- Симбиогенетика микоризы. (Лекция 6)

Содержание
- 2. Фундаментальное значение микоризных ассоциаций для наземных растений Микориза —тип микробно-растительного симбиоза, который формируется между растениями и
- 3. Типы микориз Эктомикоризы - распространение гриба ограничено межклеточными пространствами Эндомикоризы - гриб проникает в растительные клетки,
- 4. Эктомикориза, распространение и значение Эктомикориза – наиболее часто встречающийся в лесах средне-европейской полосы корневой симбиоз. Подобная
- 5. Строение эктомикоризы Гифы гриба прорастают в корковый слой корня, но, не проникая в корневые клетки, образуют
- 6. Онтогенез эктомикориз (ЭМ) 3 стадии: Преинфекционные процессы: индукция корневыми экссудатами прорастания покоящихся спор и рост гифов,
- 7. Гипотеза о ведущей роли фитогормонов, синтезируемых грибами (ауксин-подобные соединения) В ходе образования ЭМ партнеры синтезируют ряд
- 8. Метаболическая интеграция партнеров эктомикоризы (ЭM) Основная функция ЭМ заключается в обеспечении хозяина азотом (NH4+). В деградации
- 9. Мобилизация почвенного азота — это многоступенчатый процесс, включающий разрушение полимера, поглощение грибом образующихся низкомолекулярных продуктов, перевод
- 10. Везикулярно-арбускулярная микориза (ВАМ): распространение и значение Грибы-зигомицеты из порядка Glomales (200 видов) Симбиоз между корнями растения
- 11. Стадии развития арбускулярной микоризы 3 этапа: преинфекционные взаимодействия: споры гриба прорастают в почве под действием растительных
- 12. 1 стадия - преинфекционные взаимодействия Хемотаксис Растительными сигналами, активирующими прорастание спор, рост зародышевых трубок по направлению
- 13. 2 стадия - инфекционная Апрессории – специальные структуры прикрепления. Изменения в растительных клетках: Вакуоль резко уменьшается
- 14. Две морфологические формы эндомикоризной ассоциации “Arum” и “Paris” Арбускулярная микориза у клевера (арбускулы в клетке кортекса
- 15. 3 стадия — формирование внекорневых гифов Для растения-хозяина наиболее важна их трофическая функция (поглощение почвенных веществ),
- 17. Симбиогенетика АМ Анализ контроля над развитием AM, осуществляемого со стороны хозяина, был начат в конце 1980-х
- 18. Сходство генетического контроля формирования АМ и клубенькообразования Выявляется 300-400 генов, активируемых при развитии AM, причем около
- 19. Гены симбиоза у растений Один и тот же ген SYMRK необходим для успешного сожительства растений с
- 20. Эволюция генов симбиоза у растений «Генетическая программа» клубенькового симбиоза представляет собой модификацию «генетической программы» более древней
- 21. Метаболическая интеграция партнеров арбускулярной микоризы (AM) Инокуляция АМ-грибами улучшает поступление в растения всего комплекса питательных веществ,
- 22. Орхидная микориза Практически все орхидные (сем. Orchidaceae) являются облигатными микотрофами, не способными осуществлять прорастание семян и
- 24. Скачать презентацию

Фундаментальное значение микоризных ассоциаций для наземных растений
Микориза —тип микробно-растительного симбиоза,
Фундаментальное значение микоризных ассоциаций для наземных растений
Микориза —тип микробно-растительного симбиоза,
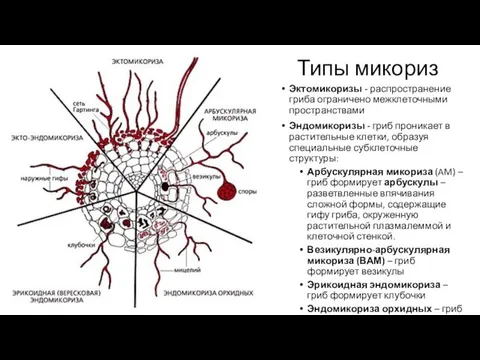
Типы микориз
Эктомикоризы - распространение гриба ограничено межклеточными пространствами
Эндомикоризы - гриб проникает
Типы микориз
Эктомикоризы - распространение гриба ограничено межклеточными пространствами
Эндомикоризы - гриб проникает

Эктомикориза, распространение и значение
Эктомикориза – наиболее часто встречающийся в лесах средне-европейской
Эктомикориза, распространение и значение
Эктомикориза – наиболее часто встречающийся в лесах средне-европейской
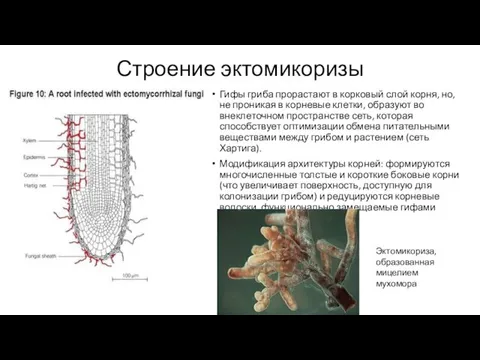
Строение эктомикоризы
Гифы гриба прорастают в корковый слой корня, но, не проникая
Строение эктомикоризы
Гифы гриба прорастают в корковый слой корня, но, не проникая

