- Митохондрии грибов

Содержание
- 2. Митохондрии (от греч. μίτος — нить и χόνδρος — зёрнышко, крупинка) - самовоспроизводящиеся полуавтономные двумембранные органоиды
- 3. Митохондрии характерны за малым исключением для всех эукариотических клеток, как автотрофных (фотосинтезирующие растения), так и гетеротрофных
- 4. Функции Митохондрии – это мультифункциональные полиморфные органеллы, выполняющие важнейшие функции в клетках эукариотических организмов. Митохондрии осуществляют
- 5. Исследование тонкой структуры митохондрий различных грибов позволило отметить принципиальное сходство с таковой высших организмов. Они окружены
- 6. Наружная мембрана митохондрий содержит белки-порины, гомологичные бактериальным, и проницаема для небольших молекул. Содержание белков в мембранах
- 7. Перенос предшественников белков через комплекс TOM40.
- 8. Перенос предшественников белков через комплекс TOM40. (а) при участии белка Tom70. Транспорт многих предшественников белков, включая
- 9. Биогенез В то время как большинство митохондриальных белков кодируется в ядре и посттрансляционно импортируется в органеллу,
- 10. Происхождение Из имеющихся данных по определению нуклеотидных последовательностей митохондриальных геномов был сделан вывод о монофилетическом (то
- 11. Методы наблюдения митохондрий у грибов Флуоресцентные (потенциалзависимые красители или встраивание GFP плазмиды в геном митохондрий) Методы
- 12. Первые наблюдения митохондрий у гриба Pustularia versispora (Tarzetta catinus) относятся к 1911 году, несколько позже описаны
- 13. Размеры митохондрий Совокупность митохондрий в клетке называют хондриом. Размеры митохондрий непостоянны у разных видов. Обычно митохондрии
- 14. Ультраструктура митохондрий в клетках поверхностно растущего вегетативного мицелия (А) и субгимения (Б) Agaricus bisporus А Б
- 15. - положением клеток в колонии - условиями роста - стадией жизненного цикла и др. Морфология и
- 16. Распределение и морфология митохондрий у мицелиальных грибов
- 17. Положение хондриома в апексе у Neurospora crassa Локализованные в апикальном кончике мицелия митохондрии (до 50 мкм)
- 18. Типы распределения хондриома в клетках мицелия штаммов рода Agaricus (Матросова, 2006) . Тип 1. Характерен для:
- 19. Базидии шампиньона двуспорового
- 20. Влияние стрессового фактора на морфологию (фрагментацию) митохондрий Podospora anserina Родамин 6Ж GFP плазмида контроль Опыт (окислительный
- 21. а – нулевая точка, б – начало фрагментации нитевидных митохондрий (формирование многочисленных перетяжек) через 10 мин
- 22. Распределение и морфология митохондрий у одноклеточных грибов
- 23. Распределение митохондрий в клетке дрожжей Saccharomyces cerevisiae
- 24. Связь клеточного и митохондриального циклов Связь с клеточным циклом. При изучении почкующихся дрожжей было продемонстрировано, что
- 25. Связь клеточного и митохондриального циклов
- 26. Динамика митохондрий и жизненный цикл При половом цикле размножения у S. cerevisiae наблюдается следующая динамика поведения
- 27. Геном митохондрий грибов ДНК в митохондриях представлена циклическими молекулами, не образующими связь с гистонными белками. Митохондриальная
- 28. Наследование митохондрий Сохранение целостности митохондриального генома в процессе наследования очень важно для дыхательной функции. Факторы, контролирующие
- 29. Движение митохондрий В клетках Saccharomyces cerevisiae, как и в клетках Aspergillus nidulans в транспорте митохондрий большую
- 30. Слияние (англ. fusion) и деление (англ.fission или division) митохондрий Динамично протекающие процессы слияния и деления (динамика)
- 31. Слияние (англ. fusion) и деление (англ.fission или division) митохондрий Основными компонентами слияния и деления митохондрий являются
- 32. Westermann, 2008 Westermann, 2008
- 33. Предложено несколько моделей, объясняющих деление митохондрий с участием белков Dnm1, Fis1, Mdv1, Caf4 и Mdm33
- 34. Модель молекулярного аппарата деления митохондрий у дрожжей Mdv1 и Caf4 имеют доменную организацию и выполняют функцию
- 37. Слияние митохондрий Теоретически возможно два пути слияния митохондрий: с одновременным и раздельным слиянием внутренних и внешних
- 38. Модель молекулярного аппарата слияния митохондрий у дрожжей. Две молекулы Fzo1 связывают две смежные митохондрии путем сборки
- 39. Процесс слияния митохондрий можно разделить на три этапа — фиксация органелл, слияние наружных и слияние внутренних
- 40. Основными регуляторами слияния митохондрий у дрожжей являются ГТФазы наружной и внутренней мембран митохондрий Fzo1 и Mgm1
- 41. Субкомпартменты внутренней мембраны митохондрий Митохондрии содержат около 800 белков/1500 у человека Субкомпартментализация внутренней мембраны митохондрий. Распространение
- 42. Дыхательная цепь митохондрий I – НАДН-дегидрогеназа II – сукцинатдегидрогеназа III – убихинол-цитохром с-редуктаза IV – цитохром
- 43. Комплексы дыхательной цепи построены из множества полипептидов и содержат ряд различных окислительно-восстановительных коферментов, связанных с белками.
- 44. На каждый комплекс действуют определенные ингибиторы, например, ротенон и пиерицидин на Комплекс I, антимицин и миксотиазол
- 45. Филогенетическое древо АО, построенное на основании анализа первичных структур.
- 46. АО найдена у многих видов дрожжей, включая Rhodotorula glutinis, Candida lipolytica (теперь Yarrowia lipolytica), C. parapsilosis,
- 47. АО обнаружена в ряде грибов, включая Ustilago ayclis, U. maydis, Tapesia acuformis, Aspergillus niger, A. fumigatus,
- 48. Метаболические процессы
- 49. Метаболические связи пероксисом и митохондрий
- 51. β-окисление жирных кислот протекает в митохондриях с участием четырех ферментов: ацил-СоА дегидрогеназа, эноил-СоА гидратаза, 3-ОН-ацил-СоА дегидрогеназа,
- 52. □ ■ β-окисление жирных кислот протекает в митохондриях (ацил-СоА дегидрогеназа, эноил-СоА гидратаза, 3-ОН-ацил-СоА дегидрогеназа, 3-кето-ацил-СоА тиолаза).
- 53. Связь митохондрий с другими органеллами: пероксисомы и ядра (митоз и мейоз) (клетки мицелия Agaricus bisporus, связь
- 54. Схема митофагии у дрожжей. Митофагия индуцируется снижением мембранного митохондриального потенциала, азотным голоданием, рапамицином или входе в
- 55. Макромитофагия Piptoporus betulinus (21 ср. мальт-агар) митохондрия лизосома
- 56. Старение (англ. аging) это фундаментальный процесс обнаруженный почти у всех биологических систем и может быть определен
- 57. Признаки старения Podospora anserina and Neurospora crassa (senescent strains) (Griffiths, 1992, Osiewacz and Hamman, 2006), которые
- 58. Митохондриальные факторы старения P. anserina Схема вырезания и накапливания последовательности α-sen в стареющих культурах P. anserina
- 59. Накопление факторов старения у Рodospora anserina Гифа сверху Модельная схема накопления и распределения между клеток условного
- 60. Накопление факторов старения у дрожжей Модельная схема накопления и распределения между клетками условного фактора старения у
- 61. Роль митохондрий в апоптозе клетки Особая роль митохондрий в системе выбора клетки между жизнью и смертью
- 62. Признаки апоптоза у дрожжей Saccharomyces cerevisiae
- 63. Схема путей апоптоза у грибов
- 64. Морфология митохондрий клеток глубинного мицелия Agaricus bisporus, находящегося в стадии апоптоза
- 65. Ультраструктура апоптотической клетки глубинного мицелия Agaricus bisporus после длительного культивирования
- 66. Гидрогеносомы
- 68. Распространение Гидрогеносомы были обнаружены примерно 40 лет назад. Эти органеллы присутствуют у разных неродственных анаэробных эукариот,
- 69. Происхождение Полагают, что эти органеллы произошли от эндосимбиотических анаэробных бактерий или архей, хотя в случае трихомонад
- 70. Функции Главная функция гидрогеносом – это окислительное декарбоксилирование пирувата, сопряженное с образованием АТФ, с электронным транспортом
- 71. Анаэробные хитридиевые (например, виды Neocallimastix и Piromyces) метаболизируют малат (образованный из глюкозы) с образованием АТФ и
- 72. Ключевые ферменты гидрогеносом: пируват:ферредоксин оксидоредуктаза и гидрогеназа
- 74. Скачать презентацию

