Очистка глутаматдегидрогеназы из листьев кукурузы и исследование ее каталитических свойств презентация
- Очистка глутаматдегидрогеназы из листьев кукурузы и исследование ее каталитических свойств

Содержание
- 2. ОБЗОР ЛИТЕРАТУРЫ
- 3. Глутаматдегидрогеназа, как правило, катализирует обратимую реакцию восстановительного аминирования 2-оксоглутарата (2-ОГ) до глутамата с использованием НАД(Ф) в
- 4. Коферментная специфичность Согласно коферментной специфичности, существуют три основных типа ГДГ: НАД-зависимая (КФ 1.4.1.2) НАДФ-зависимая (КФ 1.4.1.4)
- 5. Локализация в растениях Изоферменты ГДГ в растениях могут встречаться в корнях (GDH-2), в семядолях (GDH-1), а
- 6. Некоторые физико-химические характеристики Ph-оптимум фермента различается у различных организмов, а также в зависимости от катализируемой реакции.
- 7. Структура фермента Большинство глутаматдегидрогеназ, которые были обнаружены до сих пор – олигомерные ферменты, но они различаются
- 8. Роль в обмене азота Неорганический азот усваивается в виде аммония, который включен в качестве аминогруппы в
- 9. Схема ассимиляции молекулярного азота
- 10. ГАМК-шунт
- 11. Гены ГДГ Известно, что активный белок глутаматдегидрогеназы чаще всего представляет собой гексамер, состоящий из шести субъединиц
- 12. В геноме арабидопсиаса также обнаружено 3 гена: gdh1 и gdh2, локализованные в 5 хромосоме и gdh3,
- 13. ЭКСПЕРИМЕНТАЛЬНАЯ ЧАСТЬ
- 14. Цель и задачи Целью курсовой работы было получение высокоочищенных препаратов глутаматдегидрогеназы из листьев кукурузы и изучение
- 15. Методы исследования
- 16. Определение активности ГДГ Активность глутаматдегидрогеназы определяли спектрофотометрическим методом путем измерения оптической плотности раствора, содержащего 2,5мМ 2-оксоглутарата,
- 17. Определение количества белка по методу Лоури Общее количество белка определяли по методу Lowry с совт. Оптическую
- 18. Очистка глутаматдегидрогеназы из зеленых листьев кукурузы Очистку фермента осуществляли в несколько стадий при температуре 0-4°С. 1.
- 19. Полученный ферментативный препарат наносили на колонку, заполненную сефадексом G-25 для освобождения от низкомолекулярных примесей. Элюцию осуществляли
- 20. РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
- 21. Очистка ГДГ из зеленых листьев кукурузы Первой стадией очистки было получение гомогената. Общая активность в гомогенате
- 22. Таблица 1. Очистка ГДГ
- 23. В качестве определяющей стадии очистки осуществляли ионообменную хроматографию. Элюцию фермента в колонки осуществляли линейным градиентом хлорида
- 24. Определение кинетических характеристик
- 25. Определение оптимума pH Зависимость активности полученного препарата ГДГ отзначения pH для реакции окисления 2- оксоглутарата.
- 26. Определение константы Михаэлиса Определение Km полученного ферментного препарата по 2-оксоглутарату Km =28,57 мМ
- 27. Определение константы Михаэлиса Определение Km полученного ферментного препарата по НАДН Km = 22,075 мМ
- 28. ВЫВОДЫ 1. С помощью четырехстадийной схемы очистки получен высокоочищенный препарат глутаматдегидрогеназы из листьев кукурузы с удельной
- 30. Скачать презентацию

ОБЗОР ЛИТЕРАТУРЫ
ОБЗОР ЛИТЕРАТУРЫ

Глутаматдегидрогеназа, как правило, катализирует обратимую реакцию восстановительного аминирования 2-оксоглутарата (2-ОГ) до
Глутаматдегидрогеназа, как правило, катализирует обратимую реакцию восстановительного аминирования 2-оксоглутарата (2-ОГ) до

