- Антропогенез. Место человека в системе животного мира

Содержание
- 2. Место человека в системе животного мира
- 3. Происхождение приматов Конец мезозоя, лесные биотопы. Предки – Насекомоядные. Ближайшая родня – Рукокрылые. Примитивные черты строения
- 4. Вероятно, все известные сегодня африканские плиоценовые гоминиды были всеядными.
- 5. Место гоминид в отряде приматов
- 6. из книги «Млекопитающие» под ред. Д.Макдональда, 2007
- 7. Центр происхождения человека - Восточная Африка. Ближайший современный родственник человека - шимпанзе. Дивергенция произошла 6 -
- 8. К вопросу о двуногости и прямохождении
- 9. Для определения прямохождения у ископаемых приматов используют признаки: – положение затылочного отверстия - у прямоходящих оно
- 10. – строение таза - у прямоходящих таз широкий и низкий (такое строение известно начиная с Australopithecus
- 11. – строение стопы - у прямоходящих выражен свод стопы, пальцы прямые, короткие, большой палец не отведен
- 12. Таким образом, прямохождение возникло более 6 миллионов лет назад, но еще долго отличалось от современного варианта.
- 13. Горилла ходит, опираясь на согнутую кисть
- 14. Бонобо (карликовый шимпанзе) ходит на полусогнутых задних ногах
- 16. Орангутан ранее других человекообразных отделился от общего ствола, сохранив исходный тип двуногого хождения, возможно, характерный для
- 17. Версии происхождения бипедии На открытых равнинах саванн, среди высокой травы выпрямление тела и высоко поднятая голова
- 18. Sahelanthropus tchadensis Сахелантроп «Тумай» возраст 6–7 млн. лет (миоцен); найден в 2001 г. в в пустыне
- 19. сахелантроп
- 20. Orrorin tugenensis Оррорин возраст 6 млн лет (миоцен); найден в 2000 г. французской экспедицией в Кении,
- 21. оррорин
- 22. Australopithecus anamensis Австралопитек озерный возраст 4,1-4,2 млн лет (плиоцен); найден группой Мив Лики в Кении в
- 23. Australopithecus garhi Австралопитек гари возраст 2,5 млн лет (плейстоцен); найден в Эфиопии в 1997 году, описан
- 24. австралопитек гари
- 25. Homo erectus Человек прямоходящий Homo erectus произошел от африканского H.ergaster примерно 1.6 млн. лет назад и
- 26. Homo erectus
- 27. Орудия поздних Homo erectus
- 28. Homo neanderthalensis Неандерталец населял Европу и Западную Азию (от Испании до Алтая) 200 000 - 28
- 29. Особенности генотипа и фенотипа неандертальцев Неандертальцы, возможно, были рыжими и светлокожими. У них обнаружена мутация в
- 30. Для выяснения влияния «неандертальской» мутации гена MC1R на функцию меланокортинового рецептора, неандертальский ген был искусственно внедрен
- 31. неандерталец (слева) и кроманьонец (справа)
- 33. Скачать презентацию

Место человека в системе животного мира
Место человека в системе животного мира

Происхождение приматов
Конец мезозоя, лесные биотопы.
Предки – Насекомоядные.
Ближайшая родня – Рукокрылые.
Примитивные черты
Происхождение приматов
Конец мезозоя, лесные биотопы.
Предки – Насекомоядные.
Ближайшая родня – Рукокрылые.
Примитивные черты

