- Нарушение обмена веществ и энергии

Содержание
- 2. УРОВНИ НАРУШЕНИЯ ОБМЕНА ВЕЩЕСТВ Молекулярный уровень нарушения в генетическом аппарате действие ингибиторов ферментов недостаточное поступление незаменимых
- 3. ОБМЕН УГЛЕВОДОВ И ЕГО НАРУШЕНИЯ
- 4. Все организмы получают химическую энергию из молекул таких как глюкоза Клетки расщепляют глюкозу и накапливают освобождаемую
- 5. Наиболее важным углеводом крови является ГЛ У составляют не более 2% общего сухого веса тканей человеческого
- 6. УРОВЕНЬ ГЛ В КРОВИ Определяется балансом между: УРОВЕНЬ ГЛ всасыванием из кишечника АКТИВАЦИЯ глюконеогенеза АКТИВАЦИЯ гликогенолиза
- 7. Усвоение углеводов основным источником У для человека являются У пищи: крахмал растений гликоген животной ткани полисахара
- 8. крахмал сахароза лактоза α-амилаза слюны Крахмал Декстрины Желудок Ротовая полость ПЖ Кишечник Энтероцит Расщепление крахмала или
- 9. Фруктоза Глюкоза Na+ Галактоза Na+ 3 Na+ 2 К+ 3 Na+ 2 К+ АТФ Фн АДФ
- 10. У выполняют в организме жизненно важные функции, принимая участие: в структурной и метаболической организации органов и
- 11. гликоген гликогенсинтетаза глюкоза гексокиназа глюкокиназа Г - 6 - Ф пируват АцКоА цикл Кребса СО2 Динамика
- 12. Интеграция липидного, углеводного и белкового обмена Ацетил-КоА ЖК углеводы АК цитрат малонил-КоА ацил-ацетил-КоА СО2 окисление через
- 13. Основным клиническим биохимическим показателем нарушений У обмена является изменение концентрации в крови ГЛ гипергликемия увеличение содержания
- 14. ОСОБЕННОСТИ ОБМЕНА УГЛЕВОДОВ У ДЕТЕЙ За счет У у грудных детей покрывается 40% суточной энергетической потребности,
- 15. ГИПЕРГЛИКЕМИЯ (последствия) Центральные осморецепторы ЖАЖДА Полидипсия полиурия ГЛЮКОЗУРИЯ ПОТЕРЯ Na+ К+ Mg2+ Р Сa2+ Cl- ГИПОВОЛЕМИЯ
- 16. Н2О С Ф ГЛ
- 17. нарушение переваривания и всасывания углеводов нарушение экскреторной функции ПЖ отсутствие выработки или сниженная активность дисахаридаз в
- 18. НАСЛЕДСТВЕННЫЕ НАРУШЕНИЯ ОБМЕНА ГАЛАКТОЗЕМИЯ Галактоза галактоза- 1 - фосфат Г- 1 - Ф недостаток галактокиназы недостаток
- 19. ФРУКТОЗЕМИЯ Фруктоза фруктоза- 1 - фосфат Г- 1 - Ф недостаток фруктокиназы недостаток фруктозо-1-фосфоальдолазы Часть фруктозы
- 20. ГЛИКОГЕНОЗЫ (болезни накопления) нарушение ферментативного распада ГЛК печени и мышц из-за врожденного дефицита или отсутствия ферментов
- 21. ОСОБЕННОСТИ НАРУШЕНИЯ ОБМЕНА УГЛЕВОДОВ У ДЕТЕЙ одно из наиболее частых проявлений нарушения У обмена, обусловленное несовершенством
- 22. ОБМЕН БЕЛКОВ И ЕГО НАРУШЕНИЯ
- 23. БЕЛКИ играют центральную роль в процессах жизнедеятельности клеток и в формировании клеточных структур БЕЛКИ высокомолекулярные пептиды
- 24. Функции белков ферментативная (каталитическая) структурная (пластическая) регуляторная (эндокринная) рецепторная транспортная защитная сократительная специальная Передача генетической информации
- 25. Б, поступающие с пищей, подвергаются в ЖКТ распаду при участии протеолитических ферментов или пептидгидролаз, которые ускоряют
- 26. Ротовая полость во рту Б пищи только механически измельчаются Б пищи БЕЛКИ полипептиды и АК пепсин
- 27. ВСАСЫВАНИЕ АМИНОКИСЛОТ В КИШЕЧНИКЕ Пути использования аминокислот после их всасывания (участие в синтезе компонентов обмена некоторых
- 28. Промежуточный обмен аминокислот белки пищи белки тканей R — CH — COOH | NH2 NH3 биогенные
- 29. Катаболизм аминокислот белки пищи белки тканей R — CH — COOH | NH2 NH3 биогенные амины
- 30. NH3 СО2 Н2О ПЕЧЕНЬ мочевина экскреция глютамин глютамин ЖК Пуриновые основания NH3 Н+ глютамин NH4+ клубочек
- 31. Схема биосинтеза (трансляции) белка (А.С. Спирину) структурный ГЕН мРНК ДНК ЯДРО рибосома белок цитоплазма тРНК 75-80%
- 32. Гуаниловая кислота Инозиновая кислота Адениловая кислота 1 Рибозо-5-фосфат + АТФ 3 5-фосфорибозил-1-пирофосфат (ФРПФ) + глутамин 5-фосфорибозил-1-амин
- 33. ОСОБЕННОСТИ ОБМЕНА БЕЛКОВ У ДЕТЕЙ У новорожденных и детей 1 года жизни содержание общего Б в
- 34. НАРУШЕНИЯ ОБМЕНА БЕЛКОВ нарушения гидролиза и всасывания нарушения биосинтеза и распада нарушения общих и частных реакций
- 35. Нарушения переваривания и всасывания Б желудок Дефицит пепсина возникает при уменьшении секреции пепсиногена главными клетками слизистой,
- 36. нарушения биосинтеза и распада Б изменения генетического аппарата быстро распознаются специфическими ферментами – рестриктазами, измененные последовательности
- 37. вторичные ПЕЧЕНЬ Б гипопротеинемии диспротеинемии парапротеинемии недостаточность синтеза отдельных Б геморрагии, анемии, дистрофии, вторичные иммунодефициты ,
- 38. нарушения общих и частных реакций превращения АК нарушение транспорта структурно близких АК нарушение транспорта АК нарушение
- 39. нарушение трансаминирования АК Гиперфенилаланинемия нарушение превращения фенилаланина в тирозин обусловлено снижением активности ферментного комплекса фенилаланингидроксилазой прогрессирующие
- 40. нарушение окислительного дезаминирования АК при гипоксии, недостатке витаминов С, РР, В2, В6, поражениях печени, почек блокируется
- 41. нарушение биосинтеза мочевины нарушения конечных этапов метаболизма и выведения АК недостаточность карбомоилфосфатсинтетазы, катализирующей включение NH3 в
- 42. НАРУШЕНИЯ ПУРИНОВОГО ОБМЕНА Гиперурикемия состояние, проявляющееся повышением содержания мочевой кислоты в крови Причиной гиперурикемии может быть:
- 43. мочевая кислота плохо растворимое соединение, то при повышении концентрации происходит ее кристаллизация и накопление в суставах
- 44. ОСОБЕННОСТИ НАРУШЕНИЯ ОБМЕНА БЕЛКОВ У ДЕТЕЙ недостаток в течение длительного времени пищевых Б, богатых незаменимыми АК,
- 45. нарушения превращения АК биосинтеза Б нарушения, обусловленные мутацией гена, нередко сопровождаются появлением аномальных Б с необычными
- 46. ОБМЕН ЖИРОВ И ЕГО НАРУШЕНИЯ
- 47. совокупность процессов переваривания и всасывания нейтральных Ж (ТГЛ) и продуктов их распада в ЖКТ, промежуточного обмена
- 48. Трансформация липидов в эпителии тонкого кишечника ТГЛ – триглицериды ДГ – диглицериды МГ – моноглицериды СЖК-свободные
- 49. Метаболизм липидов печень (гепатоцит) ТГЛ ФЛ СИНТЕЗ ЛПОНП АПО В энтероцит кровеносные сосуды ХМ ТГЛ Глицерин
- 50. жировая ткань ТГЛ ТГЛ ХМ ЛПОНП ЛПЛ СЖК СЖК СЖК СЖК СЖК + АБ Глицерин +
- 51. ФУНКЦИОНАЛЬНАЯ РОЛЬ ХС 1. ХС является фактором краткосрочной адаптации клеток к изменениям окружающей водной среды путем
- 52. ГЛ пируват кетогенные АК ЖК АцКоА МХ: образование цитрата цикл Кребса ЖК ХС Цитозоль стероидные гормоны
- 53. Метаболизм ХС печень (гепатоцит) кровеносные сосуды ХМ ТГЛ Глицерин НЭЖК α-глицерофосфат АцКоА ЛППП ЛПНП ЛПНП ЛПНП
- 54. Регуляция жирового обмена а/д, на/д ГК, тироксин АКТГ, глюкагон ПГЕ1, ИНС, никотиновая кислота + + +
- 55. ОСОБЕННОСТИ ОБМЕНА ЖИРОВ У ДЕТЕЙ желудок Гидролитическое расщепление Ж в желудке играет важную роль в процессе
- 56. Одним из источников энергии является бурая жировая ткань. У новорожденного она составляет около 1% от массы
- 57. В детском возрасте синтез Ж из У идет наиболее интенсивно. Преимущественно У питание очень быстро приводит
- 58. НАРУШЕНИЯ ОБМЕНА ЖИРОВ нарушения гидролиза и всасывания нарушения транспорта и распада
- 59. Нарушения переваривания и всасывания Ж желудок ПЖ липаза недостаточное всасывание Ж в тонкой кишке может быть
- 60. нарушения транспорта ХМ ЛИМФА (d.thoracicus) Повышенное содержание ЛП – гиперлипопротеинемия ЛП ЛПЛ ГИ Гиперлипопротеинемия энтероцит ТГЛ
- 61. ЛПЛ нарушения одиночного гена многофакторные нарушения Первичные Семейная недостаточность липопротеинлипазы ХМ ЭХС ТГЛ Семейная гиперлипопротеинемия 3
- 62. ТГЛ ИНС ИР ТГЛ Вторичные Сахарный диабет Злоупотребление алкоголем ИНС ЛПОНП ХМ кетоновые тела СЖК НЭЖК
- 63. Гиполипопротеинемия желчные килоты липаза ЛП ХМ ЛПОНП ТГЛ Пониженное содержание ЛП – гипопротеинемия тироксин генетические нарушения
- 65. Скачать презентацию
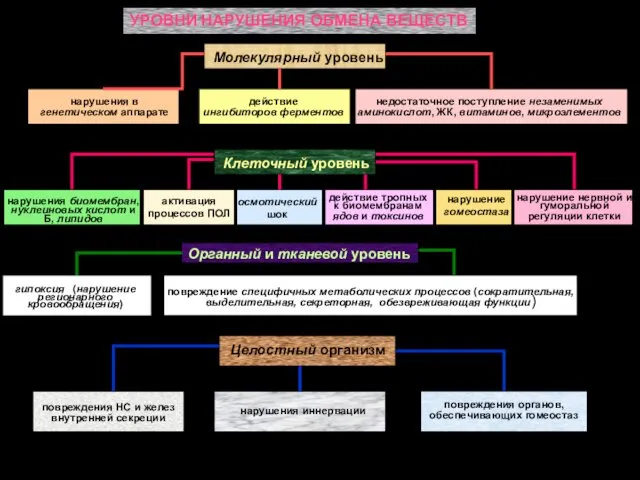
УРОВНИ НАРУШЕНИЯ ОБМЕНА ВЕЩЕСТВ
Молекулярный уровень
нарушения в генетическом аппарате
действие ингибиторов
УРОВНИ НАРУШЕНИЯ ОБМЕНА ВЕЩЕСТВ
Молекулярный уровень
нарушения в генетическом аппарате
действие ингибиторов

