- Фізіологія м‘язів

Содержание
- 2. Скелетні м'язи починають скорочуватися лише після того як отримають ПД від мотонейрона. Проведення ПД від мотонейрона
- 3. Синаптична щілина: Простір між двома мембранами Ширина - 20-30 нм Заповнена синаптичної рідиною, своїм складом нагадує
- 4. Усередині нервового закінчення є велика кількість (до 300.000) синаптичних пухирців (діаметром близько 50 нм), що містять
- 5. Вихід медіатора в нервово-м'язовому синапсі При надходженні ПД до пресинаптичної мембрани в ній відкриваються Са2 +
- 6. Медіатор дифундує по синаптичної рідини і велика частина його молекул досягає постсинаптичної мембрани, де взаимодія з
- 7. Дія медіатора Викид медіатора забезпечує взаємодію його з лігандзавісімимі структурами каналу (ХР). При цьому ворота відкриваються
- 8. ПД по нерву можуть надходити з максимальною частотою до 1000 імп / с. Зазвичай по нерву
- 9. АХ після взаємодії з холінорецептор швидко руйнується розташованої поруч ферментом (холінестеразою). Тому для продовження ефекту (посилення
- 10. Явище суммації Зазвичай для передачі тільки одного ПД вивільняється до мільйона молекул АХ (200-300 везикул). При
- 11. У нервовому волокні відбувається постійне поповнення медіатора. Тут є кілька механізмів відновлення везикул з медіатором. медіатор
- 12. Проведення ПД по мембрані НА малюнку я нагадую як ПД проводиться по сарколемі від точки до
- 13. СПР – депо Са2+ Під впливом ПД, що проводиться по мембрані м'язового волокна і заходить на
- 14. Скелетний м'яз складається з м'язових волокон. Всередині їх містяться найважливіші органели: мітохондрії, саркоплазматический ретикулум. А так
- 15. Актин складається з двох скручених ланцюжків актінових білків. Між ними розташовується білок тропомиозин. Його частина (тропонин
- 16. Схема взаємодії актинових и міозинових філаментів
- 17. Взаємодія актинових и міозинових філаментів
- 18. Вихід кальцію забезпечує відкриття активних центрів актину і взаємодія міозинової головки з актином. Після цього головка
- 19. Крок Зазначене вище іменується «кроковим» механізмом.
- 20. Етапи «крокової» механізму Послідовні етапи: а - розслаблення, б - з'єднання миозинових головок з активним центром
- 21. Тропонін С, наявний в тонких волокнах, ідентичний кальмодуліном. Приєднуючи Са2 +, кальмодулін сприяє активації АТФази і
- 22. Деполяризація мембрани цистерн відкриває тут Електровозбудімость кальцієві канали. У зв'язку з тим, що в саркоплазме концентрація
- 23. 1 - викид медіатора в синаптичну щілина. 2 - звільнення активного центру актину. 3 - розслаблення
- 24. Співвідношення ПД і рефрактерності у різних м'язів Тривалість періоду рефрактерності у скелетного м'яза невелика, тому при
- 25. Різні режими скорочення скелетного м'яза а - одиночне скорочення, б - неповний тетанус, в - повний
- 26. а) скорочення (утворення містків); б) розслаблення (розриву містків); в) роботи Са2 + -насоса (2 АТФ на
- 27. Шляхи ресинтезу АТФ 1) креатинфосфокіназний (КФ): АДФ + КФ АТФ + К 2) гліколітичний, З) аеробне
- 28. а) фосфогенний (КФ) - 3,6 моль АТФ/хв, б) гліколітічний - 1,2 моль АТФ/хв, в) окислювальний -
- 29. Рухова одиниця - одиничне нервове волокно мотонейрона а також іннервуючі ним м'язові волокна, складають одну ДЕ
- 30. Швидкі і повільні ДЕ Швидкі Великий мотонейрон (висока швидкість проведення ПД). Багато АТФ. Багато КФ. Активний
- 31. Гладкі м’язи
- 33. Скачать презентацию