Вероятно, все известные сегодня африканские плиоценовые гоминиды были всеядными.
Вероятно, все известные сегодня африканские плиоценовые гоминиды были всеядными.
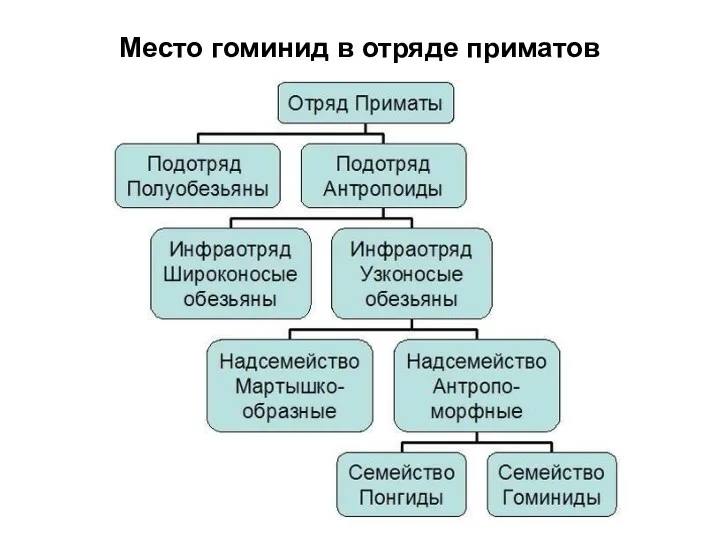
Место гоминид в отряде приматов
Место гоминид в отряде приматов
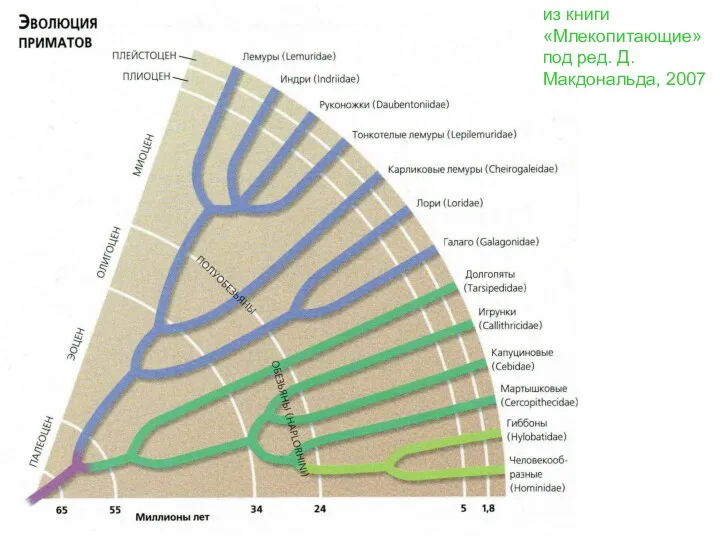
из книги «Млекопитающие» под ред. Д.Макдональда, 2007
из книги «Млекопитающие» под ред. Д.Макдональда, 2007

Центр происхождения человека - Восточная Африка.
Ближайший современный родственник человека - шимпанзе.
Дивергенция
Центр происхождения человека - Восточная Африка.
Ближайший современный родственник человека - шимпанзе.
Дивергенция

К вопросу о двуногости
и прямохождении
К вопросу о двуногости
и прямохождении

Для определения прямохождения у ископаемых приматов используют признаки:
– положение затылочного отверстия
Для определения прямохождения у ископаемых приматов используют признаки:
– положение затылочного отверстия

– строение таза - у прямоходящих таз широкий и низкий (такое

– строение стопы - у прямоходящих выражен свод стопы, пальцы прямые,
– строение стопы - у прямоходящих выражен свод стопы, пальцы прямые,

Таким образом, прямохождение возникло более 6 миллионов лет назад, но еще

Горилла ходит, опираясь на согнутую кисть
Горилла ходит, опираясь на согнутую кисть

Бонобо (карликовый шимпанзе) ходит на полусогнутых задних ногах
Бонобо (карликовый шимпанзе) ходит на полусогнутых задних ногах


Орангутан ранее других человекообразных отделился от общего ствола, сохранив исходный тип
Орангутан ранее других человекообразных отделился от общего ствола, сохранив исходный тип

Версии происхождения бипедии
На открытых равнинах саванн, среди высокой травы выпрямление тела
Версии происхождения бипедии
На открытых равнинах саванн, среди высокой травы выпрямление тела

Sahelanthropus tchadensis
Сахелантроп «Тумай»
возраст 6–7 млн. лет (миоцен);
найден в 2001 г.
Sahelanthropus tchadensis
Сахелантроп «Тумай»
возраст 6–7 млн. лет (миоцен);
найден в 2001 г.

