- Физиология мышц. Нервно-мышечный синапс. Двигательные единицы. (Лекция 2)

Содержание
- 2. Мышечное волокно (миоцит) основная структурная и функциональная единица соматической мышечной ткани;
- 3. Три вида мышц человека: поперечно-полосатые скелетные мышцы; поперечно-полосатая сердечная мышца; гладкие мышцы внутренних органов, кожи, сосудов.
- 5. Поперечно-полосатая мышца Сердечная мышца Гладкая мышца
- 6. Физиологические свойства мышц: 1. возбудимость 2. сократимость 3. рефрактерность 4. лабильность 5. проводимость 6. автоматизм
- 7. Возбудимость способность приходить в состояние возбуждения при действии раздражителей.
- 8. Сократимость способность мышцы изменять свою длину или напряжение в ответ на действие раздражителя.
- 9. Рефрактерность кратковременное снижение возбудимости нервной и мышечной тканей непосредственно вслед за потенциалом действия. Рефрактерность обнаруживается при
- 10. Лабильность функциональная подвижность, скорость протекания элементарных циклов возбуждения в нервной и мышечной тканях.
- 11. Проводимость способность проводить возбуждение.
- 12. Автоматизм (Автоматия) способность клетки, ткани или органа к ритмической, периодической или апериодической самопроизвольной деятельности без очевидной
- 13. ФИЗИЧЕСКИЕ СВОЙСТВА МЫШЦ Растяжимость; Эластичность; Пластичность; Способность развивать силу; Способность совершать работу;
- 14. Растяжимость способность мышцы изменять свою длину под действием (влиянием) приложенной растягивающей её силы.
- 15. Эластичность способность мышцы принимать (восстанавливать) свою первоначальную длину после прекращения действия растягивающей или деформирующей её силы.
- 16. Сила мышцы. определяется максимальным грузом, который мышца в состоянии поднять.
- 17. Способность мышцы совершать работу. Работа мышцы определяется произведением величины поднятого груза на высоту подъема.
- 18. Пластичность (для гладких мышц) способность удерживать исходное напряжение при увеличении растяжения (пример: наполнение желудка или желчного
- 19. Функции скелетных мышц: передвижение тела в пространстве; перемещение частей тела относительно друг друга (динамическая функция); поддержание
- 20. Функции скелетных мышц: двигательная активность важнейший антистрессовый фактор - «движение - это жизнь»; депонирование воды и
- 21. Мышца комплекс мышечных волокон (клеток), связанных соединительной тканью.
- 22. Скелетная мышца. Скелетные мышцы составляют 40% от массы тела.
- 23. Мышечное волокно - основная структурная и функциональная единица соматической мышечной ткани
- 24. Мышечное волокно
- 25. Сарколемма мембрана окружающая мышечное волокно, сходна по электрическим свойствам с мембраной нервных клеток.
- 27. В состав мышечных волокон входит большое количество миофибрилл, которые и создают характерную поперечную исчерченность.
- 29. Миофибрилла нитевидная структура, состоящая из саркомеров.
- 30. Миофибриллы органеллы клеток поперечнополосатых мышц, обеспечивающие их сокращение. Служат для сокращений мышечных волокон. Миофибриллы создают характерную
- 31. Структура миофибрилл Сведения о белковой структуре компонентов поперечнополосатых мышц получены в середине XIX века В. Кюне.
- 32. Саркомер базовая структурно-функциональная сократительная единица поперечнополосатой мышечной ткани, участок миофибриллы между двумя соседними Z-линиями (Z-мебранами). комплекс
- 33. Расположение саркомеров совпадают, это и создает картину "полосатости", или поперечной исчерченности волокна.
- 34. Митохондрия двумембранный органоид эукариотической клетки, основная функция которого синтез АТФ –источника энергии для жизнедеятельности клетки. Большое
- 36. Саркоплазматический ретикулум (СР) пространство между миофибриллами заполненное саркоплазмой, в которой находится сложная система трубок и цистерн.
- 38. Актин глобулярный белок, состоящий из 376 аминокислотных остатков.
- 39. Актиновая (тонкая) нить образована двумя спиральными тяжами из глобулярных молекул актина (G - актин), закрученных один
- 40. В состав актинового филамента входят два вспомогательных белка - тропомиозин и тропонин Тропомиозин - белок, располагающийся
- 41. Миозин фибриллярный белок, один из главных компонентов сократительных волокон мышц - миофибрилл.
- 42. Миозин составляет 40-60 % общего количества мышечных белков. При соединении миозина с другим белком миофибрилл (актином)
- 43. Миозиновые нити структуры эукариотической клетки, прямо или косвенно участвующие в различных функциях клетки, связанных с движением.
- 44. Миозиновые нити имеют поперечные мостики с головками, которые отходят от нити биполярно. Головка обладает АТФ-азной активностью,
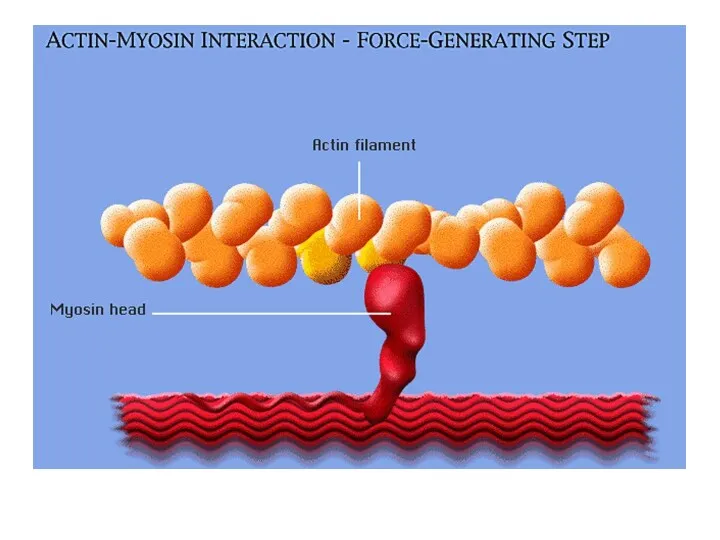
- 45. Взаимодействие актиновых и миозиновых филаментов
- 46. В процессе сокращения мышечного волокна в нем происходит электромеханическое сопряжение, включающее: а) электрохимическое; б) хемомеханическое преобразования;
- 47. Этапы генерирования сокращения мышцы: Стимуляция мышечного волокна. Возникновение потенциала действия. Проведение возбуждения по Т-системе. Высвобождение Са2+
- 48. ДВИГАТЕЛЬНЫЕ ЕДИНИЦЫ
- 49. Двигательная единица мотонейрон передних рогов спинного мозга и иннервируемые им мышечные волокна. Аксон мотонейрона внутри мышцы
- 50. Морфологической единицей мышцы является мышечное волокно. Функциональной единицей скелетной мускулатуры является двигательная единица.
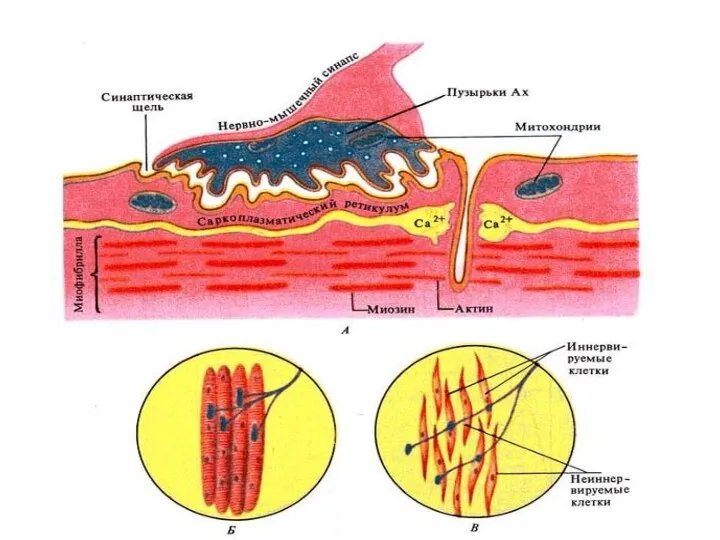
- 51. Схема двигательной единицы: а – мотонейрон; б – нервно-мышечные синапсы; в – мышечные волокна. Концевые веточки
- 52. НЕРВНО-МЫШЕЧНЫЙ СИНАПС
- 53. Нервно-мышечный синапс.
- 54. Нервно-мышечный синапс эффекторное нервное окончание на скелетном мышечном волокне. Входит в состав нервно-мышечного веретена. Нейромедиатором в
- 55. Эффекторный передающий импульсы от нервных центров к рабочим органам, тканям.
- 56. Медиатор (нейромедиатор) (химический посредник) биологически активное химическое вещество для передачи нервного импульса от одной клетки к
- 57. Взаимодействие медиатора с постсинаптической мембраной Медиатор диффундирует по синаптической жидкости и большая часть молекул его достигает
- 58. Нервно-мышечный синапс.
- 59. Внутри нервного окончания имеется большое количество (до 300.000) синаптических пузырьков (диаметром около 50 нм), содержащих химическое
- 60. Взаимодействие медиатора с постсинаптической мембраной Результатом взаимодействия АХ с ХР является открытие хемовозбудимых ионных каналов. Селективный
- 61. Выброс медиатора обеспечивает взаимодействие его с лигандзависимыми структурами канала
- 62. Лиганд - химическое соединение, образующее комплекс с биомолекулой (чаще белком , пример клеточным рецептором) и производящее
- 63. Передача возбуждения в нервно-мышечном синапсе. Нервный импульс достигает концевой веточки аксона и вызывает выделение медиатора ацетилхолина
- 77. Восстановление медиатора в синаптической бляшке В нервном волокне происходит постоянное пополнение медиатора. Механизмы восстановления везикул с
- 78. ТЕОРИЯ МЫШЕЧНОГО СОКРАЩЕНИЯ И РАССЛАБЛЕНИЯ
- 79. Теория скольжения нитей (Хаксли) В 1954 г. А. Хаксли при использовании микроскопа установил, что актиновые и
- 80. Механизмы сокращения мышечного волокна В покое мышечные волокна находятся в расслабленном состоянии. Концентрация ионов Са2+ в
- 81. При возбуждении мышечного волокна потенциал действия, возникающий в области концевой пластинки, распространяется по Т-системе вглубь волокна,
- 82. Под влиянием ионов Са2+ молекула тропонина изменяет свою форму и выталкивает тропомиозин в желобок между двумя
- 83. В покое миозиновая головка присоединяет АТФ, гидролизует её, оставаясь связанной с АДФ и фосфат-ионом. Миозиновые головки,
- 84. Этапы «шагового» механизма Последовательные этапы: а – расслабление, б – соединение миозиновых головок с активным центром
- 85. В состоянии покоя В условиях покоя головка миозина не прикреплена к активной зоне актиновой нити -
- 86. Шаг 1. Ионы кальция, выходя из СР запускают процесс сокращения. Под влиянием ионов Са2+ молекула тропонина
- 87. Шаг 2: Головка миозина прикрепляется к активному участку актина
- 88. Шаг 3: Фермент миозиновая АТФ-аза гидролизует АТФ, расположенный на головке миозина, что обеспечивает энергией поперечные мостики.
- 89. Шаг 4: Освобождающиеся при гидролизе АТФ молекула АДФ и неорганический фосфат используются для последующего ресинтеза АТФ
- 90. Шаг 5: Повторное прикрепление и отсоединение мостиков продолжается до тех пор, пока концентрация ионов Са2+ внутри
- 103. Саркомер в расслабленном состоянии Саркомер в состоянии сокращения
- 104. Роль кальция в мышечном сокращении 1 – Выброс медиатора в синаптическую щель. 2 – Освобождение активного
- 105. Механизм расслабления мышцы [Са+2] 107 М + кальциевый насос в СПР СПР Са+2 Са+2 Са+2 АТФ
- 106. Затраты энергии АТФ во время мышечного сокращения На поддержание ионной асимметрии (натрий-калиевй насос) Образование мостиков («приклеивание»
- 107. Роль АТФ в мышечном сокращении а) сокращения (образования мостиков); б) расслабления (разрыва мостиков); в) работы Са-насоса
- 108. Пути ресинтеза АТФ 1) креатинфосфокиназный (КФ): АДФ + КФ АТФ + К (3,6 моль АТФ/мин) 2)
- 109. Типы мышечного сокращения 1. Изотоническое — волокна мышцы укорачиваются без изменения их напряжения 2. Изометрическое -
- 110. Виды раздражения мышцы в эксперименте Непрямое раздражение – импульс воздействует на нервное окончание мотонейрона Прямое раздражение
- 111. Режимы мышечных сокращений Режим мышечных сокращений зависит от частоты импульсации мотонейронов.
- 112. Одиночное сокращение - - механический ответ мышечного волокна на однократное раздражение. Выделяют три фазы сокращения 1.
- 113. Фазы одиночного мышечного сокращения (ОМС)
- 114. Одиночное мышечное сокращение 1 2 3 1 – латентный период 2 – период напряжения 3 –
- 115. Соотношения фаз ОМС с фазами возбудимости в скелетных мышцах Красный график- фазы сокращения: 1 – латентный
- 117. Соотношение фаз ПД с фазами сокращения и возбудимости
- 118. Тетанус В естественных условиях к мышечным волокнам поступают не одиночные нервные импульсы, а их ряды. Когда
- 119. Суммация одиночных сокращений
- 120. Тетанус возникает вследствие суммации одиночных мышечных сокращений. Для того, чтобы возник тетанус, необходимо действие повторных раздражителей
- 121. Суммация мышечных сокращений
- 122. Если раздражающие импульсы сближены и каждый из них приходится на расслабления одиночного сокращения, то возникает зубчатый
- 123. Если раздражающие импульсы сближены настолько, что каждый последующий приходится на фазу сокращения, т.е. когда мышца не
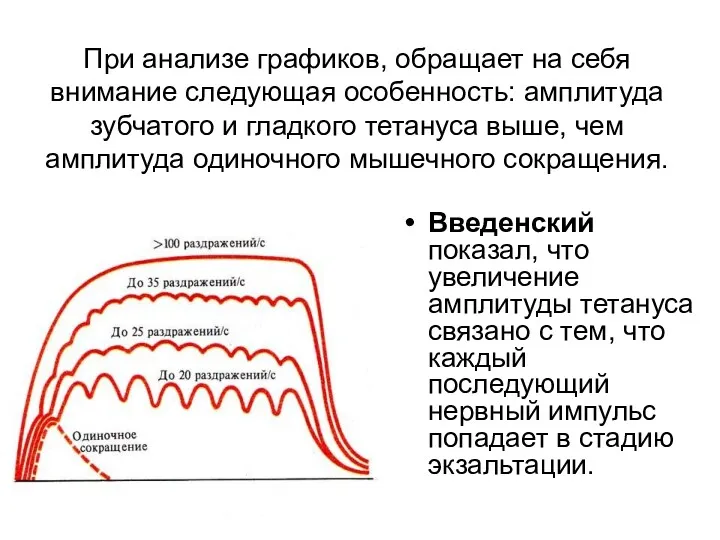
- 124. При анализе графиков, обращает на себя внимание следующая особенность: амплитуда зубчатого и гладкого тетануса выше, чем
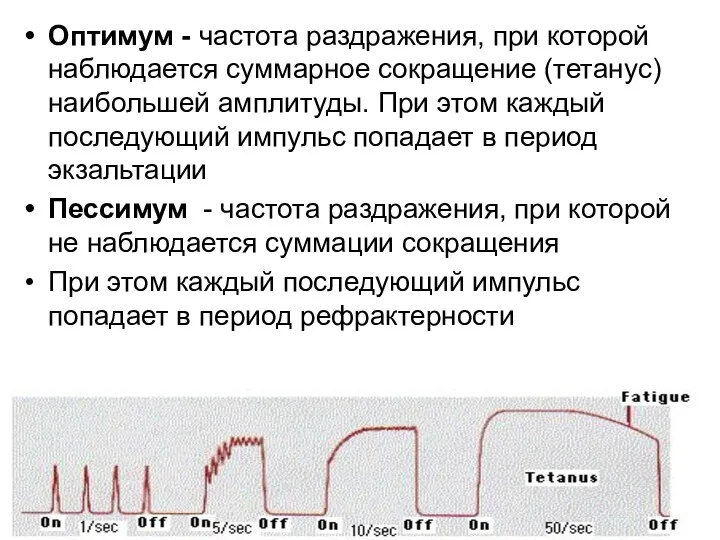
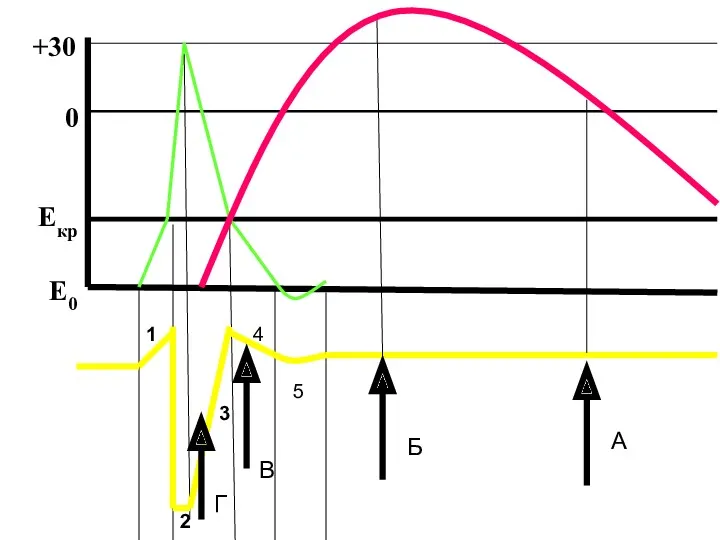
- 125. Оптимум - частота раздражения, при которой наблюдается суммарное сокращение (тетанус) наибольшей амплитуды. При этом каждый последующий
- 126. 1 2 3 4 5 Е0 Екр 0 +30 В Г А Б
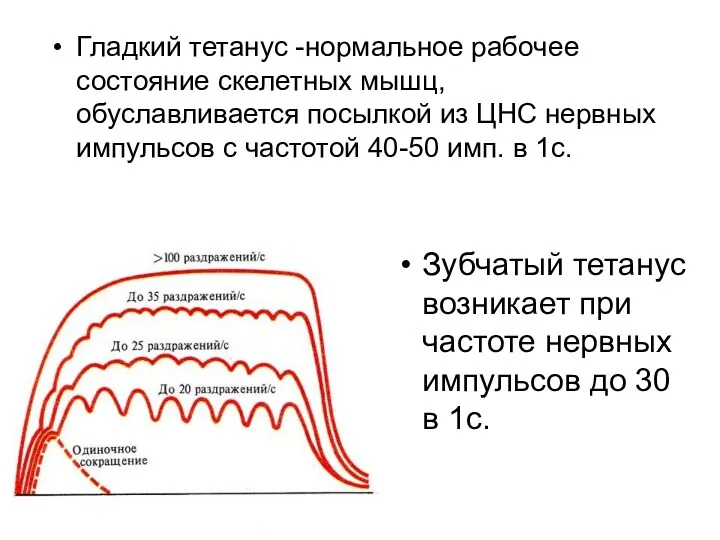
- 127. Зубчатый тетанус возникает при частоте нервных импульсов до 30 в 1с. Гладкий тетанус -нормальное рабочее состояние
- 128. Т.о., выделяют режимы мышечных сокращений: 1)….2)…3)…..4)….5)…. Режимы мышечных сокращений зависят от частоты импульсации мотонейронов
- 129. Сила мышц Зависит от толщины мышцы и её поперечного физиологического сечения
- 130. Работа мышцы Это энергия, затрачиваемая на перемещение тела с определенной силой на определенное расстояние: A =
- 131. утомление Процесс временного снижения работоспособности мышцы. Возникает в связи с уменьшением энергетических запасов (АТФ) в мышечном
- 132. Двигательные единицы - Единичное нервное волокно мотонейрона и, иннервируемые им мышечные волокна, составляют одну ДЕ 1
- 133. Виды двигательных единиц: 1. медленные, малоутомляемые (красные волокна) 2. быстрые, легко утомляемые (белые волокна) 3. быстрые,
- 134. Функциональные особенности медленных ДЕ 1. Иннервируются высоко возбудимыми а-мотонейронами с низкой скоростью проведения возбуждения по аксону
- 135. Функциональные особенности медленных ДЕ 4. Имеют хорошее кровоснабжение, много митохондрий, миоглобина, высокий аэробный обмен, поэтому обладают
- 136. Функциональные особенности быстрых ДЕ. 1. Иннервируются крупными, менее возбудимыми а-мотонейронами с высокой скоростью проведения ПД по
- 137. Функциональные особенности быстрых ДЕ. 4. Имеют слаборазвитую капиллярную сеть, мало митохондрий, миоглобина, но содержат много гликолитических
- 138. Функциональные особенности быстрых, устойчивых к утомлению ДЕ. По структурно-функциональным свойствам занимают среднее положение между медленными и
- 139. ФИЗИОЛОГИЯ ГЛАДКИХ МЫШЦ гладкая (непроизвольная) мускулатура находятся в стенках внутренних органах, сосудах и коже обеспечивают функцию
- 140. Структурные особенности ГМК миофиламенты содержат актин и миозин, которые распределены менее упорядочено, чем в волокнах скелетной
- 141. Физиологические особенности ГМК Потенциал покоя -30 до -70 мВ Потенциал действия двух типов: пикоподобные и плато.
- 142. Свойства гладких мышц сила сокращения зависит от степени растяжения мышцы (прямо-пропорционально) пластичность - способность сохранять приданную
- 145. Скачать презентацию