ОБМЕН УГЛЕВОДОВ И ЕГО НАРУШЕНИЯ
ОБМЕН УГЛЕВОДОВ И ЕГО НАРУШЕНИЯ
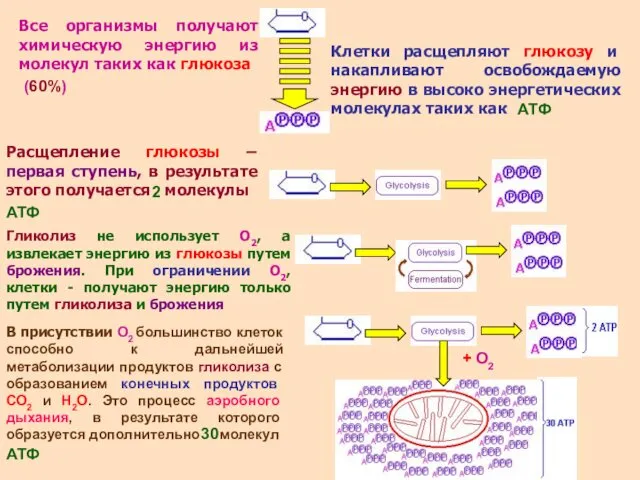
Все организмы получают химическую энергию из молекул таких как глюкоза
Клетки расщепляют
Все организмы получают химическую энергию из молекул таких как глюкоза
Клетки расщепляют

Наиболее важным углеводом крови является ГЛ
У составляют не более 2%
Наиболее важным углеводом крови является ГЛ
У составляют не более 2%
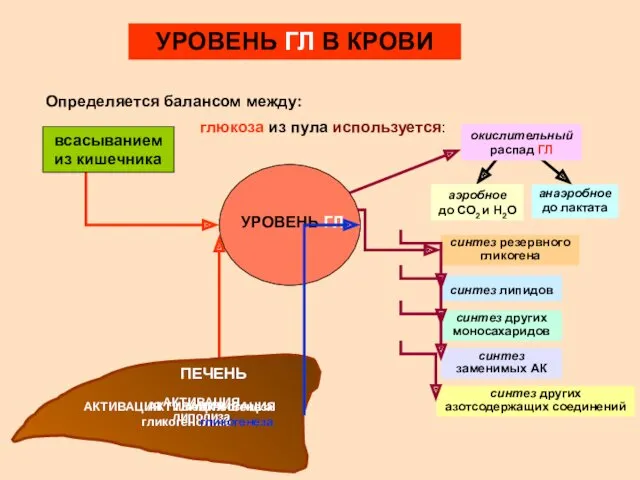
УРОВЕНЬ ГЛ В КРОВИ
Определяется балансом между:
УРОВЕНЬ ГЛ
всасыванием из кишечника
АКТИВАЦИЯ глюконеогенеза
АКТИВАЦИЯ
УРОВЕНЬ ГЛ В КРОВИ
Определяется балансом между:
УРОВЕНЬ ГЛ
всасыванием из кишечника
АКТИВАЦИЯ глюконеогенеза
АКТИВАЦИЯ