сахелантроп
сахелантроп

Orrorin tugenensis
Оррорин
возраст 6 млн лет (миоцен);
найден в 2000 г. французской экспедицией
Orrorin tugenensis
Оррорин
возраст 6 млн лет (миоцен);
найден в 2000 г. французской экспедицией

оррорин
оррорин

Australopithecus anamensis
Австралопитек озерный
возраст 4,1-4,2 млн лет (плиоцен);
найден группой Мив
Australopithecus anamensis
Австралопитек озерный
возраст 4,1-4,2 млн лет (плиоцен);
найден группой Мив

Australopithecus garhi
Австралопитек гари
возраст 2,5 млн лет (плейстоцен);
найден в Эфиопии
Australopithecus garhi
Австралопитек гари
возраст 2,5 млн лет (плейстоцен);
найден в Эфиопии
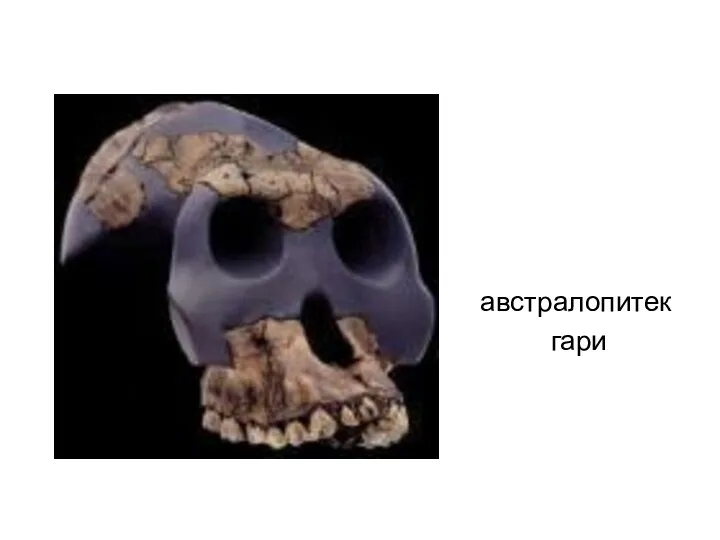
австралопитек
гари
австралопитек
гари

Homo erectus
Человек прямоходящий
Homo erectus произошел от африканского H.ergaster примерно 1.6 млн.
Homo erectus
Человек прямоходящий
Homo erectus произошел от африканского H.ergaster примерно 1.6 млн.

Homo erectus
Homo erectus

Орудия поздних Homo erectus
Орудия поздних Homo erectus

Homo neanderthalensis
Неандерталец
населял Европу и Западную Азию (от Испании до Алтая) 200
Homo neanderthalensis
Неандерталец
населял Европу и Западную Азию (от Испании до Алтая) 200

Особенности генотипа и фенотипа неандертальцев
Неандертальцы, возможно, были рыжими и светлокожими.
Особенности генотипа и фенотипа неандертальцев
Неандертальцы, возможно, были рыжими и светлокожими.