Скелетні м'язи починають скорочуватися лише після того як отримають ПД від
Скелетні м'язи починають скорочуватися лише після того як отримають ПД від
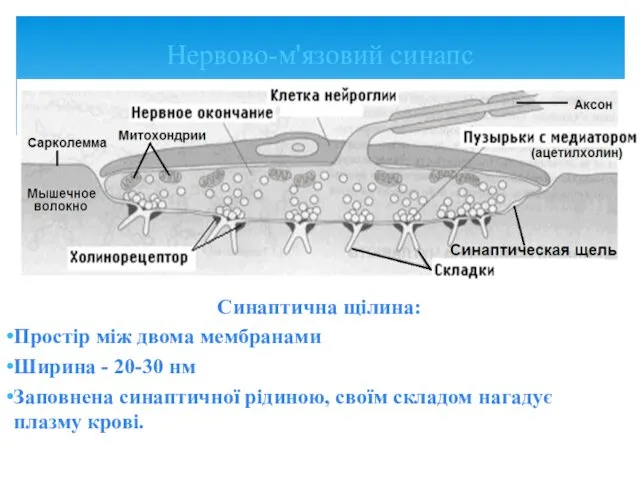
Синаптична щілина:
Простір між двома мембранами
Ширина - 20-30 нм
Заповнена синаптичної рідиною, своїм
Синаптична щілина:
Простір між двома мембранами
Ширина - 20-30 нм
Заповнена синаптичної рідиною, своїм

Усередині нервового закінчення є велика кількість (до 300.000) синаптичних пухирців (діаметром
Усередині нервового закінчення є велика кількість (до 300.000) синаптичних пухирців (діаметром
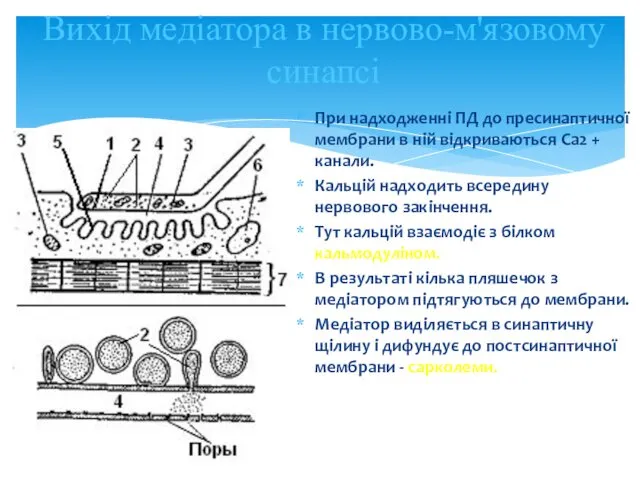
Вихід медіатора в нервово-м'язовому синапсі
При надходженні ПД до пресинаптичної мембрани в
Вихід медіатора в нервово-м'язовому синапсі
При надходженні ПД до пресинаптичної мембрани в

Медіатор дифундує по синаптичної рідини і велика частина його молекул досягає
Медіатор дифундує по синаптичної рідини і велика частина його молекул досягає
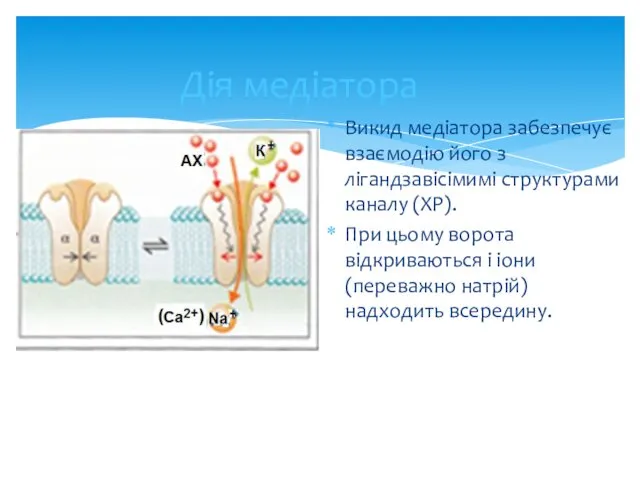
Дія медіатора
Викид медіатора забезпечує взаємодію його з лігандзавісімимі структурами каналу (ХР).
При
Дія медіатора
Викид медіатора забезпечує взаємодію його з лігандзавісімимі структурами каналу (ХР).
При