Усвоение углеводов
основным источником У для человека являются У пищи:
крахмал
Усвоение углеводов
основным источником У для человека являются У пищи:
крахмал
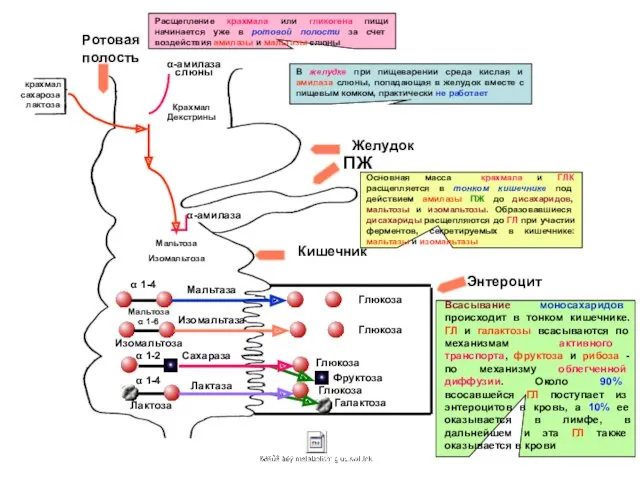
крахмал
сахароза
лактоза
α-амилаза слюны
Крахмал
Декстрины
Желудок
Ротовая полость
ПЖ
Кишечник
Энтероцит
Расщепление крахмала или гликогена пищи начинается уже в ротовой
крахмал
сахароза
лактоза
α-амилаза слюны
Крахмал
Декстрины
Желудок
Ротовая полость
ПЖ
Кишечник
Энтероцит
Расщепление крахмала или гликогена пищи начинается уже в ротовой
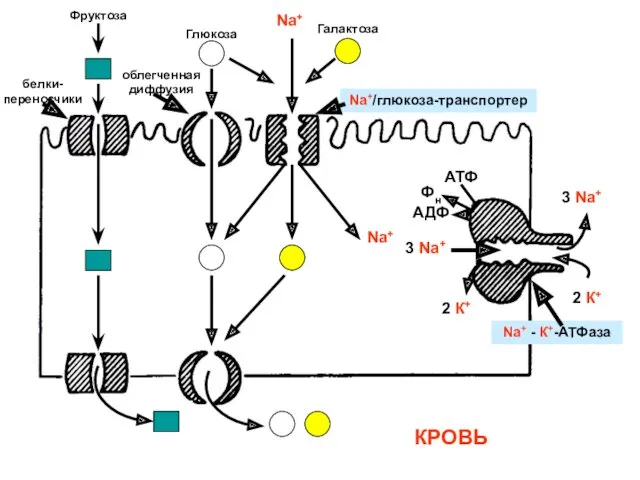
Фруктоза
Глюкоза
Na+
Галактоза
Na+
3 Na+
2 К+
3 Na+
2 К+
АТФ
Фн
АДФ
Na+ - К+-АТФаза
Na+/глюкоза-транспортер
белки-переносчики
КРОВЬ
облегченная диффузия
Фруктоза
Глюкоза
Na+
Галактоза
Na+
3 Na+
2 К+
3 Na+
2 К+
АТФ
Фн
АДФ
Na+ - К+-АТФаза
Na+/глюкоза-транспортер
белки-переносчики
КРОВЬ
облегченная диффузия
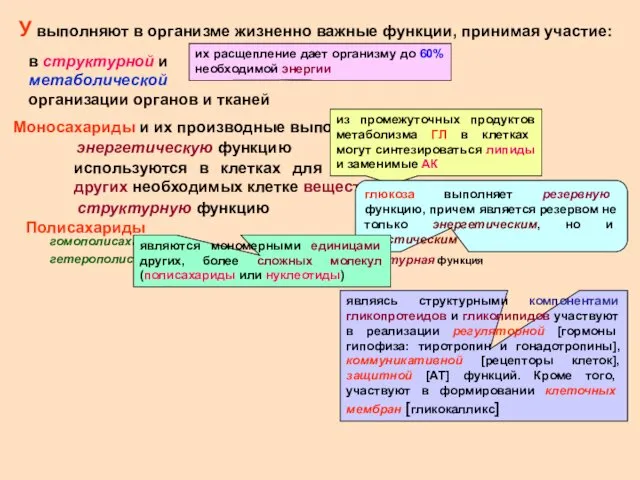
У выполняют в организме жизненно важные функции, принимая участие:
в структурной и
У выполняют в организме жизненно важные функции, принимая участие:
в структурной и
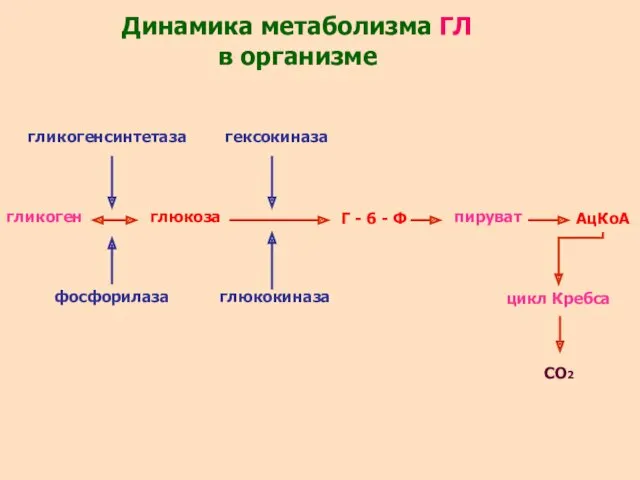
гликоген
гликогенсинтетаза
глюкоза
гексокиназа
глюкокиназа
Г - 6 - Ф
пируват
АцКоА
цикл Кребса
СО2
Динамика метаболизма ГЛ в организме
фосфорилаза
гликоген
гликогенсинтетаза
глюкоза
гексокиназа
глюкокиназа
Г - 6 - Ф
пируват
АцКоА
цикл Кребса
СО2
Динамика метаболизма ГЛ в организме
фосфорилаза
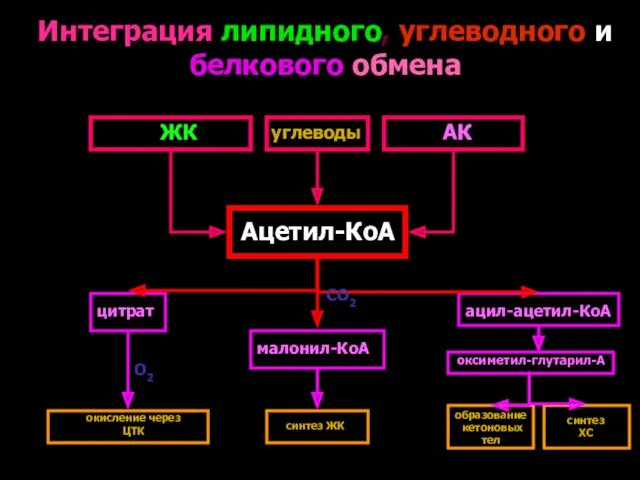
Интеграция липидного, углеводного и белкового обмена
Ацетил-КоА
ЖК
углеводы
АК
цитрат
малонил-КоА
ацил-ацетил-КоА
СО2
окисление через ЦТК
синтез ЖК
образование
кетоновых
тел
синтез
ХС
О2
оксиметил-глутарил-А
Интеграция липидного, углеводного и белкового обмена
Ацетил-КоА
ЖК
углеводы
АК
цитрат
малонил-КоА
ацил-ацетил-КоА
СО2
окисление через ЦТК
синтез ЖК
образование
кетоновых
тел
синтез
ХС
О2
оксиметил-глутарил-А