Коферментная специфичность
Согласно коферментной специфичности, существуют три основных типа ГДГ:
НАД-зависимая (КФ
Коферментная специфичность
Согласно коферментной специфичности, существуют три основных типа ГДГ:
НАД-зависимая (КФ

Локализация в растениях
Изоферменты ГДГ в растениях могут встречаться в корнях (GDH-2),
Локализация в растениях
Изоферменты ГДГ в растениях могут встречаться в корнях (GDH-2),

Некоторые физико-химические характеристики
Ph-оптимум фермента различается у различных организмов, а также в
Некоторые физико-химические характеристики
Ph-оптимум фермента различается у различных организмов, а также в

Структура фермента
Большинство глутаматдегидрогеназ, которые были обнаружены до сих пор – олигомерные
Структура фермента
Большинство глутаматдегидрогеназ, которые были обнаружены до сих пор – олигомерные

Роль в обмене азота
Неорганический азот усваивается в виде аммония, который включен
Роль в обмене азота
Неорганический азот усваивается в виде аммония, который включен

Схема ассимиляции молекулярного азота
Схема ассимиляции молекулярного азота

ГАМК-шунт
ГАМК-шунт

Гены ГДГ
Известно, что активный белок глутаматдегидрогеназы чаще всего представляет собой гексамер,
Гены ГДГ
Известно, что активный белок глутаматдегидрогеназы чаще всего представляет собой гексамер,

В геноме арабидопсиаса также обнаружено 3 гена: gdh1 и gdh2, локализованные
В геноме арабидопсиаса также обнаружено 3 гена: gdh1 и gdh2, локализованные

ЭКСПЕРИМЕНТАЛЬНАЯ ЧАСТЬ
ЭКСПЕРИМЕНТАЛЬНАЯ ЧАСТЬ

Цель и задачи
Целью курсовой работы было получение высокоочищенных препаратов глутаматдегидрогеназы из
Цель и задачи
Целью курсовой работы было получение высокоочищенных препаратов глутаматдегидрогеназы из

Методы исследования
Методы исследования

Определение активности ГДГ
Активность глутаматдегидрогеназы определяли спектрофотометрическим методом путем измерения оптической плотности
Определение активности ГДГ
Активность глутаматдегидрогеназы определяли спектрофотометрическим методом путем измерения оптической плотности

Определение количества белка по методу Лоури
Общее количество белка определяли по методу
Определение количества белка по методу Лоури
Общее количество белка определяли по методу

Очистка глутаматдегидрогеназы из зеленых листьев кукурузы
Очистку фермента осуществляли в несколько стадий
Очистка глутаматдегидрогеназы из зеленых листьев кукурузы
Очистку фермента осуществляли в несколько стадий

Полученный ферментативный препарат наносили на колонку, заполненную сефадексом G-25 для освобождения
Полученный ферментативный препарат наносили на колонку, заполненную сефадексом G-25 для освобождения

РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ

Очистка ГДГ из зеленых листьев кукурузы
Первой стадией очистки было получение гомогената.
Очистка ГДГ из зеленых листьев кукурузы
Первой стадией очистки было получение гомогената.

Таблица 1. Очистка ГДГ
Таблица 1. Очистка ГДГ

В качестве определяющей стадии очистки осуществляли ионообменную хроматографию. Элюцию фермента в
В качестве определяющей стадии очистки осуществляли ионообменную хроматографию. Элюцию фермента в

Определение кинетических характеристик
Определение кинетических характеристик
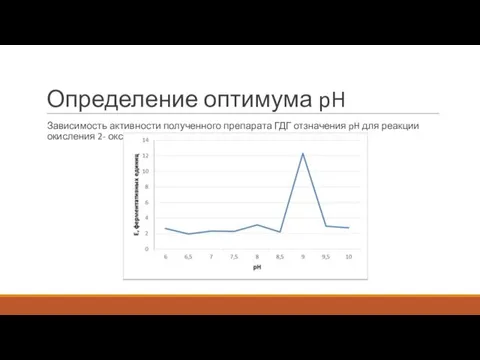
Определение оптимума pH
Зависимость активности полученного препарата ГДГ отзначения pH для реакции
Определение оптимума pH
Зависимость активности полученного препарата ГДГ отзначения pH для реакции