Митохондрии (от греч. μίτος — нить и χόνδρος — зёрнышко, крупинка) - самовоспроизводящиеся
Митохондрии (от греч. μίτος — нить и χόνδρος — зёрнышко, крупинка) - самовоспроизводящиеся

Митохондрии характерны за малым исключением для всех эукариотических клеток, как автотрофных
Митохондрии характерны за малым исключением для всех эукариотических клеток, как автотрофных

Функции
Митохондрии – это мультифункциональные полиморфные органеллы, выполняющие важнейшие функции в клетках
Функции
Митохондрии – это мультифункциональные полиморфные органеллы, выполняющие важнейшие функции в клетках

Исследование тонкой структуры митохондрий различных грибов позволило отметить принципиальное сходство с
Исследование тонкой структуры митохондрий различных грибов позволило отметить принципиальное сходство с

Наружная мембрана митохондрий содержит белки-порины, гомологичные бактериальным, и проницаема для небольших
Наружная мембрана митохондрий содержит белки-порины, гомологичные бактериальным, и проницаема для небольших
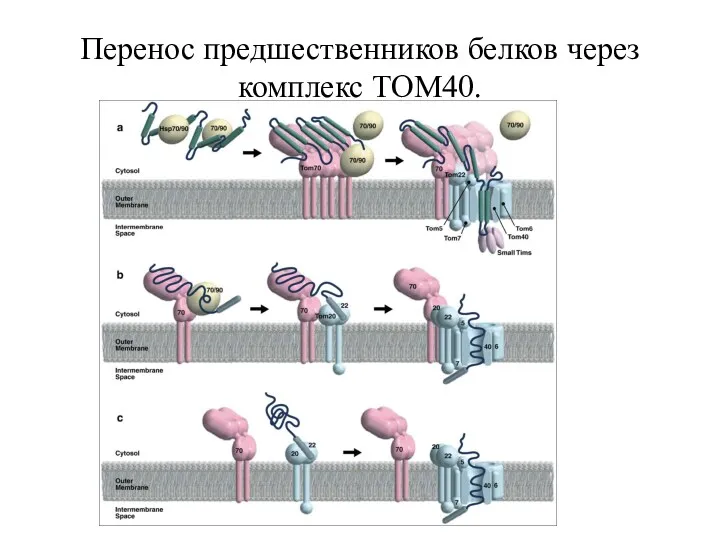
Перенос предшественников белков через комплекс TOM40.
Перенос предшественников белков через комплекс TOM40.

Перенос предшественников белков через комплекс TOM40.
(а) при участии белка Tom70. Транспорт
Перенос предшественников белков через комплекс TOM40.
(а) при участии белка Tom70. Транспорт

Биогенез
В то время как большинство митохондриальных белков кодируется в ядре и
Биогенез
В то время как большинство митохондриальных белков кодируется в ядре и

Происхождение
Из имеющихся данных по определению нуклеотидных последовательностей митохондриальных геномов был сделан
Происхождение
Из имеющихся данных по определению нуклеотидных последовательностей митохондриальных геномов был сделан
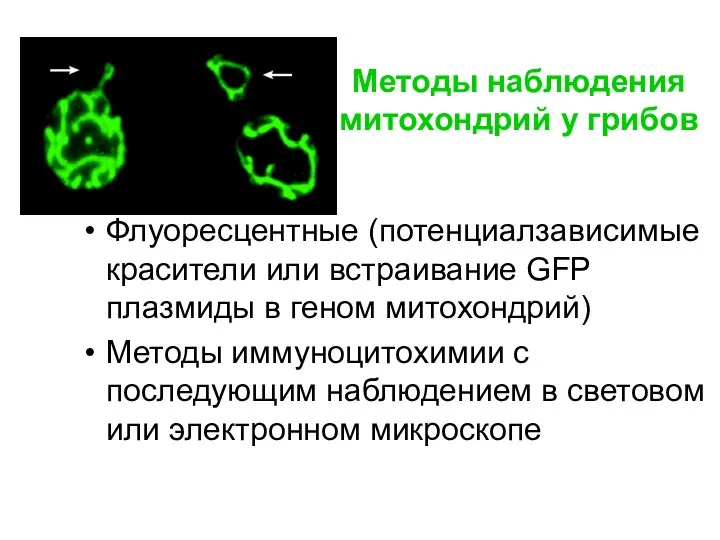
Методы наблюдения митохондрий у грибов
Флуоресцентные (потенциалзависимые красители или встраивание GFP плазмиды
Методы наблюдения митохондрий у грибов
Флуоресцентные (потенциалзависимые красители или встраивание GFP плазмиды

Первые наблюдения митохондрий у гриба Pustularia versispora (Tarzetta catinus) относятся к
Первые наблюдения митохондрий у гриба Pustularia versispora (Tarzetta catinus) относятся к

Размеры митохондрий
Совокупность митохондрий в клетке называют хондриом.
Размеры митохондрий непостоянны
Размеры митохондрий
Совокупность митохондрий в клетке называют хондриом.
Размеры митохондрий непостоянны
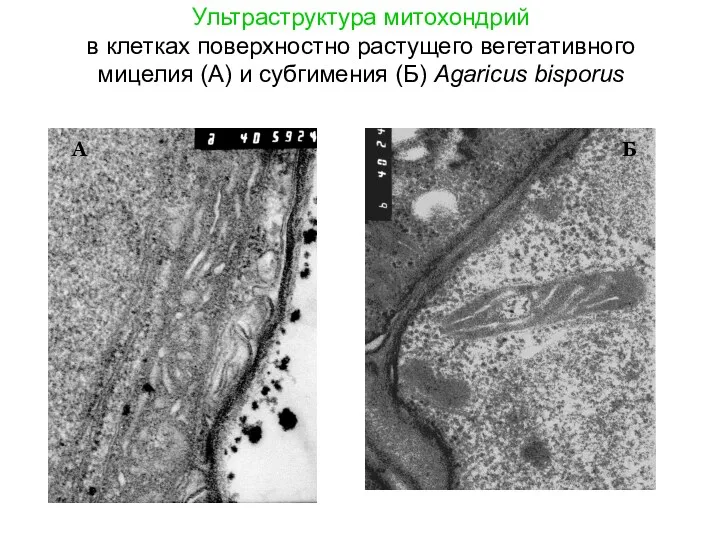
Ультраструктура митохондрий
в клетках поверхностно растущего вегетативного мицелия (А) и субгимения
Ультраструктура митохондрий в клетках поверхностно растущего вегетативного мицелия (А) и субгимения

