- Митохондриальный протеом. Структура мтДНК. Лекция 2

Содержание
- 3. Часть белков эндосимбионта до сих пор функционирует в митохондриях (ферменты окислительного фосфорилирования, компоненты рибосом) Часть белков
- 4. Потеря белков митохондрией в ходе эволюции в основном происходила на ранних этапах: Из 370 белков бактериального
- 5. Потеря белков митохондрией: Часть белков потеряна полностью в связи с потерей функции (ферменты, ответственные за синтез
- 6. Потерянные бактериальные белки часто заменялись другими неортологичными белками Например, DNA polymerase γ, POLRMT и TWINKLE имеют
- 7. Одна из гипотез: перенос происходит из-за высокой частоты мутаций в связи с близостью дыхательной цепи. Ортологичные
- 8. Все белки, необходимые для митохондриальной репликации, транскрипции и трансляции, расположены в ядерном геноме.
- 9. Для репликации и экспрессии мт генома, в котором сохранились всего несколько генов, необходима сложная ферментативная система.
- 10. В ходе эволюции происходило увеличение числа субъединиц в больших мультиферментных митохондриальных комплексах Это может компенсировать мутации
- 11. Митохондрии произошли от α-протеобактерий Часть белков эндосимбионта до сих пор функционирует в митохондриях Большая часть предковых
- 12. МтДНК организована в ДНК-белковые комплексы – нуклеоиды. Нуклеоид содержит 1-10 копий мтДНК и белки, участвующие в
- 13. PMID:24076128
- 14. TFAM (mitochondrial transcription factor A) участвует в пространственной организации нуклеоида: Имеет гомологию с ДНК-связывающими белками HMG
- 15. TFAM (mitochondrial transcription factor A) участвует в пространственной организации нуклеоида. PMID: 22465614
- 16. Open circle Supercoiled circle Эти две формы составляют большинство у многоклеточных животных Head-to-tail circular dimer В
- 17. Catenane Их количество значительно варьирует: 10% у мыши, более 30% в клетках человека линии НЕК. У
- 18. Так же организована мтДНК у некоторых растений, малярийного плазмодия и некоторых грибов: линейные геномы, соединенные Head-to-tail
- 19. МтДНК Млекопитающих организована в ДНК-белковые комплексы – нуклеоиды В одной клетке около 1000 нуклеоидов TFAM (mitochondrial
- 20. Генетический код в мтДНК несколько отличается от универсального
- 21. Для каких тРНК в мт геноме по 2 гена? PMID:22369973 PMID:24822055
- 22. D-arm-lacking tRNA (bovine mt tRNASer(GCU) T-arm-lacking tRNA (Ascaris suum mt tRNAMet PMID:24822055
- 23. Некоторые кодоны редки или отсутствуют в мтДНК: Из 111 геномов: в не менее, чем 76 нет
- 24. Мт ДНК кольцевая двуцепочечная молекула. Её цепи называются L (light) и H (heavy) из-за разницы в
- 25. Митохондриальный геном человека содержит 37 генов: На L-цепи – 8 тРНК + 1 мРНК На Н-цепи
- 26. 1.Мт ДНК кольцевая двуцепочечная молекула. Её цепи сильно отличаются по нуклеотидному составу и называются L (light)
- 27. Регуляторные участки в мтДНК: D-loop
- 28. D-loop впервые обнаружен на ЭМ мтДНК мыши и цыпленка более 40 лет назад (Arnberg et al,
- 29. В мт ДНК есть протяженный некодирующий участок NCR (non-coding region), расположенный между генами tRNA Pro и
- 30. PMID:24709344 D-loop содержится не во всех молекулах ДНК. Его содержание колеблется в широких пределах 1-95%. Есть
- 31. NCR содержит регуляторные элементы: Ориджин репликации OH и дополнительные ориджины (ori b) Промоторы для обеих цепей
- 32. Образование D-loop: репликация с участием ДНК-полимеразы ɣ (PolgA+PolgB), TFAM, mtSSB иTwinkle Деградация D-loop: нуклеаза MGME1 C
- 33. Предположительные функции D-loop: D-loop - преждевременно терминированная Н-цепь, образованная при репликации (модель Strand displacement) D-loop необходима
- 34. NCR содержит регуляторные элементы: ориджин репликации OH, промоторы LSP и HSP1 участки регуляции репликации CSB, участок
- 35. Генетика митохондрий МтДНК подвержена мутациям. Гомоплазмия – ни в одной молекуле мтДНК нет мутаций ИЛИ во
- 36. МтДНК реплицируется в течение всего клеточного цикла, независимо от репликации в ядре, поэтому мутации в мтДНК
- 37. Пациенты с гетероплазмией часто имеют разный уровень содержания мутантной ДНК в разных органах и даже в
- 38. При гомоплазмии все потомки больной матери будут также больными. 90% наследственной оптической нейропатии Лебера Мт ДНК
- 39. Мать с гетероплазмией может передать потомству разный уровень мутантной мтДНК, а может вообще не передать мутацию.