Определение константы Михаэлиса
Определение Km полученного ферментного препарата по 2-оксоглутарату
Km =28,57 мМ
Определение константы Михаэлиса
Определение Km полученного ферментного препарата по 2-оксоглутарату
Km =28,57 мМ
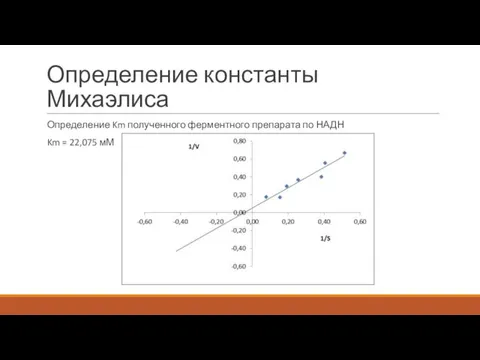
Определение константы Михаэлиса
Определение Km полученного ферментного препарата по НАДН
Km = 22,075
Определение константы Михаэлиса
Определение Km полученного ферментного препарата по НАДН
Km = 22,075

ВЫВОДЫ
1. С помощью четырехстадийной схемы очистки получен высокоочищенный препарат глутаматдегидрогеназы из
ВЫВОДЫ
1. С помощью четырехстадийной схемы очистки получен высокоочищенный препарат глутаматдегидрогеназы из
 Семинар Добро пожаловать в проектные технологии!
Семинар Добро пожаловать в проектные технологии! Из опыта использования интерактивных методов обучения
Из опыта использования интерактивных методов обучения Создатели и потребители моды
Создатели и потребители моды Урок города Что такое Кузбасс Диск
Урок города Что такое Кузбасс Диск Заболачивание
Заболачивание Умный дом. Передача данных с датчиков умного дома
Умный дом. Передача данных с датчиков умного дома Устройство компьютера
Устройство компьютера Учимся читать с паровозиком - обучение плавному слоговому чтению.
Учимся читать с паровозиком - обучение плавному слоговому чтению. Окружность!
Окружность! Род местоимений 3 лица
Род местоимений 3 лица Презентация Алюминий
Презентация Алюминий Информация для родителей
Информация для родителей Закон об ОСАГО 2012 года. Связь технического осмотра и страхования. Получение полиса ОСАГО
Закон об ОСАГО 2012 года. Связь технического осмотра и страхования. Получение полиса ОСАГО Гардероб для тварини. Проект
Гардероб для тварини. Проект Презентация из опыта работы
Презентация из опыта работы Проблемы науки и производства. Биосферная совместимость. Архитектура
Проблемы науки и производства. Биосферная совместимость. Архитектура Защитник прав человеческих А.Д. Сахаров
Защитник прав человеческих А.Д. Сахаров Презентация Рассказы и сказки Н. Носова
Презентация Рассказы и сказки Н. Носова Интеллектуальный марафон. Занятие 3 (3 класс)
Интеллектуальный марафон. Занятие 3 (3 класс) Vitaminele
Vitaminele Агротехника виноградного куста. Посадка
Агротехника виноградного куста. Посадка Презентация Организация предметно-развивающей среды часть II
Презентация Организация предметно-развивающей среды часть II Участники процесса, отстаивающие свои или представляемые интересы
Участники процесса, отстаивающие свои или представляемые интересы Климат нашей местности. 6 класс
Климат нашей местности. 6 класс Технология кладочных работ. Основные типы конструкций наружных стен
Технология кладочных работ. Основные типы конструкций наружных стен Изменение форм взаимодействия воспитателя с родителями в соответствии с требованиями ФГОС ДО
Изменение форм взаимодействия воспитателя с родителями в соответствии с требованиями ФГОС ДО Презентация урока черчения: Чертежи шпоночных и штифтовых соединений
Презентация урока черчения: Чертежи шпоночных и штифтовых соединений Многообразие ракообразных в природе
Многообразие ракообразных в природе