Основным клиническим биохимическим показателем нарушений У обмена является изменение концентрации в
Основным клиническим биохимическим показателем нарушений У обмена является изменение концентрации в

ОСОБЕННОСТИ ОБМЕНА УГЛЕВОДОВ У ДЕТЕЙ
За счет У у грудных детей покрывается
ОСОБЕННОСТИ ОБМЕНА УГЛЕВОДОВ У ДЕТЕЙ
За счет У у грудных детей покрывается
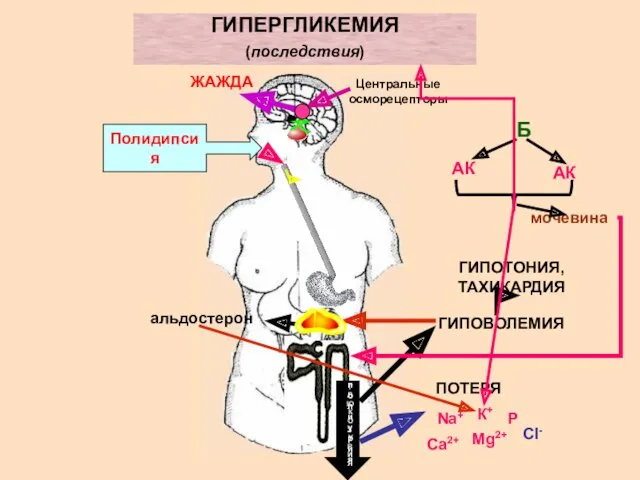
ГИПЕРГЛИКЕМИЯ
(последствия)
Центральные осморецепторы
ЖАЖДА
Полидипсия
полиурия
ГЛЮКОЗУРИЯ
ПОТЕРЯ
Na+
К+
Mg2+
Р
Сa2+
Cl-
ГИПОВОЛЕМИЯ
ГИПОТОНИЯ, ТАХИКАРДИЯ
альдостерон
Б
АК
АК
мочевина
ГИПЕРГЛИКЕМИЯ
(последствия)
Центральные осморецепторы
ЖАЖДА
Полидипсия
полиурия
ГЛЮКОЗУРИЯ
ПОТЕРЯ
Na+
К+
Mg2+
Р
Сa2+
Cl-
ГИПОВОЛЕМИЯ
ГИПОТОНИЯ, ТАХИКАРДИЯ
альдостерон
Б
АК
АК
мочевина
Н2О
С
Ф
ГЛ
Н2О
С
Ф
ГЛ
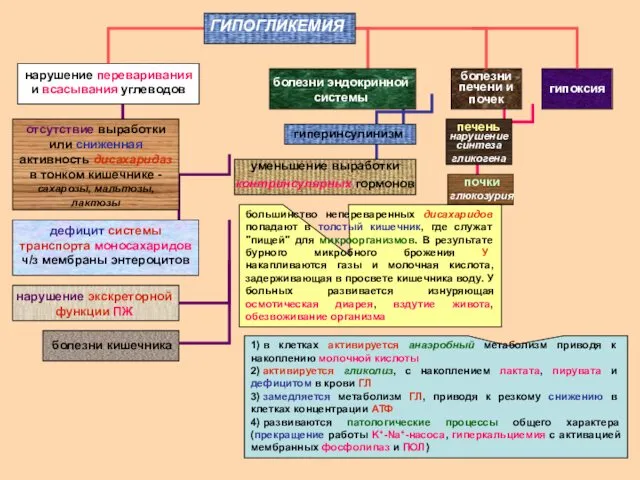
нарушение переваривания и всасывания углеводов
нарушение экскреторной функции ПЖ
отсутствие выработки или
нарушение переваривания и всасывания углеводов
нарушение экскреторной функции ПЖ
отсутствие выработки или
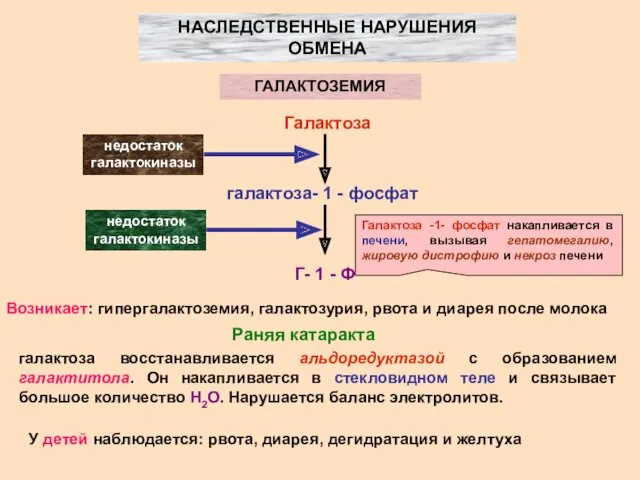
НАСЛЕДСТВЕННЫЕ НАРУШЕНИЯ ОБМЕНА
ГАЛАКТОЗЕМИЯ
Галактоза
галактоза- 1 - фосфат
Г- 1 - Ф
недостаток галактокиназы
недостаток галактокиназы
Возникает:
НАСЛЕДСТВЕННЫЕ НАРУШЕНИЯ ОБМЕНА
ГАЛАКТОЗЕМИЯ
Галактоза
галактоза- 1 - фосфат
Г- 1 - Ф
недостаток галактокиназы
недостаток галактокиназы
Возникает:
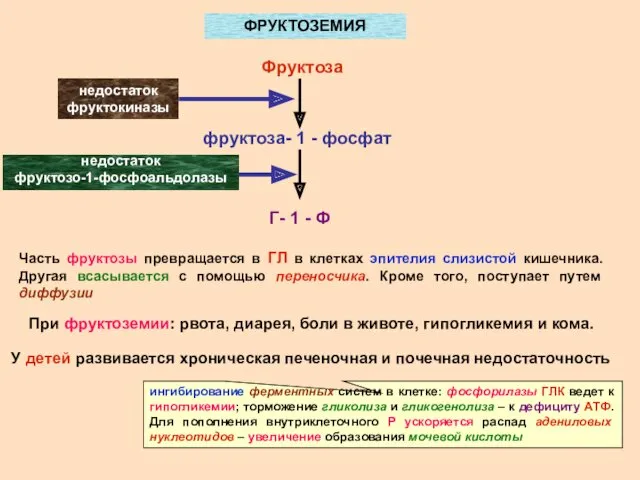
ФРУКТОЗЕМИЯ
Фруктоза
фруктоза- 1 - фосфат
Г- 1 - Ф
недостаток фруктокиназы
недостаток
фруктозо-1-фосфоальдолазы
Часть фруктозы превращается
ФРУКТОЗЕМИЯ
Фруктоза
фруктоза- 1 - фосфат
Г- 1 - Ф
недостаток фруктокиназы
недостаток
фруктозо-1-фосфоальдолазы
Часть фруктозы превращается

ГЛИКОГЕНОЗЫ (болезни накопления)
нарушение ферментативного распада ГЛК печени и мышц из-за
ГЛИКОГЕНОЗЫ (болезни накопления)
нарушение ферментативного распада ГЛК печени и мышц из-за

ОСОБЕННОСТИ НАРУШЕНИЯ ОБМЕНА УГЛЕВОДОВ У ДЕТЕЙ
одно из наиболее частых проявлений нарушения
ОСОБЕННОСТИ НАРУШЕНИЯ ОБМЕНА УГЛЕВОДОВ У ДЕТЕЙ
одно из наиболее частых проявлений нарушения