Мышечное волокно (миоцит)
основная структурная и функциональная единица соматической мышечной ткани;
Мышечное волокно (миоцит)
основная структурная и функциональная единица соматической мышечной ткани;

Три вида мышц человека:
поперечно-полосатые скелетные мышцы;
поперечно-полосатая сердечная мышца;
гладкие мышцы внутренних органов,
Три вида мышц человека:
поперечно-полосатые скелетные мышцы;
поперечно-полосатая сердечная мышца;
гладкие мышцы внутренних органов,
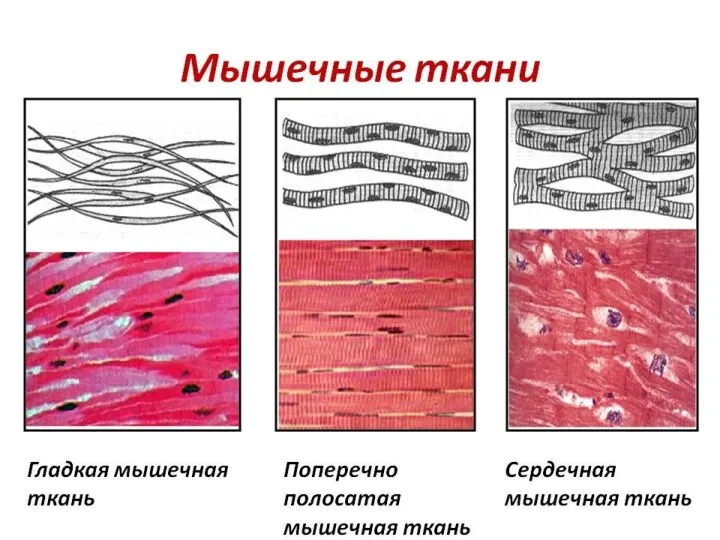
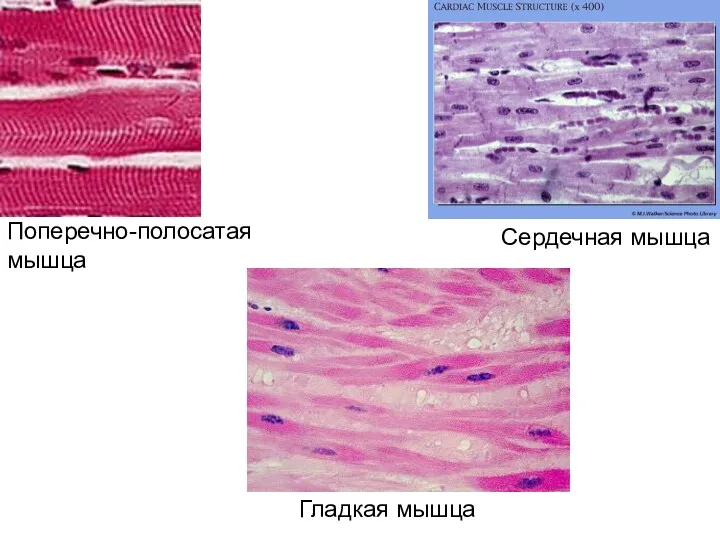
Поперечно-полосатая мышца
Сердечная мышца
Гладкая мышца
Поперечно-полосатая мышца
Сердечная мышца
Гладкая мышца

Физиологические свойства мышц:
1. возбудимость
2. сократимость
3. рефрактерность
4. лабильность
5. проводимость
6. автоматизм
Физиологические свойства мышц:
1. возбудимость
2. сократимость
3. рефрактерность
4. лабильность
5. проводимость
6. автоматизм

Возбудимость
способность приходить в состояние возбуждения при действии раздражителей.
Возбудимость
способность приходить в состояние возбуждения при действии раздражителей.

Сократимость
способность мышцы изменять свою длину или напряжение в ответ на
Сократимость
способность мышцы изменять свою длину или напряжение в ответ на

Рефрактерность
кратковременное снижение возбудимости нервной и мышечной тканей непосредственно вслед за потенциалом
Рефрактерность
кратковременное снижение возбудимости нервной и мышечной тканей непосредственно вслед за потенциалом

Лабильность
функциональная подвижность, скорость протекания элементарных циклов возбуждения в нервной и мышечной
Лабильность
функциональная подвижность, скорость протекания элементарных циклов возбуждения в нервной и мышечной

Проводимость
способность проводить возбуждение.
Проводимость
способность проводить возбуждение.

Автоматизм (Автоматия)
способность клетки, ткани или органа к ритмической, периодической или
Автоматизм (Автоматия)
способность клетки, ткани или органа к ритмической, периодической или

ФИЗИЧЕСКИЕ СВОЙСТВА МЫШЦ
Растяжимость;
Эластичность;
Пластичность;
Способность развивать силу;
Способность совершать работу;
ФИЗИЧЕСКИЕ СВОЙСТВА МЫШЦ
Растяжимость;
Эластичность;
Пластичность;
Способность развивать силу;
Способность совершать работу;

Растяжимость
способность мышцы изменять свою длину под действием (влиянием) приложенной растягивающей её
Растяжимость
способность мышцы изменять свою длину под действием (влиянием) приложенной растягивающей её

Эластичность
способность мышцы принимать (восстанавливать) свою первоначальную длину после прекращения действия растягивающей
Эластичность
способность мышцы принимать (восстанавливать) свою первоначальную длину после прекращения действия растягивающей

Сила мышцы.
определяется максимальным грузом, который мышца в состоянии поднять.
Сила мышцы.
определяется максимальным грузом, который мышца в состоянии поднять.

Способность мышцы совершать работу.
Работа мышцы определяется произведением величины поднятого груза на высоту
Способность мышцы совершать работу.
Работа мышцы определяется произведением величины поднятого груза на высоту

Пластичность (для гладких мышц)
способность удерживать исходное напряжение при увеличении растяжения
Пластичность (для гладких мышц)
способность удерживать исходное напряжение при увеличении растяжения

Функции скелетных мышц:
передвижение тела в пространстве;
перемещение частей тела относительно друг друга
Функции скелетных мышц:
передвижение тела в пространстве;
перемещение частей тела относительно друг друга

Функции скелетных мышц:
двигательная активность важнейший антистрессовый фактор - «движение - это
Функции скелетных мышц:
двигательная активность важнейший антистрессовый фактор - «движение - это

Мышца
комплекс мышечных волокон (клеток), связанных соединительной тканью.
Мышца
комплекс мышечных волокон (клеток), связанных соединительной тканью.
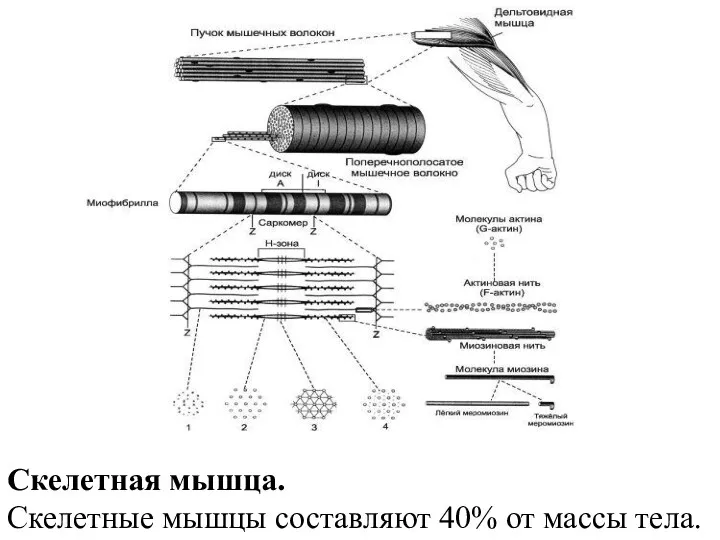
Скелетная мышца.
Скелетные мышцы составляют 40% от массы тела.
Скелетная мышца.
Скелетные мышцы составляют 40% от массы тела.
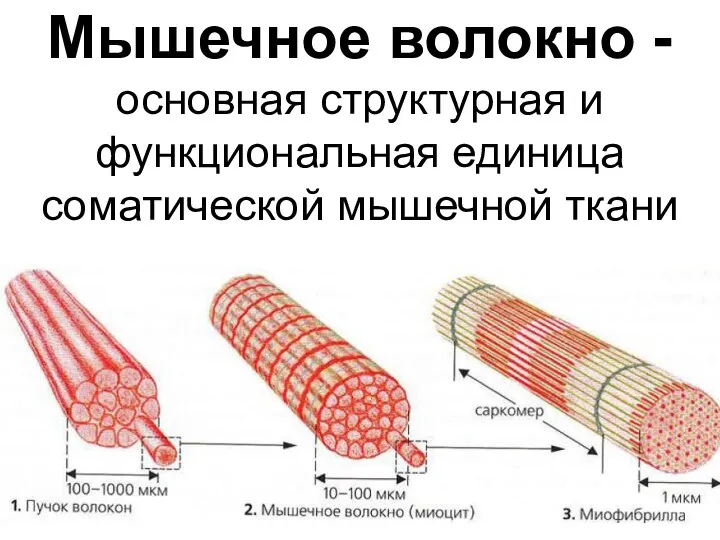
Мышечное волокно - основная структурная и функциональная единица соматической мышечной ткани
Мышечное волокно - основная структурная и функциональная единица соматической мышечной ткани
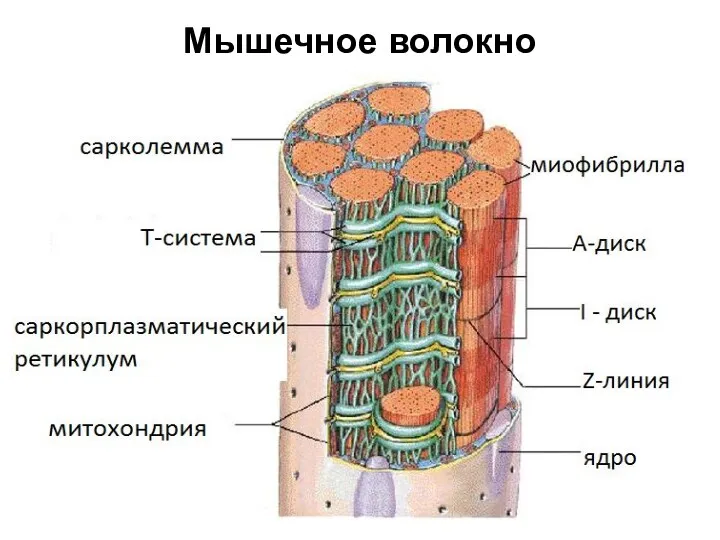
Мышечное волокно
Мышечное волокно