- положением клеток в колонии - условиями роста
- стадией
- положением клеток в колонии - условиями роста - стадией

Распределение и морфология митохондрий у мицелиальных грибов
Распределение и морфология митохондрий у мицелиальных грибов
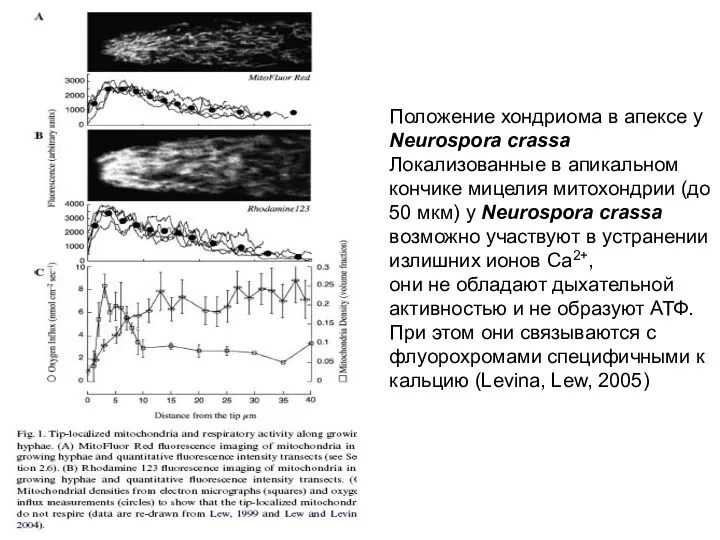
Положение хондриома в апексе у
Neurospora crassa Локализованные в апикальном кончике
Положение хондриома в апексе у Neurospora crassa Локализованные в апикальном кончике
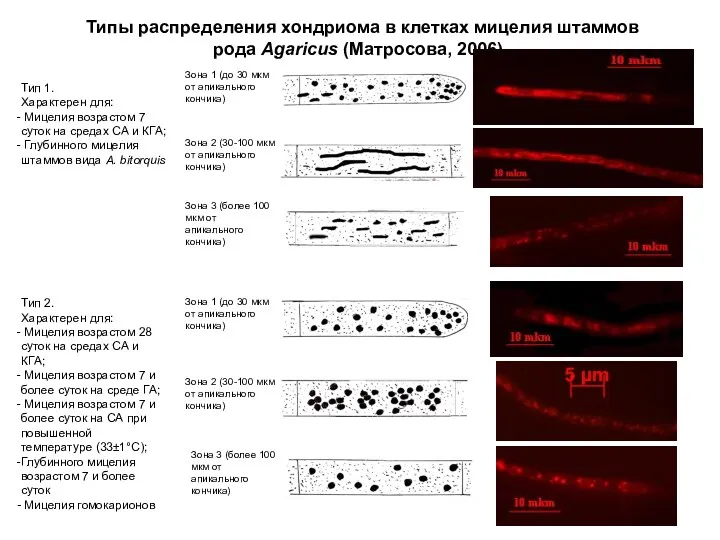
Типы распределения хондриома в клетках мицелия штаммов рода Agaricus (Матросова, 2006)
Типы распределения хондриома в клетках мицелия штаммов рода Agaricus (Матросова, 2006)

Базидии шампиньона двуспорового
Базидии шампиньона двуспорового
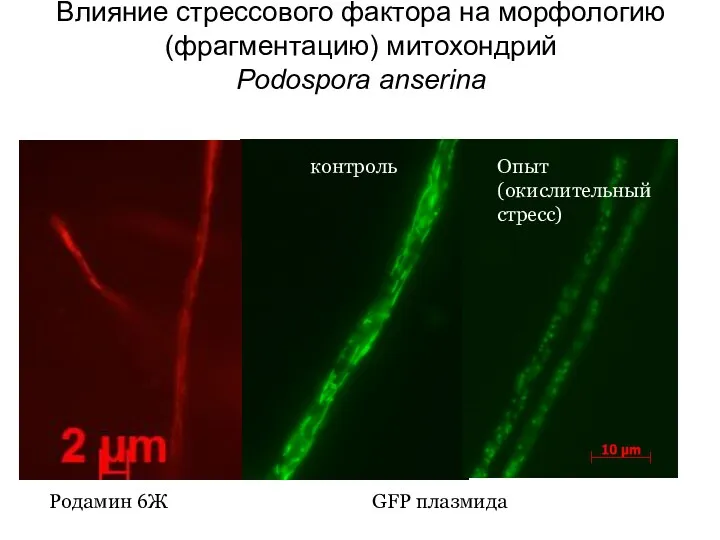
Влияние стрессового фактора на морфологию (фрагментацию) митохондрий
Podospora anserina
Родамин 6Ж
GFP плазмида
контроль
Опыт
Влияние стрессового фактора на морфологию (фрагментацию) митохондрий
Podospora anserina
Родамин 6Ж
GFP плазмида
контроль
Опыт
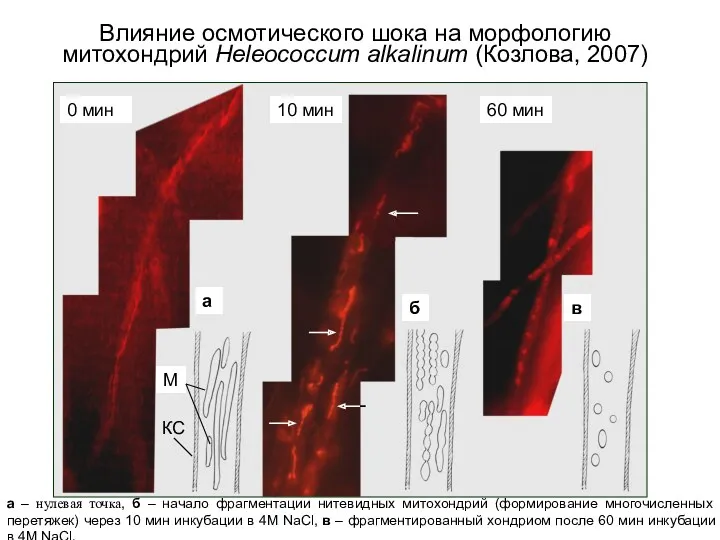
а – нулевая точка, б – начало фрагментации нитевидных митохондрий (формирование
а – нулевая точка, б – начало фрагментации нитевидных митохондрий (формирование

Распределение и морфология митохондрий у одноклеточных грибов
Распределение и морфология митохондрий у одноклеточных грибов
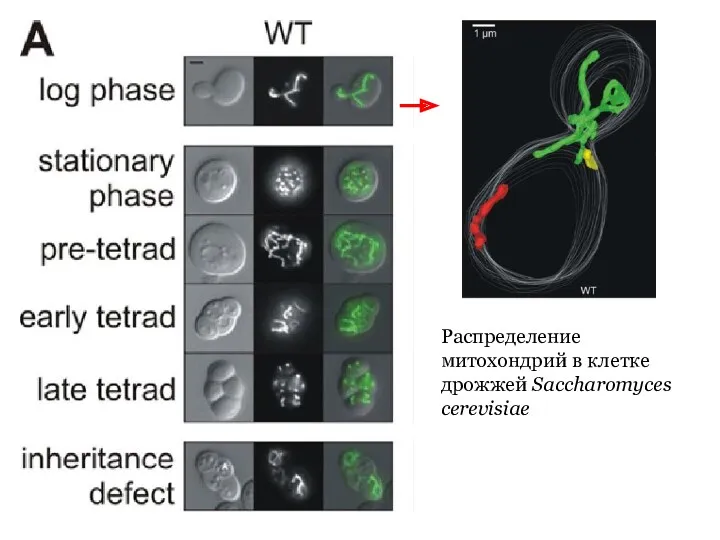
Распределение митохондрий в клетке дрожжей Saccharomyces cerevisiae
Распределение митохондрий в клетке дрожжей Saccharomyces cerevisiae

