- Метаболизм нуклеотидов

Содержание
- 2. Вопросы из билетов по теме Биосинтез и распад пиримидиновых нуклеотидов. Источники атомов пуринового кольца. Распад пуриновых
- 3. Структура темы Классификация нуклеотидов Пуриновые нуклеотиды Синтез Распад Нарушения распада и синтеза: подагра Пиримидиновые нуклеотиды Синтез
- 4. Структура нуклеотида (АТФ)
- 5. Классификация азотистых оснований Пурины: Аденин Гуанин Пиримидины Урацил Цитозин Тимин У пуринов 2 кольца: 6-членное 5-
- 7. СИНТЕЗ ПУРИНОВ
- 8. Два пути синтеза нуклеотидов de novo (с нуля): из простых предшественников Запасной путь (путь спасения): из
- 9. СИНТЕЗ DE NOVO
- 10. Синтез de novo: источники атомов пуринового кольца Глутамин (амидный азот) Глицин ТГФК (одноуглеродный фрагмент) Аспартат (азот
- 11. Особенности синтеза Пуриновое кольцо строится постепенным добавлением новых атомов в строго определенном порядке Пуриновое кольцо строится
- 12. Синтез 1-я реакция: образование ФРПФ (фосфорибозилпирофосфат) из рибозо-5-фосфата (см. пентозофосфатный путь) и АТФ. Фермент: ФРПФ-синтаза (регуляторный
- 13. 1-я реакция: образование фосфорибозилпирофосфата (ФРПФ = ФРДФ) Фермент: фосфорибозилпирофосфатсинтетаза (ФРПФсинтаза) Это регуляторный фермент синтеза пуриновых нуклеотидов
- 14. Дальнейшие стадии синтеза, до ИМФ
- 15. Синтез АМФ и ГМФ из ИМФ
- 16. Регуляция синтеза de novo Основной регуляторный фермент: ФРПФ-синтетаза Другие регуляторные ферменты: глутамин-фосфорибозиламидотрансфераза, ИМФ-дегидрогеназа, аденилосукцинатсинтаза ФРПФ-синтетатза ингибируется
- 17. Регуляторные ферменты ФРПФ-синтетаза амидотрансфераза ИМФ-дегидрогеназа Аденилосукцинатсинтаза
- 18. ЗАПАСНОЙ ПУТЬ СИНТЕЗА (ПУТЬ СПАСЕНИЯ)
- 19. Функция: использовать уже синтезированные азотистые основания и нуклеозиды Основные ферменты: ГГФРТ (гипоксантингуанинфосфорибозилтрансфераза) и АФРТ (аденозинфосфорибозилтрансфераза) Действие
- 20. Рреакция, катализируемая АФРТ (аденинфосфорибозилтрансфераза)
- 21. Первая реакция, катализируемая ГГФРТ (гипоксантин-гуанинфосфорибозилтрансфераза): превращение гипоксантина в ИМФ
- 22. Вторая реакция, катализируемая ГГФРТ (гипоксантин-гуанинфосфорибозилтрансфераза): превращение гуанина в ГМФ
- 23. РАСПАД ПУРИНОВ
- 24. Конечным продуктом распада является мочевая кислота (содержит пуриновое кольцо, т.е. сама является пурином) При распаде АТФ
- 25. Распад пуринов На схеме не показано, как сначала АТФ и ГТФ дефосфорилируются и превращаются в аденозин
- 26. НАРУШЕНИЯ МЕТАБОЛИЗМА ПУРИНОВ
- 27. Гиперурикемия: повышение содержания мочевой кислоты в крови Мочевая кислота плохо растворима в воде. Накапливаясь в синовиальной
- 28. СИНТЕЗ ПИРИМИДИНОВ
- 29. Источники атомов пиримидинового кольца: Аспартат Карбамоилфосфат: Глутамин (амидный азот) СО2
- 30. Сначала синтезируется свободное азотистое основание, затем оно прикрепляется к рибозе первое азотистое основание – оротовая кислота
- 31. Начало синтеза пиримидинов: образование карбамоилфосфата глутамин глутамат карбамоилфосфат 1
- 32. Аспартат + карбамоилфосфат N-карбамоиласпартат дигидрооротат оротат Оротидин-5-монофосфат 2 3 Синтез пиримидинов: продолжение 4 5
- 33. Оротидин-5-монофосфат УМФ (уридинмонофосфат) УТФ (уридинтрифосфат) ЦТФ (цитидинтрифосфат) Синтез пиримидинов: окончание 6 7 8
- 34. Полифункциональные ферменты синтеза пиримидинов Реакции 1, 2 и 3 катализируются одним ферментом, у которого есть 3
- 35. Регуляция Регуляторный фермент: карбамоилфосфатсинтетаза II (часть полифункционального фермента, катализирующего 3 первые реакции синтеза) Ингибируется УМФ и
- 36. Регуляция синтеза пиримидинов
- 37. Нарушения: оротацидурия Причина: мутации в ферменте УМФ-синтаза Следствия: гиперпродукция оротата Мегалобластная анемия Нарушения умственного развития, двигательной
- 38. РАСПАД ПИРИМИДИНОВ
- 39. Конечные продукты распада УМФ и ЦМФ: СО2 Аммиак β-аланин Конечные продукты распада дТМФ: СО2 Аммиак β-аминоизомасляная
- 40. Распад пиримидинов На схеме не показано, как от ЦТФ, УТФ и дТТФ отщепляются фосфаты и рибоза,
- 41. СИНТЕЗ ДЕЗОКСИРИБОНУКЛЕОТИДОВ
- 42. Основные пути синтеза ведут к появлению только РИБОнуклеотидов (для РНК) 1. Путь синтеза пиримидинов de novo
- 43. Фермент рибонуклеотидредуктаза: Синтезирует дезоксирибонуклеотиды (дАДФ, дГДФ, дГДФ, дУДФ) из соответствующих рибонуклеотидов Для этого он катализирует превращение
- 44. Схема реакции, катализируемой рибонуклеотидредуктазой: НДФ (нуклеотиддифосфат) превращается в дНДФ (дезоксинуклеотиддифосфат) НДФ дНДФ Восстановленный тиоредоксин Окисленный тиоредоксин
- 46. Скачать презентацию

