- Химия и обмен углеводов

Содержание
- 2. План: ХИМИЯ И ОБМЕН УГЛЕВОДОВ 1. Химия углеводов Классификация Моносахариды Олигосахариды Полисахариды 2. Обмен углеводов Переваривание
- 3. Распад гликогена и освобождение глюкозы (глюкогенез) Гликолиз Спиртовое брожение 3. Включение других углеводов в процесс гликолиза
- 4. Углеводы, наряду с белками и липидами, являются важнейшими химическими соединениями живых организмов. В организме животных и
- 5. Углеводы также используются для синтеза нуклеиновых кислот (рибоза и дезоксирибоза), они являются составными компонентами нуклеотидных коферментов,
- 6. Эти вещества выполняют сложные и важные функции в организме. В составе тела человека и животных углеводы
- 7. Впервые термин "углеводы" был предложен профессором Дерптского (ныне Тартуского) университета К.Г.Шмидтом в 1844 г. В то
- 8. Необходимо отметить, что химия углеводов занимает одно из ведущих мест в истории развития органической химии. Тростниковый
- 9. Следует отметить также, что значительный вклад в изучение углеводов внесли отечественные ученые А.А.Колли, П.П.Шорыгин, Н.К.Кочетков и
- 11. Моносахариды Моносахариды можно рассматривать как производные многоатомных спиртов, содержащих карбонильную (альдегидную или кетонную) группу. Если карбонильная
- 12. Стереохимия моносахаридов. Все моносахариды содержат один или более асимметричных атомов углерода: альдотриоза – один центр асимметрии,
- 13. Изомер глицеральдегида, у которого при проекции модели на плоскость ОН-группа у асимметричного атома углерода расположена с
- 14. Альдогексозы содержат четыре асимметричных атома углерода и могут существовать в виде 24=16 стереоизомеров, представителем которых является
- 15. Свойство вращать плоскость поляризованного луча вправо обозначают знаком (+), а в противоположную сторону – знаком (–).
- 16. Циклические (полуацетальные) формы моносахаридов. Любой моносахарид, обладая рядом конкретных физических свойств (т. пл., растворимость и т.д.),
- 17. Изменение удельного вращения растворов моносахаридов при стоянии (во времени) называется мутаротацией. Очевидно, мутаротация должна вызываться изменением
- 18. α-Форма – это форма, у которой расположение полуацетального гидроксила такое же, как гидроксила у асимметричного углеродного
- 19. При этом удельное вращение, характерное для исходной циклической формы, постепенно меняется, и наконец, устанавливается постоянное удельное
- 20. Находящиеся ближе к читателю связи изображают более жирными линиями (углеродные атомы цикла не пишут). Заместители, расположенные
- 21. Наконец, следует помнить, что при написании структурных формул по Хеуорсу гидроксильная группа при С1 будет расположена
- 22. Проекционные формулы Хеуорса не отражают подлинной конформации моносахаридов. Подобно циклогексану, пиранозное кольцо может принимать две конфигурации
- 23. При действии на β-D-глюкопиранозу уксусной кислоты образуется продукт ацилирования – ацетил-β-D-глюкопиранозид
- 24. Ацилированию и метилированию способны подвергаться и остальные группы моносахаридов, хотя это требует намного более жестких условий.
- 25. К N-гликозидам принадлежат исключительно важные в обмене веществ продукты расщепления нуклеиновых кислот и нуклеотидов (нуклеотиды и
- 26. Другой класс называется альдуроновыми, или уроновыми, кислотами. В уроновых кислотах окислена первичная спиртовая группа, а альдегидная
- 27. Большой интерес представляют пирофосфорные эфиры моносахаридов, например, 5-фосфорибозил-1-пирофосфат (ФРПФ), который участвует в синтезе пуриновых и пиримидиновых
- 28. Аминосахара – производные моносахаридов, гидроксильная группа которых замещена аминогруппой. В зависимости от положения аминогруппы в молекуле
- 29. В организме человека и животных аминосахарами являются D-глкюкозамин и D-галактозамин:
- 30. Аминосахара входят в состав мукополисахаридов животного, растительного и бактериального происхождения, являются углеводными компонентами различных гликопротеинов и
- 32. Олигосахариды Олигосахариды – углеводы, молекулы которых содержат от 2 до 10 остатков моносахаридов, соединенных гликозидными связями.
- 33. при действии амилаз на крахмал (или гликоген), содержит два остатка α-D-глюкозы (название сахара, полуацетальный гидроксил которого
- 34. В отличие от большинства дисахаридов сахароза не имеет свободного полуацетального гидроксила и не обладает восстановительными свойствами.
- 35. Полисахариды Известны две группы полисахаридов: гомополисахариды, состоящие из моносахаридных единиц только одного типа и гетерополисахариды, для
- 36. Крахмал Крахмал представляет собой смесь двух гомополисахаридов: линейного – амилозы и разветвленного – амилопектина, общая формула
- 38. В молекуле амилозы связано в среднем около 1000 остатков глюкозы, отдельные участки молекулы амилопектина состоят из
- 39. Целлюлоза
- 40. Целлюлоза (клетчатка) – наиболее распространенный структурный полисахарид растительного мира. Она состоит из α-глюкозных остатков в их
- 41. Хитин Хитин – структурный полисахарид низших растений, особенно грибов, а также беспозвоночных животных. Состоит из остатков
- 42. Обмен углеводов
- 43. Обмен углеводов складывается из следующих процессов. 1.Расщепление в желудочно-кишечном тракте поступающих с пищей полисахаридов и дисахаридов
- 44. 2.Синтез и распад гликогена в тканях, прежде всего, в печени. 3.Анаэробное и аэробное расщепление глюкозы. В
- 45. Переваривание и всасывание Расщепление крахмала (и гликогена) начинается в ротовой полости под действием амилазы слюны. Имеются
- 46. Под действием α-амилазы проходят первые фазы распада крахмала (гликогена) с образованием декстринов. Затем пища, смешиваясь со
- 47. Образующаяся мальтоза оказывается только временным продуктом, так как она быстро гидролизуется под влиянием фермента мальтазы (α-глюкозидазы)
- 48. Синтез гликогена Считалось, что гликоген-фосфорилаза (фосфорилаза а) катализирует как распад гликогена, так и его синтез, потому
- 49. Образовавшийся глюкозо-1-фосфат уже непосредственно участвует в синтезе гликогена. На первой стадии синтеза он вступает во взаимодействие
- 50. Данная реакция катализируется ферментом глюкозо-1-фосфатуридин-трансферазой (УДФГ-пирофосфорилазой).
- 51. На второй стадии, приводящей к образованию гликогена, происходит перенос глюкозного остатка, входящего в состав УДФ-глюкозы, на
- 52. Образующийся УДФ затем вновь фосфорилируется в УТФ за счет АТФ, и таким образом, весь цикл превращений
- 53. (С6Н10О5)п + Н3РО4 (С6Н10О5)п-1 + глюкозо-1-фосфат Фермент фосфорилаза существует в двух формах, одна из которых (фосфорилаза
- 55. Образование свободной глюкозы из глюкозо-6-фосфата в печени происходит под влиянием глюкозо-6-фосфатазы. Данный фермент катализирует гидролитическое отщепление
- 56. Гликолиз
- 57. Гликолиз (glycys – сладкий и lysis – растворение, распад) – сложный ферментативный процесс последовательных превращений глюкозы,
- 58. сопровождается освобождением значительного количества свободной энергии. 2 реакция – превращение во фруктозо-6-фосфат (в присутствии кофакторов). 3
- 59. Это наиболее медленная стадия гликолиза. 4 реакция катализируется ферментом альдолазой. При этом образуются 2 фосфотриозы. Реакция
- 60. Равновесие данной изомеразной реакции сдвинуто в сторону диоксиацетонфосфата: 95 % диоксиацетонфосфата и около 5 % глицеральдегид-3-фосфата.
- 61. Это соединение представляет собой высокоэнергетическое соединение. 7 реакция катализируется фосфоглицераткиназой 8 реакция сопровождается внутримолекулярным переносом оставшейся
- 62. 9 реакция катализируется ферментом енолазой, при этом 2-фосфо-глицериновая кислота переходит в фосфоенолпировиноградную кислоту (фосфоенолпируват), а фосфатная
- 63. Для действия пируваткиназы необходимы ионы Mg2+, а также одновалентные катионы металлов (К или др.). Внутри клетки
- 64. Гликогенолиз Процесс анаэробного распада гликогена получил название гликогенолиза. Вовлечение D-глюкозных единиц гликогена в процесс гликолиза происходит
- 65. Спиртовое брожение
- 66. Спиртовое брожение осуществляется так называемыми дрожжеподобными организмами, а также некоторыми плесневыми грибами С6Н12О6 2 С2Н5ОН +
- 67. восстанавливаясь при этом в этанол, реакция катализируется ферментом алкогольдегидрогеназой Н2 СН3СНО СН3СН2ОН Таким образом, конечными продуктами
- 69. Галактоза. Основной источник – лактоза пищи. Ее брожение
- 70. Глюконеогенез
- 71. Глюконеогенез – синтез глюкозы из неуглеводных продуктов. Такими продуктами, или метаболитами, являются в первую очередь молочная
- 73. Между гликолизом, интенсивно протекающим в мышечной ткани при ее активной деятельности, и глюконеогенезом, особенно характерным для
- 74. Окислительное декарбоксилирование пировиноградной кислоты Окисление пирувата до ацетил-КоА (коэнзим-А) происходит при участии ряда ферментов и коферментов,
- 75. На четвертой стадии регенерируется окисленная форма липоамида из восстановленного комплекса дигидролипоамида-E1. При участии фермента дигидролипоилдегидрогеназы (E3)
- 76. Образовавшийся в процессе окислительного декарбоксилирования ацетил-КоА подвергается дальнейшему окислению с образованием в конечном итоге СО2 и
- 77. Цикл трикарбоновых кислот (цикл Кребса) Цикл Кребса – общий конечный путь окисления ацетильных групп (в виде
- 78. Первая реакция катализируется ферментом нитратсинтетазой. При этом ацетильная группа ацетил-КоА конденсируется с оксалоацетатом, в результате чего
- 79. В результате второй реакции образовавшаяся лимонная кислота подвергается дегидратированию с образованием цис-аконитовой кислоты, которая, присоединяя молекулу
- 80. В ходе изоцитратдегидрогеназной реакции изолимонная кислота одновременно декарбоксилируется. Кроме того, фермент для проявления своей активности нуждается
- 81. Шестая реакция – сукцинат дегидрируется в фумаровую кислоту. Седьмая реакция осуществляется под влиянием фермента фумаратгидратазы (фумаразы).
- 82. В ходе восьмой реакции цикла трикарбоновых кислот под влиянием митохондриапьной НАД-зависимой малатдегидрогеназы происходит окисление L-малата в
- 83. Пентозофосфатный путь окисления углеводов Расхождение путей окисления углеводов – классического (цикл Кребса) и пентозофосфатного – начинается
- 87. Регуляция углеводного обмена
- 88. Регуляция углеводного обмена Факторы, влияющие на активность ферментов, участвующих в реакциях углеводного обмена: концентрация субстратов, содержание
- 89. При некоторых состояниях можно наблюдать повышение содержания глюкозы в крови – гипергликемия, а также понижение концентрации
- 90. В печени при общем снижении интенсивности биосинтетических процессов (биосинтеза белков, синтеза жирных кислот из продуктов распада
- 91. А — уровень глюкозы в крови натощак; В — максимальное содержание глюкозы в крови после нагрузки
- 93. Скачать презентацию