Связь клеточного и митохондриального циклов
Связь с клеточным циклом. При изучении почкующихся
Связь клеточного и митохондриального циклов
Связь с клеточным циклом. При изучении почкующихся
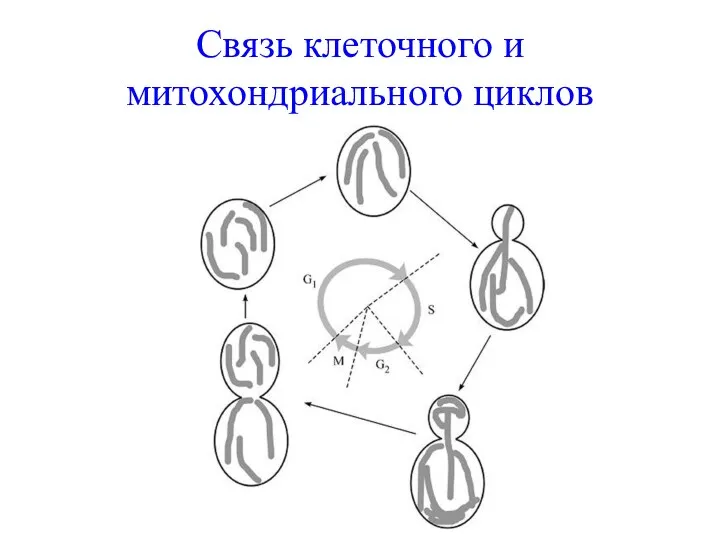
Связь клеточного и митохондриального циклов
Связь клеточного и митохондриального циклов

Динамика митохондрий и жизненный цикл
При половом цикле размножения у S. cerevisiae
Динамика митохондрий и жизненный цикл
При половом цикле размножения у S. cerevisiae

Геном митохондрий грибов
ДНК в митохондриях представлена циклическими молекулами, не образующими связь
Геном митохондрий грибов
ДНК в митохондриях представлена циклическими молекулами, не образующими связь

Наследование митохондрий
Сохранение целостности митохондриального генома в процессе наследования очень важно для
Наследование митохондрий
Сохранение целостности митохондриального генома в процессе наследования очень важно для

Движение митохондрий
В клетках Saccharomyces cerevisiae, как и в клетках Aspergillus nidulans
Движение митохондрий
В клетках Saccharomyces cerevisiae, как и в клетках Aspergillus nidulans

Слияние (англ. fusion) и деление (англ.fission или division) митохондрий
Динамично протекающие процессы
Слияние (англ. fusion) и деление (англ.fission или division) митохондрий
Динамично протекающие процессы

Слияние (англ. fusion) и деление (англ.fission или division) митохондрий
Основными компонентами слияния
Слияние (англ. fusion) и деление (англ.fission или division) митохондрий
Основными компонентами слияния

Westermann, 2008
Westermann, 2008
Westermann, 2008
Westermann, 2008

Предложено несколько моделей, объясняющих деление митохондрий с участием белков Dnm1, Fis1,
Предложено несколько моделей, объясняющих деление митохондрий с участием белков Dnm1, Fis1,
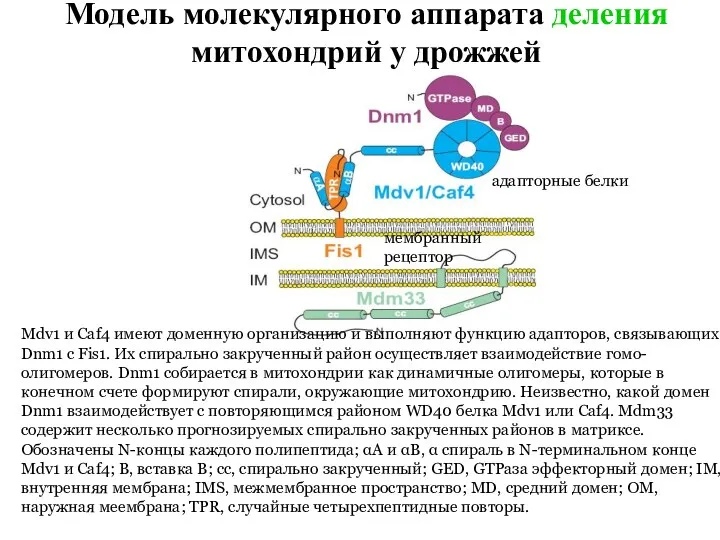
Модель молекулярного аппарата деления митохондрий у дрожжей
Mdv1 и Caf4 имеют доменную
Модель молекулярного аппарата деления митохондрий у дрожжей
Mdv1 и Caf4 имеют доменную

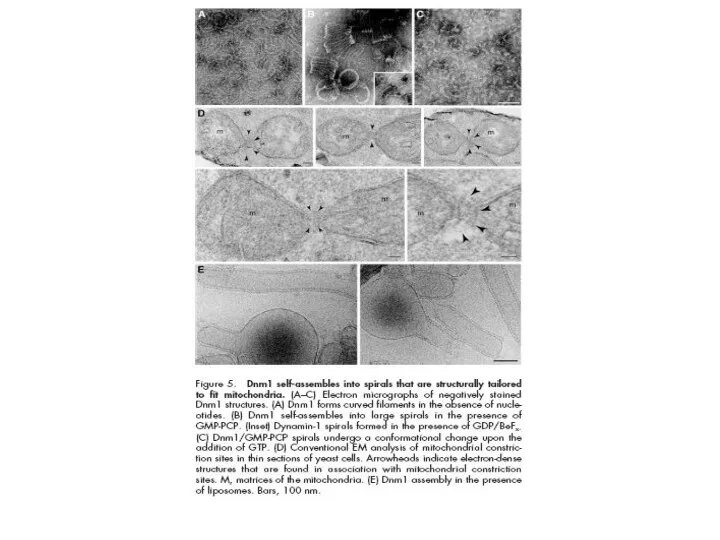

Слияние митохондрий
Теоретически возможно два пути слияния митохондрий: с одновременным и раздельным
Слияние митохондрий
Теоретически возможно два пути слияния митохондрий: с одновременным и раздельным
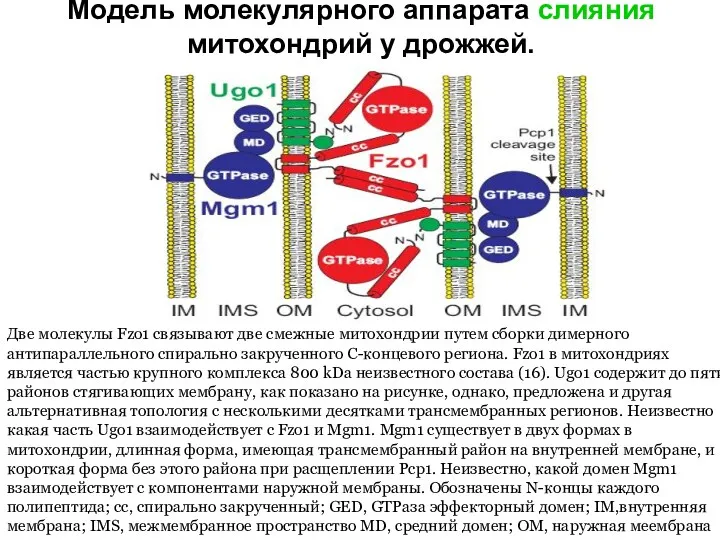
Модель молекулярного аппарата слияния митохондрий у дрожжей.
Две молекулы Fzo1 связывают две
Модель молекулярного аппарата слияния митохондрий у дрожжей.
Две молекулы Fzo1 связывают две

