезозой
Кайнозой
Криптозой
«ЕСТЕСТВЕННАЯ ИСТОРИЯ»

Биоэволюция
гоминид
71
Биоэволюция
гоминид
71

Некоторые теории антропогенеза
«Научный» креационизм
72
Некоторые теории антропогенеза
«Научный» креационизм
72
 Тестовая работа по химии.Тема Жиры.
Тестовая работа по химии.Тема Жиры. Применение солнечных электростанций в энергоэффективном (энергоактивном) архитектурном проектировании
Применение солнечных электростанций в энергоэффективном (энергоактивном) архитектурном проектировании Складки – собственная геометрия. Собственные геометрические параметры складок
Складки – собственная геометрия. Собственные геометрические параметры складок Автоматическое регулирование технологических процессов. Системы автоматического регулирования
Автоматическое регулирование технологических процессов. Системы автоматического регулирования Философия XIX века
Философия XIX века Педагогика профессионального образования (занятие 1)
Педагогика профессионального образования (занятие 1) Образовательное пространство в дошкольном учреждении
Образовательное пространство в дошкольном учреждении Правление Ивана IV
Правление Ивана IV Качество медицинских товаров и его свойства
Качество медицинских товаров и его свойства Профессии воды.
Профессии воды. Презентация 8 Марта
Презентация 8 Марта Лакокрасочные товары
Лакокрасочные товары PPE_ JOCAP 3
PPE_ JOCAP 3 Різноманітність комах та хребетних тварин луки
Різноманітність комах та хребетних тварин луки Культура Руси XIII-XIV веков
Культура Руси XIII-XIV веков Рыцари
Рыцари Эпоха гуннов
Эпоха гуннов Препарация зубов под Targis/Vectris
Препарация зубов под Targis/Vectris Происхождение человека
Происхождение человека Понятие и система показателей качества продукции
Понятие и система показателей качества продукции Время реакции человека
Время реакции человека Антарктида. Природа.
Антарктида. Природа. Объёмные цветы
Объёмные цветы Презентация к рассказу К. Паустовского Кот- ворюга
Презентация к рассказу К. Паустовского Кот- ворюга Основы теории государства. Конституционный строй Российской Федерации
Основы теории государства. Конституционный строй Российской Федерации Музыкально - дидактическая интерактивная игра Музыкальные инструменты
Музыкально - дидактическая интерактивная игра Музыкальные инструменты Размерный анализ. Точность технологических операций
Размерный анализ. Точность технологических операций проект игрушечный город
проект игрушечный город