ПД по нерву можуть надходити з максимальною частотою до 1000 імп
ПД по нерву можуть надходити з максимальною частотою до 1000 імп
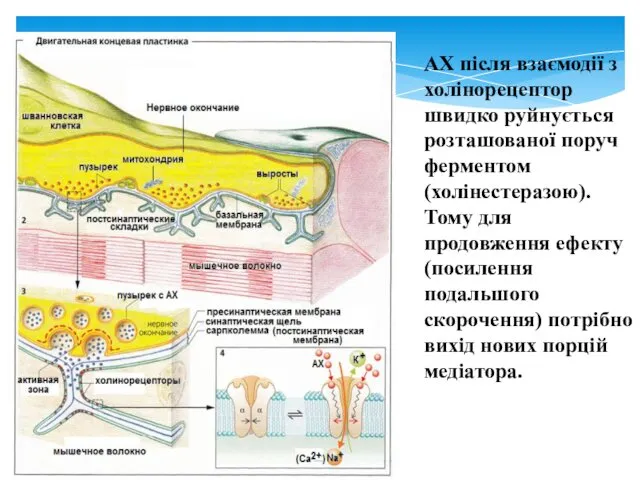
АХ після взаємодії з холінорецептор швидко руйнується розташованої поруч ферментом (холінестеразою).
Тому
АХ після взаємодії з холінорецептор швидко руйнується розташованої поруч ферментом (холінестеразою).
Тому
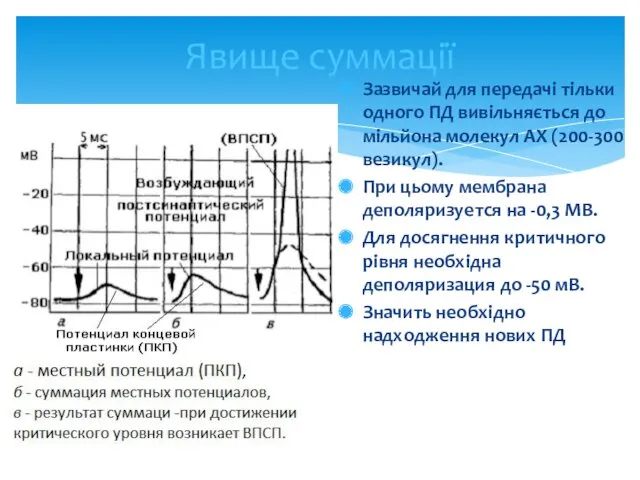
Явище суммації
Зазвичай для передачі тільки одного ПД вивільняється до мільйона молекул
Явище суммації
Зазвичай для передачі тільки одного ПД вивільняється до мільйона молекул