Сарколемма
мембрана окружающая мышечное волокно, сходна по электрическим свойствам с мембраной нервных
Сарколемма
мембрана окружающая мышечное волокно, сходна по электрическим свойствам с мембраной нервных
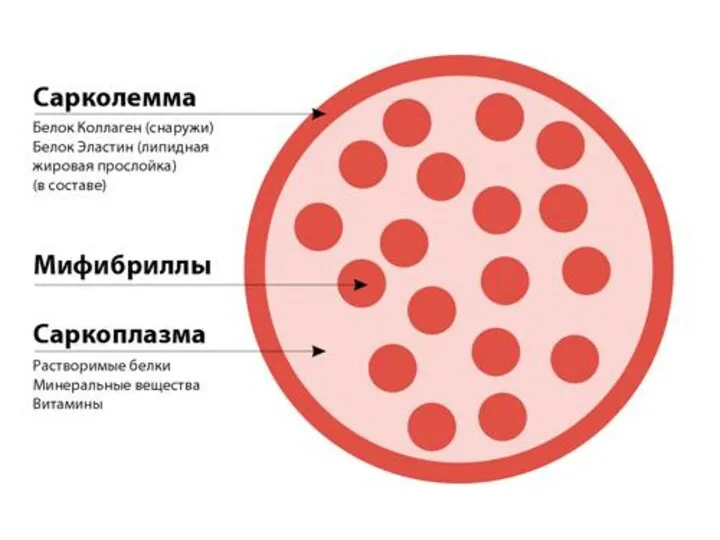

В состав мышечных волокон
входит большое количество миофибрилл, которые и создают
В состав мышечных волокон
входит большое количество миофибрилл, которые и создают
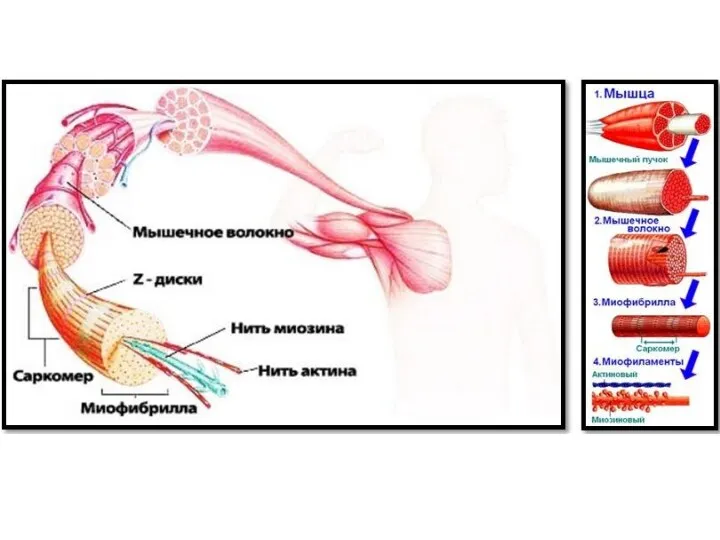

Миофибрилла
нитевидная структура, состоящая из саркомеров.
Миофибрилла
нитевидная структура, состоящая из саркомеров.

Миофибриллы
органеллы клеток поперечнополосатых мышц, обеспечивающие их сокращение.
Служат для сокращений мышечных волокон.
Миофибриллы
органеллы клеток поперечнополосатых мышц, обеспечивающие их сокращение.
Служат для сокращений мышечных волокон.

Структура миофибрилл
Сведения о белковой структуре компонентов поперечнополосатых мышц получены в середине
Структура миофибрилл
Сведения о белковой структуре компонентов поперечнополосатых мышц получены в середине

Саркомер
базовая структурно-функциональная сократительная единица поперечнополосатой мышечной ткани, участок миофибриллы между двумя
Саркомер
базовая структурно-функциональная сократительная единица поперечнополосатой мышечной ткани, участок миофибриллы между двумя
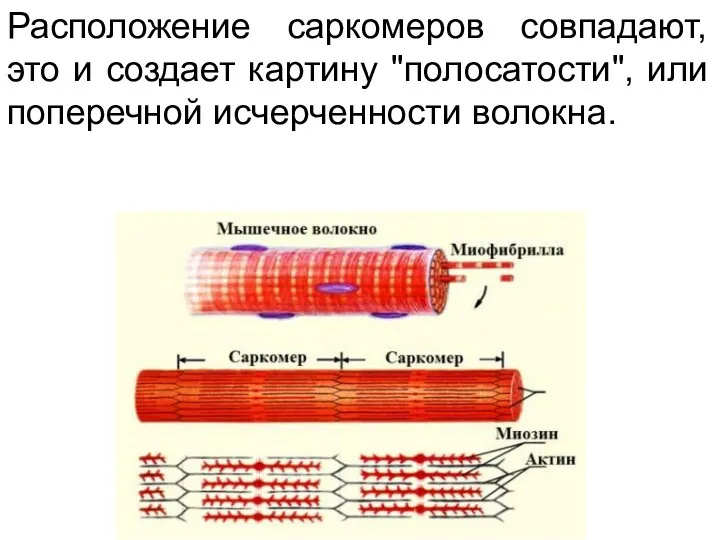
Расположение саркомеров совпадают, это и создает картину "полосатости", или поперечной исчерченности
Расположение саркомеров совпадают, это и создает картину "полосатости", или поперечной исчерченности

Митохондрия
двумембранный органоид эукариотической клетки, основная функция которого синтез АТФ –источника энергии для жизнедеятельности клетки.
Большое
Митохондрия
двумембранный органоид эукариотической клетки, основная функция которого синтез АТФ –источника энергии для жизнедеятельности клетки.
Большое
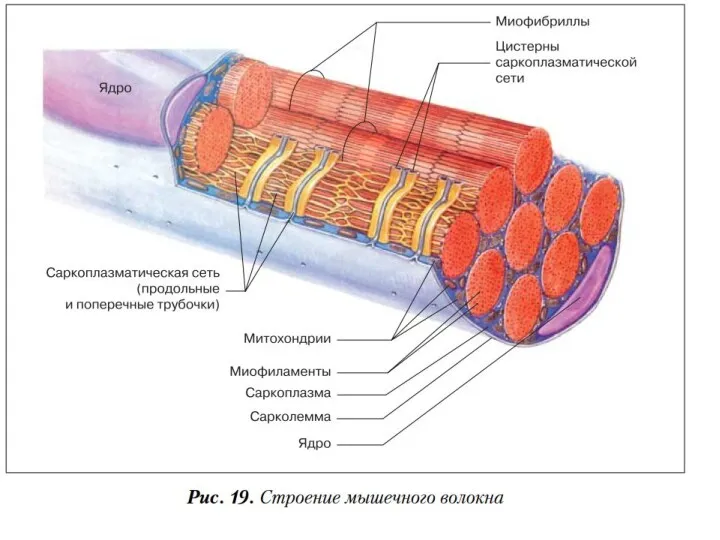

Саркоплазматический ретикулум (СР)
пространство между миофибриллами заполненное саркоплазмой, в которой находится
Саркоплазматический ретикулум (СР)
пространство между миофибриллами заполненное саркоплазмой, в которой находится


Актин
глобулярный белок, состоящий из 376 аминокислотных остатков.
Актин
глобулярный белок, состоящий из 376 аминокислотных остатков.
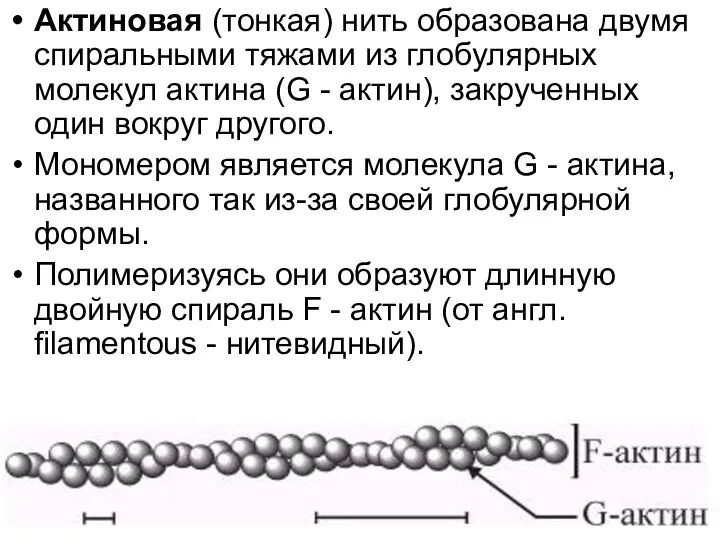
Актиновая (тонкая) нить образована двумя спиральными тяжами из глобулярных молекул актина
Актиновая (тонкая) нить образована двумя спиральными тяжами из глобулярных молекул актина
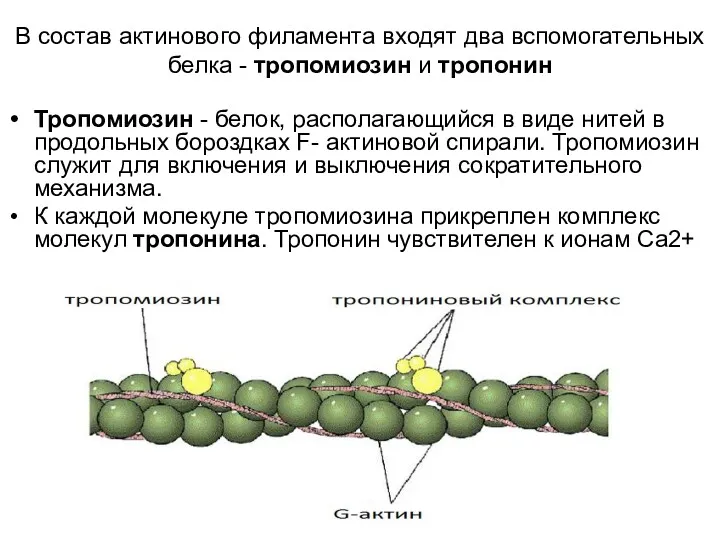
В состав актинового филамента входят два вспомогательных белка - тропомиозин и
В состав актинового филамента входят два вспомогательных белка - тропомиозин и

Миозин
фибриллярный белок, один из главных компонентов сократительных волокон мышц - миофибрилл.
Миозин
фибриллярный белок, один из главных компонентов сократительных волокон мышц - миофибрилл.

Миозин составляет 40-60 % общего количества мышечных белков.
При соединении миозина с другим
Миозин составляет 40-60 % общего количества мышечных белков.
При соединении миозина с другим

Миозиновые нити
структуры эукариотической клетки, прямо или косвенно участвующие в различных функциях клетки,
Миозиновые нити
структуры эукариотической клетки, прямо или косвенно участвующие в различных функциях клетки,
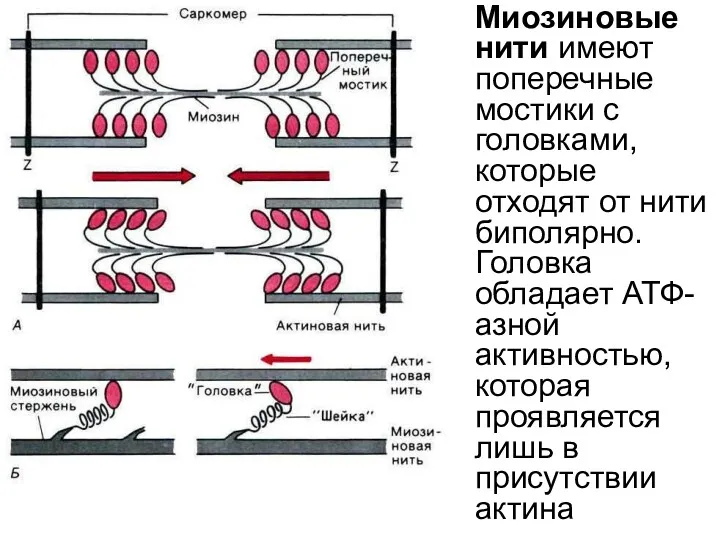
Миозиновые нити имеют поперечные мостики с головками, которые отходят от нити
Миозиновые нити имеют поперечные мостики с головками, которые отходят от нити
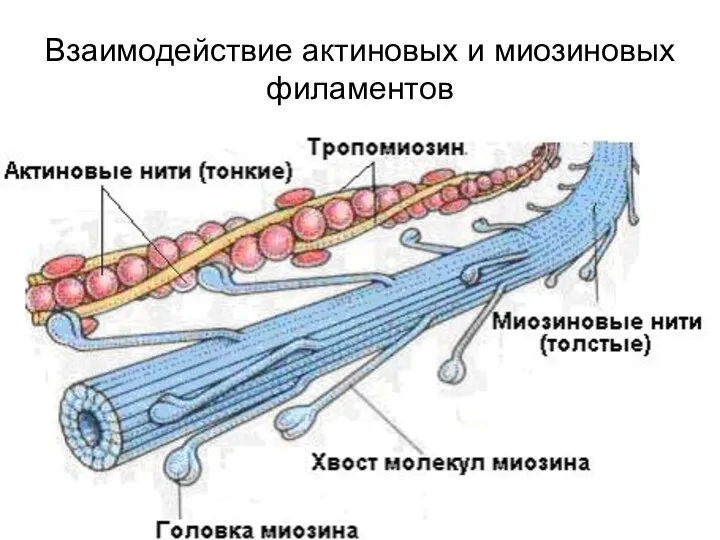
Взаимодействие актиновых и миозиновых филаментов
Взаимодействие актиновых и миозиновых филаментов