Процесс слияния митохондрий можно разделить на три этапа — фиксация органелл,
Процесс слияния митохондрий можно разделить на три этапа — фиксация органелл,

Основными регуляторами слияния митохондрий у дрожжей являются ГТФазы наружной и внутренней
Основными регуляторами слияния митохондрий у дрожжей являются ГТФазы наружной и внутренней
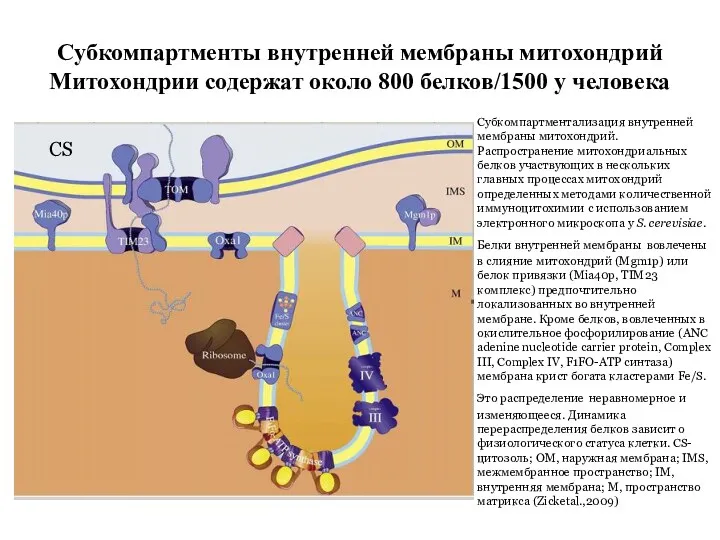
Субкомпартменты внутренней мембраны митохондрий
Митохондрии содержат около 800 белков/1500 у человека
Субкомпартментализация внутренней
Субкомпартменты внутренней мембраны митохондрий
Митохондрии содержат около 800 белков/1500 у человека
Субкомпартментализация внутренней
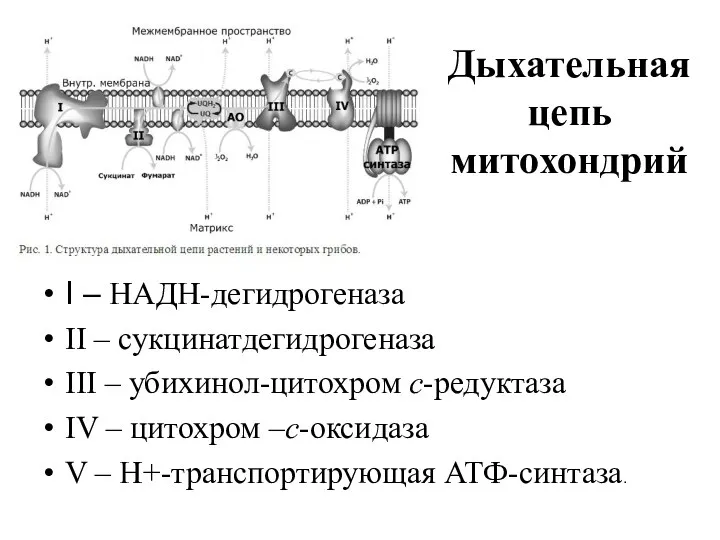
Дыхательная цепь митохондрий
I – НАДН-дегидрогеназа
II – сукцинатдегидрогеназа
III – убихинол-цитохром с-редуктаза
IV
Дыхательная цепь митохондрий
I – НАДН-дегидрогеназа
II – сукцинатдегидрогеназа
III – убихинол-цитохром с-редуктаза
IV

Комплексы дыхательной цепи построены из множества полипептидов и содержат ряд различных
Комплексы дыхательной цепи построены из множества полипептидов и содержат ряд различных

На каждый комплекс действуют определенные ингибиторы, например, ротенон и пиерицидин на
На каждый комплекс действуют определенные ингибиторы, например, ротенон и пиерицидин на
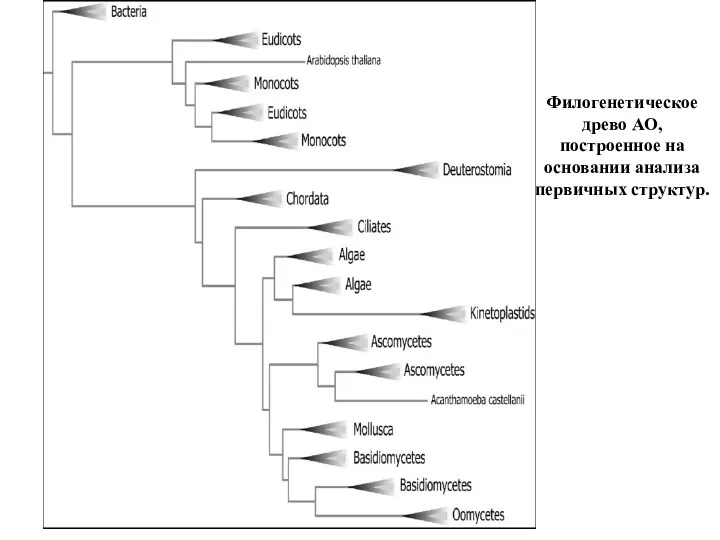
Филогенетическое древо АО, построенное на основании анализа первичных структур.
Филогенетическое древо АО, построенное на основании анализа первичных структур.
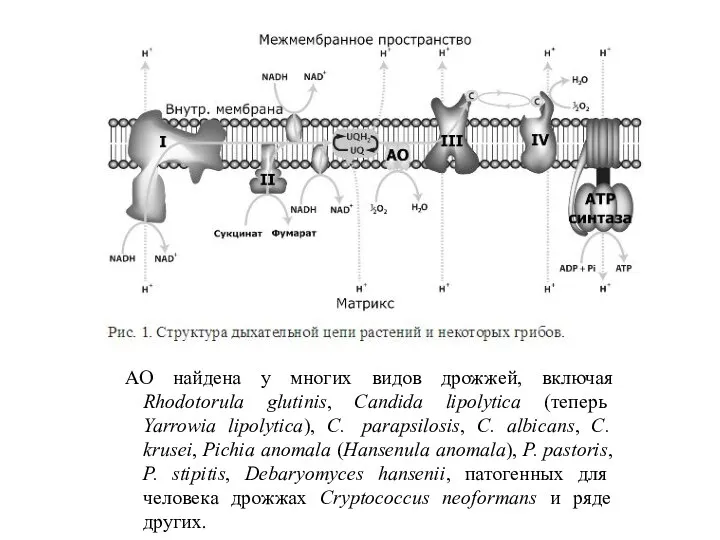
АО найдена у многих видов дрожжей, включая Rhodotorula glutinis, Candida lipolytica
АО найдена у многих видов дрожжей, включая Rhodotorula glutinis, Candida lipolytica