У нервовому волокні відбувається постійне поповнення медіатора. Тут є кілька механізмів
У нервовому волокні відбувається постійне поповнення медіатора. Тут є кілька механізмів
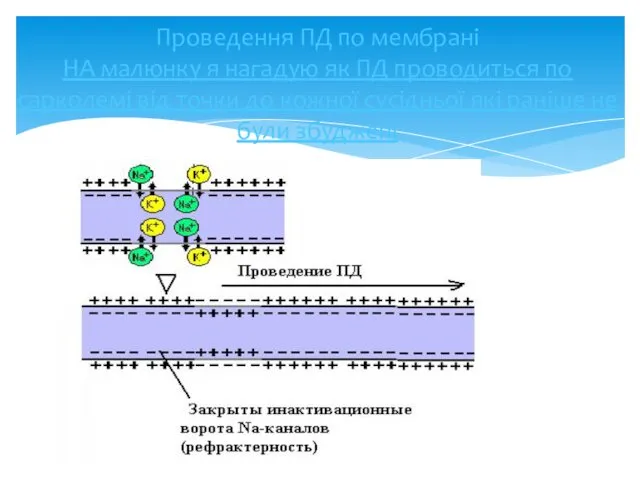
Проведення ПД по мембрані
НА малюнку я нагадую як ПД проводиться по
Проведення ПД по мембрані НА малюнку я нагадую як ПД проводиться по
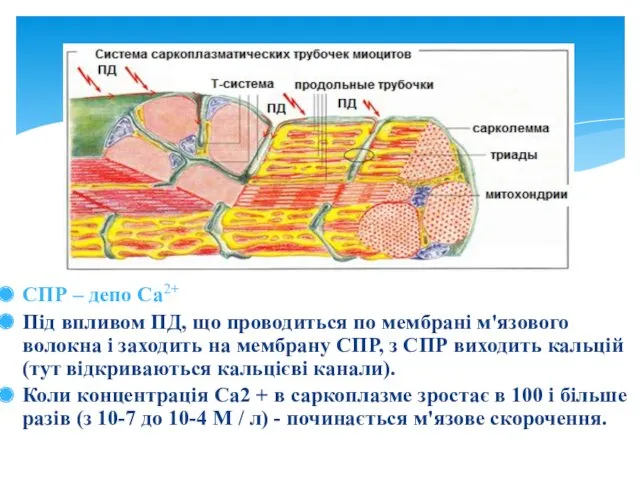
СПР – депо Са2+
Під впливом ПД, що проводиться по мембрані
СПР – депо Са2+
Під впливом ПД, що проводиться по мембрані
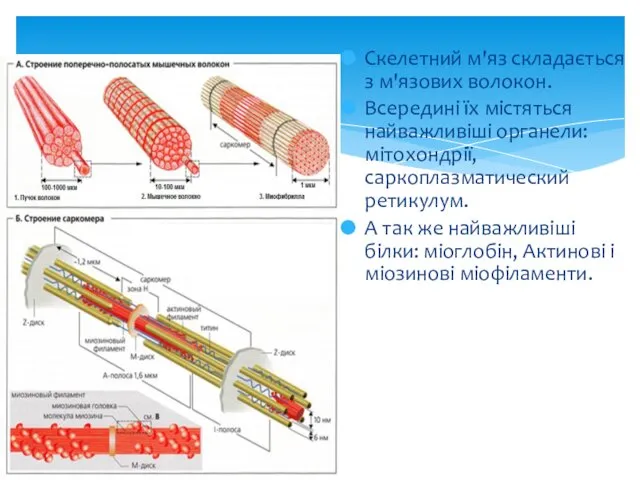
Скелетний м'яз складається з м'язових волокон.
Всередині їх містяться найважливіші органели: мітохондрії,
Скелетний м'яз складається з м'язових волокон.
Всередині їх містяться найважливіші органели: мітохондрії,
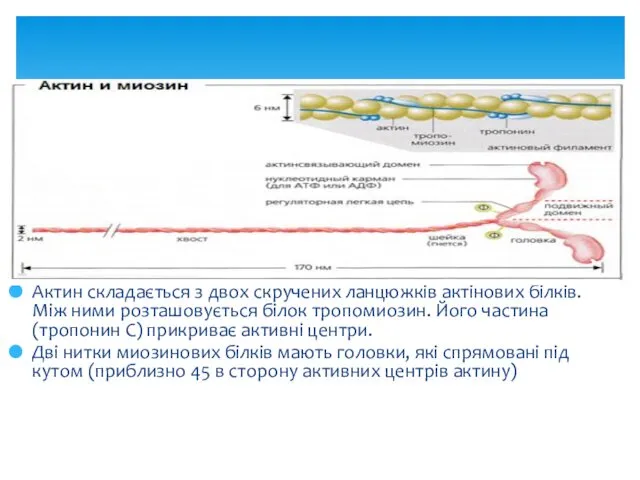