Вопросы из билетов по теме
Биосинтез и распад пиримидиновых нуклеотидов.
Источники атомов пуринового
Вопросы из билетов по теме
Биосинтез и распад пиримидиновых нуклеотидов.
Источники атомов пуринового

Структура темы
Классификация нуклеотидов
Пуриновые нуклеотиды
Синтез
Распад
Нарушения распада и синтеза: подагра
Пиримидиновые нуклеотиды
Синтез
Распад
Нарушения синтеза
Синтез дезоксирибонуклеотидов
Структура темы
Классификация нуклеотидов
Пуриновые нуклеотиды
Синтез
Распад
Нарушения распада и синтеза: подагра
Пиримидиновые нуклеотиды
Синтез
Распад
Нарушения синтеза
Синтез дезоксирибонуклеотидов
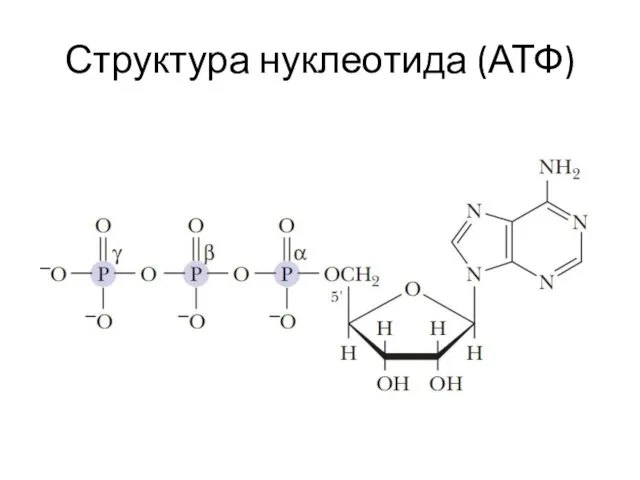
Структура нуклеотида (АТФ)
Структура нуклеотида (АТФ)
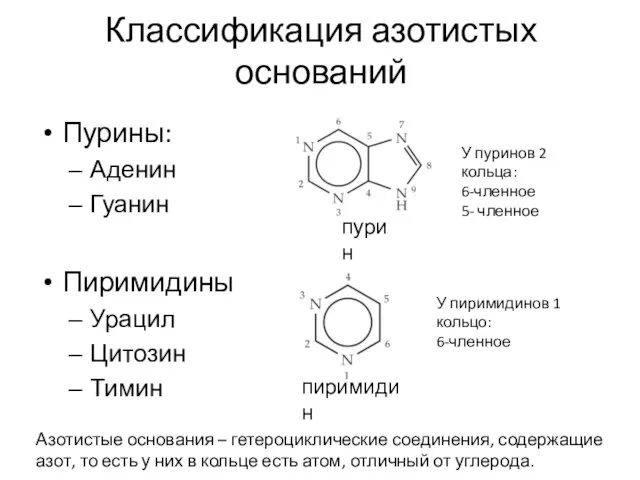
Классификация азотистых оснований
Пурины:
Аденин
Гуанин
Пиримидины
Урацил
Цитозин
Тимин
У пуринов 2 кольца:
6-членное
5- членное
У пиримидинов 1 кольцо:
6-членное
Азотистые основания
Классификация азотистых оснований
Пурины:
Аденин
Гуанин
Пиримидины
Урацил
Цитозин
Тимин
У пуринов 2 кольца:
6-членное
5- членное
У пиримидинов 1 кольцо:
6-членное
Азотистые основания