План:
ХИМИЯ И ОБМЕН УГЛЕВОДОВ
1. Химия углеводов
Классификация
Моносахариды
Олигосахариды
Полисахариды
2. Обмен углеводов
Переваривание и всасывание углеводов
Синтез
План:
ХИМИЯ И ОБМЕН УГЛЕВОДОВ
1. Химия углеводов
Классификация
Моносахариды
Олигосахариды
Полисахариды
2. Обмен углеводов
Переваривание и всасывание углеводов
Синтез

Распад гликогена и освобождение глюкозы (глюкогенез)
Гликолиз
Спиртовое брожение
3. Включение других углеводов в
Распад гликогена и освобождение глюкозы (глюкогенез)
Гликолиз
Спиртовое брожение
3. Включение других углеводов в

Углеводы, наряду с белками и липидами, являются важнейшими химическими соединениями живых
Углеводы, наряду с белками и липидами, являются важнейшими химическими соединениями живых

Углеводы также используются для синтеза нуклеиновых кислот (рибоза и дезоксирибоза), они
Углеводы также используются для синтеза нуклеиновых кислот (рибоза и дезоксирибоза), они
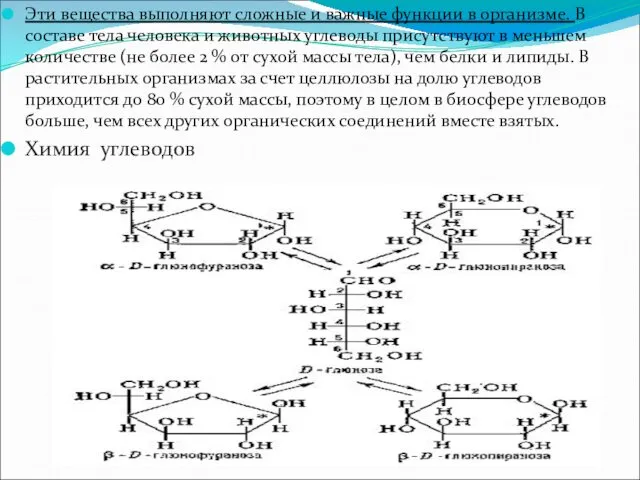
Эти вещества выполняют сложные и важные функции в организме. В составе
Эти вещества выполняют сложные и важные функции в организме. В составе
Впервые термин "углеводы" был предложен профессором Дерптского (ныне Тартуского) университета К.Г.Шмидтом
Впервые термин "углеводы" был предложен профессором Дерптского (ныне Тартуского) университета К.Г.Шмидтом

Необходимо отметить, что химия углеводов
занимает одно из ведущих мест в
Необходимо отметить, что химия углеводов
занимает одно из ведущих мест в