Актин складається з двох скручених ланцюжків актінових білків. Між ними розташовується
Актин складається з двох скручених ланцюжків актінових білків. Між ними розташовується
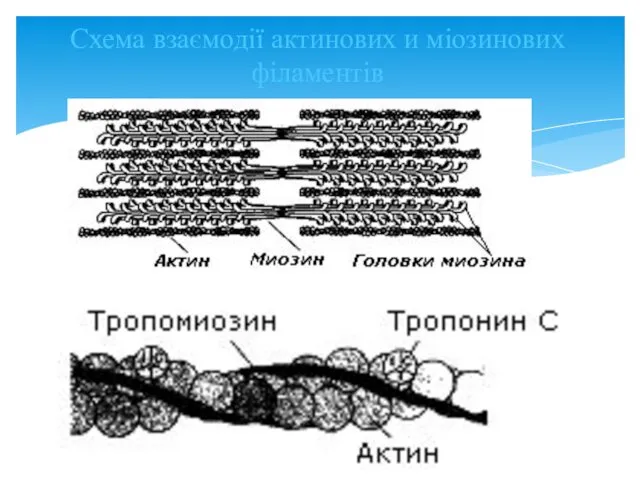
Схема взаємодії актинових и міозинових філаментів
Схема взаємодії актинових и міозинових філаментів
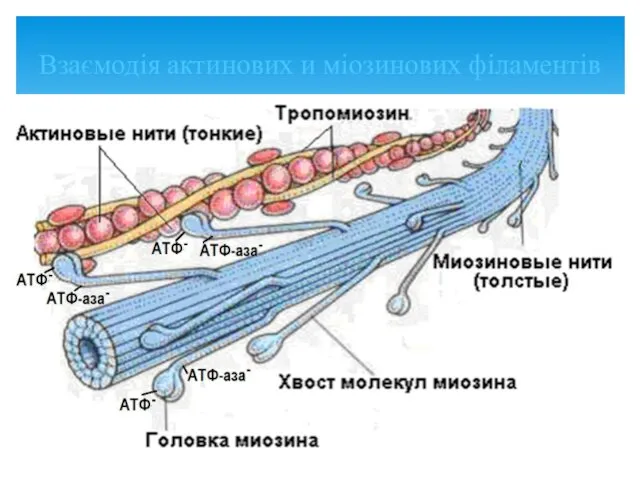
Взаємодія актинових и міозинових філаментів
Взаємодія актинових и міозинових філаментів
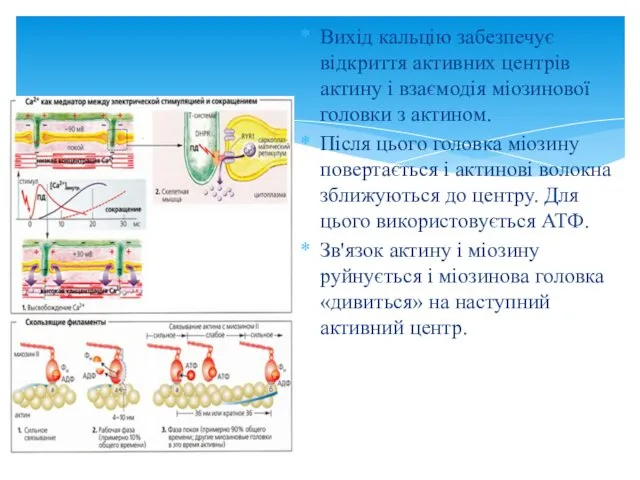
Вихід кальцію забезпечує відкриття активних центрів актину і взаємодія міозинової головки
Вихід кальцію забезпечує відкриття активних центрів актину і взаємодія міозинової головки
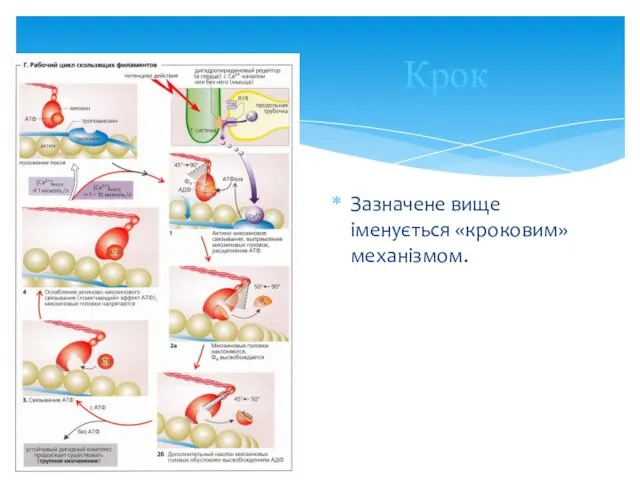
Крок
Зазначене вище іменується «кроковим» механізмом.
Крок
Зазначене вище іменується «кроковим» механізмом.