В процессе сокращения мышечного волокна
в нем происходит электромеханическое сопряжение, включающее:
а)
В процессе сокращения мышечного волокна
в нем происходит электромеханическое сопряжение, включающее:
а)

Этапы генерирования сокращения мышцы:
Стимуляция мышечного волокна.
Возникновение потенциала действия.
Проведение возбуждения по Т-системе.
Высвобождение
Этапы генерирования сокращения мышцы:
Стимуляция мышечного волокна.
Возникновение потенциала действия.
Проведение возбуждения по Т-системе.
Высвобождение

ДВИГАТЕЛЬНЫЕ ЕДИНИЦЫ
ДВИГАТЕЛЬНЫЕ ЕДИНИЦЫ

Двигательная единица
мотонейрон передних рогов спинного мозга и иннервируемые им мышечные
Двигательная единица
мотонейрон передних рогов спинного мозга и иннервируемые им мышечные

Морфологической единицей мышцы является мышечное волокно.
Функциональной единицей скелетной мускулатуры является
Функциональной единицей скелетной мускулатуры является

Схема двигательной единицы:
а – мотонейрон;
б – нервно-мышечные синапсы;
в
Схема двигательной единицы:
а – мотонейрон;
б – нервно-мышечные синапсы;
в

НЕРВНО-МЫШЕЧНЫЙ СИНАПС
НЕРВНО-МЫШЕЧНЫЙ СИНАПС
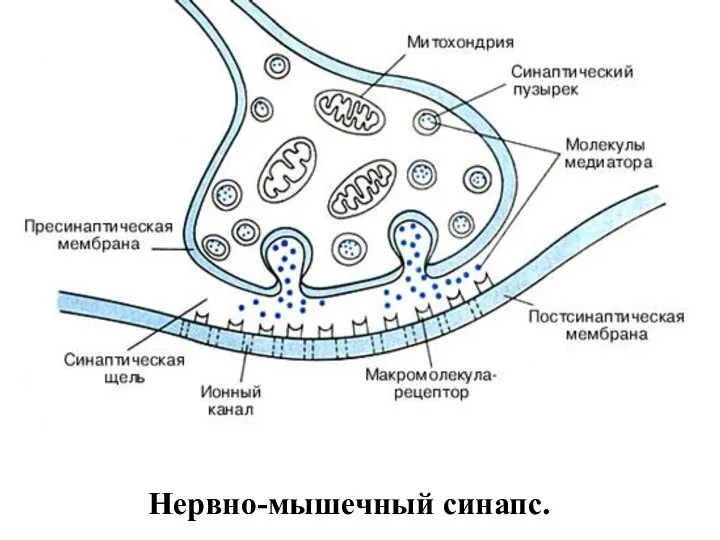
Нервно-мышечный синапс.
Нервно-мышечный синапс.

Нервно-мышечный синапс
эффекторное нервное окончание на скелетном мышечном волокне.
Входит в
Нервно-мышечный синапс
эффекторное нервное окончание на скелетном мышечном волокне.
Входит в

Эффекторный
передающий импульсы от нервных центров к рабочим органам, тканям.
Эффекторный
передающий импульсы от нервных центров к рабочим органам, тканям.

Медиатор (нейромедиатор)
(химический посредник)
биологически активное химическое вещество для передачи нервного импульса от
Медиатор (нейромедиатор)
(химический посредник)
биологически активное химическое вещество для передачи нервного импульса от

Взаимодействие медиатора с постсинаптической мембраной
Медиатор диффундирует по синаптической жидкости и большая
Взаимодействие медиатора с постсинаптической мембраной
Медиатор диффундирует по синаптической жидкости и большая
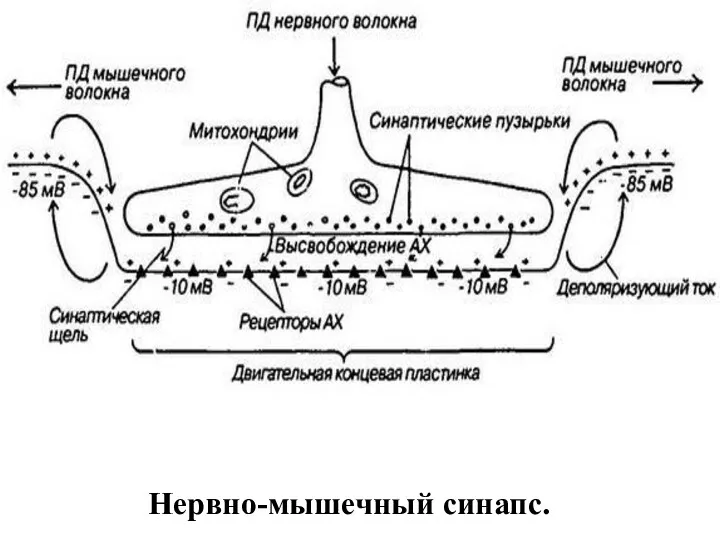
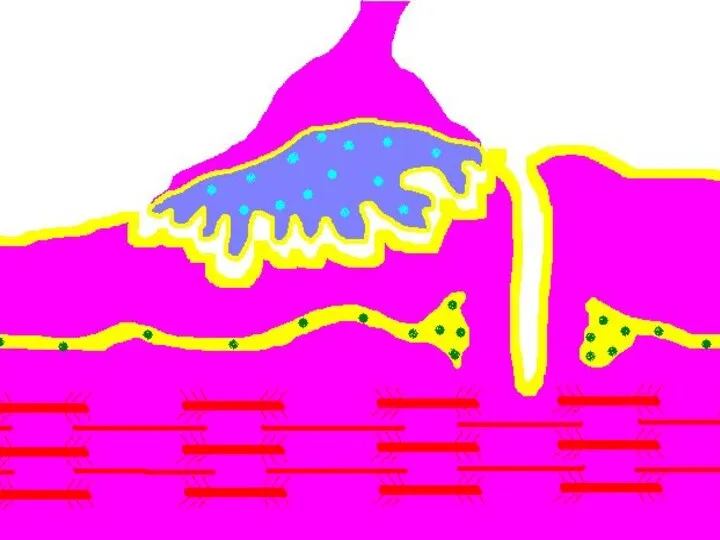
Нервно-мышечный синапс.
Нервно-мышечный синапс.

Внутри нервного окончания имеется большое количество (до 300.000) синаптических пузырьков (диаметром
Внутри нервного окончания имеется большое количество (до 300.000) синаптических пузырьков (диаметром

Взаимодействие медиатора с постсинаптической мембраной
Результатом взаимодействия АХ с ХР является открытие
Взаимодействие медиатора с постсинаптической мембраной
Результатом взаимодействия АХ с ХР является открытие
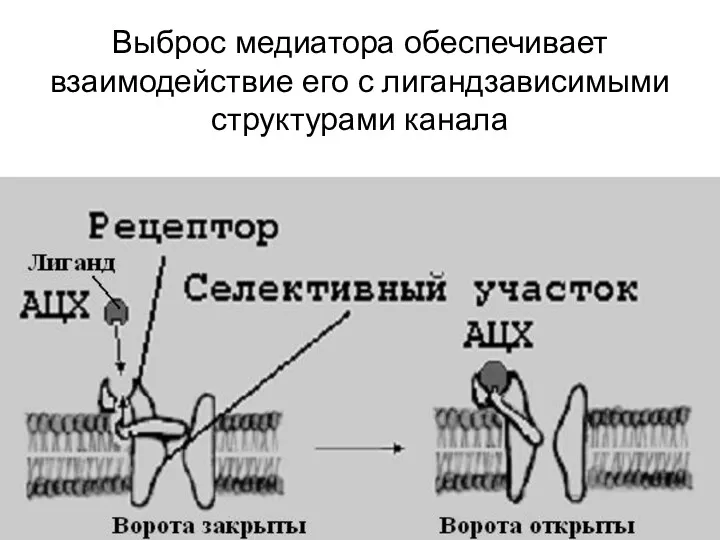
Выброс медиатора обеспечивает взаимодействие его с лигандзависимыми структурами канала
Выброс медиатора обеспечивает взаимодействие его с лигандзависимыми структурами канала

Лиганд
- химическое соединение, образующее комплекс с биомолекулой (чаще белком , пример
Лиганд
- химическое соединение, образующее комплекс с биомолекулой (чаще белком , пример

Передача возбуждения в нервно-мышечном синапсе.
Нервный импульс достигает концевой веточки аксона
Передача возбуждения в нервно-мышечном синапсе.
Нервный импульс достигает концевой веточки аксона
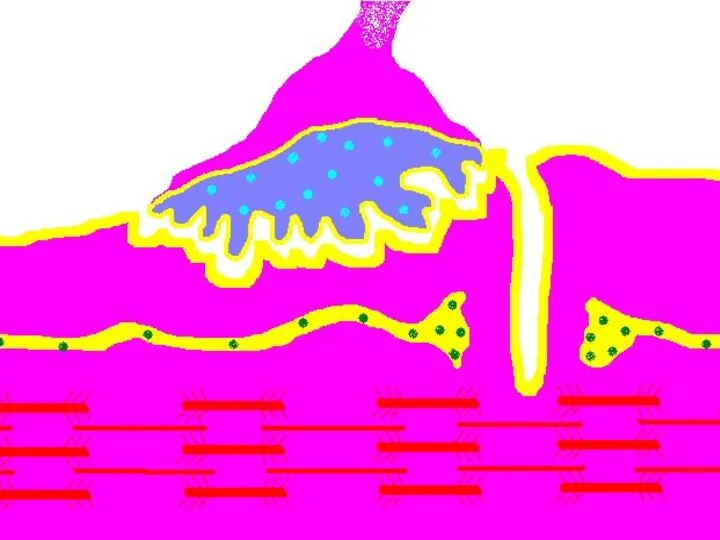


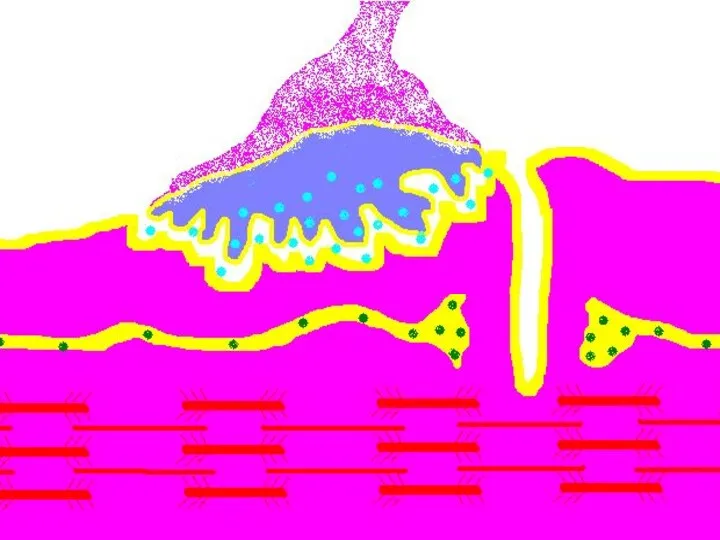
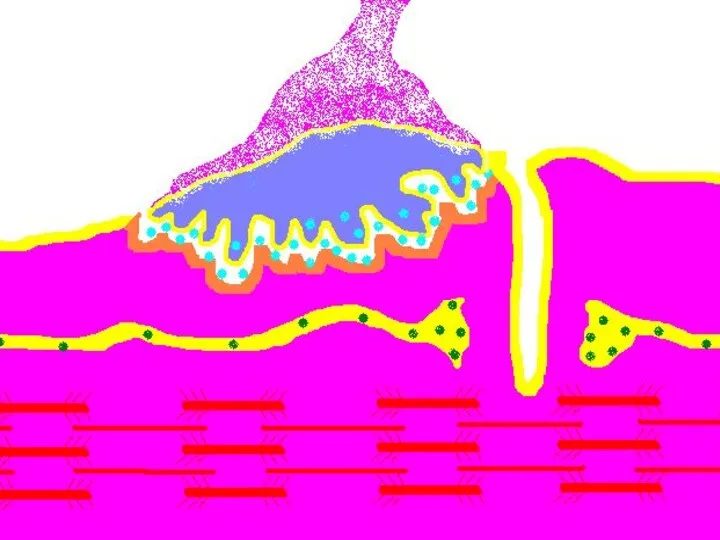
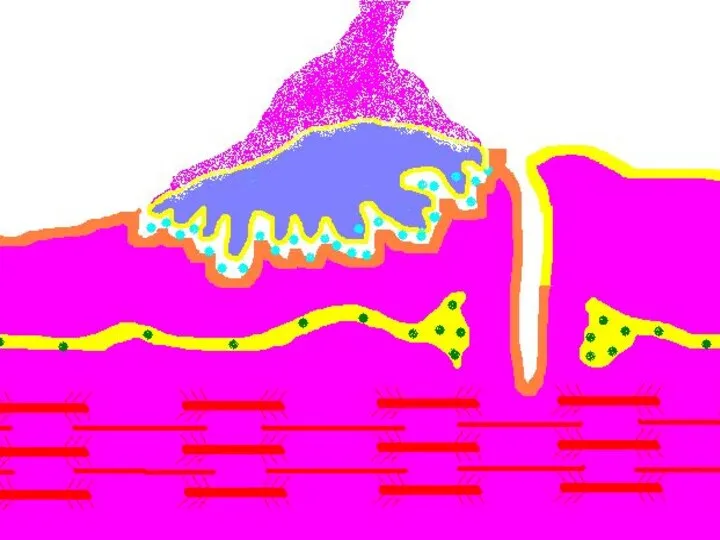
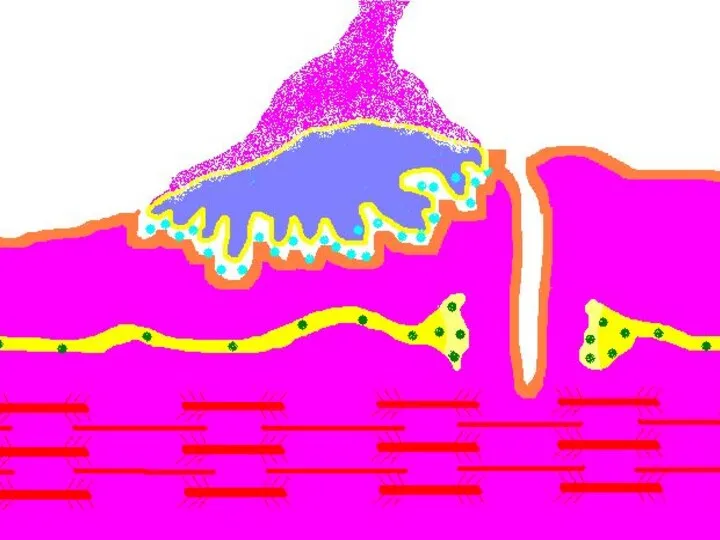
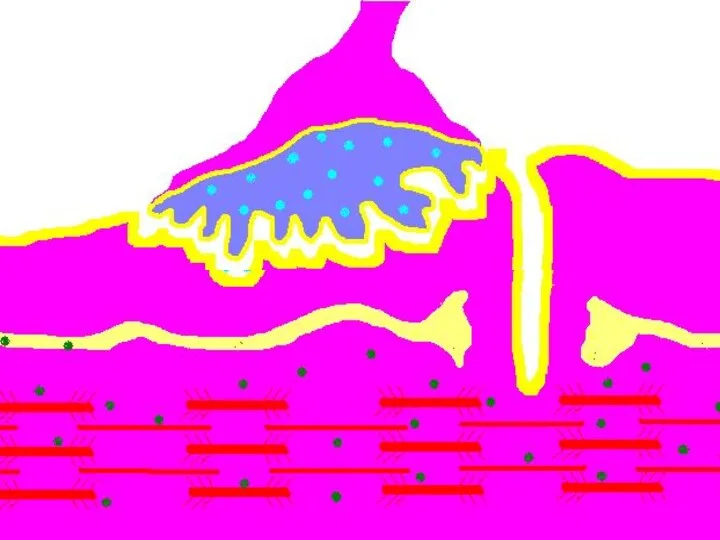
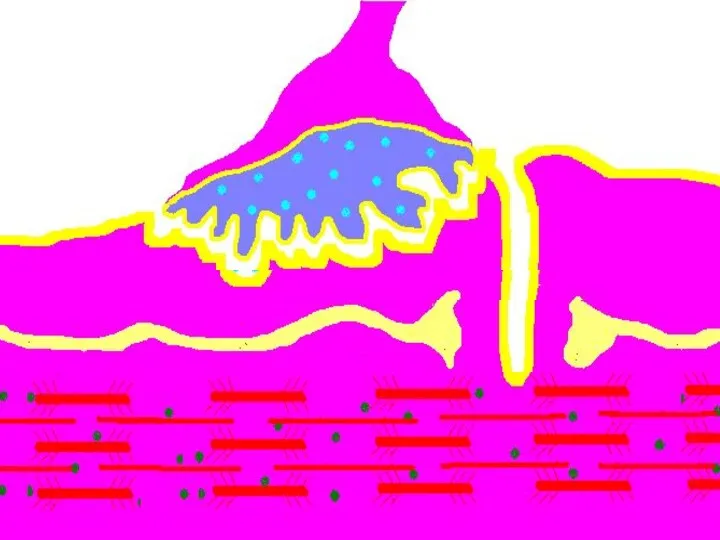
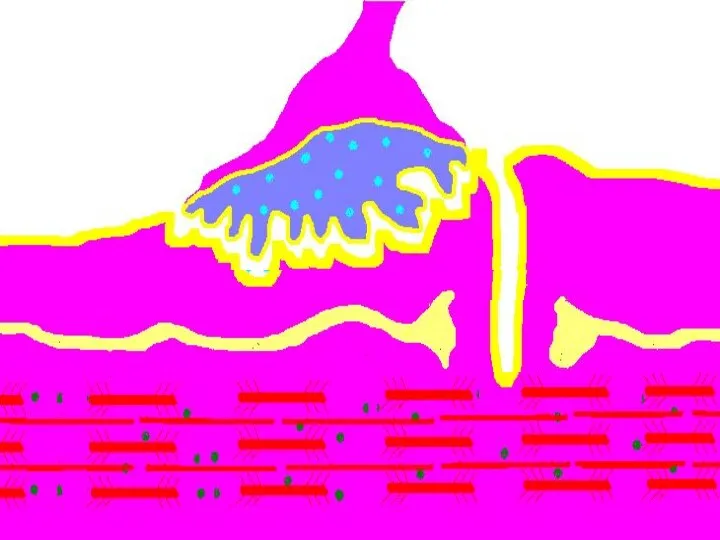
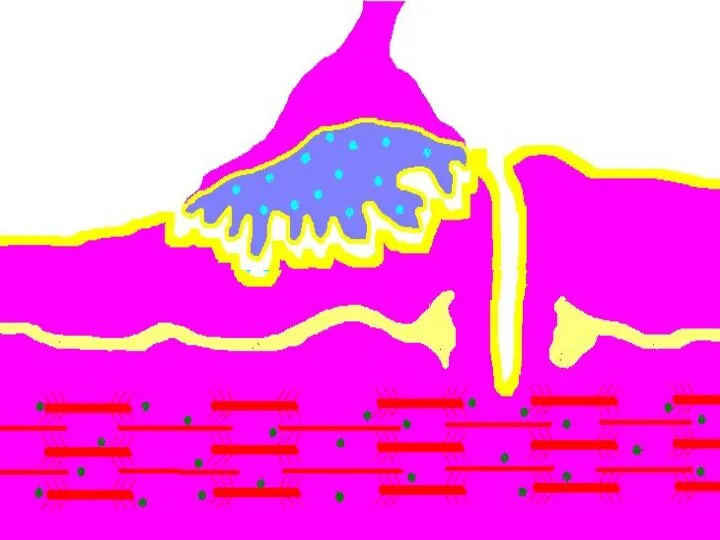