ОБМЕН БЕЛКОВ И ЕГО НАРУШЕНИЯ
ОБМЕН БЕЛКОВ И ЕГО НАРУШЕНИЯ
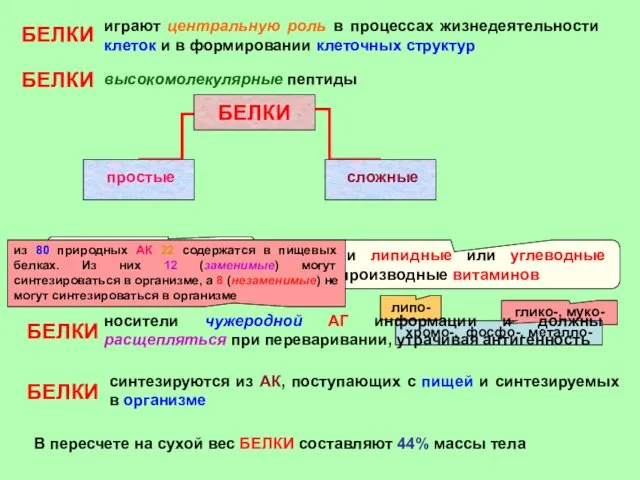
БЕЛКИ
играют центральную роль в процессах жизнедеятельности клеток и в формировании клеточных
БЕЛКИ
играют центральную роль в процессах жизнедеятельности клеток и в формировании клеточных

Функции белков
ферментативная (каталитическая)
структурная (пластическая)
регуляторная (эндокринная)
рецепторная
транспортная
защитная
сократительная
Функции белков
ферментативная (каталитическая)
структурная (пластическая)
регуляторная (эндокринная)
рецепторная
транспортная
защитная
сократительная

Б, поступающие с пищей, подвергаются в ЖКТ распаду при участии протеолитических
Б, поступающие с пищей, подвергаются в ЖКТ распаду при участии протеолитических
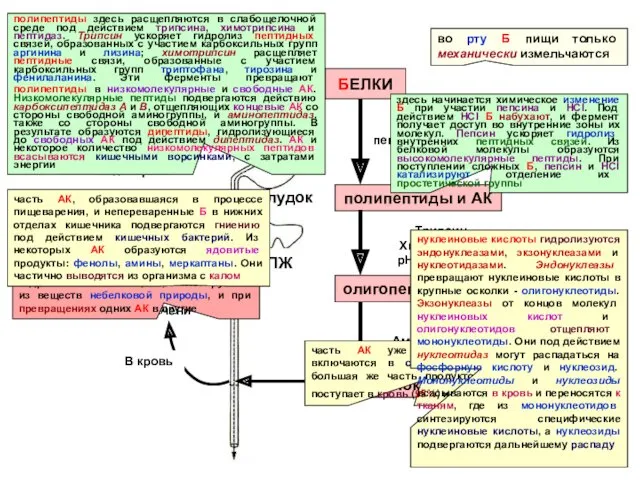
Ротовая полость
во рту Б пищи только механически измельчаются
Б пищи
БЕЛКИ
полипептиды и АК
пепсин
Ротовая полость
во рту Б пищи только механически измельчаются
Б пищи
БЕЛКИ
полипептиды и АК
пепсин
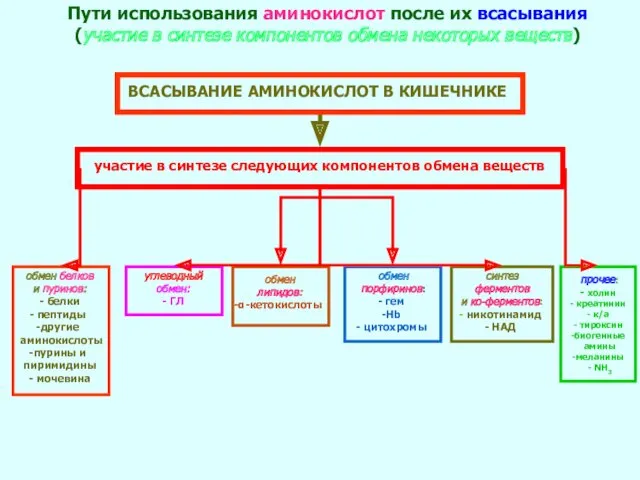
ВСАСЫВАНИЕ АМИНОКИСЛОТ В КИШЕЧНИКЕ
Пути использования аминокислот после их всасывания (участие в
ВСАСЫВАНИЕ АМИНОКИСЛОТ В КИШЕЧНИКЕ
Пути использования аминокислот после их всасывания (участие в
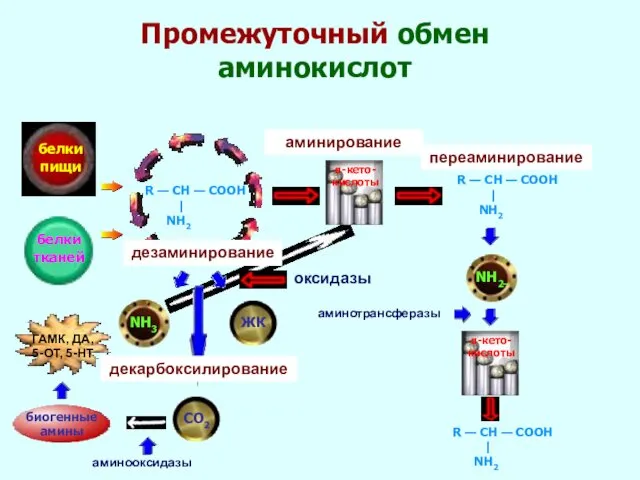
Промежуточный обмен аминокислот
белки
пищи
белки
тканей
R — CH — COOH
|
NH2
NH3
биогенные
амины
СО2
α-кето-
кислоты
ЖК
оксидазы
аминирование
R — CH
Промежуточный обмен аминокислот
белки
пищи
белки
тканей
R — CH — COOH
|
NH2
NH3
биогенные
амины
СО2
α-кето-
кислоты
ЖК
оксидазы
аминирование
R — CH
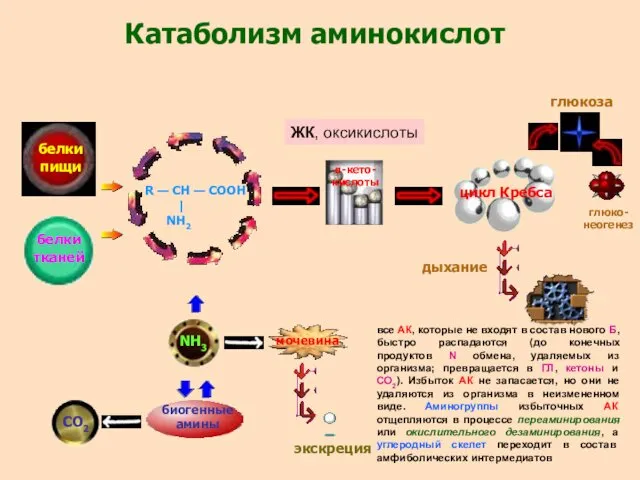
Катаболизм аминокислот
белки
пищи
белки
тканей
R — CH — COOH
|
NH2
NH3
биогенные
амины
СО2
мочевина
экскреция
α-кето-
кислоты
цикл Кребса
глюкоза
глюко-
неогенез
дыхание
ЖК, оксикислоты
все АК,
Катаболизм аминокислот
белки
пищи
белки
тканей
R — CH — COOH
|
NH2
NH3
биогенные
амины
СО2
мочевина
экскреция
α-кето-
кислоты
цикл Кребса
глюкоза
глюко-
неогенез
дыхание
ЖК, оксикислоты
все АК,