Следует
отметить также, что значительный вклад в
изучение углеводов внесли отечественные ученые
Следует
отметить также, что значительный вклад в
изучение углеводов внесли отечественные ученые
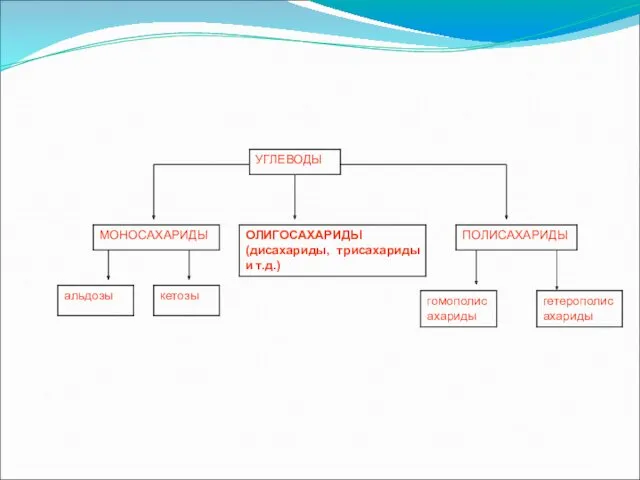

Моносахариды
Моносахариды можно рассматривать как производные многоатомных спиртов, содержащих карбонильную (альдегидную
Моносахариды
Моносахариды можно рассматривать как производные многоатомных спиртов, содержащих карбонильную (альдегидную

Стереохимия моносахаридов. Все моносахариды содержат один или более асимметричных атомов углерода:
Стереохимия моносахаридов. Все моносахариды содержат один или более асимметричных атомов углерода:
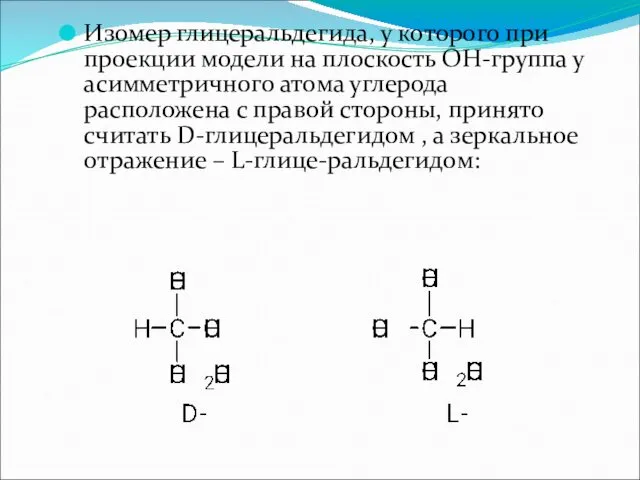
Изомер глицеральдегида, у которого при проекции модели на плоскость ОН-группа у
Изомер глицеральдегида, у которого при проекции модели на плоскость ОН-группа у

Альдогексозы содержат четыре асимметричных атома углерода и могут существовать в виде
Альдогексозы содержат четыре асимметричных атома углерода и могут существовать в виде

Свойство вращать плоскость поляризованного луча вправо обозначают знаком (+), а в
Свойство вращать плоскость поляризованного луча вправо обозначают знаком (+), а в

Циклические (полуацетальные) формы моносахаридов. Любой моносахарид, обладая рядом конкретных физических свойств
Циклические (полуацетальные) формы моносахаридов. Любой моносахарид, обладая рядом конкретных физических свойств

Изменение удельного вращения растворов моносахаридов при стоянии (во времени) называется мутаротацией.
Изменение удельного вращения растворов моносахаридов при стоянии (во времени) называется мутаротацией.

α-Форма – это форма, у которой расположение полуацетального гидроксила такое же,
α-Форма – это форма, у которой расположение полуацетального гидроксила такое же,

При этом удельное вращение, характерное для исходной циклической формы, постепенно меняется,
При этом удельное вращение, характерное для исходной циклической формы, постепенно меняется,

Находящиеся ближе к читателю связи изображают более жирными линиями (углеродные атомы
Находящиеся ближе к читателю связи изображают более жирными линиями (углеродные атомы

Наконец, следует помнить, что при написании структурных формул по Хеуорсу гидроксильная
Наконец, следует помнить, что при написании структурных формул по Хеуорсу гидроксильная

Проекционные формулы Хеуорса не отражают подлинной конформации моносахаридов. Подобно циклогексану, пиранозное
Проекционные формулы Хеуорса не отражают подлинной конформации моносахаридов. Подобно циклогексану, пиранозное
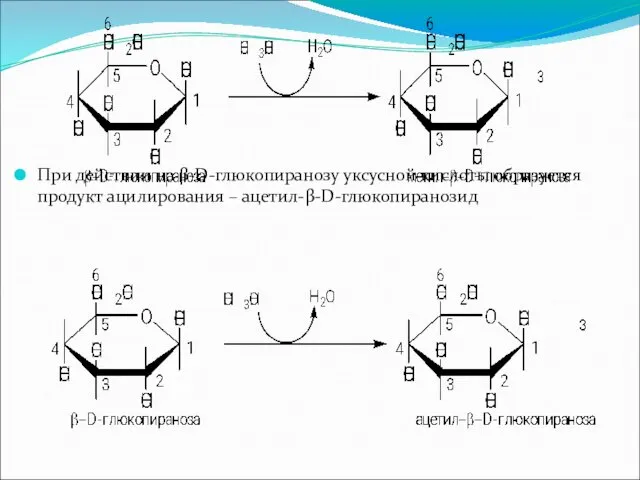
При действии на β-D-глюкопиранозу уксусной кислоты образуется продукт ацилирования – ацетил-β-D-глюкопиранозид
При действии на β-D-глюкопиранозу уксусной кислоты образуется продукт ацилирования – ацетил-β-D-глюкопиранозид

Ацилированию и метилированию способны подвергаться и остальные группы моносахаридов, хотя это
Ацилированию и метилированию способны подвергаться и остальные группы моносахаридов, хотя это

К N-гликозидам принадлежат исключительно важные в обмене веществ продукты расщепления нуклеиновых
К N-гликозидам принадлежат исключительно важные в обмене веществ продукты расщепления нуклеиновых

Другой класс называется альдуроновыми, или уроновыми, кислотами. В уроновых кислотах окислена
Другой класс называется альдуроновыми, или уроновыми, кислотами. В уроновых кислотах окислена

Большой интерес представляют пирофосфорные эфиры моносахаридов, например, 5-фосфорибозил-1-пирофосфат (ФРПФ), который участвует
Большой интерес представляют пирофосфорные эфиры моносахаридов, например, 5-фосфорибозил-1-пирофосфат (ФРПФ), который участвует
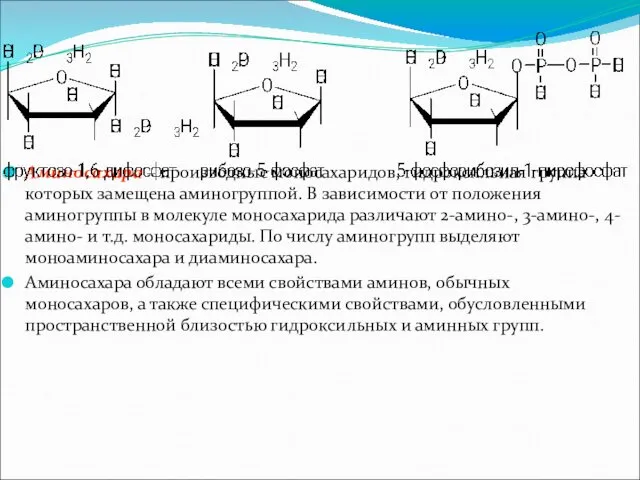
Аминосахара – производные моносахаридов, гидроксильная группа которых замещена аминогруппой. В зависимости
Аминосахара – производные моносахаридов, гидроксильная группа которых замещена аминогруппой. В зависимости
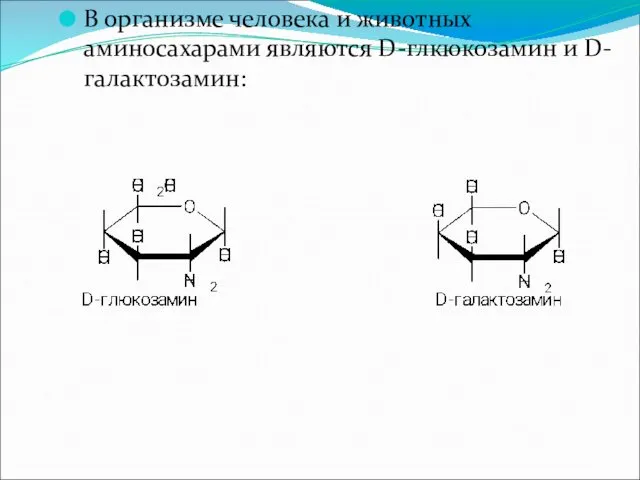
В организме человека и животных аминосахарами являются D-глкюкозамин и D-галактозамин:
В организме человека и животных аминосахарами являются D-глкюкозамин и D-галактозамин:

Аминосахара входят в состав мукополисахаридов животного, растительного и бактериального происхождения, являются


Олигосахариды
Олигосахариды – углеводы, молекулы которых содержат от 2 до 10 остатков
Олигосахариды
Олигосахариды – углеводы, молекулы которых содержат от 2 до 10 остатков
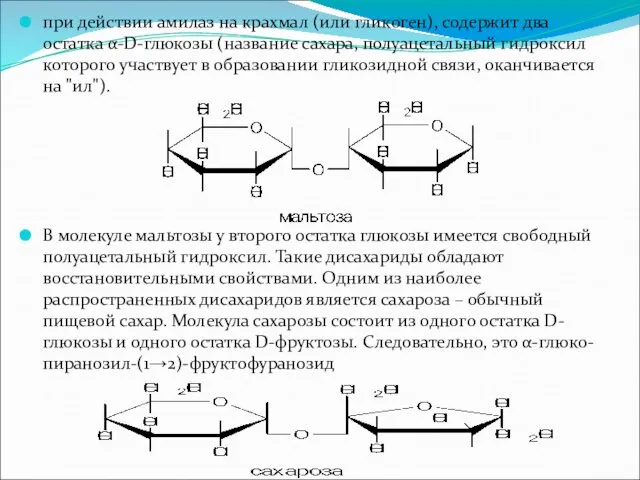
при действии амилаз на крахмал (или гликоген), содержит два остатка α-D-глюкозы
при действии амилаз на крахмал (или гликоген), содержит два остатка α-D-глюкозы

В отличие от большинства дисахаридов сахароза не имеет свободного полуацетального гидроксила
В отличие от большинства дисахаридов сахароза не имеет свободного полуацетального гидроксила

Полисахариды
Известны две группы полисахаридов: гомополисахариды, состоящие из моносахаридных единиц только одного
Полисахариды
Известны две группы полисахаридов: гомополисахариды, состоящие из моносахаридных единиц только одного

Крахмал
Крахмал представляет собой смесь двух гомополисахаридов: линейного – амилозы и
Крахмал
Крахмал представляет собой смесь двух гомополисахаридов: линейного – амилозы и
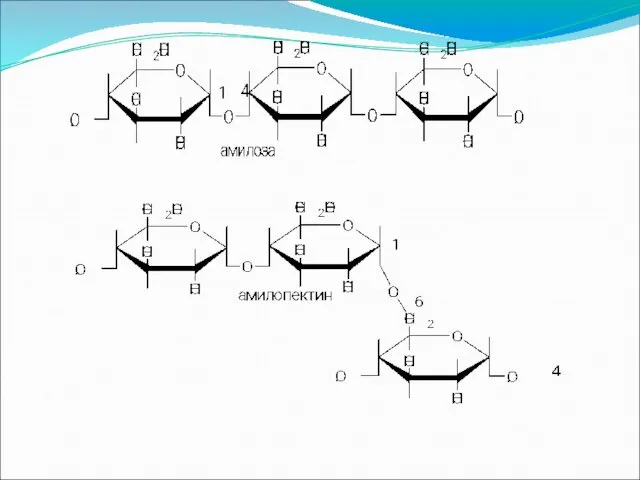

В молекуле амилозы связано в среднем около 1000 остатков глюкозы, отдельные
В молекуле амилозы связано в среднем около 1000 остатков глюкозы, отдельные

Целлюлоза
Целлюлоза

Целлюлоза (клетчатка) – наиболее распространенный структурный полисахарид растительного мира. Она состоит
Целлюлоза (клетчатка) – наиболее распространенный структурный полисахарид растительного мира. Она состоит
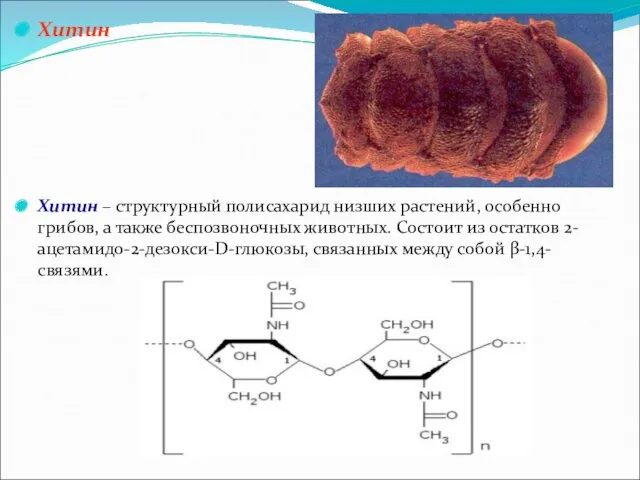
Хитин
Хитин – структурный полисахарид низших растений, особенно грибов, а также
Хитин
Хитин – структурный полисахарид низших растений, особенно грибов, а также
Обмен углеводов
Обмен углеводов
Обмен углеводов складывается из следующих процессов.
1.Расщепление в желудочно-кишечном тракте поступающих с
Обмен углеводов складывается из следующих процессов.
1.Расщепление в желудочно-кишечном тракте поступающих с

2.Синтез и распад гликогена в тканях, прежде всего, в печени.
2.Синтез и распад гликогена в тканях, прежде всего, в печени.