- 41. Скачать презентацию
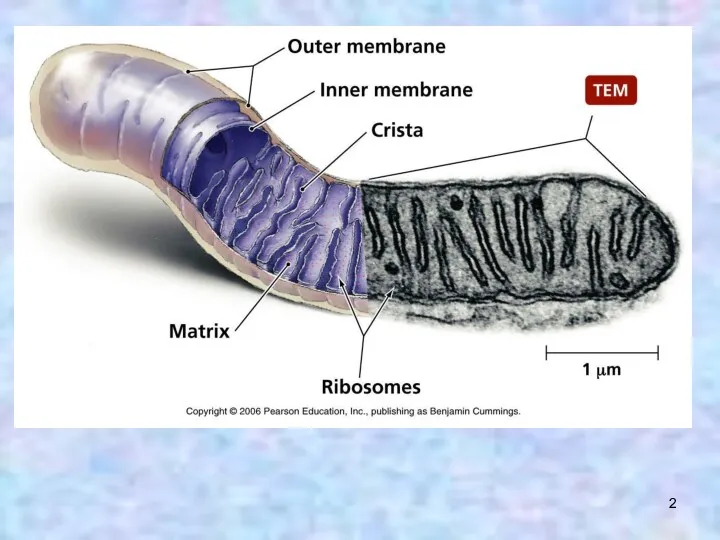
Часть белков эндосимбионта до сих пор функционирует в митохондриях (ферменты окислительного
Часть белков эндосимбионта до сих пор функционирует в митохондриях (ферменты окислительного
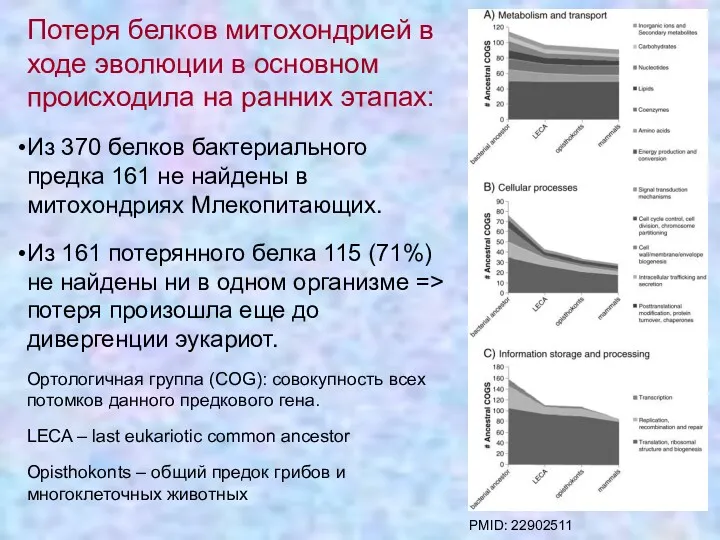
Потеря белков митохондрией в ходе эволюции в основном происходила на ранних
Потеря белков митохондрией в ходе эволюции в основном происходила на ранних

Потеря белков митохондрией:
Часть белков потеряна полностью в связи с потерей функции
Потеря белков митохондрией:
Часть белков потеряна полностью в связи с потерей функции
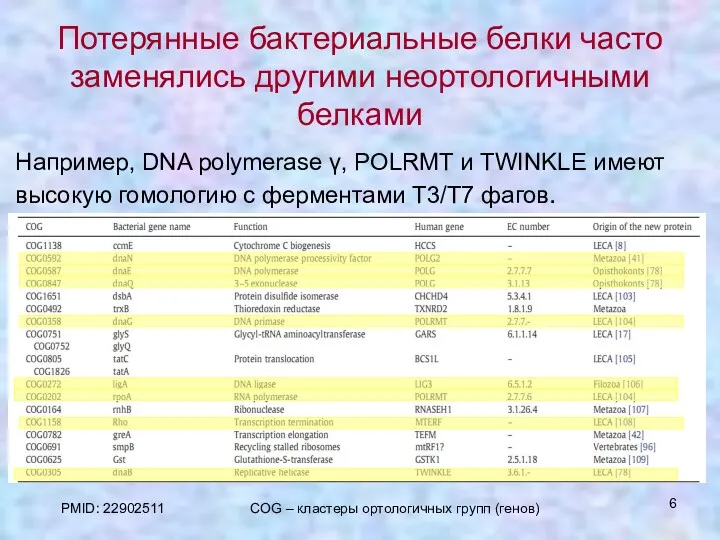
Потерянные бактериальные белки часто заменялись другими неортологичными белками
Например, DNA polymerase
Потерянные бактериальные белки часто заменялись другими неортологичными белками
Например, DNA polymerase
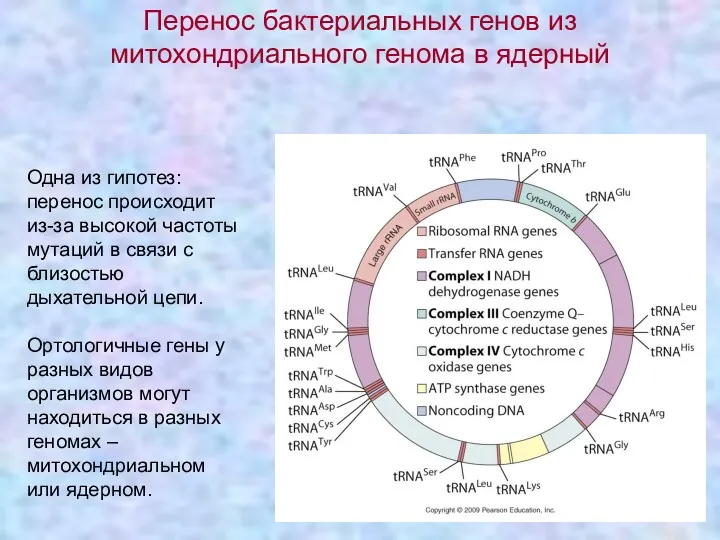
Одна из гипотез: перенос происходит из-за высокой частоты мутаций в связи
Одна из гипотез: перенос происходит из-за высокой частоты мутаций в связи
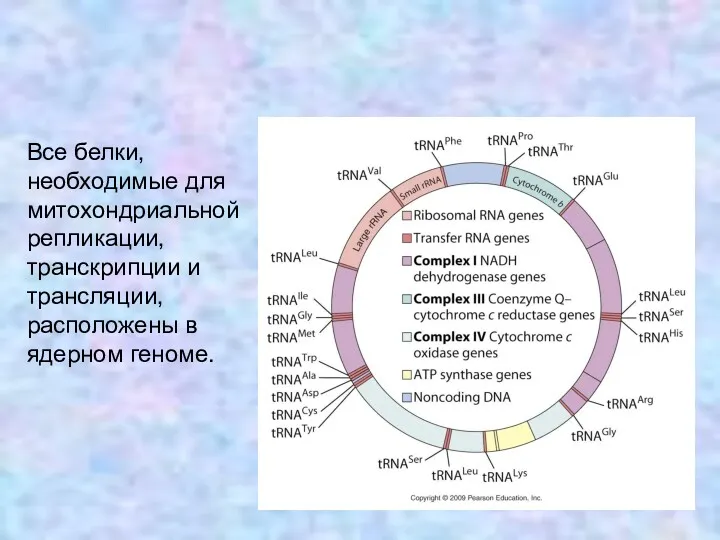
Все белки, необходимые для митохондриальной репликации, транскрипции и трансляции, расположены в