NH3
СО2
Н2О
ПЕЧЕНЬ
мочевина
экскреция
глютамин
глютамин
ЖК
Пуриновые основания
NH3
Н+
глютамин
NH4+
клубочек
глютаминаза
СО2
СО2
Н2О
Н2О
85%
5%
3%
амидирование
цитруллин-аргинино-орнитиновый цикл
NH3
СО2
Н2О
ПЕЧЕНЬ
мочевина
экскреция
глютамин
глютамин
ЖК
Пуриновые основания
NH3
Н+
глютамин
NH4+
клубочек
глютаминаза
СО2
СО2
Н2О
Н2О
85%
5%
3%
амидирование
цитруллин-аргинино-орнитиновый цикл
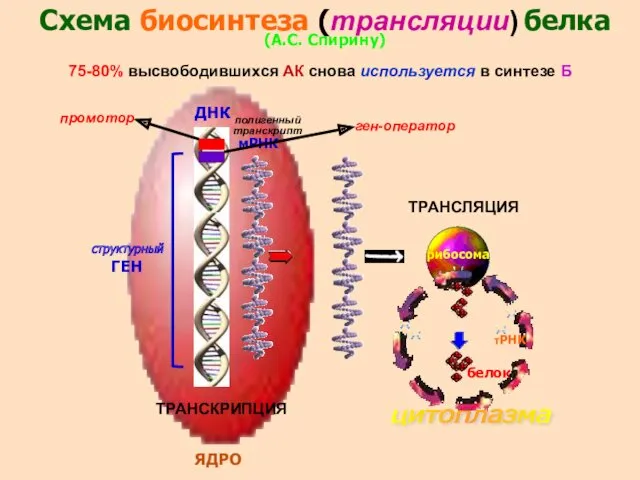
Схема биосинтеза (трансляции) белка
(А.С. Спирину)
структурный ГЕН
мРНК
ДНК
ЯДРО
рибосома
белок
цитоплазма
тРНК
75-80% высвободившихся АК снова используется в
Схема биосинтеза (трансляции) белка
(А.С. Спирину)
структурный ГЕН
мРНК
ДНК
ЯДРО
рибосома
белок
цитоплазма
тРНК
75-80% высвободившихся АК снова используется в
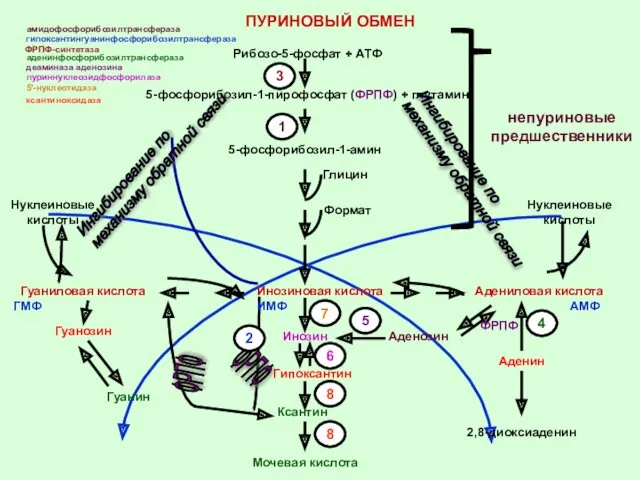
Гуаниловая кислота
Инозиновая кислота
Адениловая кислота
1
Рибозо-5-фосфат + АТФ
3
5-фосфорибозил-1-пирофосфат (ФРПФ) + глутамин
5-фосфорибозил-1-амин
Глицин
Формат
Гуанозин
Гуанин
Нуклеиновые кислоты
ФРПФ
ФРПФ
2
ИМФ
7
Инозин
Гипоксантин
6
8
Ксантин
8
Мочевая кислота
Аденозин
ФРПФ
4
Аденин
2,8-диоксиаденин
АМФ
Нуклеиновые
Гуаниловая кислота
Инозиновая кислота
Адениловая кислота
1
Рибозо-5-фосфат + АТФ
3
5-фосфорибозил-1-пирофосфат (ФРПФ) + глутамин
5-фосфорибозил-1-амин
Глицин
Формат
Гуанозин
Гуанин
Нуклеиновые кислоты
ФРПФ
ФРПФ
2
ИМФ
7
Инозин
Гипоксантин
6
8
Ксантин
8
Мочевая кислота
Аденозин
ФРПФ
4
Аденин
2,8-диоксиаденин
АМФ
Нуклеиновые