Онтогенез эктомикориз (ЭМ)
3 стадии:
Преинфекционные процессы: индукция корневыми экссудатами прорастания покоящихся спор
Онтогенез эктомикориз (ЭМ)
3 стадии:
Преинфекционные процессы: индукция корневыми экссудатами прорастания покоящихся спор

Гипотеза о ведущей роли фитогормонов, синтезируемых грибами (ауксин-подобные соединения)
В ходе образования
Гипотеза о ведущей роли фитогормонов, синтезируемых грибами (ауксин-подобные соединения)
В ходе образования

Метаболическая интеграция партнеров эктомикоризы (ЭM)
Основная функция ЭМ заключается в обеспечении хозяина
Метаболическая интеграция партнеров эктомикоризы (ЭM)
Основная функция ЭМ заключается в обеспечении хозяина

Мобилизация почвенного азота — это многоступенчатый процесс, включающий разрушение полимера, поглощение
Мобилизация почвенного азота — это многоступенчатый процесс, включающий разрушение полимера, поглощение

Везикулярно-арбускулярная микориза (ВАМ): распространение и значение
Грибы-зигомицеты из порядка Glomales (200 видов)
Симбиоз
Везикулярно-арбускулярная микориза (ВАМ): распространение и значение
Грибы-зигомицеты из порядка Glomales (200 видов)
Симбиоз
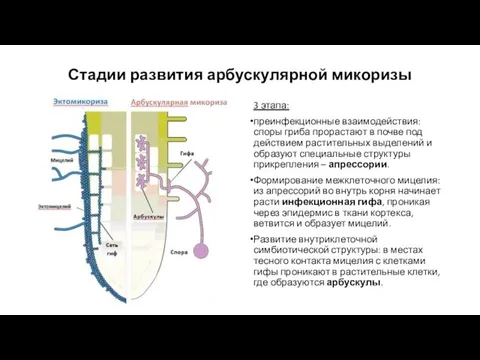
Стадии развития арбускулярной микоризы
3 этапа:
преинфекционные взаимодействия: споры гриба прорастают в
Стадии развития арбускулярной микоризы
3 этапа:
преинфекционные взаимодействия: споры гриба прорастают в

1 стадия - преинфекционные взаимодействия
Хемотаксис
Растительными сигналами, активирующими прорастание спор, рост зародышевых
1 стадия - преинфекционные взаимодействия
Хемотаксис
Растительными сигналами, активирующими прорастание спор, рост зародышевых

2 стадия - инфекционная
Апрессории – специальные структуры прикрепления.
Изменения в растительных
2 стадия - инфекционная
Апрессории – специальные структуры прикрепления.
Изменения в растительных
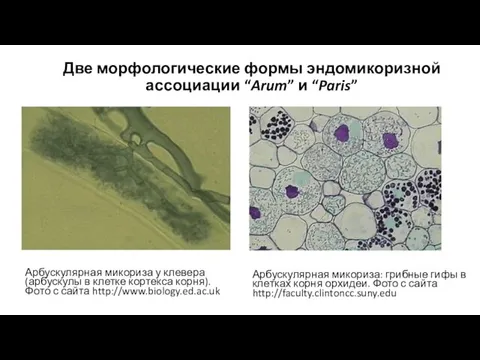
Две морфологические формы эндомикоризной ассоциации “Arum” и “Paris”
Арбускулярная микориза у клевера
Две морфологические формы эндомикоризной ассоциации “Arum” и “Paris”
Арбускулярная микориза у клевера

3 стадия — формирование внекорневых гифов
Для растения-хозяина наиболее важна их трофическая
3 стадия — формирование внекорневых гифов
Для растения-хозяина наиболее важна их трофическая
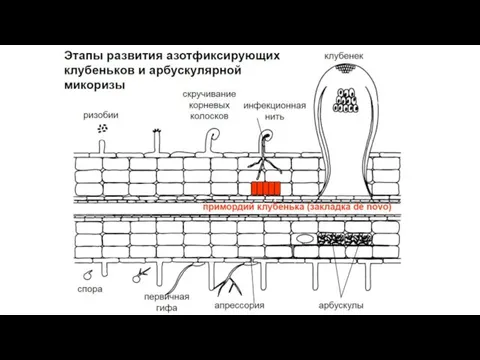

Симбиогенетика АМ
Анализ контроля над развитием AM, осуществляемого со стороны хозяина, был
Симбиогенетика АМ
Анализ контроля над развитием AM, осуществляемого со стороны хозяина, был

Сходство генетического контроля формирования АМ и клубенькообразования
Выявляется 300-400 генов, активируемых при
Сходство генетического контроля формирования АМ и клубенькообразования
Выявляется 300-400 генов, активируемых при