Етапи «крокової» механізму
Послідовні етапи:
а - розслаблення,
б - з'єднання миозинових головок з
Етапи «крокової» механізму
Послідовні етапи:
а - розслаблення,
б - з'єднання миозинових головок з

Тропонін С, наявний в тонких волокнах, ідентичний кальмодуліном.
Приєднуючи Са2
Тропонін С, наявний в тонких волокнах, ідентичний кальмодуліном.
Приєднуючи Са2

Деполяризація мембрани цистерн відкриває тут Електровозбудімость кальцієві канали.
У зв'язку з тим,
Деполяризація мембрани цистерн відкриває тут Електровозбудімость кальцієві канали.
У зв'язку з тим,

1 - викид медіатора в синаптичну щілина.
2 - звільнення активного центру
1 - викид медіатора в синаптичну щілина.
2 - звільнення активного центру
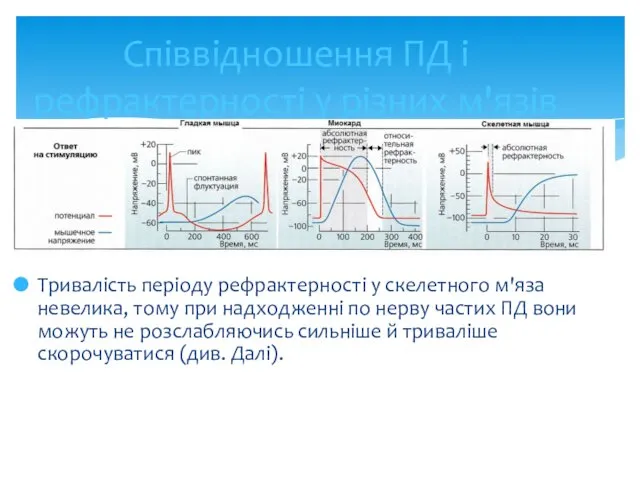
Співвідношення ПД і рефрактерності у різних м'язів
Тривалість періоду рефрактерності у скелетного
Співвідношення ПД і рефрактерності у різних м'язів
Тривалість періоду рефрактерності у скелетного
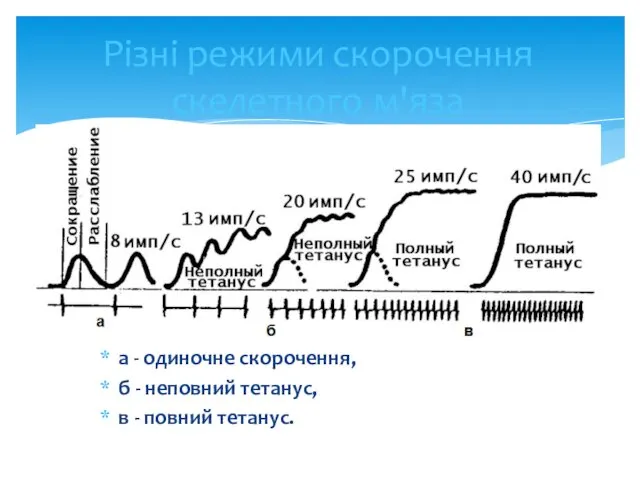
Різні режими скорочення скелетного м'яза
а - одиночне скорочення,
б - неповний тетанус,
в
Різні режими скорочення скелетного м'яза
а - одиночне скорочення,
б - неповний тетанус,
в

а) скорочення (утворення містків);
б) розслаблення (розриву містків);
в) роботи Са2 + -насоса
а) скорочення (утворення містків);
б) розслаблення (розриву містків);
в) роботи Са2 + -насоса
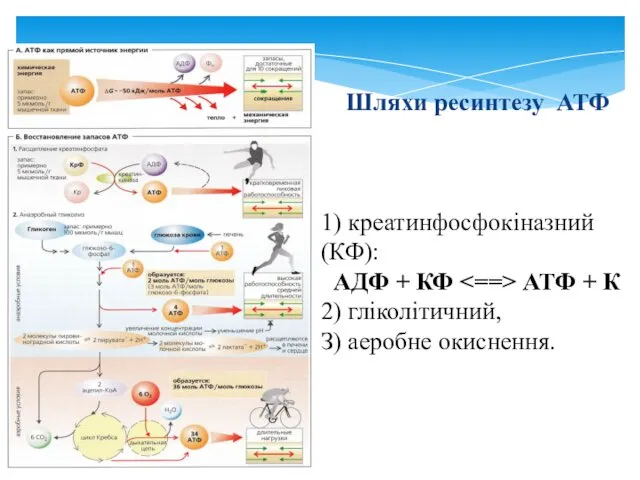
Шляхи ресинтезу АТФ
1) креатинфосфокіназний (КФ):
АДФ + КФ <==> АТФ
Шляхи ресинтезу АТФ
1) креатинфосфокіназний (КФ):
АДФ + КФ <==> АТФ