Переваривание и всасывание
Расщепление крахмала (и гликогена) начинается в ротовой полости под
Переваривание и всасывание
Расщепление крахмала (и гликогена) начинается в ротовой полости под

Под действием α-амилазы проходят первые фазы распада крахмала (гликогена) с образованием
Под действием α-амилазы проходят первые фазы распада крахмала (гликогена) с образованием

Образующаяся мальтоза оказывается только временным продуктом, так как она быстро гидролизуется
Образующаяся мальтоза оказывается только временным продуктом, так как она быстро гидролизуется

Синтез гликогена
Считалось, что гликоген-фосфорилаза (фосфорилаза а) катализирует как распад гликогена, так
Синтез гликогена
Считалось, что гликоген-фосфорилаза (фосфорилаза а) катализирует как распад гликогена, так
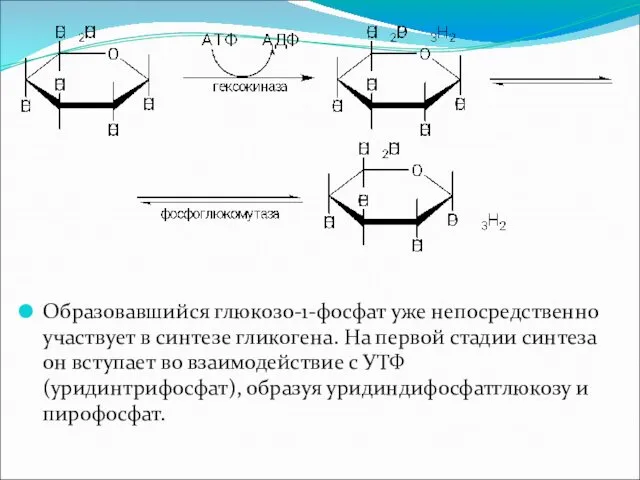
Образовавшийся глюкозо-1-фосфат уже непосредственно участвует в синтезе гликогена. На первой стадии
Образовавшийся глюкозо-1-фосфат уже непосредственно участвует в синтезе гликогена. На первой стадии
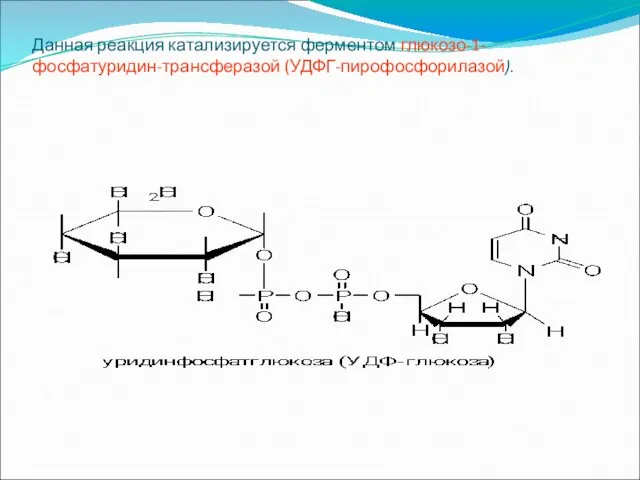
Данная реакция катализируется ферментом глюкозо-1-фосфатуридин-трансферазой (УДФГ-пирофосфорилазой).
Данная реакция катализируется ферментом глюкозо-1-фосфатуридин-трансферазой (УДФГ-пирофосфорилазой).
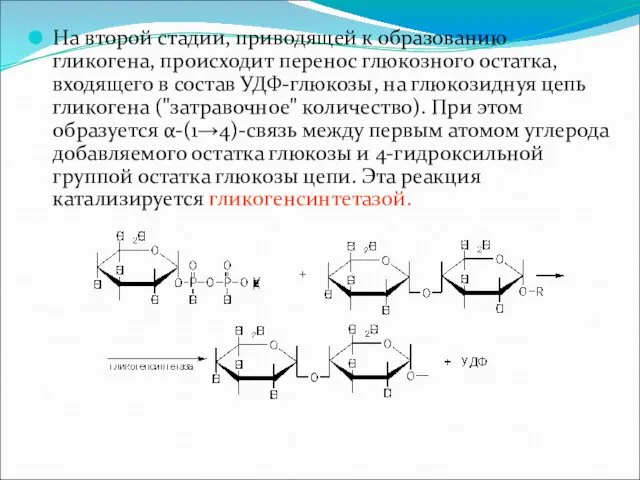
На второй стадии, приводящей к образованию гликогена, происходит перенос глюкозного остатка,
На второй стадии, приводящей к образованию гликогена, происходит перенос глюкозного остатка,

Образующийся УДФ затем вновь фосфорилируется в УТФ за счет АТФ, и
Образующийся УДФ затем вновь фосфорилируется в УТФ за счет АТФ, и

(С6Н10О5)п + Н3РО4 (С6Н10О5)п-1 + глюкозо-1-фосфат
Фермент фосфорилаза существует в двух формах,
(С6Н10О5)п + Н3РО4 (С6Н10О5)п-1 + глюкозо-1-фосфат
Фермент фосфорилаза существует в двух формах,


Образование свободной глюкозы из глюкозо-6-фосфата в печени происходит под влиянием глюкозо-6-фосфатазы.
Образование свободной глюкозы из глюкозо-6-фосфата в печени происходит под влиянием глюкозо-6-фосфатазы.
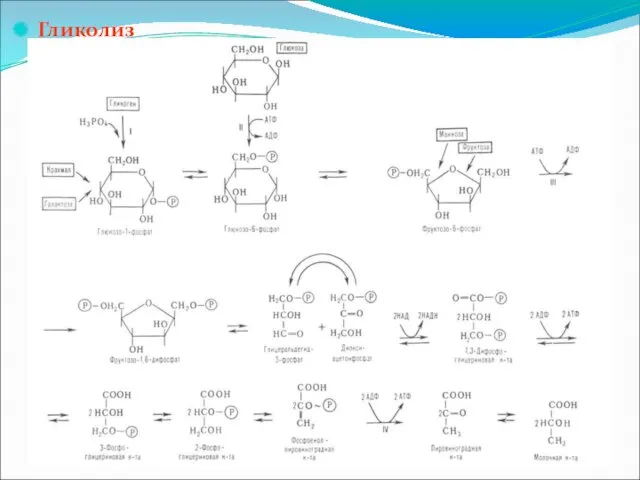
Гликолиз
Гликолиз
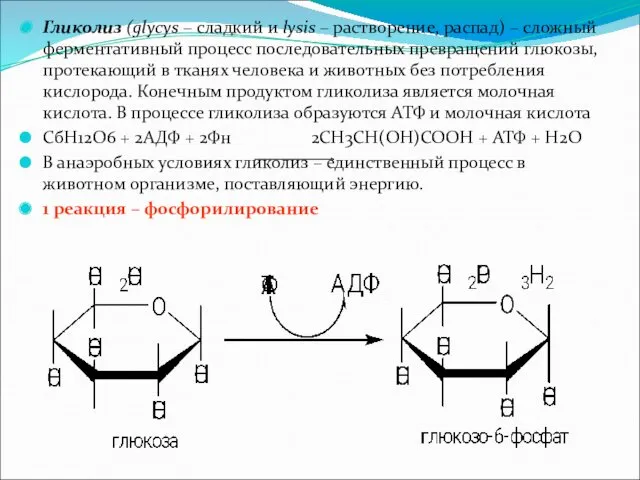
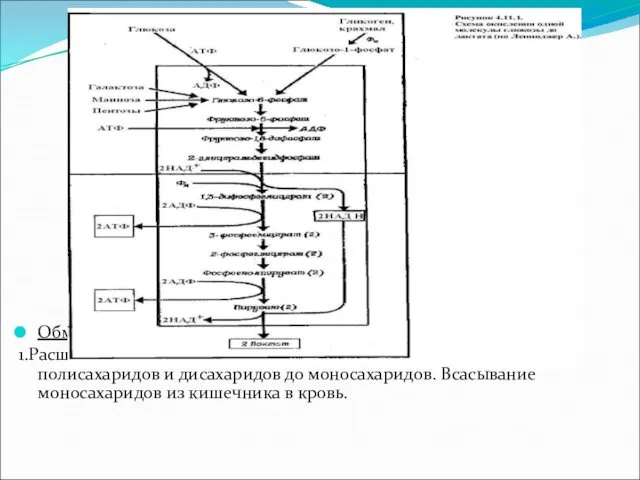
Гликолиз (glycys – сладкий и lysis – растворение, распад) – сложный
Гликолиз (glycys – сладкий и lysis – растворение, распад) – сложный
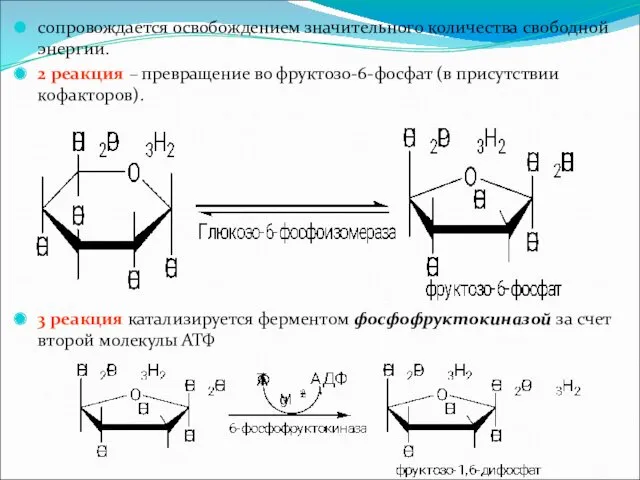
сопровождается освобождением значительного количества свободной энергии.
2 реакция – превращение во фруктозо-6-фосфат
сопровождается освобождением значительного количества свободной энергии.
2 реакция – превращение во фруктозо-6-фосфат