Восстановление медиатора в синаптической бляшке
В нервном волокне происходит постоянное пополнение
Восстановление медиатора в синаптической бляшке
В нервном волокне происходит постоянное пополнение

ТЕОРИЯ
МЫШЕЧНОГО СОКРАЩЕНИЯ
И
РАССЛАБЛЕНИЯ
ТЕОРИЯ
МЫШЕЧНОГО СОКРАЩЕНИЯ
И
РАССЛАБЛЕНИЯ

Теория скольжения нитей (Хаксли)
В 1954 г. А. Хаксли при использовании микроскопа
Теория скольжения нитей (Хаксли)
В 1954 г. А. Хаксли при использовании микроскопа

Механизмы сокращения мышечного волокна
В покое мышечные волокна находятся в расслабленном
Механизмы сокращения мышечного волокна
В покое мышечные волокна находятся в расслабленном

При возбуждении мышечного волокна потенциал действия, возникающий в области концевой пластинки,
При возбуждении мышечного волокна потенциал действия, возникающий в области концевой пластинки,

Под влиянием ионов Са2+ молекула тропонина изменяет свою форму и выталкивает
Под влиянием ионов Са2+ молекула тропонина изменяет свою форму и выталкивает

В покое миозиновая головка присоединяет АТФ, гидролизует её, оставаясь связанной с
В покое миозиновая головка присоединяет АТФ, гидролизует её, оставаясь связанной с
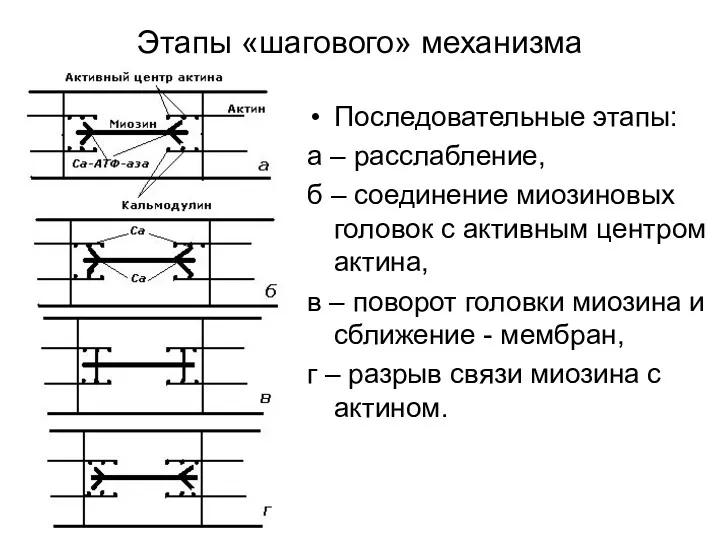
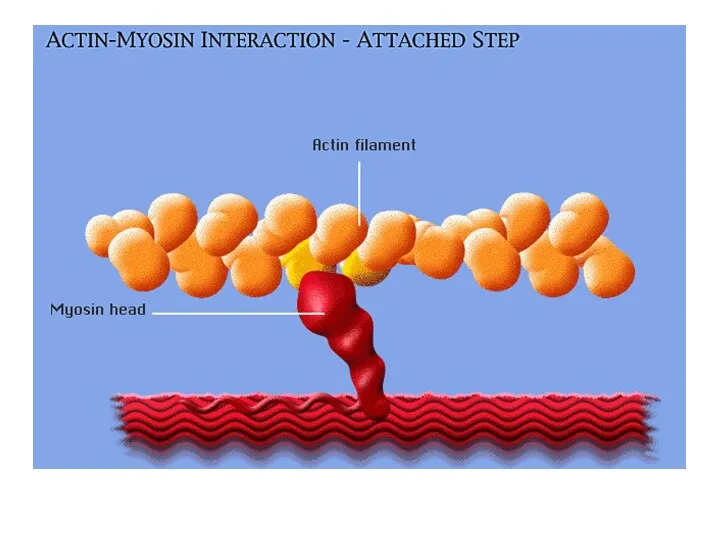
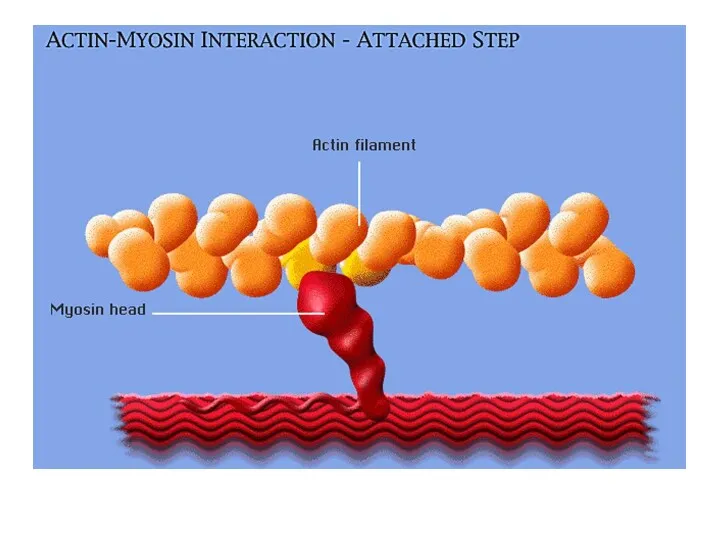
Этапы «шагового» механизма
Последовательные этапы:
а – расслабление,
б – соединение миозиновых головок с
Этапы «шагового» механизма
Последовательные этапы:
а – расслабление,
б – соединение миозиновых головок с
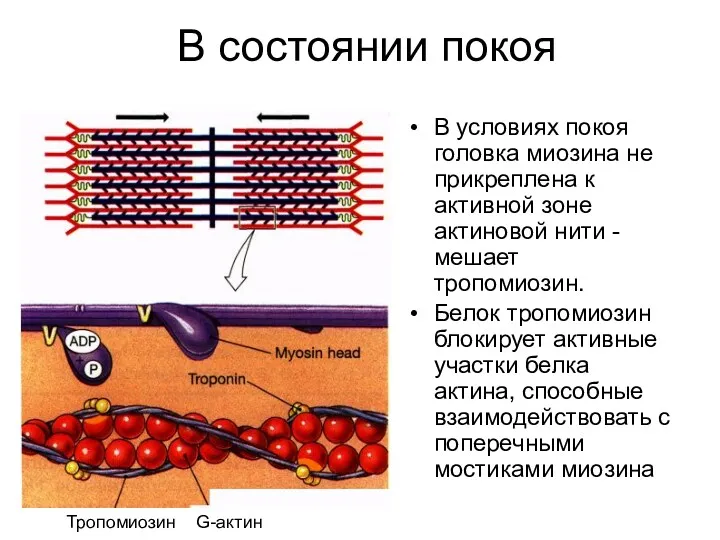
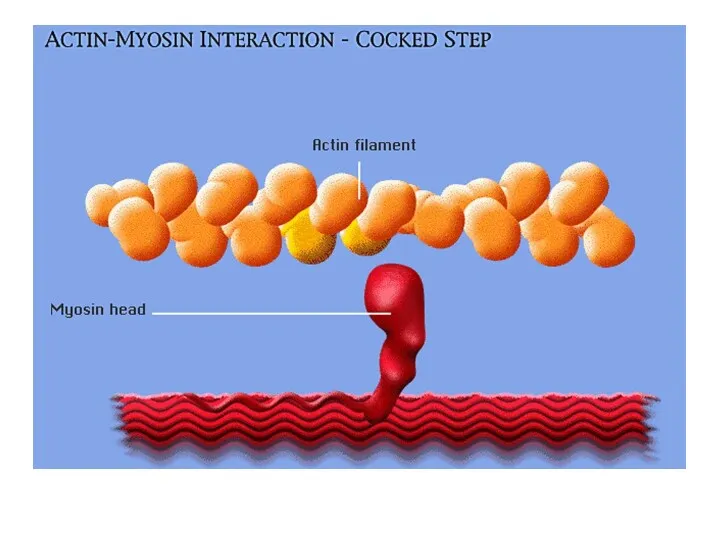
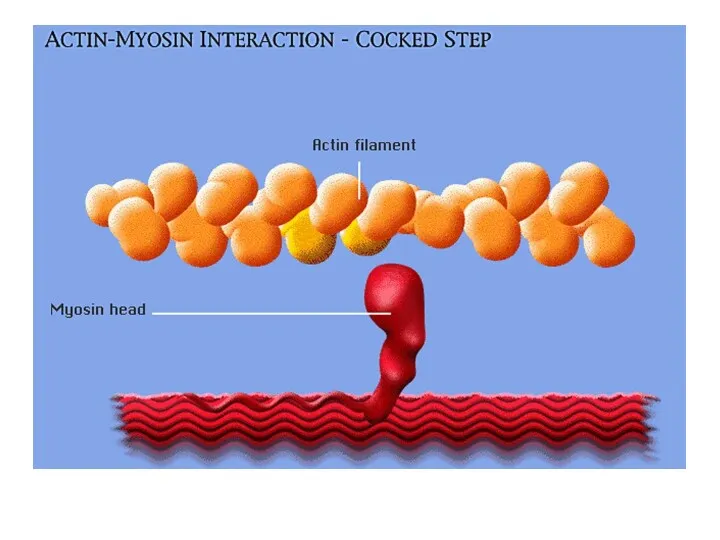