АО обнаружена в ряде грибов, включая Ustilago ayclis, U. maydis, Tapesia
АО обнаружена в ряде грибов, включая Ustilago ayclis, U. maydis, Tapesia
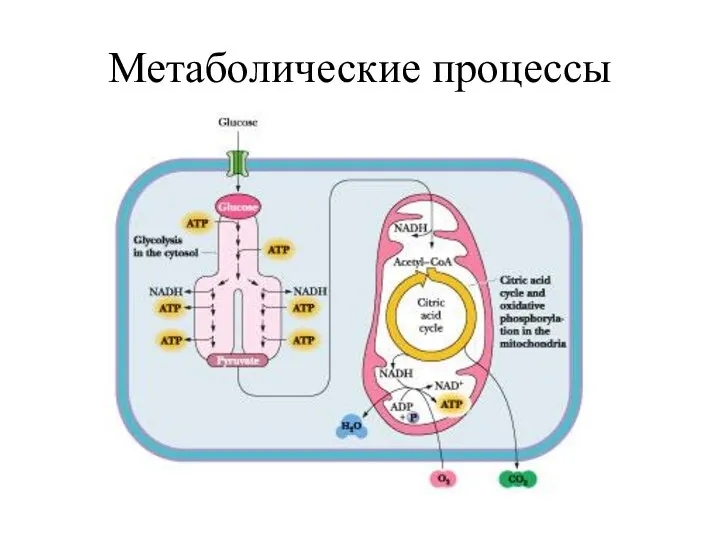
Метаболические процессы
Метаболические процессы

Метаболические связи пероксисом и митохондрий
Метаболические связи пероксисом и митохондрий
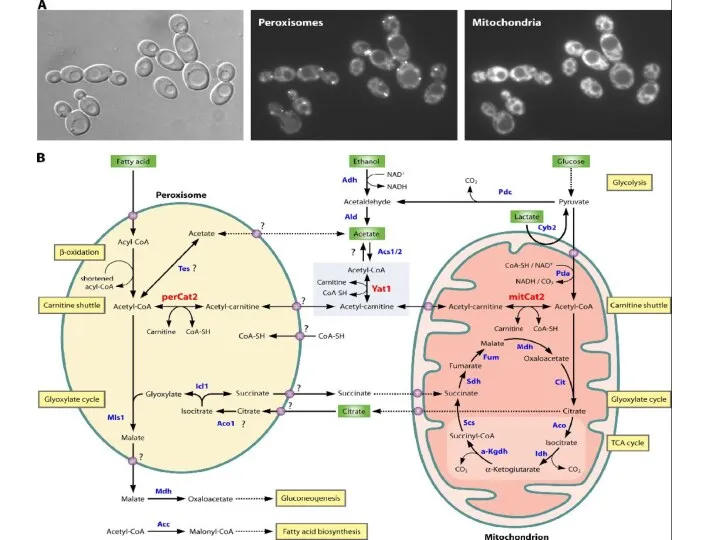

β-окисление жирных кислот протекает в митохондриях с участием четырех ферментов: ацил-СоА
β-окисление жирных кислот протекает в митохондриях с участием четырех ферментов: ацил-СоА
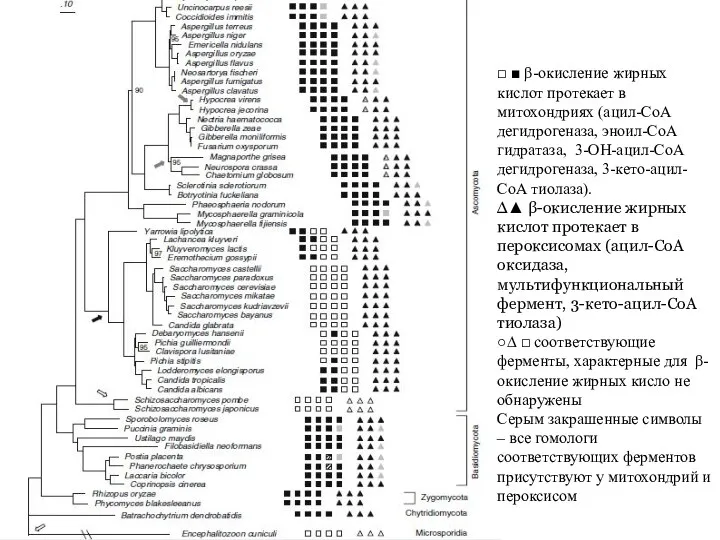
□ ■ β-окисление жирных кислот протекает в митохондриях (ацил-СоА дегидрогеназа, эноил-СоА

Связь митохондрий с другими органеллами: пероксисомы и ядра
(митоз и мейоз) (клетки
Связь митохондрий с другими органеллами: пероксисомы и ядра (митоз и мейоз) (клетки
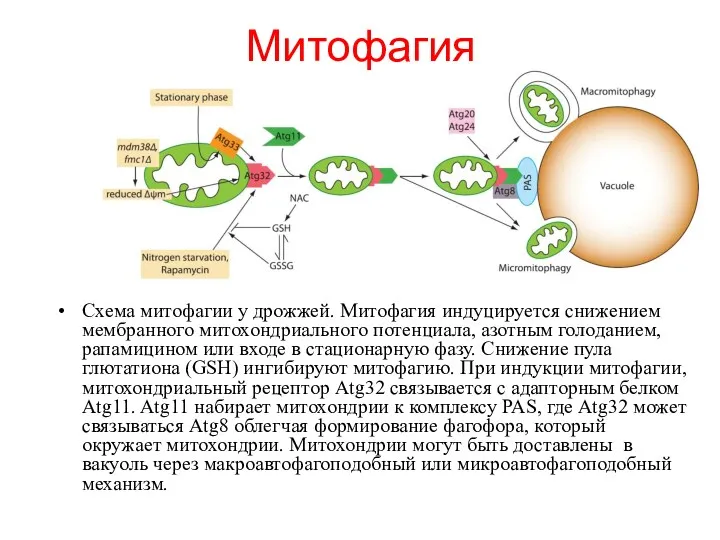
Схема митофагии у дрожжей. Митофагия индуцируется снижением мембранного митохондриального потенциала, азотным
Схема митофагии у дрожжей. Митофагия индуцируется снижением мембранного митохондриального потенциала, азотным
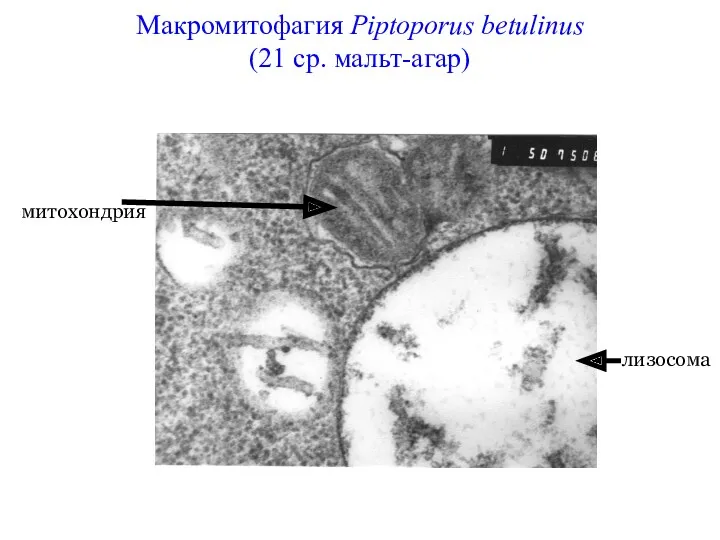
Макромитофагия Piptoporus betulinus
(21 ср. мальт-агар)
митохондрия
лизосома
Макромитофагия Piptoporus betulinus
(21 ср. мальт-агар)
митохондрия
лизосома
Старение (англ. аging) это фундаментальный процесс обнаруженный почти у всех
Старение (англ. аging) это фундаментальный процесс обнаруженный почти у всех