Для выяснения влияния «неандертальской» мутации гена MC1R на функцию меланокортинового рецептора,
Для выяснения влияния «неандертальской» мутации гена MC1R на функцию меланокортинового рецептора,
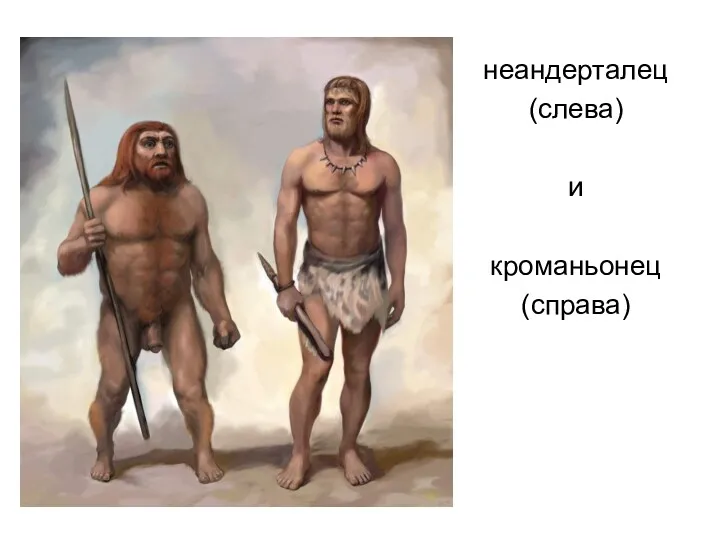
неандерталец
(слева)
и
кроманьонец
(справа)
неандерталец
(слева)
и
кроманьонец
(справа)
 Нервная система, органы чувств, эндокринная система
Нервная система, органы чувств, эндокринная система Опора и движение
Опора и движение Генетика пола. Сцепленное с полом наследование
Генетика пола. Сцепленное с полом наследование Немецкая овчарка
Немецкая овчарка Дикие и домашние животные
Дикие и домашние животные Мир комнатных растений. Окружающий мир 2 класс
Мир комнатных растений. Окружающий мир 2 класс Аурулардың патогенді факторлары. Дәнді дақылдардың саңырауқұлақ аурулары
Аурулардың патогенді факторлары. Дәнді дақылдардың саңырауқұлақ аурулары Урок по ФГОС . История изучения клетки. Методы изучения клетки. Лабораторная работа Строение клеток кожицы чешуи лука.
Урок по ФГОС . История изучения клетки. Методы изучения клетки. Лабораторная работа Строение клеток кожицы чешуи лука. Вирусы в биотехнологии
Вирусы в биотехнологии Цитология - наука о клетке
Цитология - наука о клетке Строение и функции пищеварительной системы
Строение и функции пищеварительной системы Царство Гриби. Вищі гриби. Шапкові гриби
Царство Гриби. Вищі гриби. Шапкові гриби Способы размножения. Митоз
Способы размножения. Митоз Предмет і завдання медичної мікробіології. Систематика і номенклатура мікроорганізмів
Предмет і завдання медичної мікробіології. Систематика і номенклатура мікроорганізмів Обмен липидов
Обмен липидов Мастер-класс Использование MimioStudio в учебном процессе
Мастер-класс Использование MimioStudio в учебном процессе Доказательства и основные этапы антропогенеза
Доказательства и основные этапы антропогенеза Дыхание. Значение дыхания в живых организмах
Дыхание. Значение дыхания в живых организмах Механизмы эволюции. Микроэволюция. Вид, популяция и её характеристики
Механизмы эволюции. Микроэволюция. Вид, популяция и её характеристики Бурундуки
Бурундуки Биологические полимеры – нуклеиновые кислоты
Биологические полимеры – нуклеиновые кислоты ПрезентацияХрящевые рыбы
ПрезентацияХрящевые рыбы Красная книга растений
Красная книга растений Кожа. Особенности строения. Функции кожи
Кожа. Особенности строения. Функции кожи Черенкование комнатных растений. Лабораторная работа № 5
Черенкование комнатных растений. Лабораторная работа № 5 Собственно бесполое размножение растений
Собственно бесполое размножение растений Живое клеточный уровень. Живое организменный уровень ботаника. Живое организменный уровень зоология
Живое клеточный уровень. Живое организменный уровень ботаника. Живое организменный уровень зоология Задание к теме:формы естественного отбора.
Задание к теме:формы естественного отбора.