Все белки, необходимые для митохондриальной репликации, транскрипции и трансляции, расположены в

Для репликации и экспрессии мт генома, в котором сохранились всего несколько
Для репликации и экспрессии мт генома, в котором сохранились всего несколько
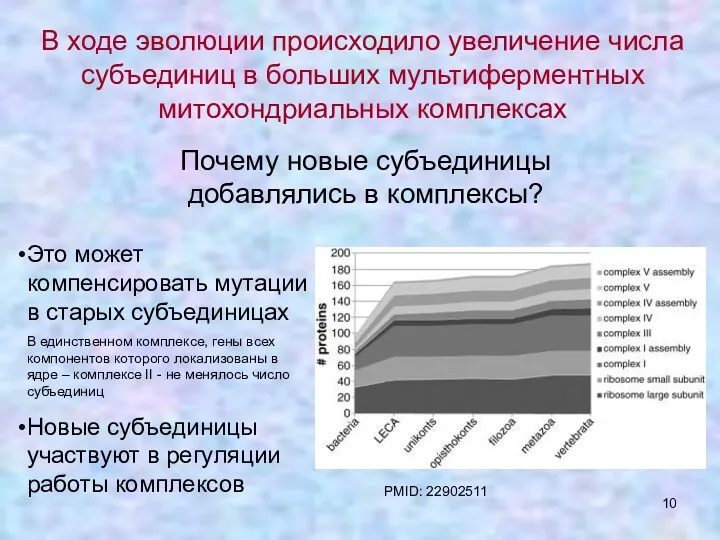
В ходе эволюции происходило увеличение числа субъединиц в больших мультиферментных митохондриальных
В ходе эволюции происходило увеличение числа субъединиц в больших мультиферментных митохондриальных

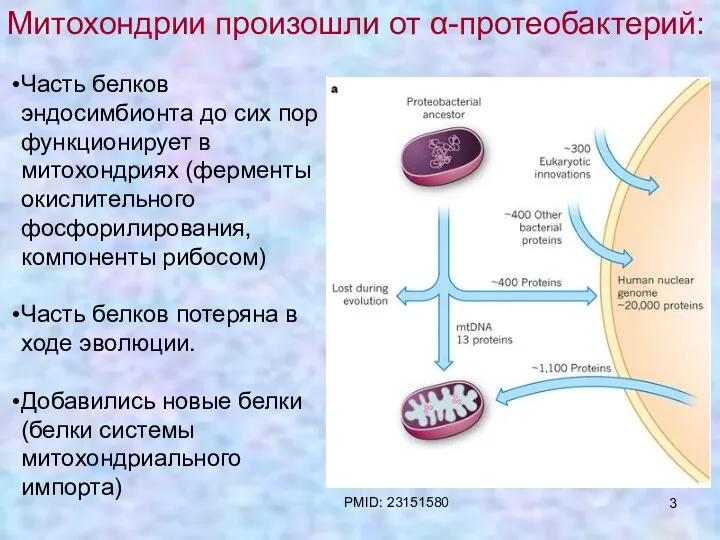
Митохондрии произошли от α-протеобактерий
Часть белков эндосимбионта до сих пор функционирует в
Митохондрии произошли от α-протеобактерий
Часть белков эндосимбионта до сих пор функционирует в
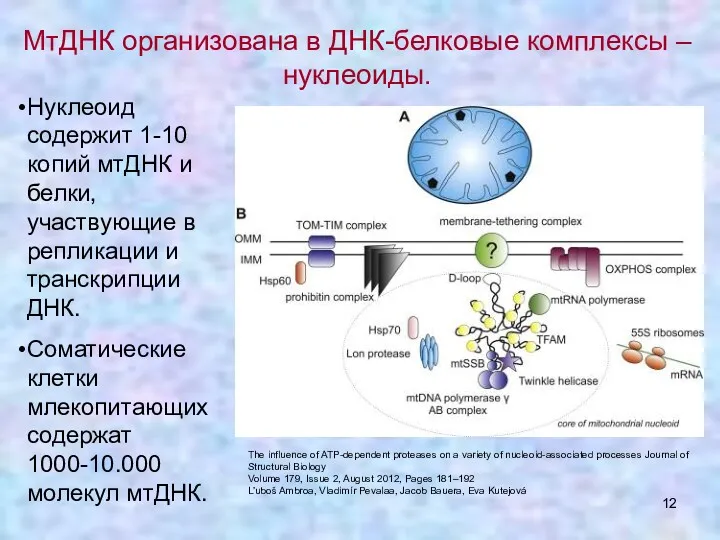
МтДНК организована в ДНК-белковые комплексы – нуклеоиды.
Нуклеоид содержит 1-10 копий
МтДНК организована в ДНК-белковые комплексы – нуклеоиды.
Нуклеоид содержит 1-10 копий
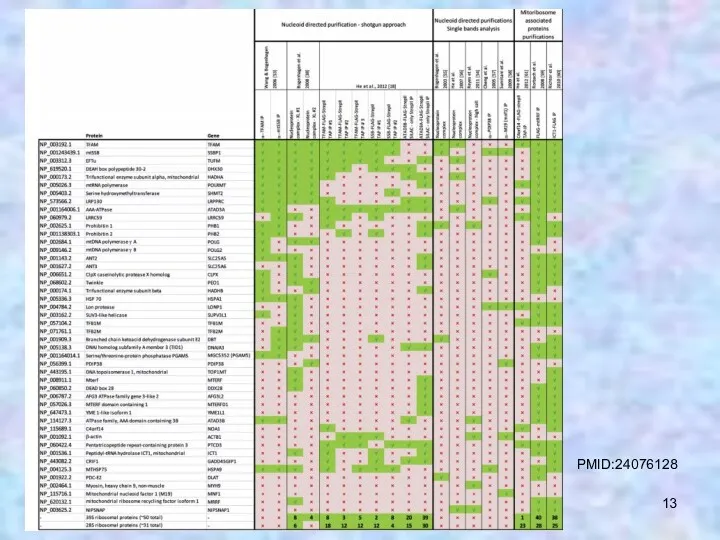
PMID:24076128
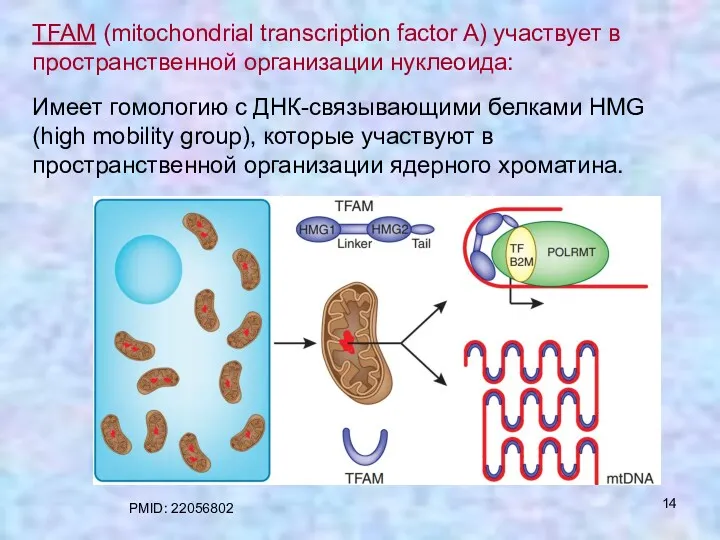
TFAM (mitochondrial transcription factor A) участвует в пространственной организации нуклеоида:
Имеет гомологию
TFAM (mitochondrial transcription factor A) участвует в пространственной организации нуклеоида:
Имеет гомологию