Признаки старения Podospora anserina and Neurospora crassa (senescent strains) (Griffiths, 1992,
Признаки старения Podospora anserina and Neurospora crassa (senescent strains) (Griffiths, 1992,
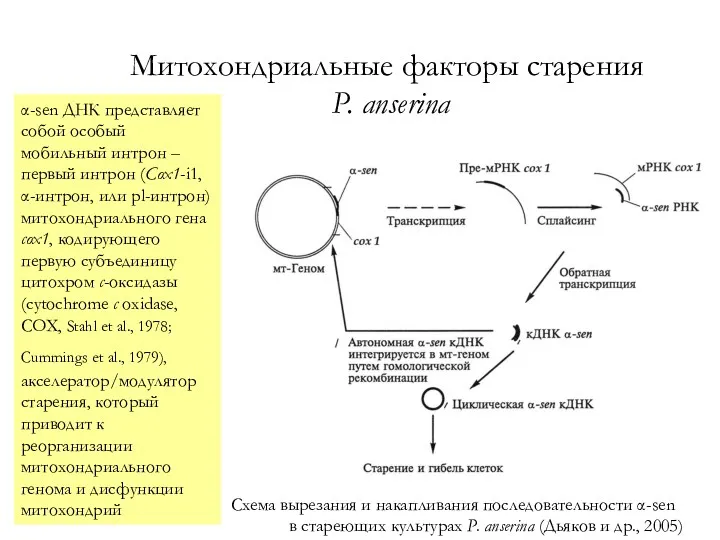
Митохондриальные факторы старения
P. anserina
Схема вырезания и накапливания последовательности α-sen
в
Митохондриальные факторы старения
P. anserina
Схема вырезания и накапливания последовательности α-sen
в

Накопление факторов старения у Рodospora anserina
Гифа сверху Модельная схема накопления и
Накопление факторов старения у Рodospora anserina
Гифа сверху Модельная схема накопления и

Накопление факторов старения у дрожжей
Модельная схема накопления и распределения между клетками
Накопление факторов старения у дрожжей
Модельная схема накопления и распределения между клетками

Роль митохондрий в апоптозе клетки
Особая роль митохондрий в системе выбора клетки
Роль митохондрий в апоптозе клетки Особая роль митохондрий в системе выбора клетки
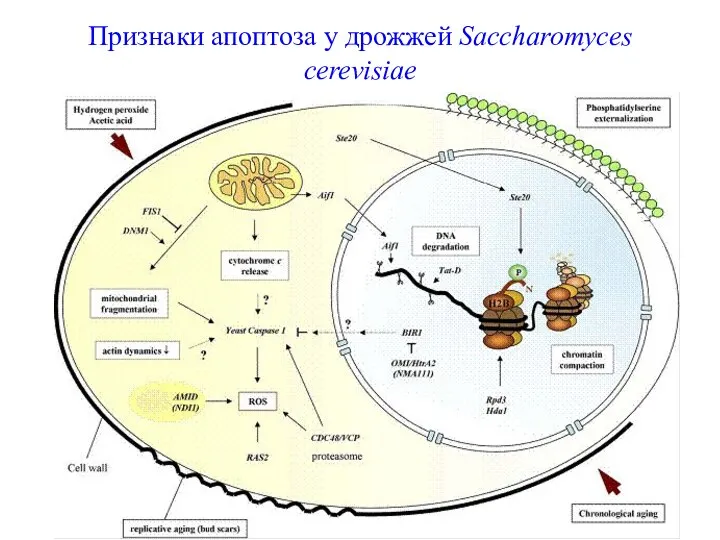
Признаки апоптоза у дрожжей Saccharomyces cerevisiae
Признаки апоптоза у дрожжей Saccharomyces cerevisiae
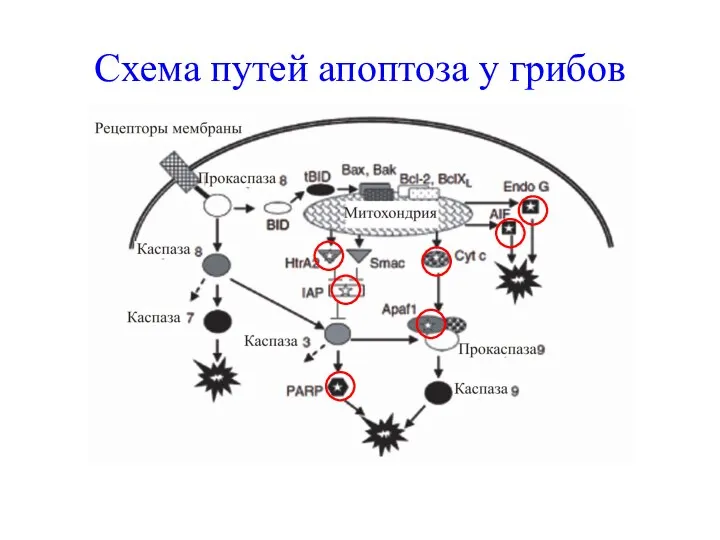
Схема путей апоптоза у грибов
Схема путей апоптоза у грибов
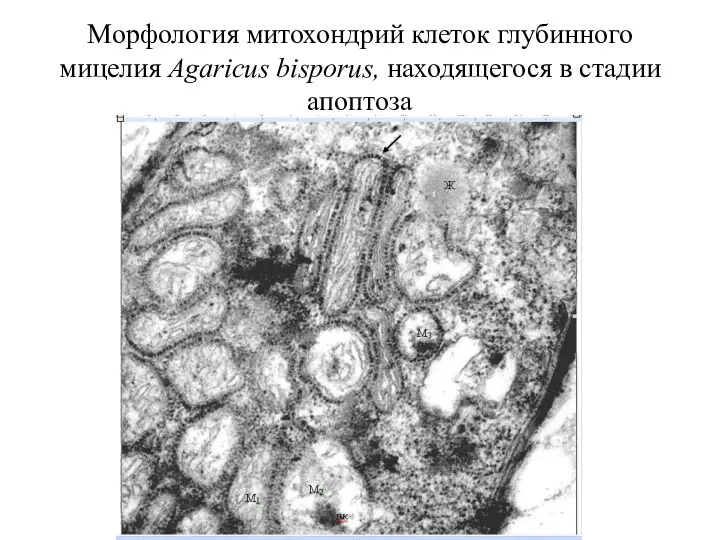
Морфология митохондрий клеток глубинного мицелия Agaricus bisporus, находящегося в стадии апоптоза
Морфология митохондрий клеток глубинного мицелия Agaricus bisporus, находящегося в стадии апоптоза
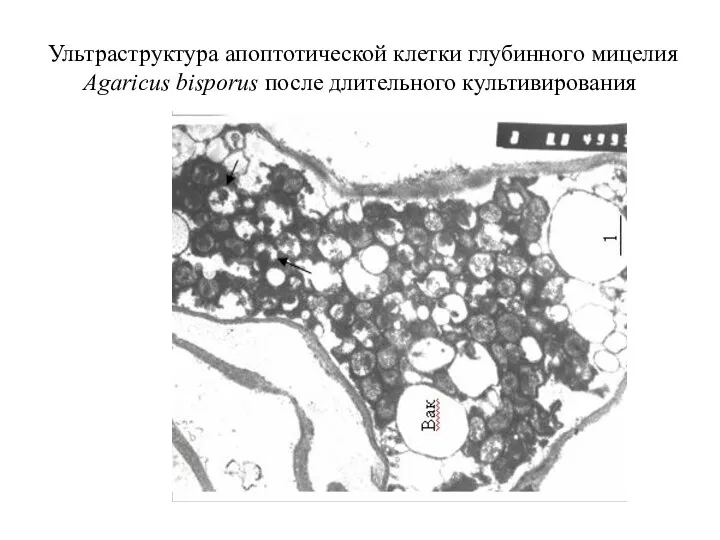
Ультраструктура апоптотической клетки глубинного мицелия Agaricus bisporus после длительного культивирования
Ультраструктура апоптотической клетки глубинного мицелия Agaricus bisporus после длительного культивирования
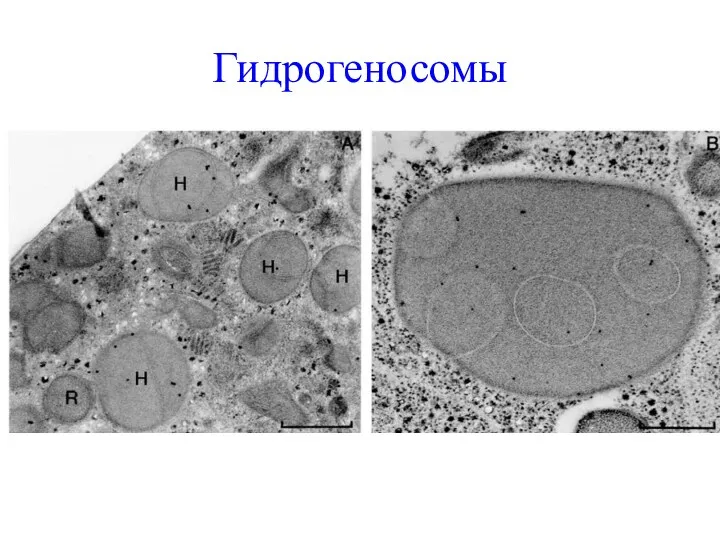
Гидрогеносомы
Гидрогеносомы
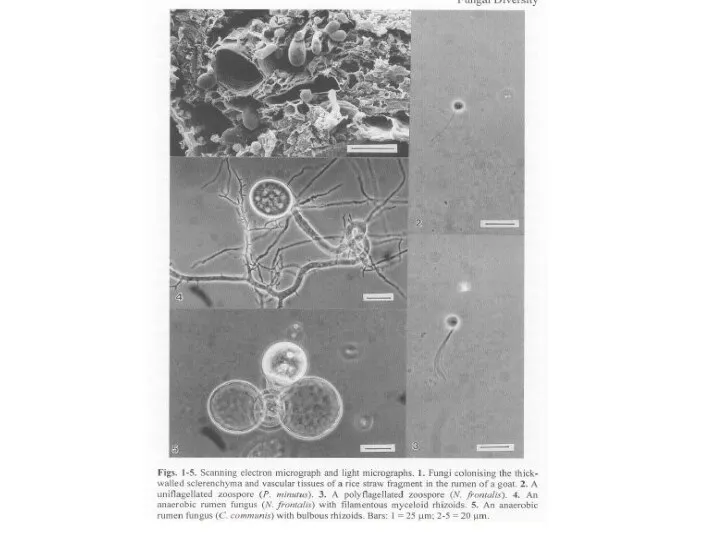