СИНТЕЗ ПУРИНОВ
СИНТЕЗ ПУРИНОВ

Два пути синтеза нуклеотидов
de novo (с нуля):
из простых предшественников
Запасной путь
Два пути синтеза нуклеотидов
de novo (с нуля):
из простых предшественников
Запасной путь

СИНТЕЗ DE NOVO
СИНТЕЗ DE NOVO
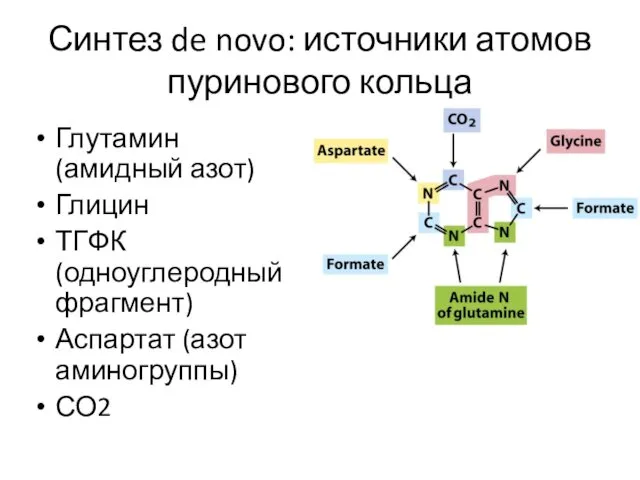
Синтез de novo: источники атомов пуринового кольца
Глутамин (амидный азот)
Глицин
ТГФК (одноуглеродный фрагмент)
Аспартат
Синтез de novo: источники атомов пуринового кольца
Глутамин (амидный азот)
Глицин
ТГФК (одноуглеродный фрагмент)
Аспартат

Особенности синтеза
Пуриновое кольцо строится постепенным добавлением новых атомов в строго определенном
Особенности синтеза
Пуриновое кольцо строится постепенным добавлением новых атомов в строго определенном

Синтез
1-я реакция: образование ФРПФ (фосфорибозилпирофосфат) из рибозо-5-фосфата (см. пентозофосфатный путь) и
Синтез
1-я реакция: образование ФРПФ (фосфорибозилпирофосфат) из рибозо-5-фосфата (см. пентозофосфатный путь) и
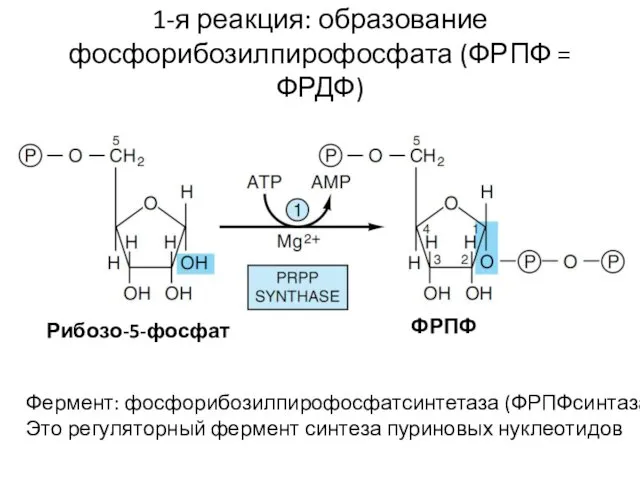
1-я реакция: образование фосфорибозилпирофосфата (ФРПФ = ФРДФ)
Фермент: фосфорибозилпирофосфатсинтетаза (ФРПФсинтаза)
Это регуляторный фермент
1-я реакция: образование фосфорибозилпирофосфата (ФРПФ = ФРДФ)
Фермент: фосфорибозилпирофосфатсинтетаза (ФРПФсинтаза)
Это регуляторный фермент
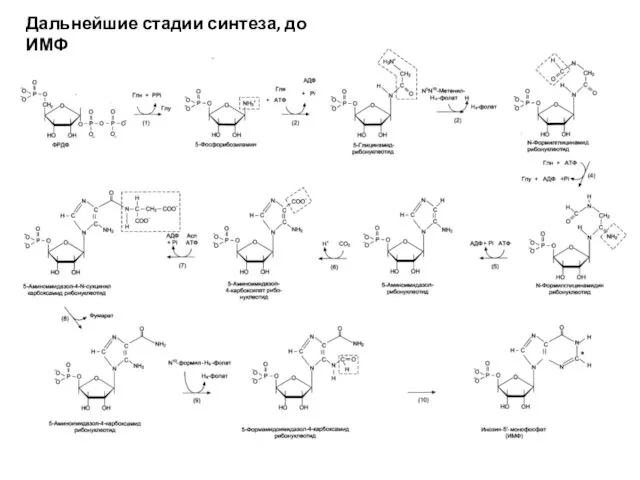
Дальнейшие стадии синтеза, до ИМФ
Дальнейшие стадии синтеза, до ИМФ
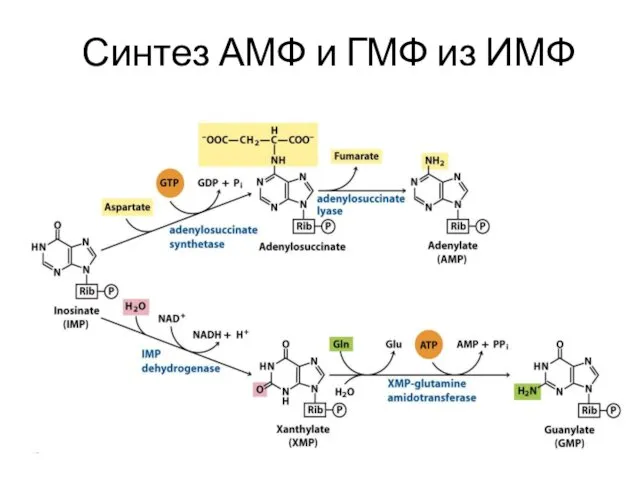
Синтез АМФ и ГМФ из ИМФ
Синтез АМФ и ГМФ из ИМФ