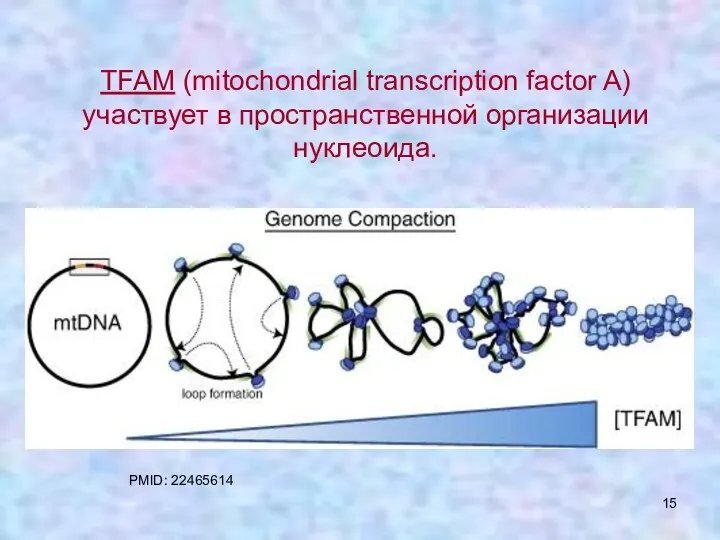
TFAM (mitochondrial transcription factor A) участвует в пространственной организации нуклеоида.
PMID: 22465614
TFAM (mitochondrial transcription factor A) участвует в пространственной организации нуклеоида.
PMID: 22465614
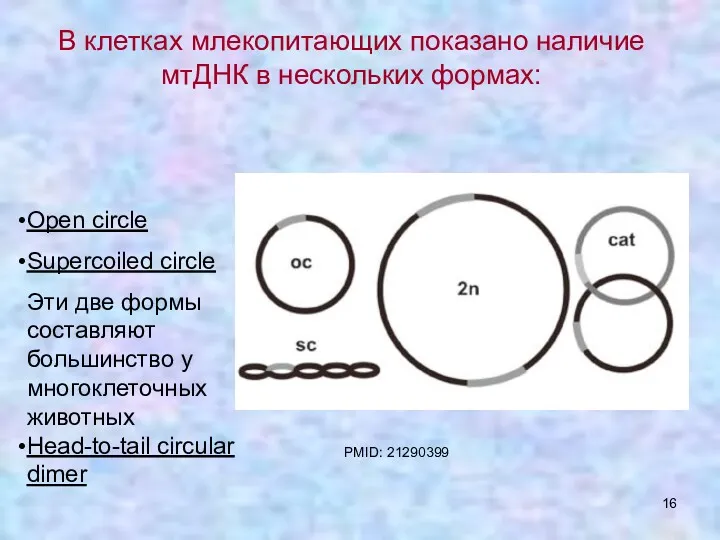
Open circle
Supercoiled circle
Эти две формы составляют большинство у многоклеточных животных
Head-to-tail
Open circle
Supercoiled circle
Эти две формы составляют большинство у многоклеточных животных
Head-to-tail

Catenane
Их количество значительно варьирует: 10% у мыши, более 30% в
Catenane
Их количество значительно варьирует: 10% у мыши, более 30% в

Так же организована мтДНК у некоторых растений, малярийного плазмодия и некоторых
Так же организована мтДНК у некоторых растений, малярийного плазмодия и некоторых

МтДНК Млекопитающих организована в ДНК-белковые комплексы – нуклеоиды
В одной клетке около
МтДНК Млекопитающих организована в ДНК-белковые комплексы – нуклеоиды
В одной клетке около
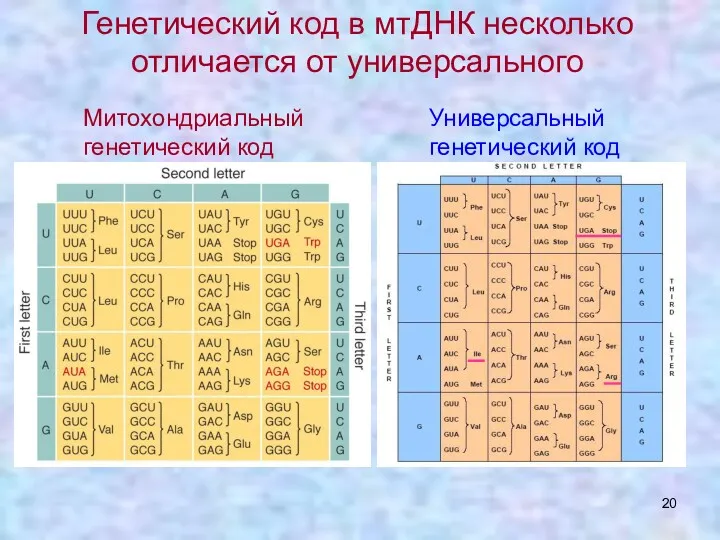
Генетический код в мтДНК несколько отличается от универсального
Генетический код в мтДНК несколько отличается от универсального
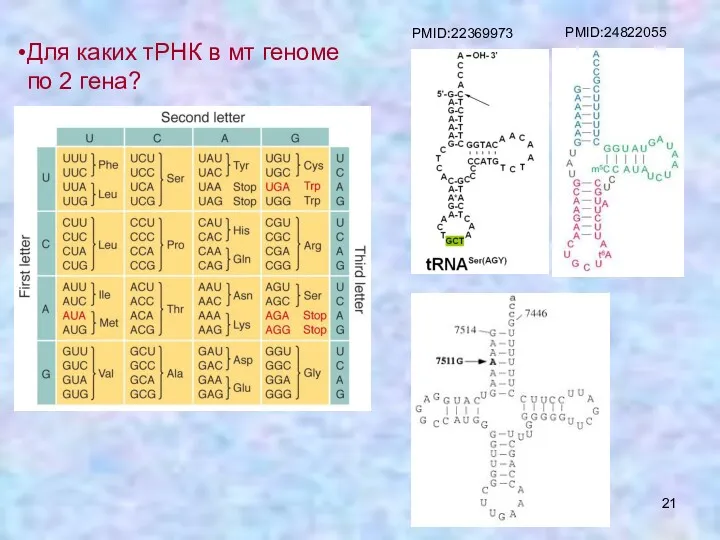
Для каких тРНК в мт геноме по 2 гена?