Гены симбиоза у растений
Один и тот же ген SYMRK необходим для
Гены симбиоза у растений
Один и тот же ген SYMRK необходим для
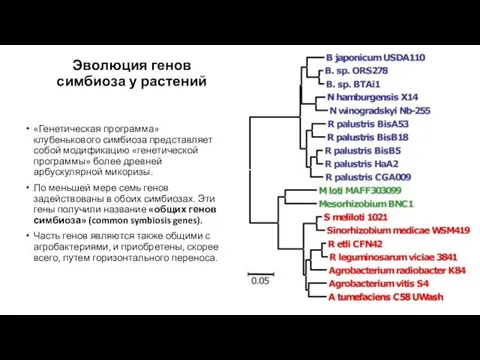
Эволюция генов симбиоза у растений
«Генетическая программа» клубенькового симбиоза представляет собой модификацию
Эволюция генов симбиоза у растений
«Генетическая программа» клубенькового симбиоза представляет собой модификацию
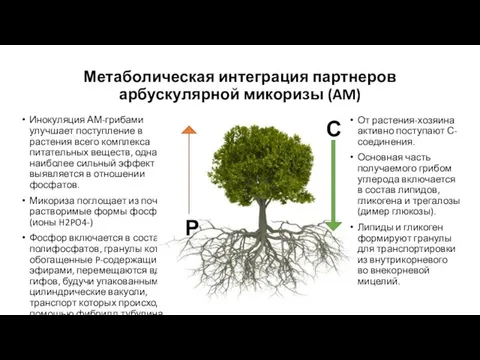
Метаболическая интеграция партнеров арбускулярной микоризы (AM)
Инокуляция АМ-грибами улучшает поступление в растения
Метаболическая интеграция партнеров арбускулярной микоризы (AM)
Инокуляция АМ-грибами улучшает поступление в растения
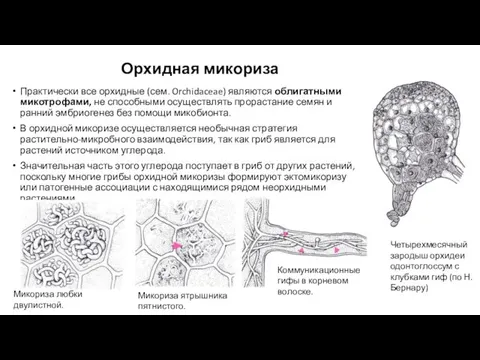
Орхидная микориза
Практически все орхидные (сем. Orchidaceae) являются облигатными микотрофами, не способными
Орхидная микориза
Практически все орхидные (сем. Orchidaceae) являются облигатными микотрофами, не способными
 Асинхронные двигатели
Асинхронные двигатели Технологии и оборудование ТКРС. ОТКРС, УСТиС
Технологии и оборудование ТКРС. ОТКРС, УСТиС Урок технологии во 2 классе Изготовление снежинки из полосок бумаги
Урок технологии во 2 классе Изготовление снежинки из полосок бумаги Правила поведения в школе
Правила поведения в школе Физиология иммунной системы
Физиология иммунной системы Интерактивные технологии в образовательном процессе
Интерактивные технологии в образовательном процессе Лес – легкие планеты
Лес – легкие планеты Arduino Uno. Цифровые контакты ввода-вывода. Широтно-импульсная модуляция
Arduino Uno. Цифровые контакты ввода-вывода. Широтно-импульсная модуляция Шаблон презентации
Шаблон презентации Классный час Их именами названы улицы города
Классный час Их именами названы улицы города Сценарий праздника 1 сентября 2 класс Таинственный остров
Сценарий праздника 1 сентября 2 класс Таинственный остров Сложение натуральных чисел
Сложение натуральных чисел Белки. Основные функции белков
Белки. Основные функции белков Руководство, органы управления, силы и средства ВСМК и ФМГСО
Руководство, органы управления, силы и средства ВСМК и ФМГСО Фундаменты под колонны
Фундаменты под колонны Презентация по теме : Решение задач с пропорциональными величинами
Презентация по теме : Решение задач с пропорциональными величинами Умножение и деление многозначных чисел
Умножение и деление многозначных чисел Этикет, контрольное занятие
Этикет, контрольное занятие РЯ 6 класс №8,9
РЯ 6 класс №8,9 Разработка урока математики в 5 классе Десятичная запись дробных чисел
Разработка урока математики в 5 классе Десятичная запись дробных чисел Воинская обязанность и формы ее реализации
Воинская обязанность и формы ее реализации Путешествие в сказку
Путешествие в сказку Презентация к часу общения.Доброта и милосердие творят чудеса.
Презентация к часу общения.Доброта и милосердие творят чудеса. foodchain
foodchain Строительство здоровой семьи
Строительство здоровой семьи Трубопроводные системы
Трубопроводные системы My native land The Republic of Tatarstan
My native land The Republic of Tatarstan Шаблоны презентаций В гостях у сказки
Шаблоны презентаций В гостях у сказки