ОСОБЕННОСТИ ОБМЕНА БЕЛКОВ У ДЕТЕЙ
У новорожденных и детей 1 года жизни
ОСОБЕННОСТИ ОБМЕНА БЕЛКОВ У ДЕТЕЙ
У новорожденных и детей 1 года жизни
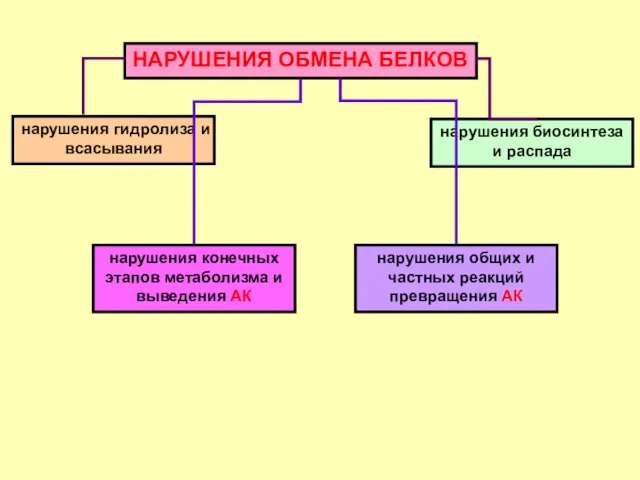
НАРУШЕНИЯ ОБМЕНА БЕЛКОВ
нарушения гидролиза и всасывания
нарушения биосинтеза и распада
нарушения общих и
НАРУШЕНИЯ ОБМЕНА БЕЛКОВ
нарушения гидролиза и всасывания
нарушения биосинтеза и распада
нарушения общих и
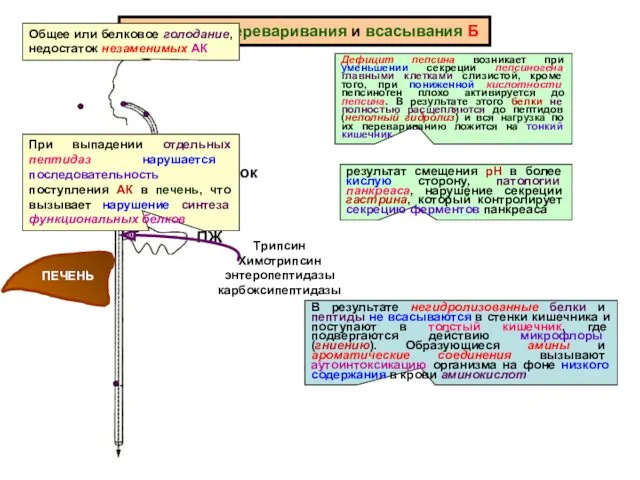
Нарушения переваривания и всасывания Б
желудок
Дефицит пепсина возникает при уменьшении секреции
Нарушения переваривания и всасывания Б
желудок
Дефицит пепсина возникает при уменьшении секреции
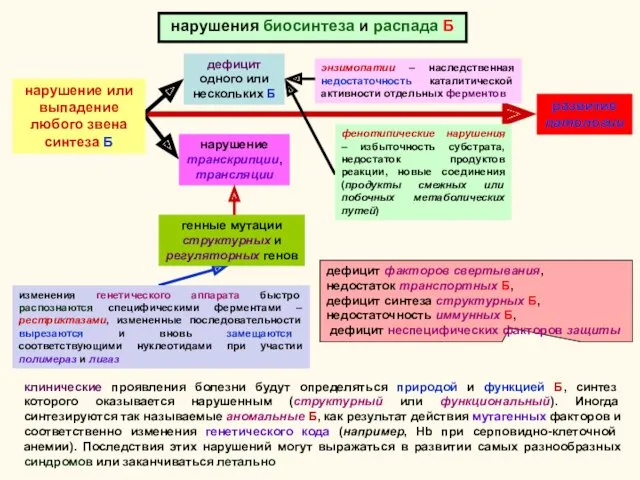
нарушения биосинтеза и распада Б
изменения генетического аппарата быстро распознаются специфическими ферментами
нарушения биосинтеза и распада Б
изменения генетического аппарата быстро распознаются специфическими ферментами
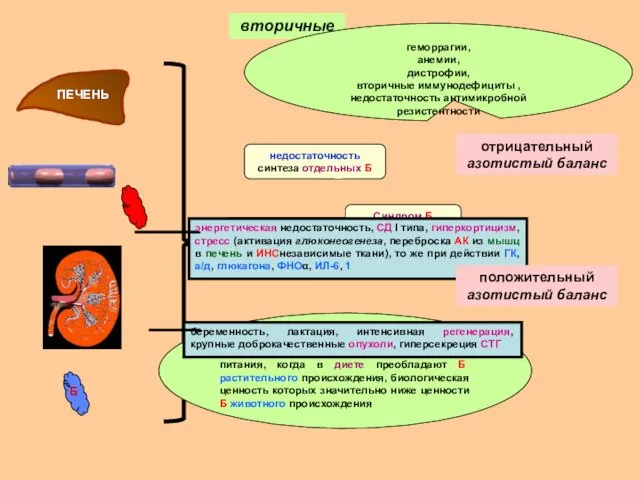
вторичные
ПЕЧЕНЬ
Б
гипопротеинемии
диспротеинемии
парапротеинемии
недостаточность синтеза отдельных Б
геморрагии,
анемии,
дистрофии,
вторичные иммунодефициты ,
недостаточность антимикробной резистентности
Синдром Б недостаточности
развивается
вторичные
ПЕЧЕНЬ
Б
гипопротеинемии
диспротеинемии
парапротеинемии
недостаточность синтеза отдельных Б
геморрагии,
анемии,
дистрофии,
вторичные иммунодефициты ,
недостаточность антимикробной резистентности
Синдром Б недостаточности
развивается
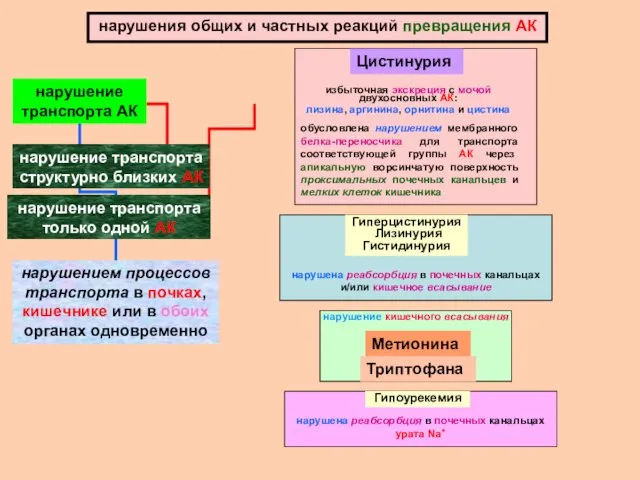
нарушения общих и частных реакций превращения АК
нарушение транспорта структурно близких АК
нарушения общих и частных реакций превращения АК
нарушение транспорта структурно близких АК
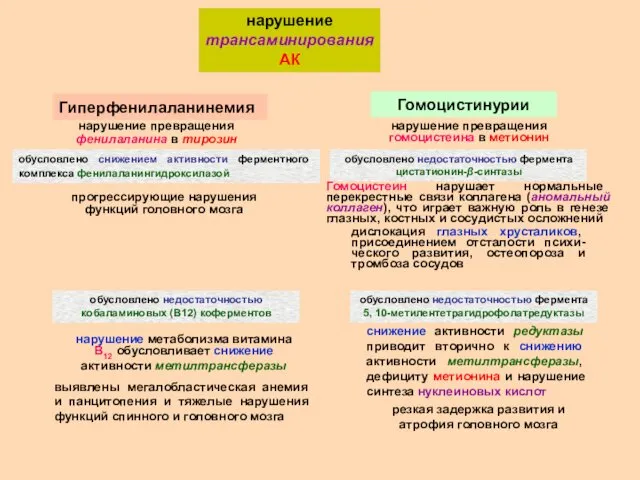
нарушение трансаминирования АК
Гиперфенилаланинемия
нарушение превращения фенилаланина в тирозин
обусловлено снижением активности
нарушение трансаминирования АК
Гиперфенилаланинемия
нарушение превращения фенилаланина в тирозин
обусловлено снижением активности

нарушение окислительного дезаминирования АК
при гипоксии, недостатке витаминов С, РР, В2, В6,
нарушение окислительного дезаминирования АК
при гипоксии, недостатке витаминов С, РР, В2, В6,
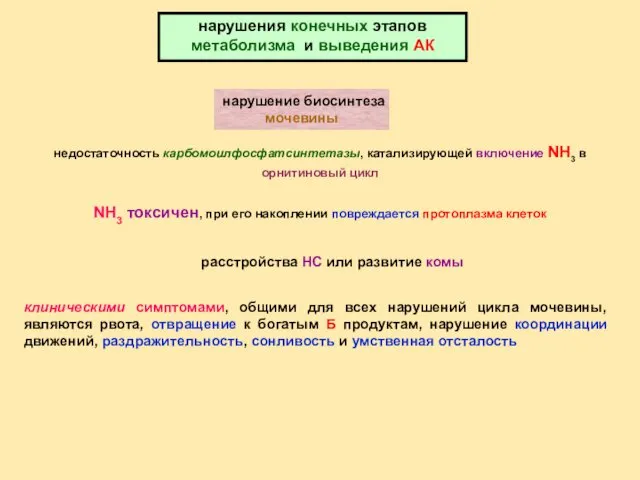
нарушение биосинтеза мочевины
нарушения конечных этапов метаболизма и выведения АК
недостаточность карбомоилфосфатсинтетазы,
нарушение биосинтеза мочевины
нарушения конечных этапов метаболизма и выведения АК
недостаточность карбомоилфосфатсинтетазы,

НАРУШЕНИЯ ПУРИНОВОГО ОБМЕНА
Гиперурикемия
состояние, проявляющееся повышением содержания мочевой кислоты в крови
НАРУШЕНИЯ ПУРИНОВОГО ОБМЕНА
Гиперурикемия
состояние, проявляющееся повышением содержания мочевой кислоты в крови