Распространение
Гидрогеносомы были обнаружены примерно 40 лет назад. Эти органеллы присутствуют у
Распространение
Гидрогеносомы были обнаружены примерно 40 лет назад. Эти органеллы присутствуют у

Происхождение
Полагают, что эти органеллы произошли от эндосимбиотических анаэробных бактерий или архей,
Происхождение
Полагают, что эти органеллы произошли от эндосимбиотических анаэробных бактерий или архей,

Функции
Главная функция гидрогеносом – это окислительное декарбоксилирование пирувата, сопряженное с образованием
Функции
Главная функция гидрогеносом – это окислительное декарбоксилирование пирувата, сопряженное с образованием

Анаэробные хитридиевые (например, виды Neocallimastix и Piromyces) метаболизируют малат (образованный из
Анаэробные хитридиевые (например, виды Neocallimastix и Piromyces) метаболизируют малат (образованный из
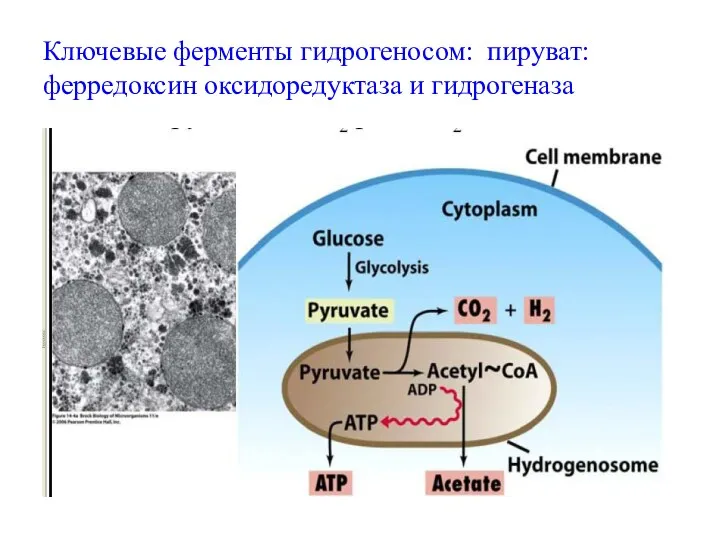
Ключевые ферменты гидрогеносом: пируват:ферредоксин оксидоредуктаза и гидрогеназа
Ключевые ферменты гидрогеносом: пируват:ферредоксин оксидоредуктаза и гидрогеназа
 Ботаника – наука о растениях
Ботаника – наука о растениях Соединительная ткань и ее виды
Соединительная ткань и ее виды Строение и работа сердца. Круги кровообращения
Строение и работа сердца. Круги кровообращения Динозавры
Динозавры Организмы Царства Грибы и Лишайники
Организмы Царства Грибы и Лишайники Хронобиология и биоритмы человека
Хронобиология и биоритмы человека Онкогены. Активирующие мутации в онкогенах. Вирусный канцерогенез (HPV, вирус Эпштейн-Барр). Гены-супрессоры
Онкогены. Активирующие мутации в онкогенах. Вирусный канцерогенез (HPV, вирус Эпштейн-Барр). Гены-супрессоры Мезозой или мезозойская эра. Флора и фауна
Мезозой или мезозойская эра. Флора и фауна Клетка – элементарная единица жизни на земле
Клетка – элементарная единица жизни на земле Физиология бактерий
Физиология бактерий Систематика як наука про класифікацію рослин
Систематика як наука про класифікацію рослин Женская половая система
Женская половая система Вредители виноградной лозы
Вредители виноградной лозы Основные понятия генетики
Основные понятия генетики Возрастная анатомия, физиология и гигиена
Возрастная анатомия, физиология и гигиена Расщепление пероксида водорода в клетках клубня картофеля. Лабораторная работа №1
Расщепление пероксида водорода в клетках клубня картофеля. Лабораторная работа №1 Молекулярно-генетические механизмы развития корня
Молекулярно-генетические механизмы развития корня Молодое растеньице, появившееся при прорастании семени
Молодое растеньице, появившееся при прорастании семени Эволюция выделительной системы
Эволюция выделительной системы Организация древесины на клеточном уровне. Лекция 4
Организация древесины на клеточном уровне. Лекция 4 Биологическое оружие
Биологическое оружие Селекцiя
Селекцiя Натуральные волокна животного происхождения. Урок 1. 6 класс
Натуральные волокна животного происхождения. Урок 1. 6 класс Экологические группы птиц презентация
Экологические группы птиц презентация Развитие жизни на Земле
Развитие жизни на Земле Прикладные аспекты. Получение антител
Прикладные аспекты. Получение антител Викторина по теме Биотические связи в природе
Викторина по теме Биотические связи в природе Отличительные признаки классов растений
Отличительные признаки классов растений