В состоянии покоя
В условиях покоя головка миозина не прикреплена к активной
В состоянии покоя
В условиях покоя головка миозина не прикреплена к активной
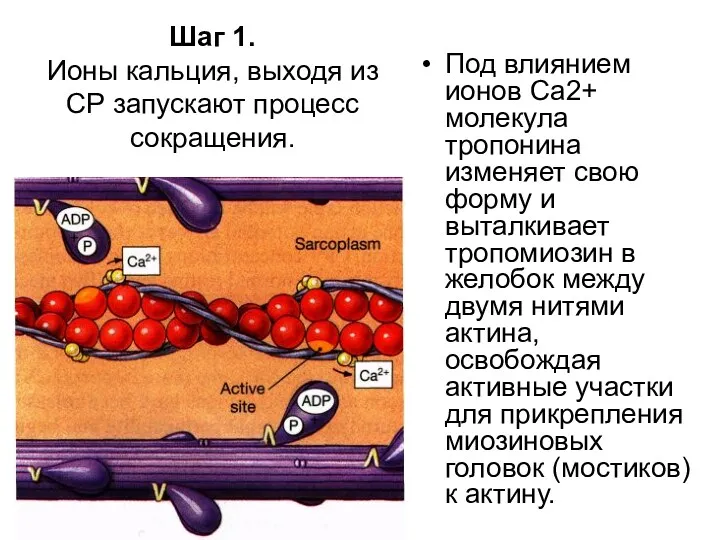
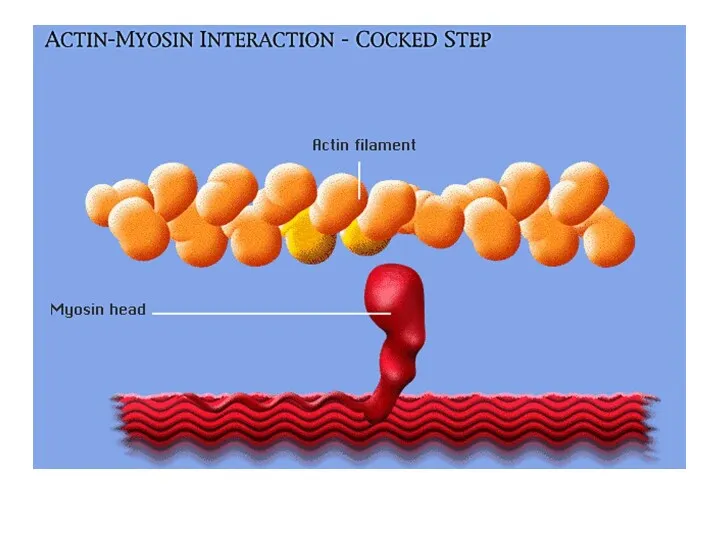
Шаг 1.
Ионы кальция, выходя из СР запускают процесс сокращения.
Под влиянием
Шаг 1.
Ионы кальция, выходя из СР запускают процесс сокращения.
Под влиянием
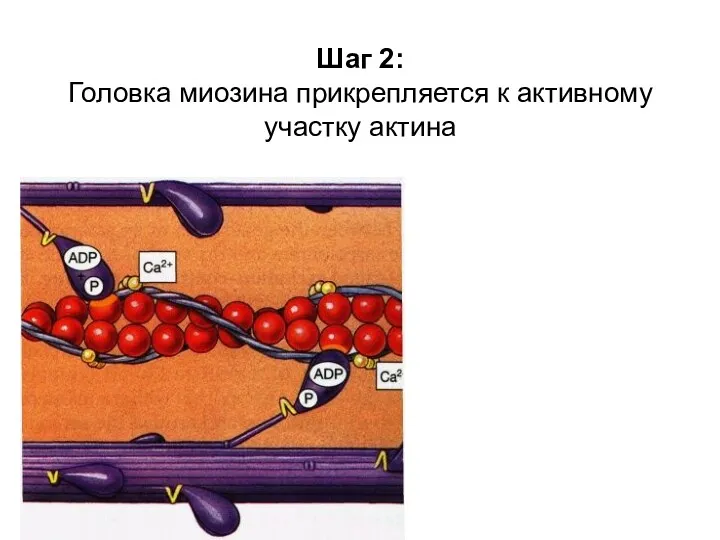
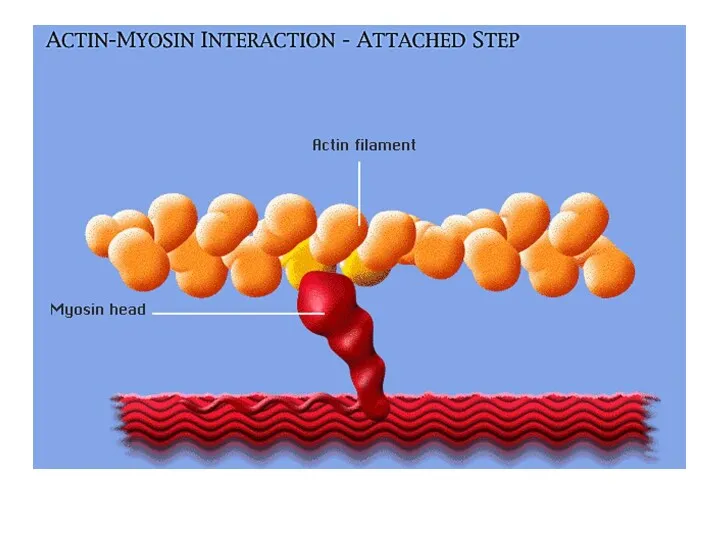
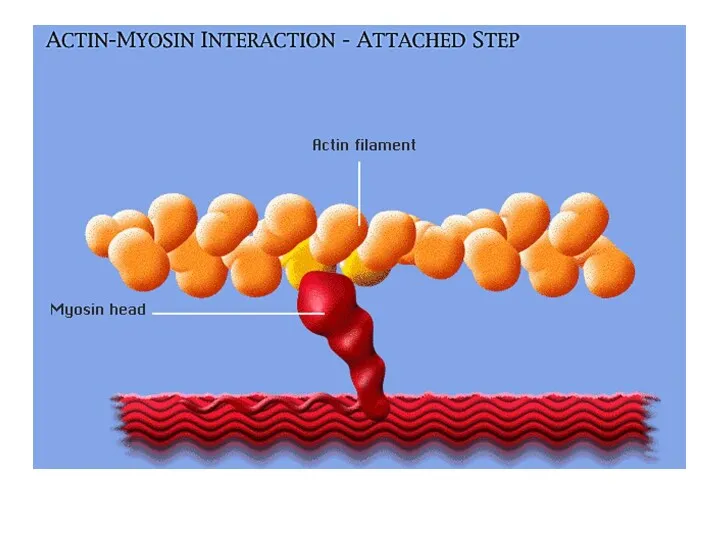
Шаг 2:
Головка миозина прикрепляется к активному участку актина
Шаг 2:
Головка миозина прикрепляется к активному участку актина
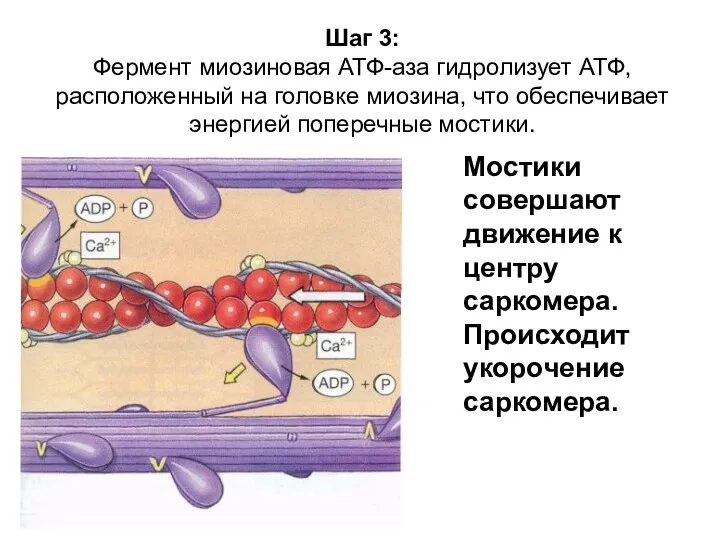
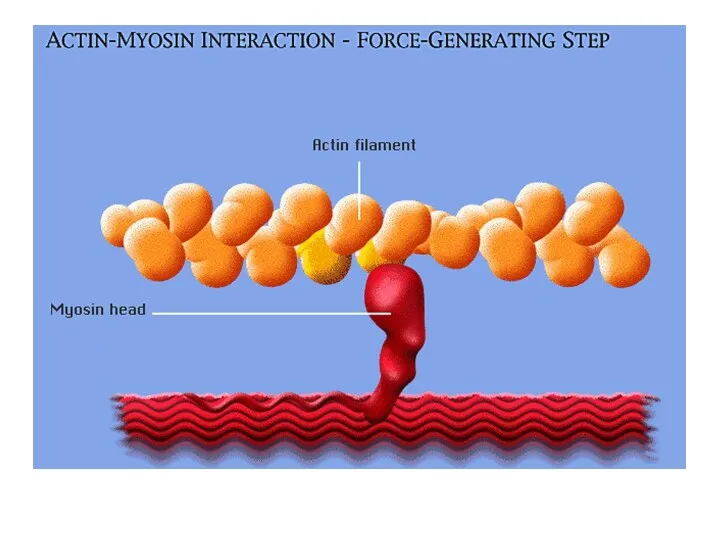
Шаг 3:
Фермент миозиновая АТФ-аза гидролизует АТФ, расположенный на головке миозина, что
Шаг 3: Фермент миозиновая АТФ-аза гидролизует АТФ, расположенный на головке миозина, что
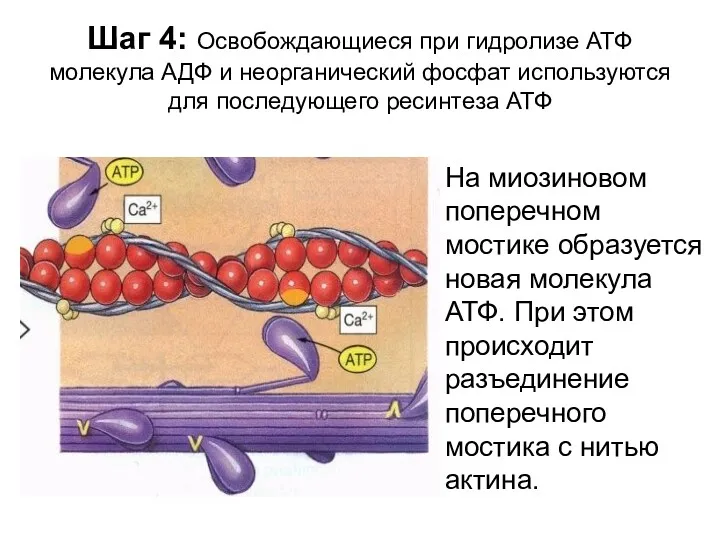
Шаг 4: Освобождающиеся при гидролизе АТФ молекула АДФ и неорганический фосфат
Шаг 4: Освобождающиеся при гидролизе АТФ молекула АДФ и неорганический фосфат
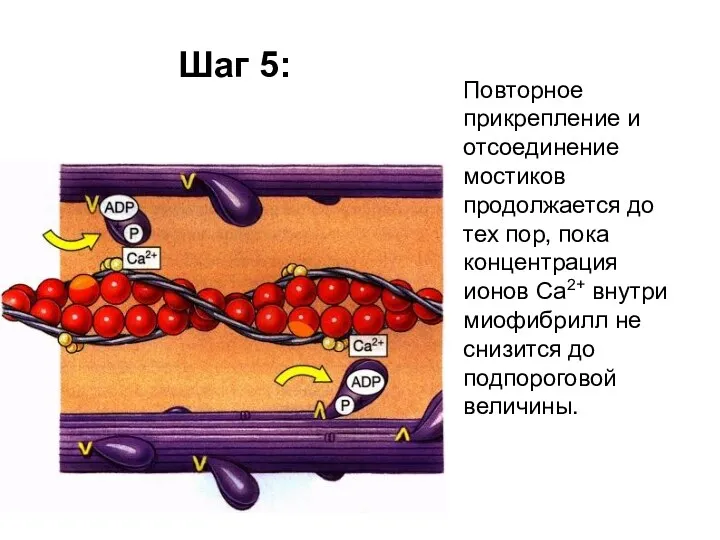
Шаг 5:
Повторное прикрепление и отсоединение мостиков продолжается до тех пор, пока
Шаг 5:
Повторное прикрепление и отсоединение мостиков продолжается до тех пор, пока
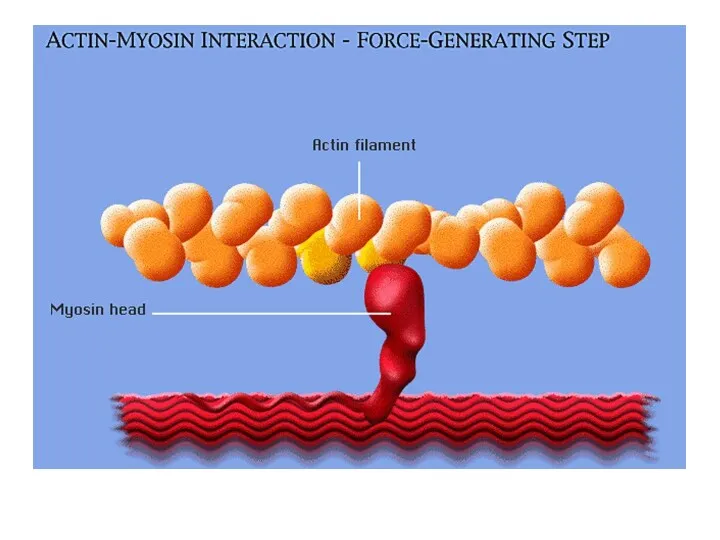
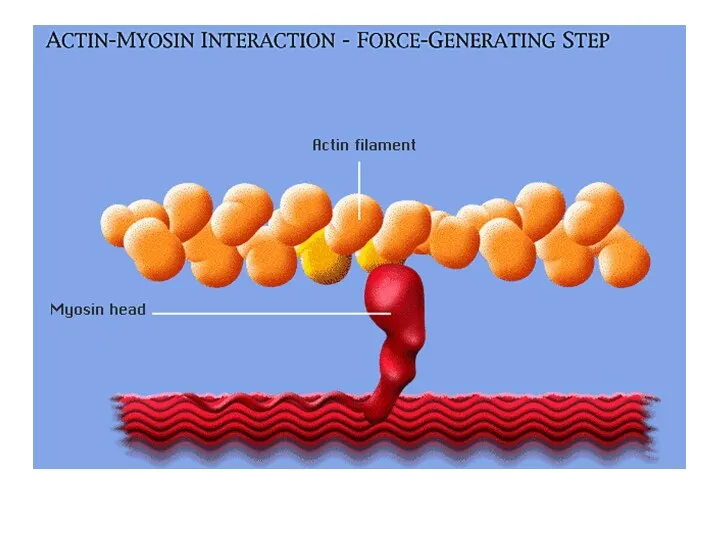

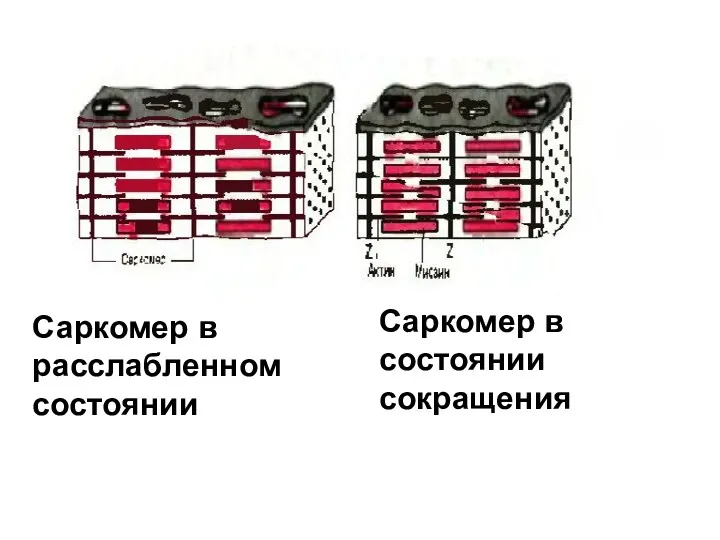
Саркомер в расслабленном
состоянии
Саркомер в состоянии
сокращения
Саркомер в расслабленном
состоянии
Саркомер в состоянии
сокращения