PMID:22369973
PMID:24822055
PMID:22369973
PMID:24822055
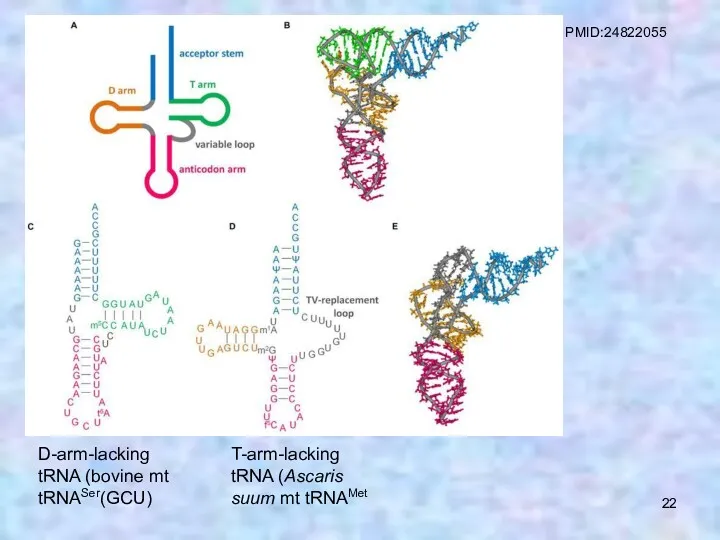
D-arm-lacking tRNA (bovine mt tRNASer(GCU)
T-arm-lacking tRNA (Ascaris suum mt tRNAMet
PMID:24822055
D-arm-lacking tRNA (bovine mt tRNASer(GCU)
T-arm-lacking tRNA (Ascaris suum mt tRNAMet
PMID:24822055

Некоторые кодоны редки или отсутствуют в мтДНК:
Из 111 геномов:
в не
Из 111 геномов:
в не
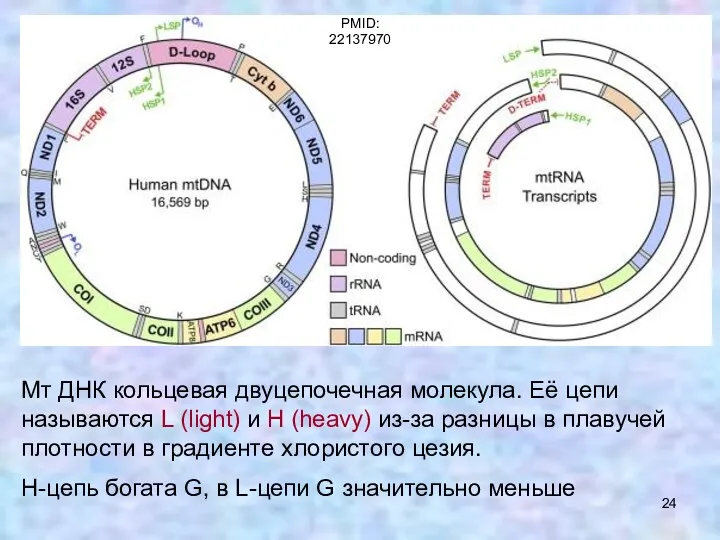
Мт ДНК кольцевая двуцепочечная молекула. Её цепи называются L (light) и
Мт ДНК кольцевая двуцепочечная молекула. Её цепи называются L (light) и
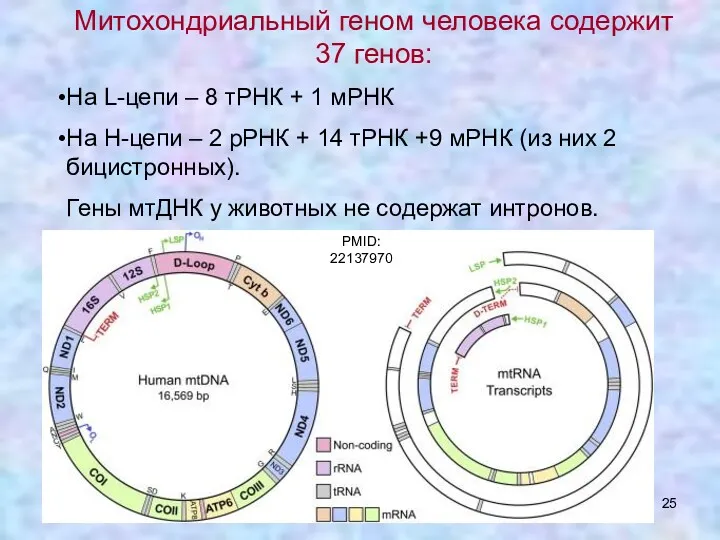
Митохондриальный геном человека содержит 37 генов:
На L-цепи – 8 тРНК +
Митохондриальный геном человека содержит 37 генов:
На L-цепи – 8 тРНК +

1.Мт ДНК кольцевая двуцепочечная молекула.
Её цепи сильно отличаются по нуклеотидному
1.Мт ДНК кольцевая двуцепочечная молекула.