Это наиболее медленная стадия гликолиза.
4 реакция катализируется ферментом альдолазой. При этом
Это наиболее медленная стадия гликолиза.
4 реакция катализируется ферментом альдолазой. При этом

Равновесие данной изомеразной реакции сдвинуто в сторону диоксиацетонфосфата: 95 % диоксиацетонфосфата и
Равновесие данной изомеразной реакции сдвинуто в сторону диоксиацетонфосфата: 95 % диоксиацетонфосфата и

Это соединение представляет собой высокоэнергетическое соединение.
7 реакция катализируется фосфоглицераткиназой
8
Это соединение представляет собой высокоэнергетическое соединение.
7 реакция катализируется фосфоглицераткиназой
8
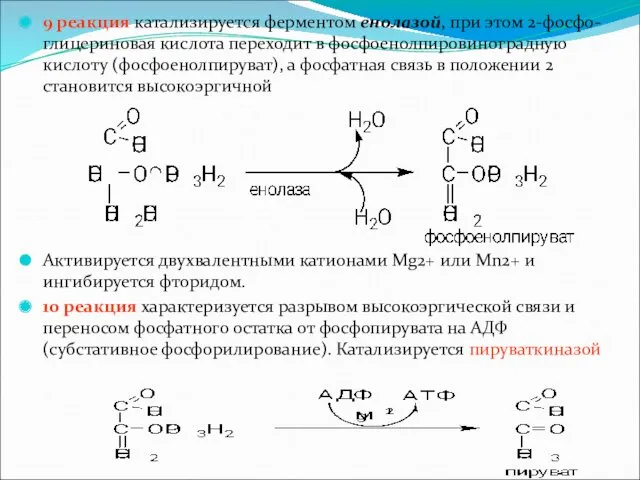
9 реакция катализируется ферментом енолазой, при этом 2-фосфо-глицериновая кислота переходит в
9 реакция катализируется ферментом енолазой, при этом 2-фосфо-глицериновая кислота переходит в

Для действия пируваткиназы необходимы ионы Mg2+, а также одновалентные катионы металлов
Для действия пируваткиназы необходимы ионы Mg2+, а также одновалентные катионы металлов

Гликогенолиз
Процесс анаэробного распада гликогена получил название гликогенолиза. Вовлечение D-глюкозных единиц гликогена
Гликогенолиз
Процесс анаэробного распада гликогена получил название гликогенолиза. Вовлечение D-глюкозных единиц гликогена

Спиртовое брожение
Спиртовое брожение

Спиртовое брожение осуществляется так называемыми дрожжеподобными организмами, а также некоторыми плесневыми
Спиртовое брожение осуществляется так называемыми дрожжеподобными организмами, а также некоторыми плесневыми

восстанавливаясь при этом в этанол, реакция катализируется ферментом алкогольдегидрогеназой
Н2
СН3СНО
восстанавливаясь при этом в этанол, реакция катализируется ферментом алкогольдегидрогеназой
Н2
СН3СНО
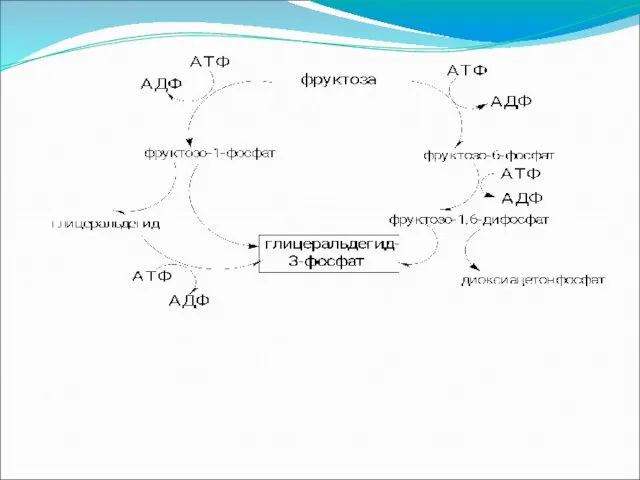
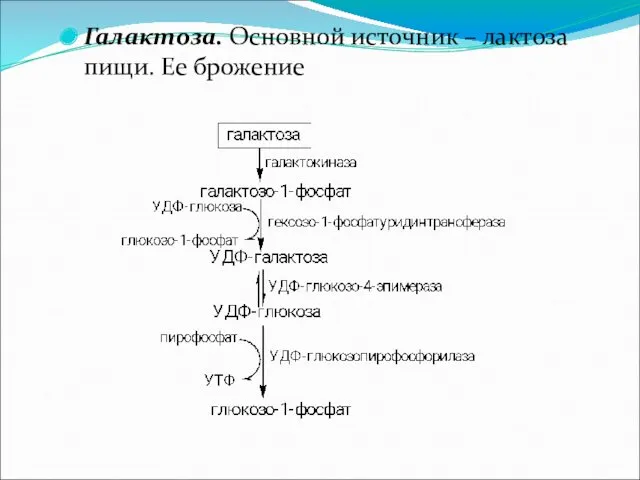
Галактоза. Основной источник – лактоза пищи. Ее брожение
Галактоза. Основной источник – лактоза пищи. Ее брожение
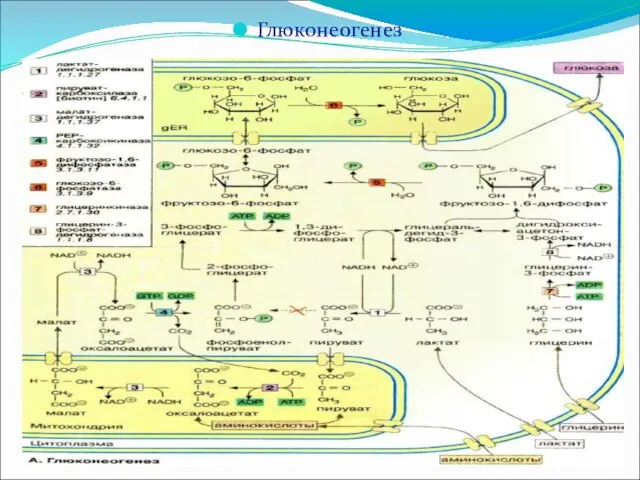
Глюконеогенез
Глюконеогенез