Регуляция синтеза de novo
Основной регуляторный фермент: ФРПФ-синтетаза
Другие регуляторные ферменты: глутамин-фосфорибозиламидотрансфераза, ИМФ-дегидрогеназа,
Регуляция синтеза de novo
Основной регуляторный фермент: ФРПФ-синтетаза
Другие регуляторные ферменты: глутамин-фосфорибозиламидотрансфераза, ИМФ-дегидрогеназа,
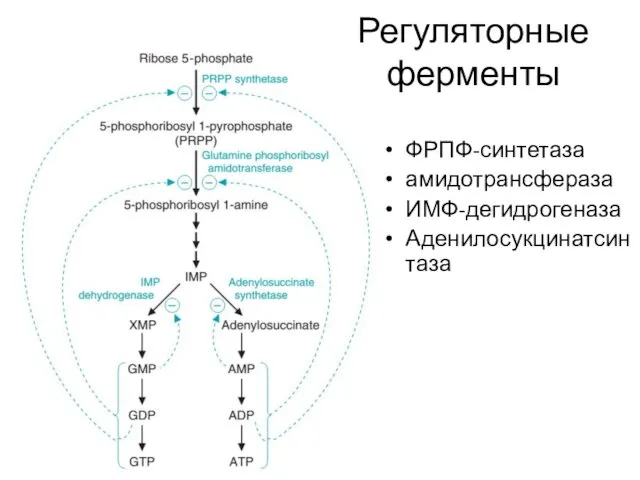
Регуляторные ферменты
ФРПФ-синтетаза
амидотрансфераза
ИМФ-дегидрогеназа
Аденилосукцинатсинтаза
Регуляторные ферменты
ФРПФ-синтетаза
амидотрансфераза
ИМФ-дегидрогеназа
Аденилосукцинатсинтаза

ЗАПАСНОЙ ПУТЬ СИНТЕЗА (ПУТЬ СПАСЕНИЯ)
ЗАПАСНОЙ ПУТЬ СИНТЕЗА (ПУТЬ СПАСЕНИЯ)