Её цепи сильно отличаются по нуклеотидному
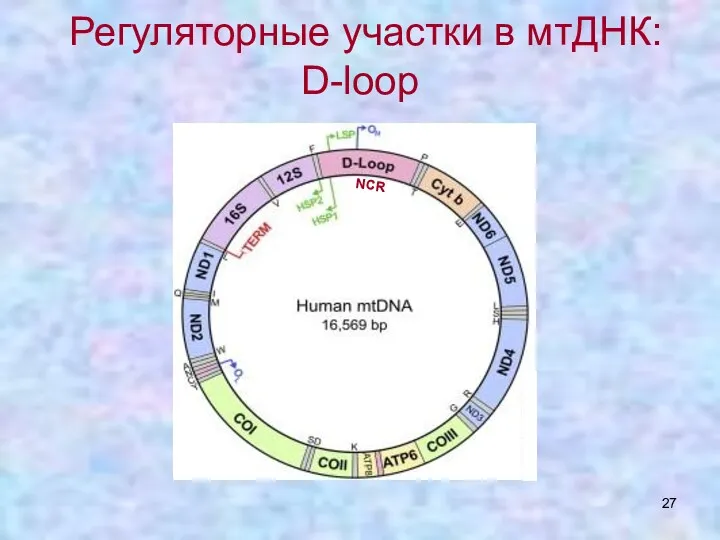
Регуляторные участки в мтДНК: D-loop
Регуляторные участки в мтДНК: D-loop
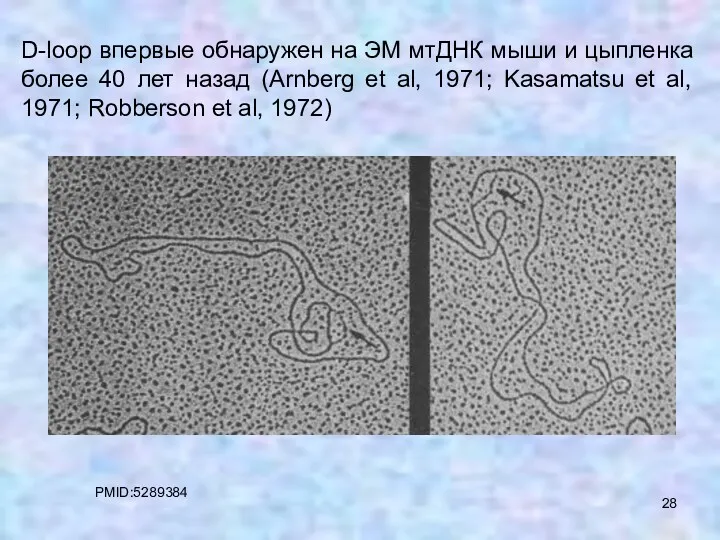
D-loop впервые обнаружен на ЭМ мтДНК мыши и цыпленка более 40
D-loop впервые обнаружен на ЭМ мтДНК мыши и цыпленка более 40
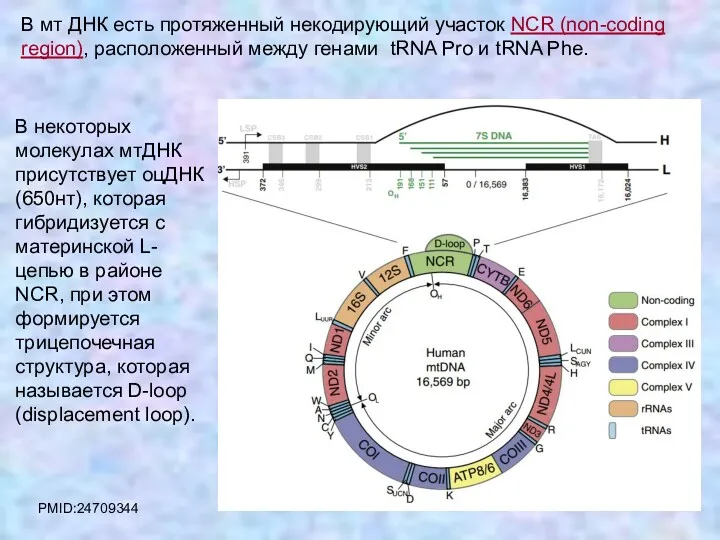
В мт ДНК есть протяженный некодирующий участок NCR (non-coding region), расположенный
В мт ДНК есть протяженный некодирующий участок NCR (non-coding region), расположенный
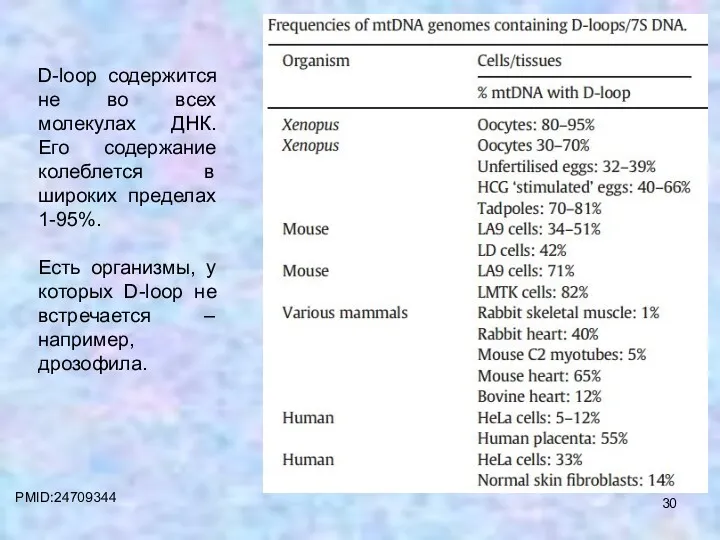
PMID:24709344
D-loop содержится не во всех молекулах ДНК. Его содержание колеблется в
PMID:24709344
D-loop содержится не во всех молекулах ДНК. Его содержание колеблется в
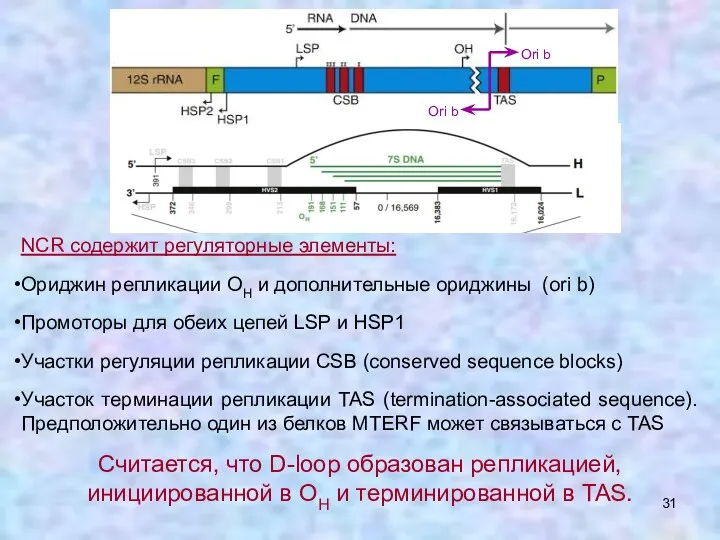
NCR содержит регуляторные элементы:
Ориджин репликации OH и дополнительные ориджины (ori b)
Промоторы
NCR содержит регуляторные элементы:
Ориджин репликации OH и дополнительные ориджины (ori b)
Промоторы
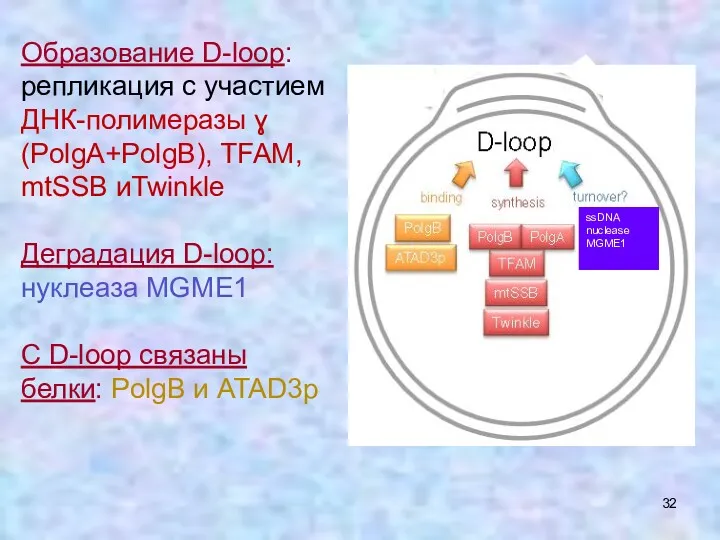
Образование D-loop: репликация с участием ДНК-полимеразы ɣ (PolgA+PolgB), TFAM, mtSSB иTwinkle
Деградация
Образование D-loop: репликация с участием ДНК-полимеразы ɣ (PolgA+PolgB), TFAM, mtSSB иTwinkle
Деградация

Предположительные функции D-loop:
D-loop - преждевременно терминированная Н-цепь, образованная при репликации (модель
Предположительные функции D-loop:
D-loop - преждевременно терминированная Н-цепь, образованная при репликации (модель
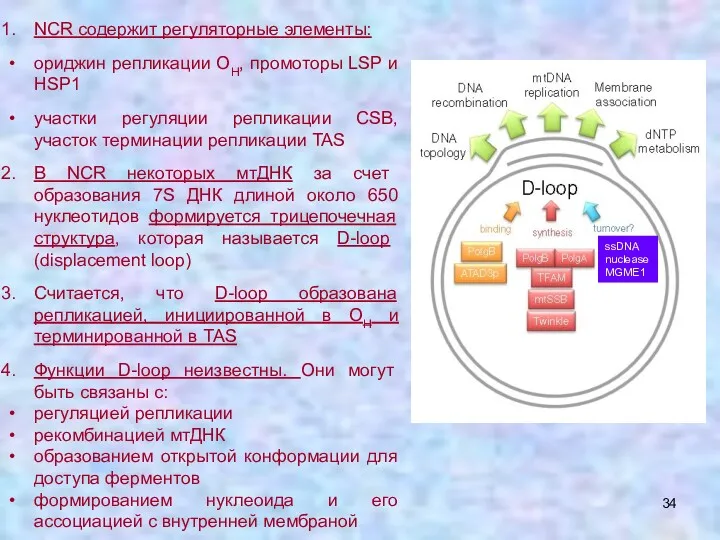
NCR содержит регуляторные элементы:
ориджин репликации OH, промоторы LSP и HSP1
участки регуляции
NCR содержит регуляторные элементы:
ориджин репликации OH, промоторы LSP и HSP1
участки регуляции
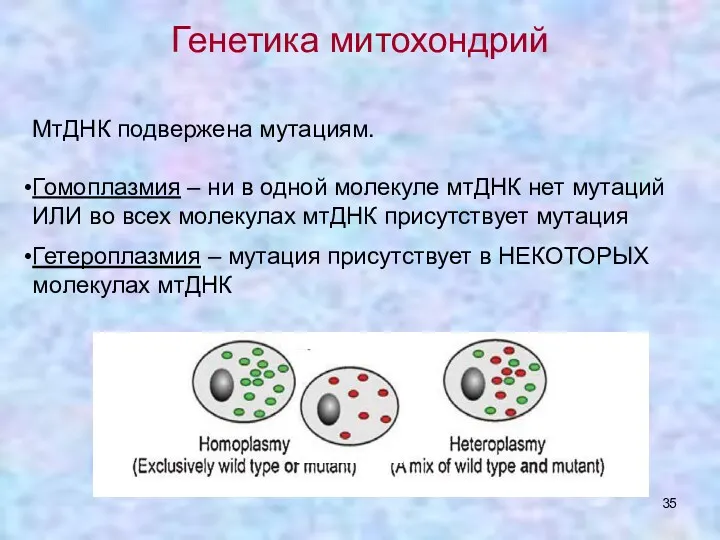
Генетика митохондрий
МтДНК подвержена мутациям.
Гомоплазмия – ни в одной молекуле мтДНК
Генетика митохондрий
МтДНК подвержена мутациям.