Глюконеогенез – синтез глюкозы из неуглеводных продуктов. Такими продуктами, или метаболитами,
Глюконеогенез – синтез глюкозы из неуглеводных продуктов. Такими продуктами, или метаболитами,


Между гликолизом, интенсивно протекающим в мышечной ткани при ее активной деятельности,
Между гликолизом, интенсивно протекающим в мышечной ткани при ее активной деятельности,

Окислительное декарбоксилирование пировиноградной кислоты
Окисление пирувата до ацетил-КоА (коэнзим-А) происходит при участии
Окислительное декарбоксилирование пировиноградной кислоты
Окисление пирувата до ацетил-КоА (коэнзим-А) происходит при участии

На четвертой стадии регенерируется окисленная форма липоамида из восстановленного комплекса дигидролипоамида-E1.
На четвертой стадии регенерируется окисленная форма липоамида из восстановленного комплекса дигидролипоамида-E1.
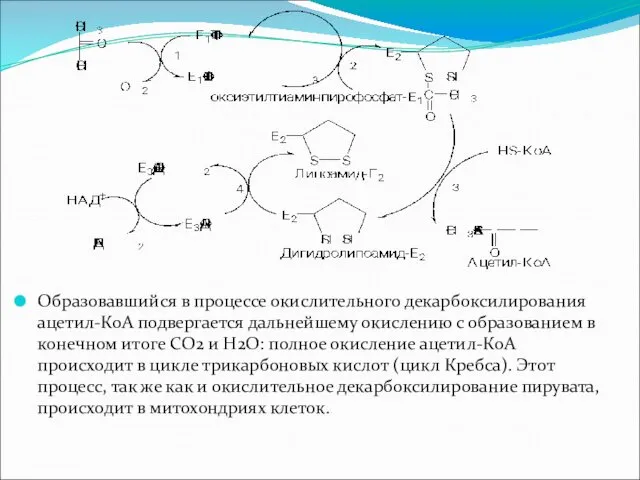
Образовавшийся в процессе окислительного декарбоксилирования ацетил-КоА подвергается дальнейшему окислению с образованием
Образовавшийся в процессе окислительного декарбоксилирования ацетил-КоА подвергается дальнейшему окислению с образованием

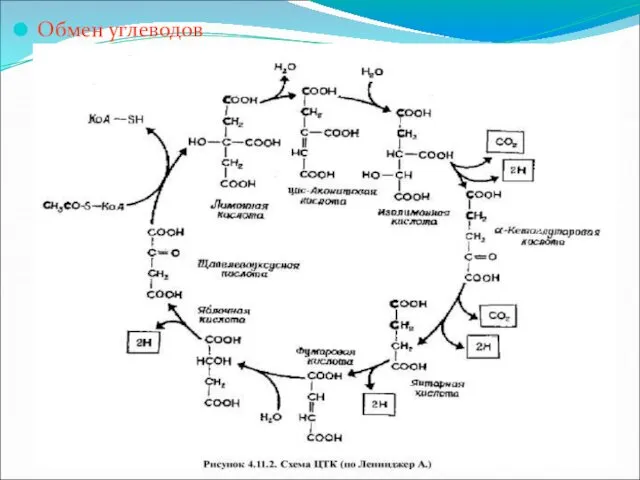
Цикл трикарбоновых кислот (цикл Кребса)
Цикл Кребса – общий конечный путь окисления
Цикл трикарбоновых кислот (цикл Кребса)
Цикл Кребса – общий конечный путь окисления
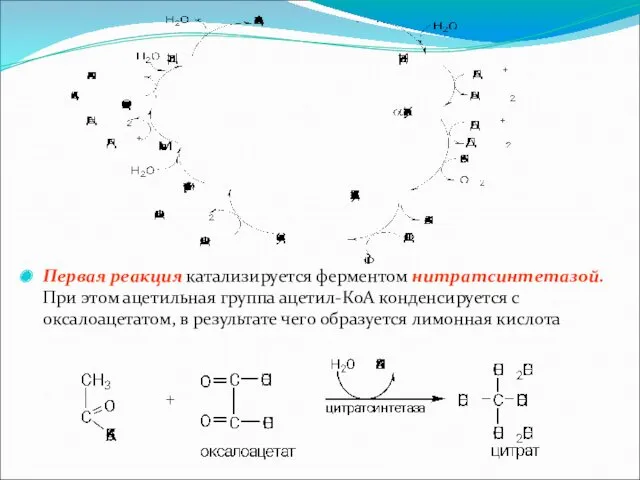
Первая реакция катализируется ферментом нитратсинтетазой. При этом ацетильная группа ацетил-КоА конденсируется
Первая реакция катализируется ферментом нитратсинтетазой. При этом ацетильная группа ацетил-КоА конденсируется

В результате второй реакции образовавшаяся лимонная кислота подвергается дегидратированию с образованием
В результате второй реакции образовавшаяся лимонная кислота подвергается дегидратированию с образованием

В ходе изоцитратдегидрогеназной реакции изолимонная кислота одновременно декарбоксилируется. Кроме того, фермент
В ходе изоцитратдегидрогеназной реакции изолимонная кислота одновременно декарбоксилируется. Кроме того, фермент
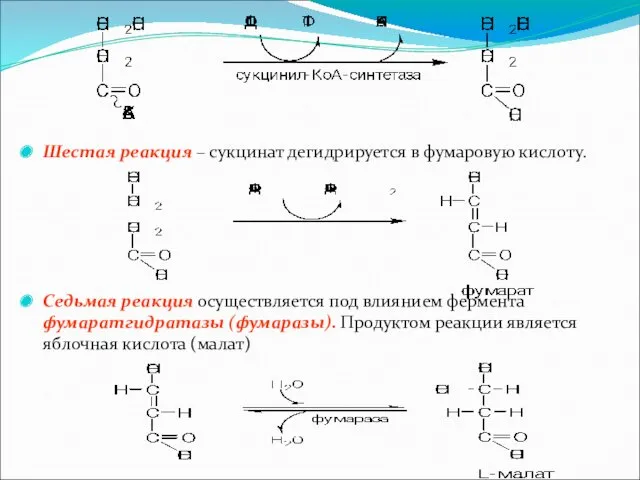
Шестая реакция – сукцинат дегидрируется в фумаровую кислоту.
Седьмая реакция осуществляется под
Шестая реакция – сукцинат дегидрируется в фумаровую кислоту.
Седьмая реакция осуществляется под

В ходе восьмой реакции цикла трикарбоновых кислот под влиянием митохондриапьной НАД-зависимой
В ходе восьмой реакции цикла трикарбоновых кислот под влиянием митохондриапьной НАД-зависимой