Функция: использовать уже синтезированные азотистые основания и нуклеозиды
Основные ферменты: ГГФРТ (гипоксантингуанинфосфорибозилтрансфераза)
Функция: использовать уже синтезированные азотистые основания и нуклеозиды
Основные ферменты: ГГФРТ (гипоксантингуанинфосфорибозилтрансфераза)
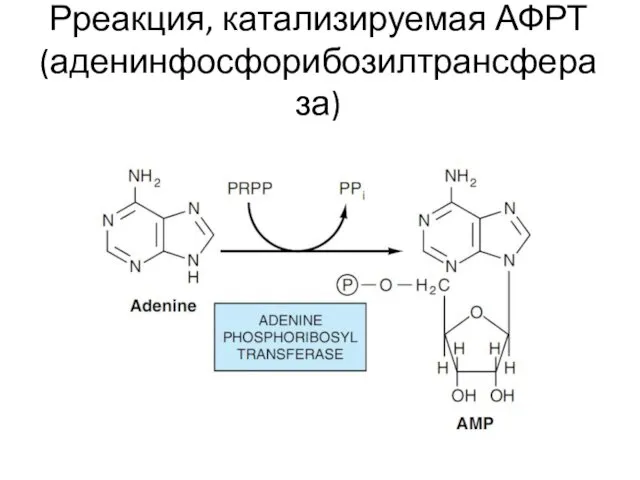
Рреакция, катализируемая АФРТ (аденинфосфорибозилтрансфераза)
Рреакция, катализируемая АФРТ (аденинфосфорибозилтрансфераза)
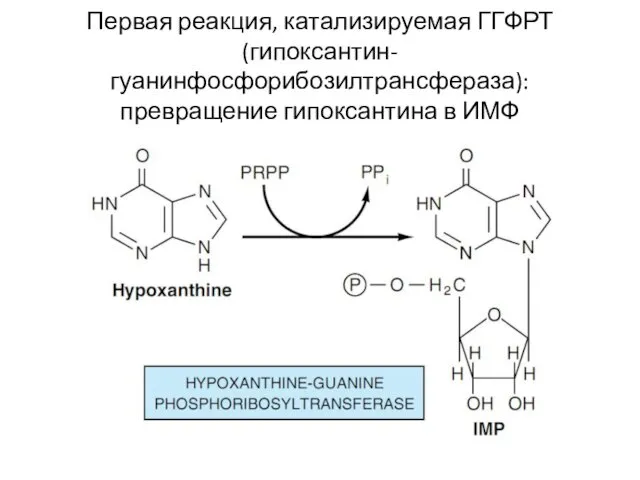
Первая реакция, катализируемая ГГФРТ (гипоксантин-гуанинфосфорибозилтрансфераза): превращение гипоксантина в ИМФ
Первая реакция, катализируемая ГГФРТ (гипоксантин-гуанинфосфорибозилтрансфераза): превращение гипоксантина в ИМФ
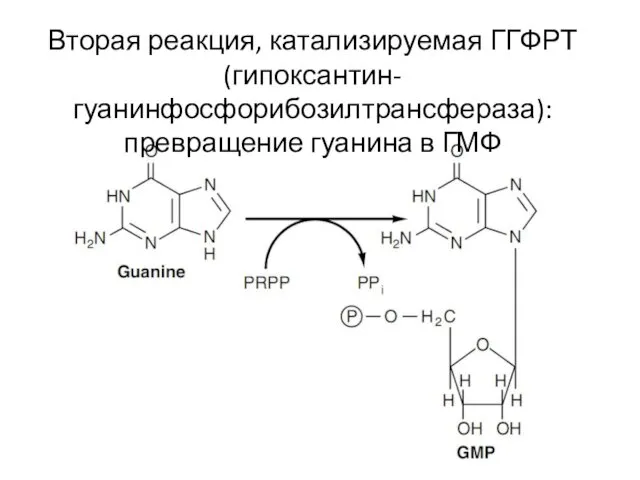
Вторая реакция, катализируемая ГГФРТ (гипоксантин-гуанинфосфорибозилтрансфераза): превращение гуанина в ГМФ
Вторая реакция, катализируемая ГГФРТ (гипоксантин-гуанинфосфорибозилтрансфераза): превращение гуанина в ГМФ

РАСПАД ПУРИНОВ
РАСПАД ПУРИНОВ

Конечным продуктом распада является мочевая кислота (содержит пуриновое кольцо, т.е. сама
Конечным продуктом распада является мочевая кислота (содержит пуриновое кольцо, т.е. сама
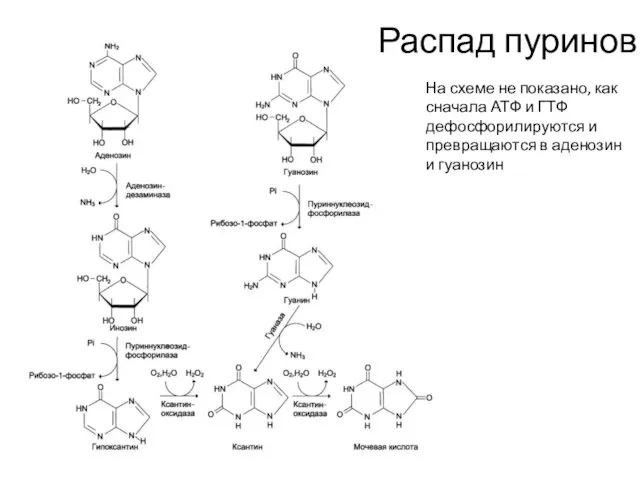
Распад пуринов
На схеме не показано, как сначала АТФ и ГТФ дефосфорилируются
Распад пуринов
На схеме не показано, как сначала АТФ и ГТФ дефосфорилируются

НАРУШЕНИЯ МЕТАБОЛИЗМА ПУРИНОВ
НАРУШЕНИЯ МЕТАБОЛИЗМА ПУРИНОВ

Гиперурикемия: повышение содержания мочевой кислоты в крови
Мочевая кислота плохо растворима в
Гиперурикемия: повышение содержания мочевой кислоты в крови
Мочевая кислота плохо растворима в