Роль кальция в мышечном сокращении
1 – Выброс медиатора в синаптическую щель.
2
Роль кальция в мышечном сокращении
1 – Выброс медиатора в синаптическую щель.
2
![Механизм расслабления мышцы [Са+2] 107 М + кальциевый насос в](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/214106/slide-104.jpg)
Механизм расслабления мышцы
[Са+2]
107 М
+ кальциевый насос в СПР
СПР
Са+2
Са+2
Са+2
АТФ
[Са+2]
>
<
105 М
Механизм расслабления мышцы
[Са+2]
107 М
+ кальциевый насос в СПР
СПР
Са+2
Са+2
Са+2
АТФ
[Са+2]
>
<
105 М

Затраты энергии АТФ во время мышечного сокращения
На поддержание ионной асимметрии (натрий-калиевй
Затраты энергии АТФ во время мышечного сокращения
На поддержание ионной асимметрии (натрий-калиевй

Роль АТФ в мышечном сокращении
а) сокращения (образования мостиков);
б) расслабления (разрыва
Роль АТФ в мышечном сокращении
а) сокращения (образования мостиков);
б) расслабления (разрыва

Пути ресинтеза АТФ
1) креатинфосфокиназный (КФ):
АДФ + КФ <==> АТФ
Пути ресинтеза АТФ
1) креатинфосфокиназный (КФ):
АДФ + КФ <==> АТФ

Типы мышечного сокращения
1. Изотоническое — волокна мышцы укорачиваются без изменения
Типы мышечного сокращения
1. Изотоническое — волокна мышцы укорачиваются без изменения

Виды раздражения мышцы в эксперименте
Непрямое раздражение – импульс воздействует на нервное
Виды раздражения мышцы в эксперименте
Непрямое раздражение – импульс воздействует на нервное
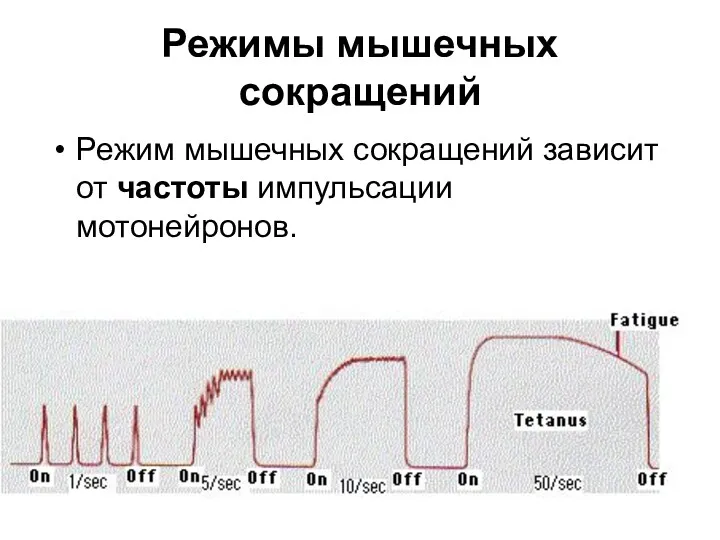
Режимы мышечных сокращений
Режим мышечных сокращений зависит от частоты импульсации мотонейронов.
Режимы мышечных сокращений
Режим мышечных сокращений зависит от частоты импульсации мотонейронов.

Одиночное сокращение -
- механический ответ мышечного волокна на однократное раздражение.
Одиночное сокращение -
- механический ответ мышечного волокна на однократное раздражение.
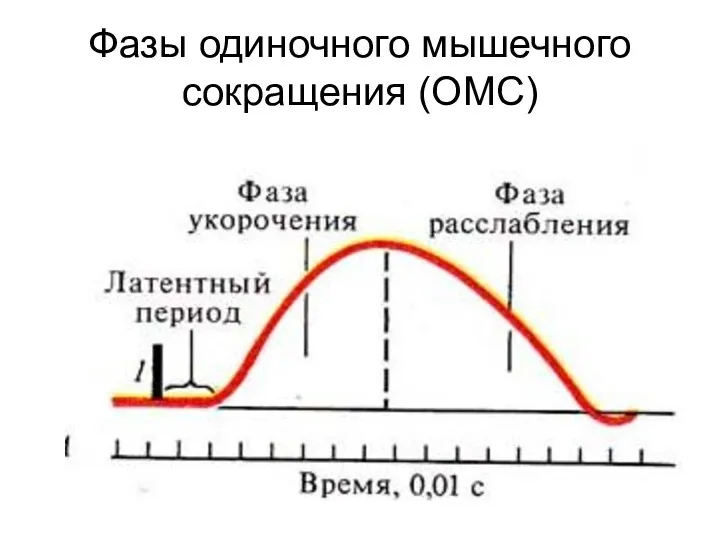
Фазы одиночного мышечного сокращения (ОМС)
Фазы одиночного мышечного сокращения (ОМС)
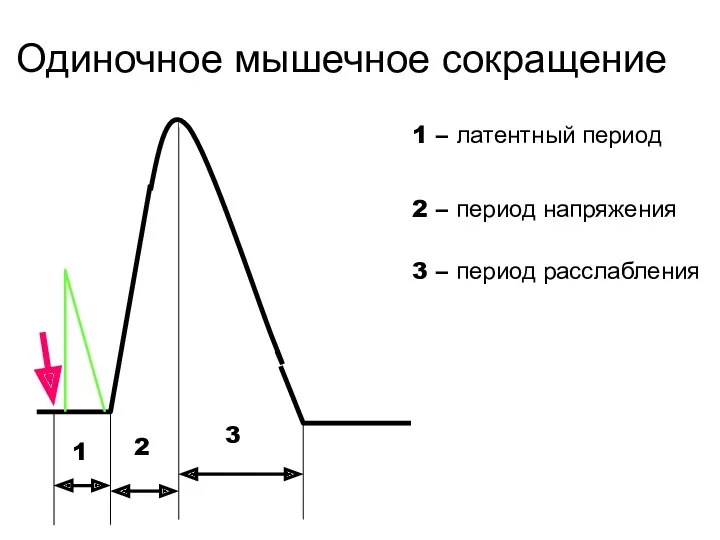
Одиночное мышечное сокращение
1
2
3
1 – латентный период
2 – период напряжения
3 – период
Одиночное мышечное сокращение
1
2
3
1 – латентный период
2 – период напряжения
3 – период
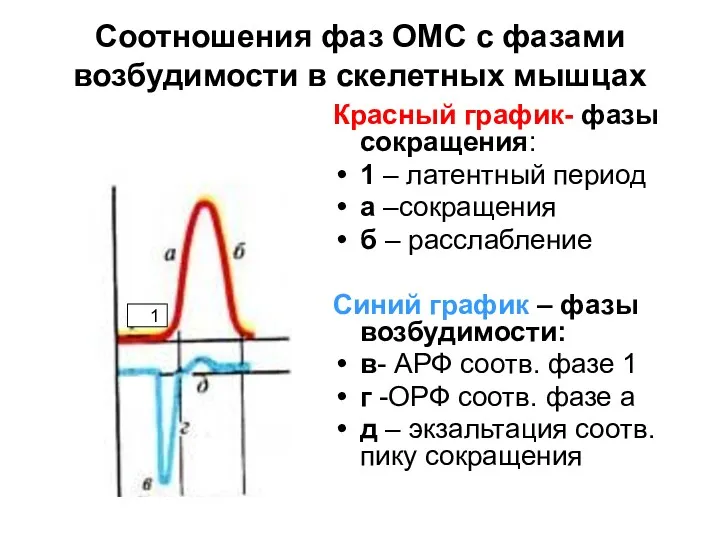
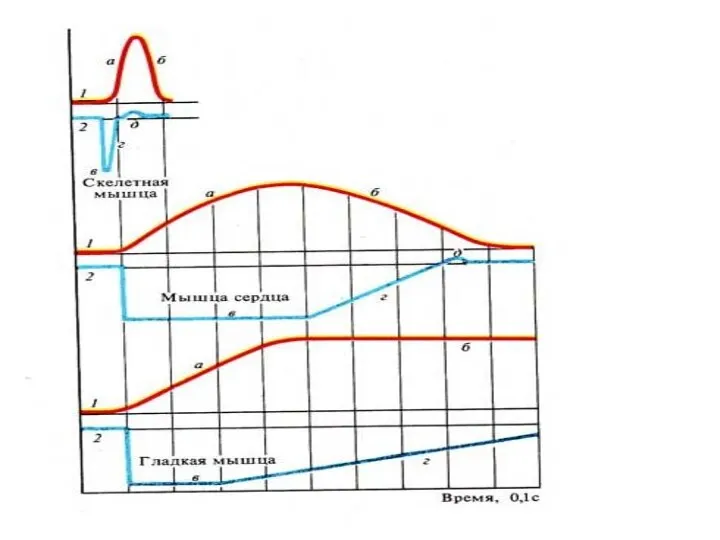
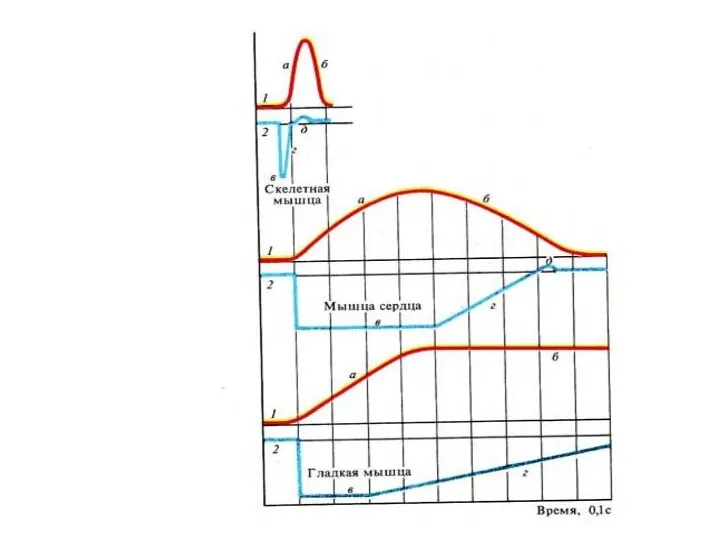
Соотношения фаз ОМС с фазами возбудимости в скелетных мышцах
Красный график- фазы
Соотношения фаз ОМС с фазами возбудимости в скелетных мышцах
Красный график- фазы
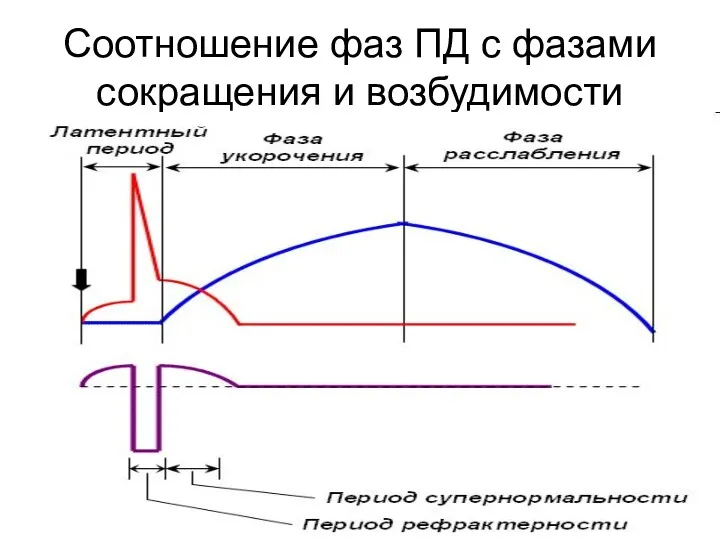
Соотношение фаз ПД с фазами сокращения и возбудимости
Соотношение фаз ПД с фазами сокращения и возбудимости

Тетанус
В естественных условиях к мышечным волокнам поступают не одиночные нервные импульсы,
Тетанус
В естественных условиях к мышечным волокнам поступают не одиночные нервные импульсы,
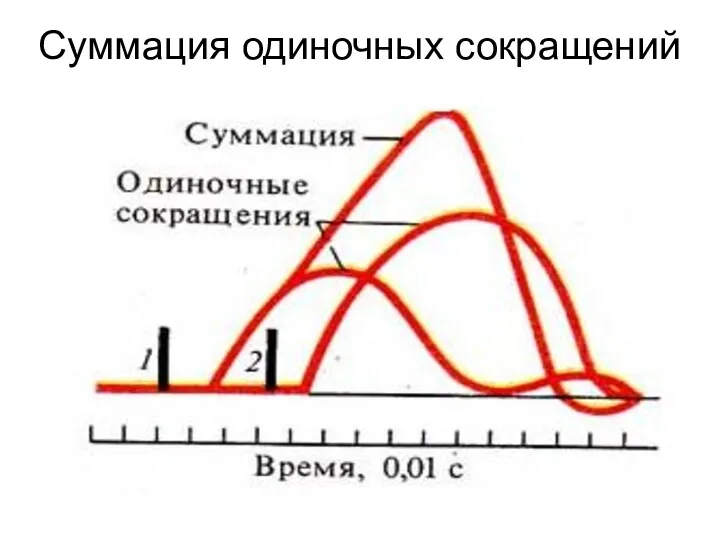
Суммация одиночных сокращений
Суммация одиночных сокращений