Гомоплазмия – ни в одной молекуле мтДНК
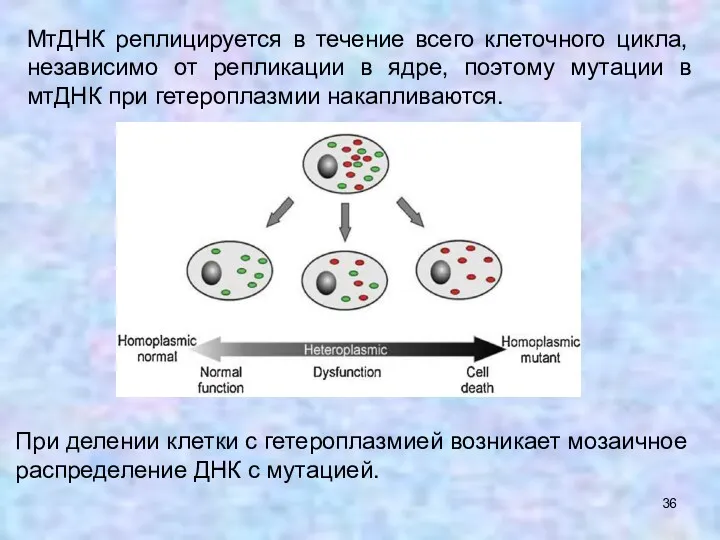
МтДНК реплицируется в течение всего клеточного цикла, независимо от репликации в
МтДНК реплицируется в течение всего клеточного цикла, независимо от репликации в
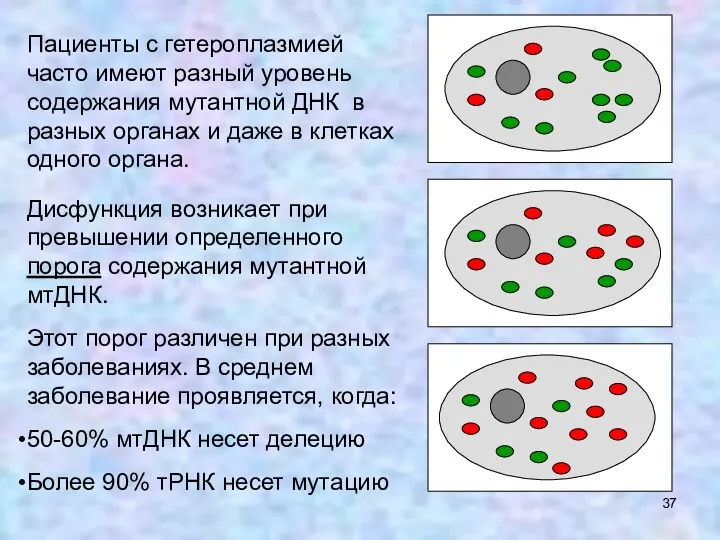
Пациенты с гетероплазмией часто имеют разный уровень содержания мутантной ДНК в
Пациенты с гетероплазмией часто имеют разный уровень содержания мутантной ДНК в
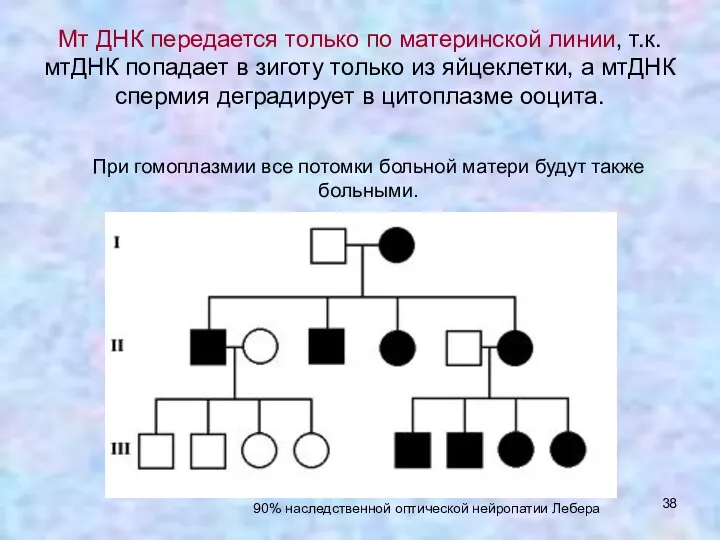
При гомоплазмии все потомки больной матери будут также больными.
90% наследственной
При гомоплазмии все потомки больной матери будут также больными.
90% наследственной
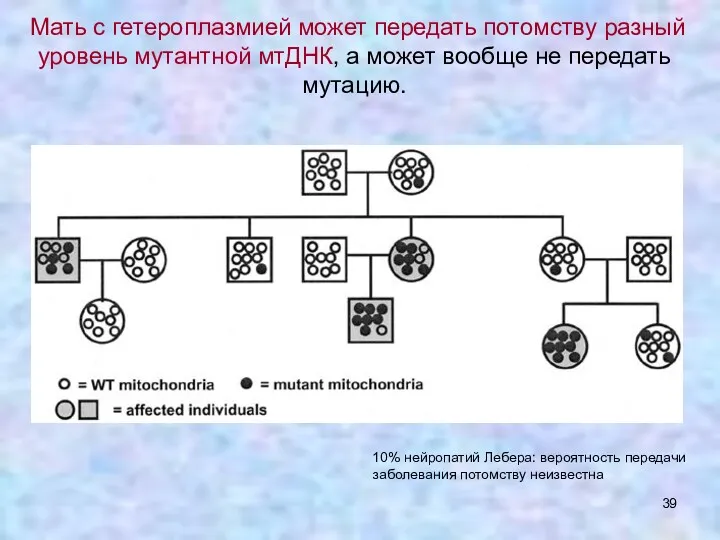
Мать с гетероплазмией может передать потомству разный уровень мутантной мтДНК, а
Мать с гетероплазмией может передать потомству разный уровень мутантной мтДНК, а
 Эволюция органов пищеварения. Обмен веществ
Эволюция органов пищеварения. Обмен веществ Состав и структура сообщества. 9 класс
Состав и структура сообщества. 9 класс Птичьи рекорды
Птичьи рекорды Питание и пищеварение
Питание и пищеварение Внешнее строение птиц, и строение скелета
Внешнее строение птиц, и строение скелета Пищевая микробиология. Дрожжи
Пищевая микробиология. Дрожжи Всё о кошках. История одомашнивания кошек
Всё о кошках. История одомашнивания кошек Археи – уникальная группа организмов
Археи – уникальная группа организмов Опорно-двигательный аппарат
Опорно-двигательный аппарат Язык. Мышечное тело языка
Язык. Мышечное тело языка Тип Хордовые. Подтип Бесчерепные
Тип Хордовые. Подтип Бесчерепные Одноклітинні організми
Одноклітинні організми Primary meristems
Primary meristems Энергетический обмен, катаболизм
Энергетический обмен, катаболизм Строение глаза
Строение глаза Расы и расизм
Расы и расизм Витамины группы В
Витамины группы В Хрящевая и костная ткани
Хрящевая и костная ткани Виртуальная выставка Во саду ли в огороде
Виртуальная выставка Во саду ли в огороде Гіпофіз, епіфіз, щитоподібна залоза
Гіпофіз, епіфіз, щитоподібна залоза Генеративные органы растения. Цветок
Генеративные органы растения. Цветок Культивирование морских брюхоногих моллюсков
Культивирование морских брюхоногих моллюсков Этапы эволюции человека
Этапы эволюции человека Классификация млекопитающих
Классификация млекопитающих Двумембранные органоиды. Ядро
Двумембранные органоиды. Ядро Геном человека. Мультифакториальные болезни
Геном человека. Мультифакториальные болезни Генетика микроорганизмов. Биотехнология
Генетика микроорганизмов. Биотехнология Презентация к внеклассному мероприятию Книга памятников природе
Презентация к внеклассному мероприятию Книга памятников природе