СИНТЕЗ ПИРИМИДИНОВ
СИНТЕЗ ПИРИМИДИНОВ
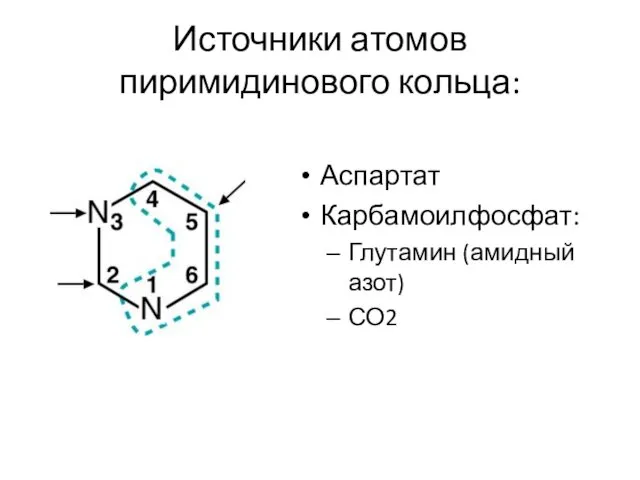
Источники атомов пиримидинового кольца:
Аспартат
Карбамоилфосфат:
Глутамин (амидный азот)
СО2
Источники атомов пиримидинового кольца:
Аспартат
Карбамоилфосфат:
Глутамин (амидный азот)
СО2

Сначала синтезируется свободное азотистое основание, затем оно прикрепляется к рибозе
первое азотистое
Сначала синтезируется свободное азотистое основание, затем оно прикрепляется к рибозе
первое азотистое
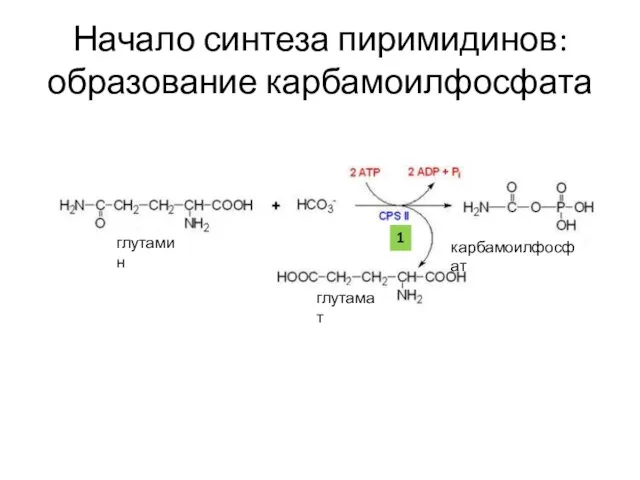
Начало синтеза пиримидинов: образование карбамоилфосфата
глутамин
глутамат
карбамоилфосфат
1
Начало синтеза пиримидинов: образование карбамоилфосфата
глутамин
глутамат
карбамоилфосфат
1
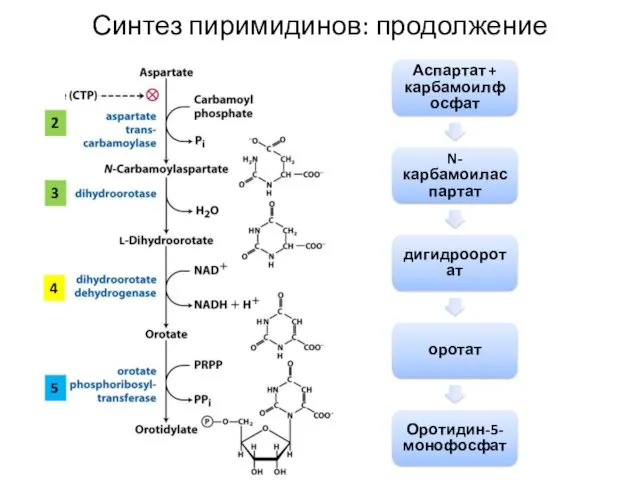
Аспартат + карбамоилфосфат
N-карбамоиласпартат
дигидрооротат
оротат
Оротидин-5-монофосфат
2
3
Синтез пиримидинов: продолжение
4
5
Аспартат + карбамоилфосфат
N-карбамоиласпартат
дигидрооротат
оротат
Оротидин-5-монофосфат
2
3
Синтез пиримидинов: продолжение
4
5
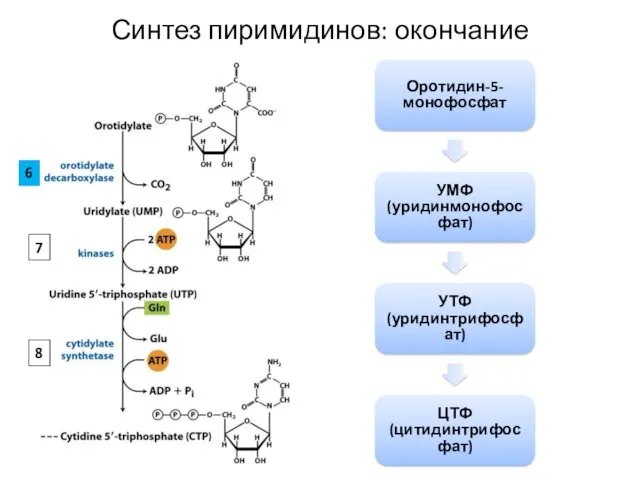
Оротидин-5-монофосфат
УМФ (уридинмонофосфат)
УТФ (уридинтрифосфат)
ЦТФ (цитидинтрифосфат)
Синтез пиримидинов: окончание
6
7
8
Оротидин-5-монофосфат
УМФ (уридинмонофосфат)
УТФ (уридинтрифосфат)
ЦТФ (цитидинтрифосфат)
Синтез пиримидинов: окончание
6
7
8