Тетанус возникает вследствие суммации одиночных мышечных сокращений.
Для того, чтобы возник
Тетанус возникает вследствие суммации одиночных мышечных сокращений.
Для того, чтобы возник
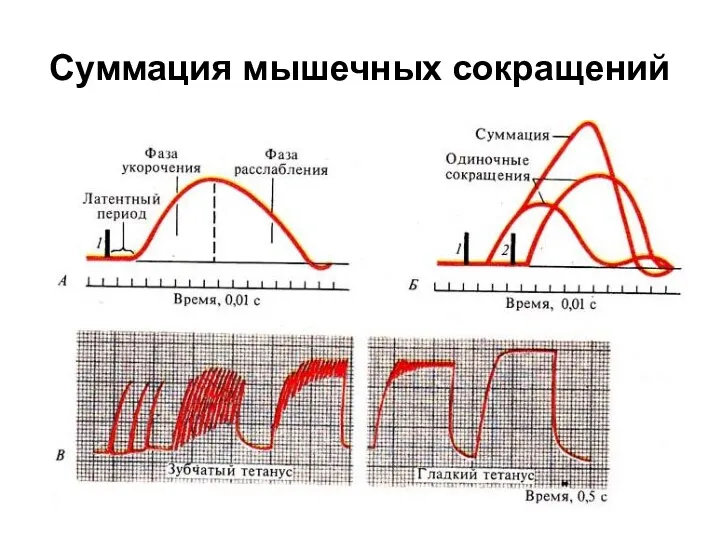
Суммация мышечных сокращений
Суммация мышечных сокращений
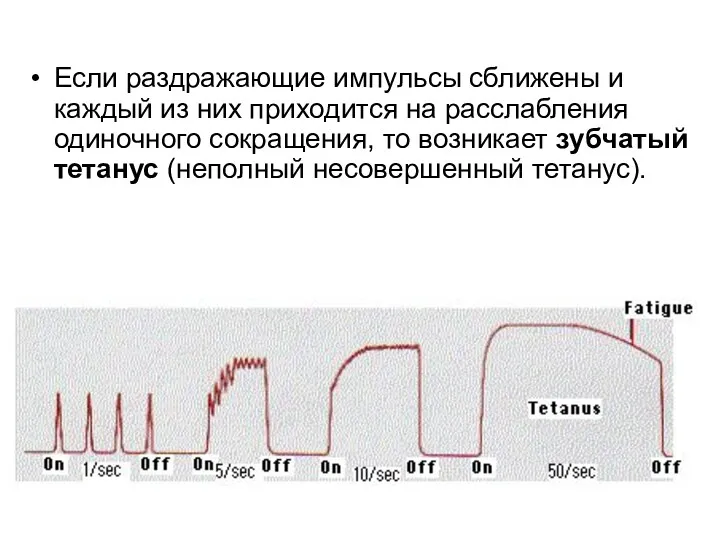
Если раздражающие импульсы сближены и каждый из них приходится на расслабления
Если раздражающие импульсы сближены и каждый из них приходится на расслабления
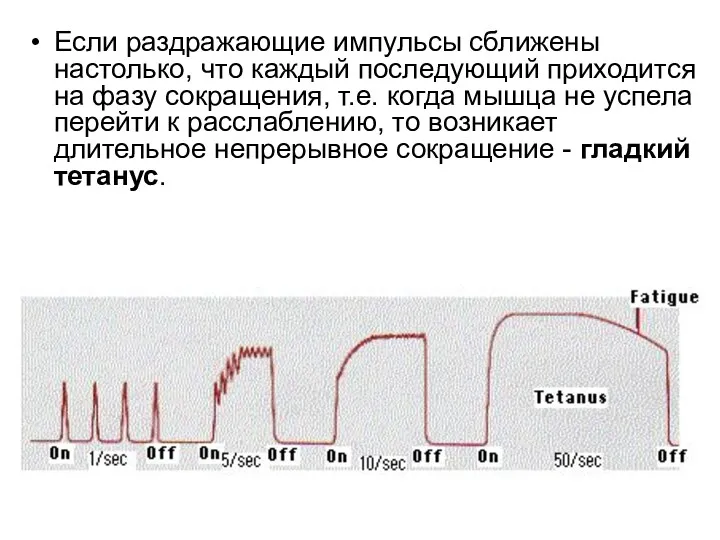
Если раздражающие импульсы сближены настолько, что каждый последующий приходится на фазу
Если раздражающие импульсы сближены настолько, что каждый последующий приходится на фазу
При анализе графиков, обращает на себя внимание следующая особенность: амплитуда зубчатого
При анализе графиков, обращает на себя внимание следующая особенность: амплитуда зубчатого
Оптимум - частота раздражения, при которой наблюдается суммарное сокращение (тетанус) наибольшей
Оптимум - частота раздражения, при которой наблюдается суммарное сокращение (тетанус) наибольшей
1
2
3
4
5
Е0
Екр
0
+30
В
Г
А
Б
1
2
3
4
5
Е0
Екр
0
+30
В
Г
А
Б
Зубчатый тетанус возникает при частоте нервных импульсов до 30 в 1с.
Зубчатый тетанус возникает при частоте нервных импульсов до 30 в 1с.
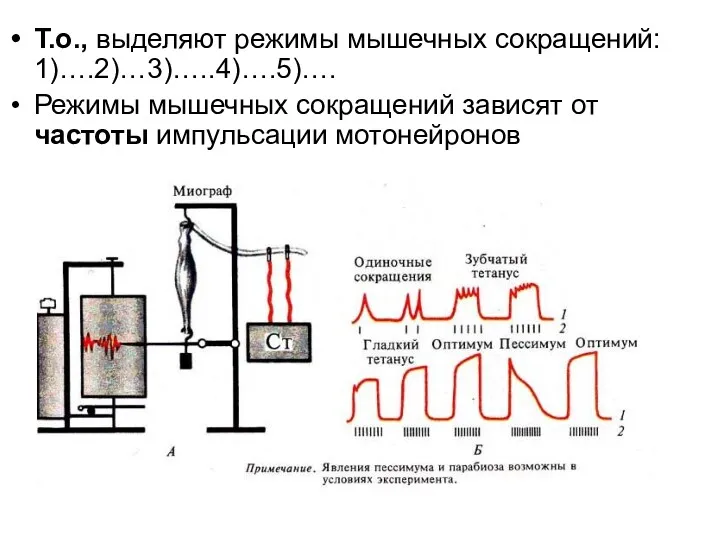
Т.о., выделяют режимы мышечных сокращений: 1)….2)…3)…..4)….5)….
Режимы мышечных сокращений зависят от частоты
Т.о., выделяют режимы мышечных сокращений: 1)….2)…3)…..4)….5)….
Режимы мышечных сокращений зависят от частоты
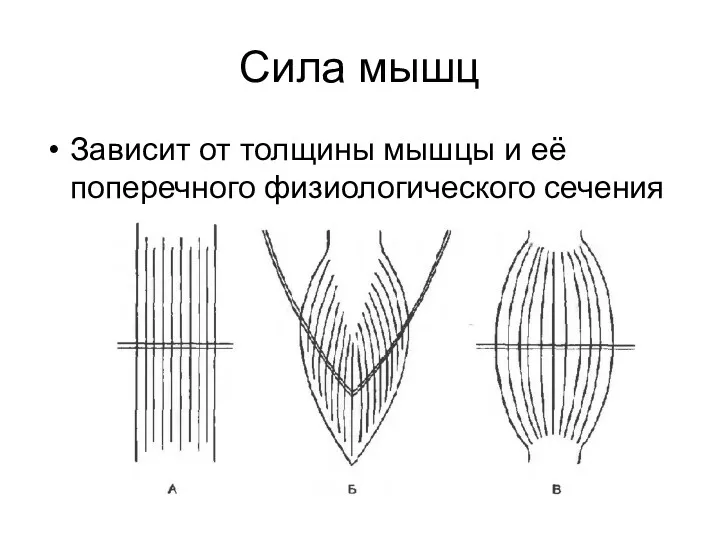
Сила мышц
Зависит от толщины мышцы и её поперечного физиологического сечения
Сила мышц
Зависит от толщины мышцы и её поперечного физиологического сечения

Работа мышцы
Это энергия, затрачиваемая на перемещение тела с определенной силой на
Работа мышцы
Это энергия, затрачиваемая на перемещение тела с определенной силой на

утомление
Процесс временного снижения работоспособности мышцы.
Возникает в связи с уменьшением энергетических запасов
утомление
Процесс временного снижения работоспособности мышцы.
Возникает в связи с уменьшением энергетических запасов
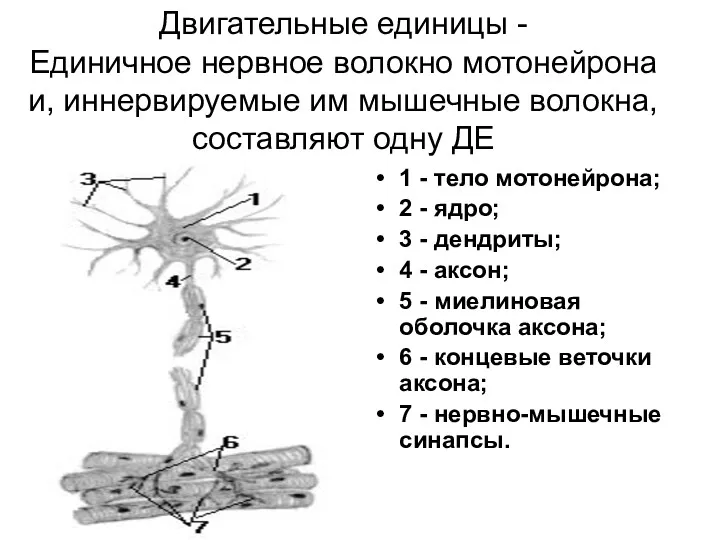
Двигательные единицы -
Единичное нервное волокно мотонейрона и, иннервируемые им мышечные волокна,
Двигательные единицы - Единичное нервное волокно мотонейрона и, иннервируемые им мышечные волокна,
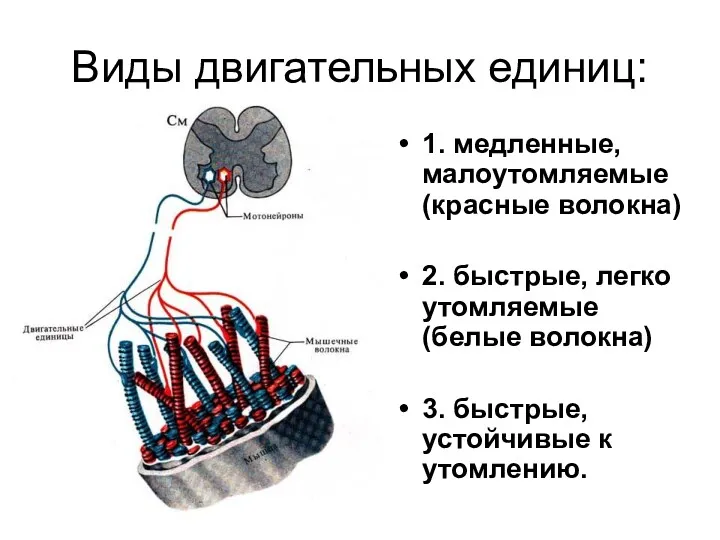
Виды двигательных единиц:
1. медленные, малоутомляемые (красные волокна)
2. быстрые, легко утомляемые (белые
Виды двигательных единиц:
1. медленные, малоутомляемые (красные волокна)
2. быстрые, легко утомляемые (белые

Функциональные особенности медленных ДЕ
1. Иннервируются высоко возбудимыми а-мотонейронами с низкой скоростью
Функциональные особенности медленных ДЕ
1. Иннервируются высоко возбудимыми а-мотонейронами с низкой скоростью

Функциональные особенности медленных ДЕ
4. Имеют хорошее кровоснабжение, много митохондрий, миоглобина, высокий
Функциональные особенности медленных ДЕ
4. Имеют хорошее кровоснабжение, много митохондрий, миоглобина, высокий

Функциональные особенности быстрых ДЕ.
1. Иннервируются крупными, менее возбудимыми а-мотонейронами с
Функциональные особенности быстрых ДЕ.
1. Иннервируются крупными, менее возбудимыми а-мотонейронами с

Функциональные особенности быстрых ДЕ.
4. Имеют слаборазвитую капиллярную сеть, мало митохондрий,
Функциональные особенности быстрых ДЕ.
4. Имеют слаборазвитую капиллярную сеть, мало митохондрий,

Функциональные особенности быстрых, устойчивых к утомлению ДЕ.
По структурно-функциональным свойствам занимают
Функциональные особенности быстрых, устойчивых к утомлению ДЕ.
По структурно-функциональным свойствам занимают
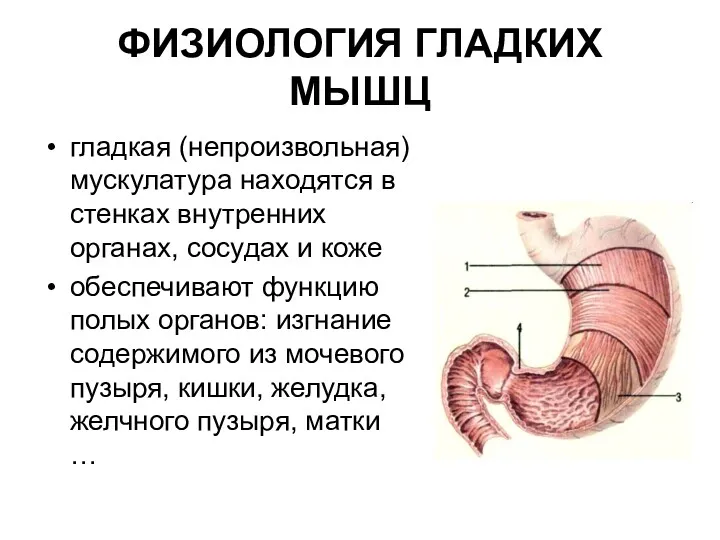
ФИЗИОЛОГИЯ ГЛАДКИХ МЫШЦ
гладкая (непроизвольная) мускулатура находятся в стенках внутренних органах, сосудах
ФИЗИОЛОГИЯ ГЛАДКИХ МЫШЦ
гладкая (непроизвольная) мускулатура находятся в стенках внутренних органах, сосудах

Структурные особенности ГМК
миофиламенты содержат актин и миозин, которые распределены менее упорядочено,
Структурные особенности ГМК
миофиламенты содержат актин и миозин, которые распределены менее упорядочено,

Физиологические особенности ГМК
Потенциал покоя -30 до -70 мВ
Потенциал действия двух
Физиологические особенности ГМК
Потенциал покоя -30 до -70 мВ
Потенциал действия двух

Свойства гладких мышц
сила сокращения зависит от степени растяжения мышцы (прямо-пропорционально)
пластичность
Свойства гладких мышц
сила сокращения зависит от степени растяжения мышцы (прямо-пропорционально)
пластичность
 Удобрения, их свойства и применение
Удобрения, их свойства и применение Понятие Сбалансированное питание
Понятие Сбалансированное питание Фізіологія занього, середнього та проміжного мозку
Фізіологія занього, середнього та проміжного мозку Соединительные ткани
Соединительные ткани Сон и сновидения
Сон и сновидения Russian desman
Russian desman Изменчивость: наследственная и ненаследственная
Изменчивость: наследственная и ненаследственная Пентозо-фосфатный путь окисления глюкозы
Пентозо-фосфатный путь окисления глюкозы Кровеносная система. Внутренняя среда организма. Кровь
Кровеносная система. Внутренняя среда организма. Кровь Презентация по биологии Своя игра (обобщение по всему курсу) 8 класс
Презентация по биологии Своя игра (обобщение по всему курсу) 8 класс Микробиология зерновых продуктов
Микробиология зерновых продуктов Строение и работа сердца
Строение и работа сердца Итоговая контрольная работа. 6 класс
Итоговая контрольная работа. 6 класс Цели физического воспитания
Цели физического воспитания Способы культивирования вирусов
Способы культивирования вирусов Термодинамика биологических процессов(new)
Термодинамика биологических процессов(new) Химический состав клетки. Нуклеиновые кислоты. ДНК
Химический состав клетки. Нуклеиновые кислоты. ДНК Мутационная изменчивость
Мутационная изменчивость Гормоны поджелудочной железы
Гормоны поджелудочной железы Отряд Перепончатокрылые
Отряд Перепончатокрылые Урок на тему :Совместная жизнь видов в биогеоценозе.
Урок на тему :Совместная жизнь видов в биогеоценозе. Неорганические вещества клетки
Неорганические вещества клетки Презентация по теме Кайнозойская эра, 11 класс
Презентация по теме Кайнозойская эра, 11 класс Водоросли и их классификация
Водоросли и их классификация Строение и функции кожи
Строение и функции кожи Генетика. Наследственность. Изменчивость
Генетика. Наследственность. Изменчивость Энзимология. Структура и механизм действия ферментов. (Лекция 2)
Энзимология. Структура и механизм действия ферментов. (Лекция 2) Подготовка к ЕГЭ по биологии. 10 класс
Подготовка к ЕГЭ по биологии. 10 класс