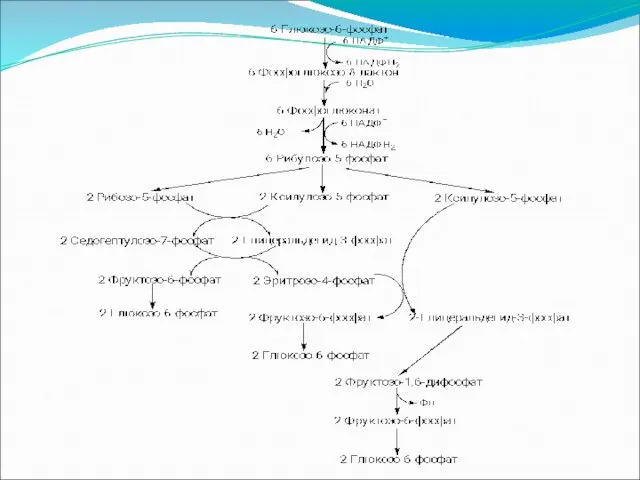
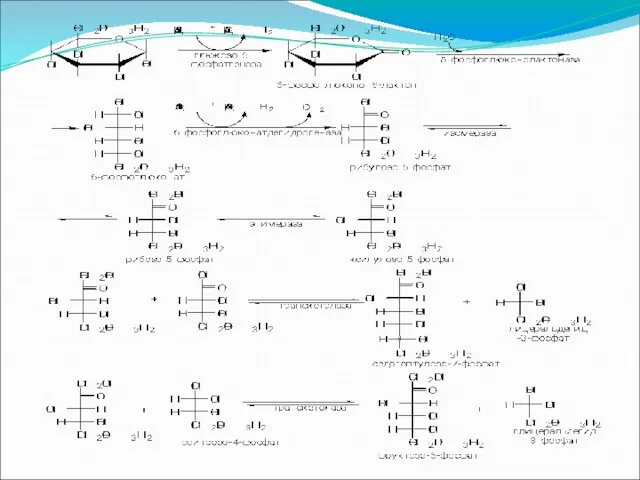
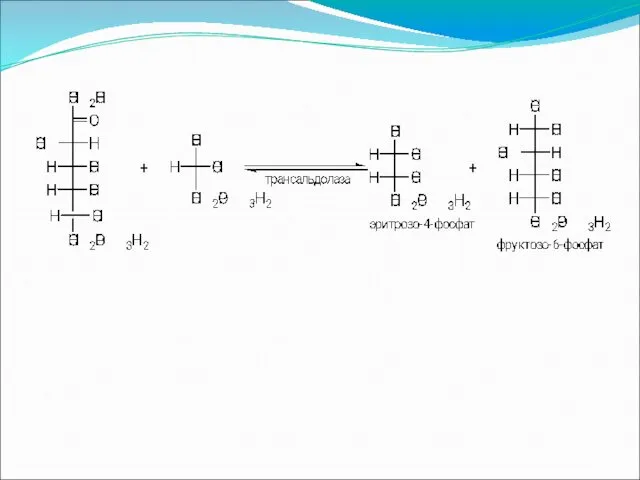
Пентозофосфатный путь окисления углеводов
Расхождение путей окисления углеводов – классического (цикл Кребса)
Пентозофосфатный путь окисления углеводов
Расхождение путей окисления углеводов – классического (цикл Кребса)
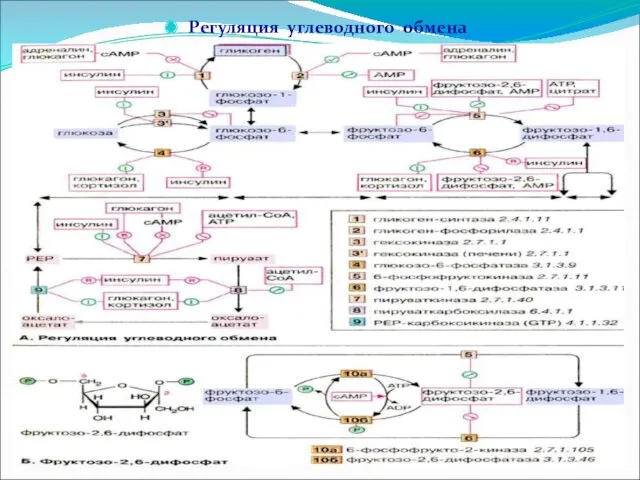
Регуляция углеводного обмена
Регуляция углеводного обмена

Регуляция углеводного обмена
Факторы, влияющие на активность ферментов, участвующих в реакциях углеводного
Регуляция углеводного обмена
Факторы, влияющие на активность ферментов, участвующих в реакциях углеводного

При некоторых состояниях можно наблюдать повышение содержания глюкозы в крови –
При некоторых состояниях можно наблюдать повышение содержания глюкозы в крови –

В печени при общем снижении интенсивности биосинтетических процессов (биосинтеза белков, синтеза
В печени при общем снижении интенсивности биосинтетических процессов (биосинтеза белков, синтеза

А — уровень глюкозы в крови натощак;
В — максимальное содержание глюкозы
А — уровень глюкозы в крови натощак;
В — максимальное содержание глюкозы
 Инструкция для участника итогового сочинения
Инструкция для участника итогового сочинения Ремонт зубчатых колес коробки подач токарных станков
Ремонт зубчатых колес коробки подач токарных станков Визитка группы раннего возраста Ладушки
Визитка группы раннего возраста Ладушки Лето, Лето к нам пришло! Средняя группа Ласточка
Лето, Лето к нам пришло! Средняя группа Ласточка Основы рекреологии
Основы рекреологии Типы химических реакций
Типы химических реакций Бытие 1-2. Сотворение и его значение
Бытие 1-2. Сотворение и его значение Обмен веществ
Обмен веществ Использование многослойных лоскутов при восстановительной хирургии в ЧЛО. Принципы микрососудистой хирургии в ЧЛО
Использование многослойных лоскутов при восстановительной хирургии в ЧЛО. Принципы микрососудистой хирургии в ЧЛО Знакомство с жителями Страны Чувств
Знакомство с жителями Страны Чувств Взаимодействие токов. Магнитное поле. Вектор магнитной индукции. Линии магнитной индукции
Взаимодействие токов. Магнитное поле. Вектор магнитной индукции. Линии магнитной индукции Знакомый и незнакомый картофель
Знакомый и незнакомый картофель Обыкновенные дроби
Обыкновенные дроби Совместная деятельность
Совместная деятельность Презентация Руссая народная кукла. Домашние обрядовые куклы
Презентация Руссая народная кукла. Домашние обрядовые куклы Презентация к внеклассному мероприятию Изготовление поделки РАДУГА
Презентация к внеклассному мероприятию Изготовление поделки РАДУГА Аппаратное обеспечение компьютера
Аппаратное обеспечение компьютера Права ребёнка. Презентация.
Права ребёнка. Презентация. Оперативная память
Оперативная память Россия советская и постсоветская 1953 – 2008 гг
Россия советская и постсоветская 1953 – 2008 гг Донецко-Криворожская республика. Артем(Ф.А.Сергеев)
Донецко-Криворожская республика. Артем(Ф.А.Сергеев) Аналоговый датчик линии
Аналоговый датчик линии Я Научу Тебя Творить
Я Научу Тебя Творить Собака - лучший друг человека!
Собака - лучший друг человека! Формы взаимодействия психолога с коллективом
Формы взаимодействия психолога с коллективом Безопасность, гигиена, эргономика, ресурсосбережение. Эксплуатационные требования к компьютерному рабочему месту
Безопасность, гигиена, эргономика, ресурсосбережение. Эксплуатационные требования к компьютерному рабочему месту Игра - путешествие
Игра - путешествие Правовой режим земель населенных пунктов
Правовой режим земель населенных пунктов