Полифункциональные ферменты синтеза пиримидинов
Реакции 1, 2 и 3 катализируются одним ферментом,
Полифункциональные ферменты синтеза пиримидинов
Реакции 1, 2 и 3 катализируются одним ферментом,

Регуляция
Регуляторный фермент: карбамоилфосфатсинтетаза II (часть полифункционального фермента, катализирующего 3 первые реакции
Регуляция
Регуляторный фермент: карбамоилфосфатсинтетаза II (часть полифункционального фермента, катализирующего 3 первые реакции
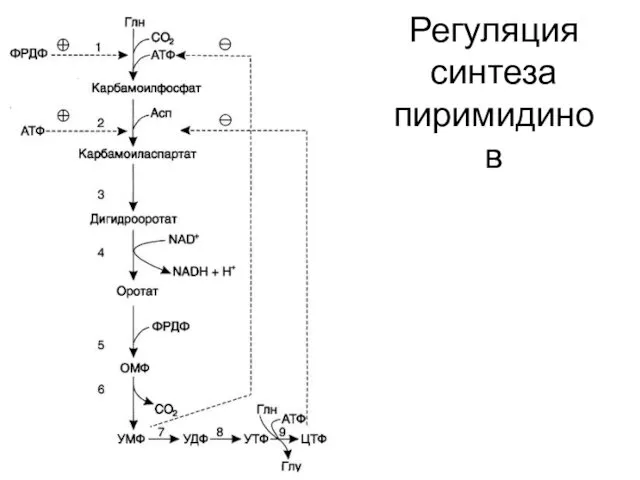
Регуляция синтеза пиримидинов
Регуляция синтеза пиримидинов

Нарушения: оротацидурия
Причина: мутации в ферменте УМФ-синтаза
Следствия:
гиперпродукция оротата
Мегалобластная анемия
Нарушения умственного развития, двигательной
Нарушения: оротацидурия
Причина: мутации в ферменте УМФ-синтаза
Следствия:
гиперпродукция оротата
Мегалобластная анемия
Нарушения умственного развития, двигательной

РАСПАД ПИРИМИДИНОВ
РАСПАД ПИРИМИДИНОВ

Конечные продукты распада УМФ и ЦМФ:
СО2
Аммиак
β-аланин
Конечные продукты распада дТМФ:
СО2
Аммиак
β-аминоизомасляная кислота (β-аминоизобутират)
β-аланин
Конечные продукты распада УМФ и ЦМФ:
СО2
Аммиак
β-аланин
Конечные продукты распада дТМФ:
СО2
Аммиак
β-аминоизомасляная кислота (β-аминоизобутират)
β-аланин
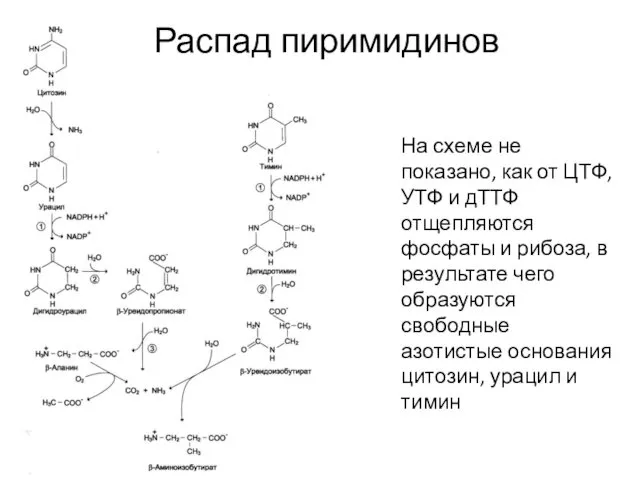
Распад пиримидинов
На схеме не показано, как от ЦТФ, УТФ и дТТФ
Распад пиримидинов
На схеме не показано, как от ЦТФ, УТФ и дТТФ