а) фосфогенний (КФ) - 3,6 моль АТФ/хв,
б) гліколітічний - 1,2
а) фосфогенний (КФ) - 3,6 моль АТФ/хв,
б) гліколітічний - 1,2

Рухова одиниця - одиничне нервове волокно мотонейрона а також іннервуючі ним
Рухова одиниця - одиничне нервове волокно мотонейрона а також іннервуючі ним

Швидкі і повільні ДЕ
Швидкі
Великий мотонейрон (висока швидкість проведення ПД).
Багато АТФ.
Багато
Швидкі і повільні ДЕ
Швидкі
Великий мотонейрон (висока швидкість проведення ПД).
Багато АТФ.
Багато
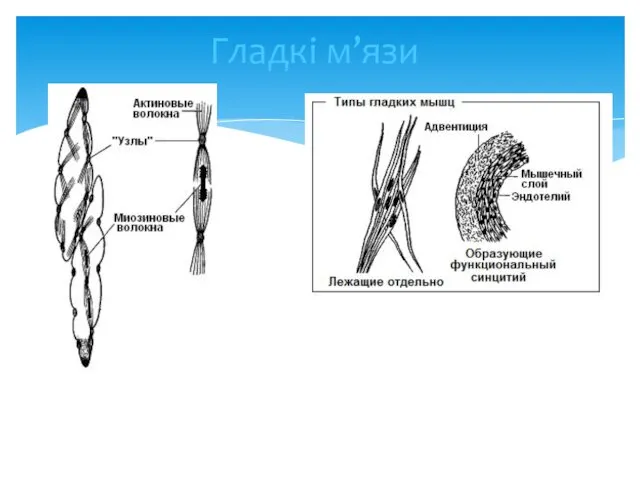
Гладкі м’язи
Гладкі м’язи
 родительское собрание в 4 классе Конфликт как путь развития личности
родительское собрание в 4 классе Конфликт как путь развития личности родительская конференция 378 лицей 13.04
родительская конференция 378 лицей 13.04 Диагностирование системы охлаждения двигателя
Диагностирование системы охлаждения двигателя Презентация Прощай, начальная школа!
Презентация Прощай, начальная школа! Компетентность родителей как условие содействия нравственному и эмоциональному благополучию ребенка.
Компетентность родителей как условие содействия нравственному и эмоциональному благополучию ребенка. “За чашкой чая”. Клуб общения
“За чашкой чая”. Клуб общения Выступление на ШМО на тему: Характеристика изменений в деятельности учащихся и их родителей в условиях введения ФГОС
Выступление на ШМО на тему: Характеристика изменений в деятельности учащихся и их родителей в условиях введения ФГОС 10 правил как научить ребенка слушаться Диск
10 правил как научить ребенка слушаться Диск Афиша презентация к уроку ИЗО в 3классе
Афиша презентация к уроку ИЗО в 3классе Гигиена детей и подростков
Гигиена детей и подростков Биохимия стресса
Биохимия стресса Перевозка полиэтилена из г. Будённовска (РФ) в г. Илкестон (Великобритания)
Перевозка полиэтилена из г. Будённовска (РФ) в г. Илкестон (Великобритания) Технико-экономическая оценка мероприятий по повышению энергоэффективности
Технико-экономическая оценка мероприятий по повышению энергоэффективности Диагностика пневмококковой инфекции
Диагностика пневмококковой инфекции Конкурс Золотая полка книг
Конкурс Золотая полка книг Clothing comparisons flashcards
Clothing comparisons flashcards Виды туризма
Виды туризма Система охлаждения двигателя автомобиля
Система охлаждения двигателя автомобиля Классификация химических реакций
Классификация химических реакций Good times ahead!
Good times ahead! Игра Путешествие по стране Букляндия
Игра Путешествие по стране Букляндия Признаки делимости чисел
Признаки делимости чисел تابع الفصل الثالث برنامج العروض التقديمية
تابع الفصل الثالث برنامج العروض التقديمية Скелетные ткани. Хрящевые ткани
Скелетные ткани. Хрящевые ткани Съемка плана местности.
Съемка плана местности. Социокультурная среда техникума
Социокультурная среда техникума Цифро-аналоговые преобразователи. (Лекция 6)
Цифро-аналоговые преобразователи. (Лекция 6) Метод координат в пространстве
Метод координат в пространстве