СИНТЕЗ ДЕЗОКСИРИБОНУКЛЕОТИДОВ
СИНТЕЗ ДЕЗОКСИРИБОНУКЛЕОТИДОВ

Основные пути синтеза ведут к появлению только РИБОнуклеотидов (для РНК)
1. Путь
Основные пути синтеза ведут к появлению только РИБОнуклеотидов (для РНК)
1. Путь

Фермент рибонуклеотидредуктаза:
Синтезирует дезоксирибонуклеотиды (дАДФ, дГДФ, дГДФ, дУДФ) из соответствующих рибонуклеотидов
Для этого
Фермент рибонуклеотидредуктаза:
Синтезирует дезоксирибонуклеотиды (дАДФ, дГДФ, дГДФ, дУДФ) из соответствующих рибонуклеотидов
Для этого
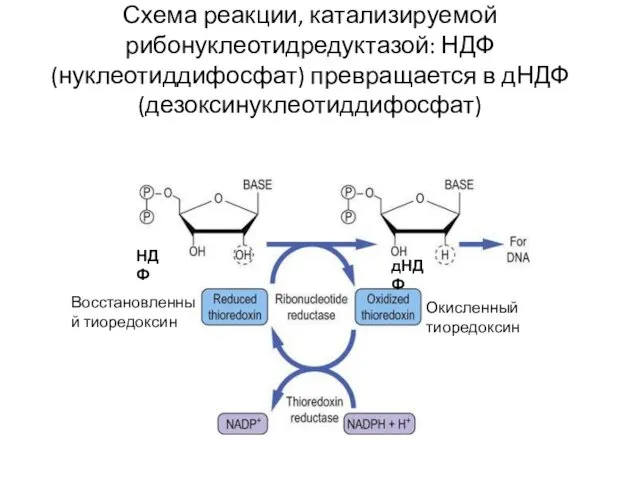
Схема реакции, катализируемой рибонуклеотидредуктазой: НДФ (нуклеотиддифосфат) превращается в дНДФ (дезоксинуклеотиддифосфат)
НДФ
дНДФ
Восстановленный тиоредоксин
Окисленный
Схема реакции, катализируемой рибонуклеотидредуктазой: НДФ (нуклеотиддифосфат) превращается в дНДФ (дезоксинуклеотиддифосфат)
НДФ
дНДФ
Восстановленный тиоредоксин
Окисленный
 С днем мамы
С днем мамы Семья пастора
Семья пастора Комплексные числа
Комплексные числа ФГОС в дошкольном образовании
ФГОС в дошкольном образовании Пластиковые окна или старые деревянные
Пластиковые окна или старые деревянные Битва под Сталинградом
Битва под Сталинградом Устный журнал Путешествие по планете Памяти
Устный журнал Путешествие по планете Памяти Бойове застосування КЗА 86Ж6. Алгоритми обробки радiолокацiйної iнформацiї, виявлення та захоплення цiлей. (Тема 8.3)
Бойове застосування КЗА 86Ж6. Алгоритми обробки радiолокацiйної iнформацiї, виявлення та захоплення цiлей. (Тема 8.3) Саванны. Субэкваториальный климатический пояс
Саванны. Субэкваториальный климатический пояс Воспитательная система класса
Воспитательная система класса Презентация Семья и школа в формировании личности ребенка
Презентация Семья и школа в формировании личности ребенка Clothes and Fashion
Clothes and Fashion Ограничение облучения от природных источников
Ограничение облучения от природных источников Презентация к викторине У сказки тихий голосок.
Презентация к викторине У сказки тихий голосок. Сюжетно-ролевая игра
Сюжетно-ролевая игра Мастер моего села
Мастер моего села Блиц-опрос Я знаю лес
Блиц-опрос Я знаю лес Концепция педагогической деятельности
Концепция педагогической деятельности System administracji publicznej w Hondurasie
System administracji publicznej w Hondurasie GP USB DownLoad Manual (English_Version)
GP USB DownLoad Manual (English_Version) Параллельность прямых в пространстве
Параллельность прямых в пространстве Хранение дел организации. (Тема 4)
Хранение дел организации. (Тема 4) Формирование комфортной городской среды
Формирование комфортной городской среды PML30_SummerPractice2022_Presentation_Pattern
PML30_SummerPractice2022_Presentation_Pattern Декоративный пейзаж
Декоративный пейзаж Пищевая сода. Необычное в обычном
Пищевая сода. Необычное в обычном Как научить своего ребёнка жить в мире людей. Уроки этики поведения для детей и родителей
Как научить своего ребёнка жить в мире людей. Уроки этики поведения для детей и родителей Выдающиеся открытия в области химии
Выдающиеся открытия в области химии