мочевая кислота плохо растворимое соединение, то при повышении концентрации происходит ее
мочевая кислота плохо растворимое соединение, то при повышении концентрации происходит ее
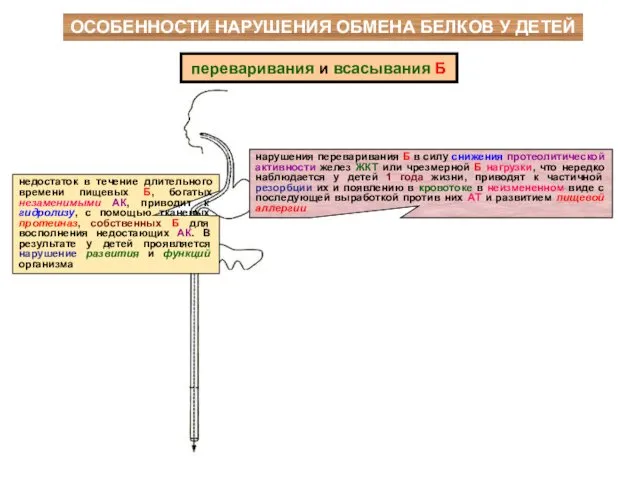
ОСОБЕННОСТИ НАРУШЕНИЯ ОБМЕНА БЕЛКОВ У ДЕТЕЙ
недостаток в течение длительного времени пищевых
ОСОБЕННОСТИ НАРУШЕНИЯ ОБМЕНА БЕЛКОВ У ДЕТЕЙ
недостаток в течение длительного времени пищевых

нарушения превращения АК
биосинтеза Б
нарушения, обусловленные мутацией гена, нередко сопровождаются появлением аномальных
нарушения превращения АК
биосинтеза Б
нарушения, обусловленные мутацией гена, нередко сопровождаются появлением аномальных

ОБМЕН ЖИРОВ И ЕГО НАРУШЕНИЯ
ОБМЕН ЖИРОВ И ЕГО НАРУШЕНИЯ
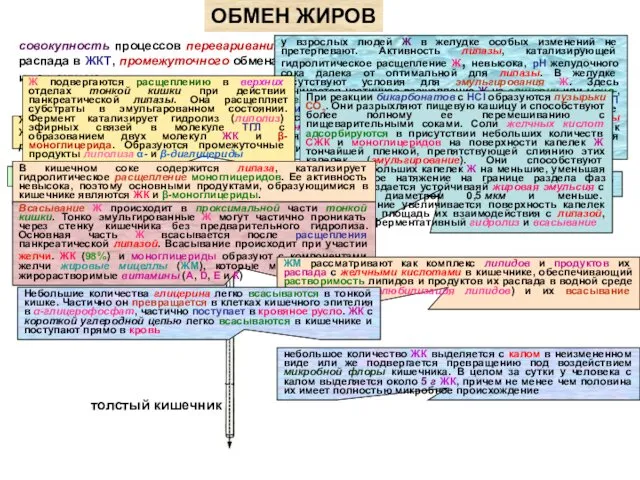
совокупность процессов переваривания и всасывания нейтральных Ж (ТГЛ) и продуктов их
совокупность процессов переваривания и всасывания нейтральных Ж (ТГЛ) и продуктов их
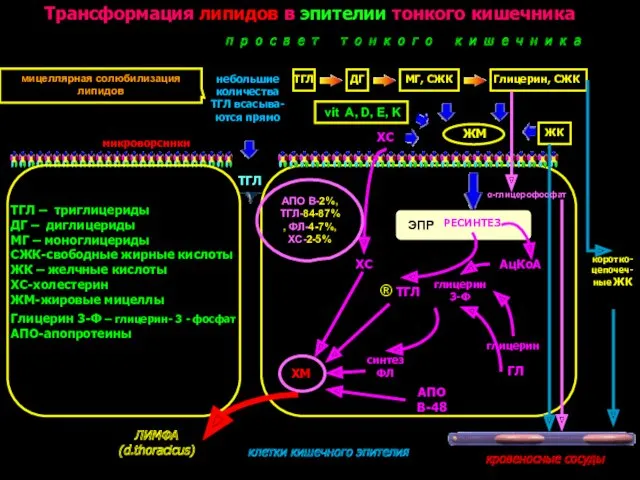
Трансформация липидов в эпителии тонкого кишечника
ТГЛ – триглицериды
ДГ – диглицериды
МГ
Трансформация липидов в эпителии тонкого кишечника
ТГЛ – триглицериды
ДГ – диглицериды
МГ
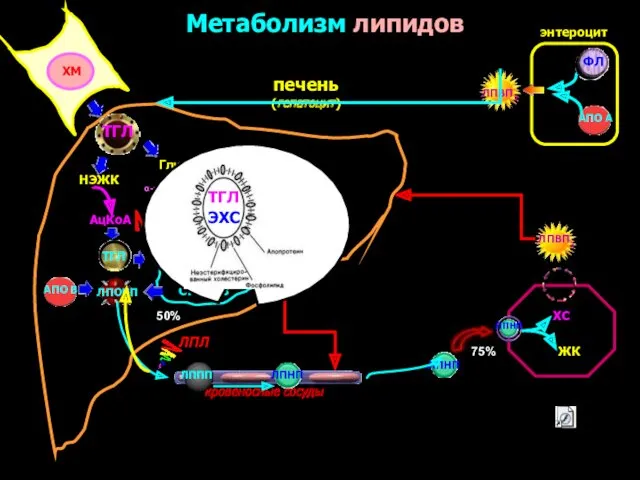
Метаболизм липидов
печень
(гепатоцит)
ТГЛ
ФЛ
СИНТЕЗ
ЛПОНП
АПО В
энтероцит
кровеносные сосуды
ХМ
ТГЛ
Глицерин
НЭЖК
α-глицерофосфат
АцКоА
ФЛ
АПО А
ЛПВП2
ЛППП
ЛПНП
ЛПНП
ЛПНП
ЖК
ХС
ЛПВП3
ГЛ
ТГЛ
ЭХС
ЛПЛ
50%
75%
Метаболизм липидов
печень
(гепатоцит)
ТГЛ
ФЛ
СИНТЕЗ
ЛПОНП
АПО В
энтероцит
кровеносные сосуды
ХМ
ТГЛ
Глицерин
НЭЖК
α-глицерофосфат
АцКоА
ФЛ
АПО А
ЛПВП2
ЛППП
ЛПНП
ЛПНП
ЛПНП
ЖК
ХС
ЛПВП3
ГЛ
ТГЛ
ЭХС
ЛПЛ
50%
75%
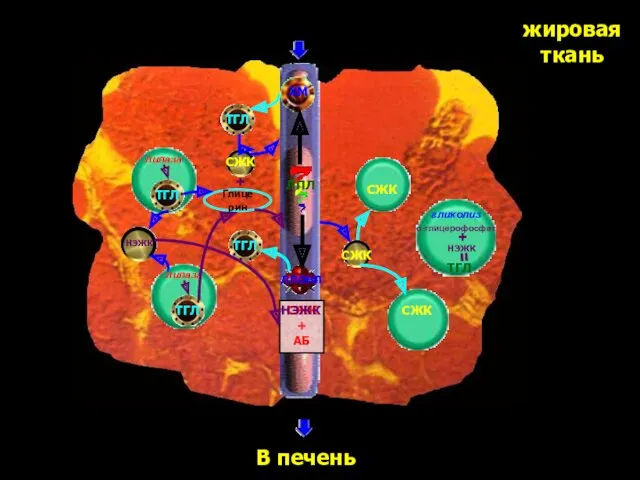
жировая
ткань
ТГЛ
ТГЛ
ХМ
ЛПОНП
ЛПЛ
СЖК
СЖК
СЖК
СЖК
СЖК
+
АБ
Глицерин
+
В печень
НЭЖК
ТГЛ
липаза
ТГЛ
липаза
α-глицерофосфат
гликолиз
+
НЭЖК
ТГЛ
=
НЭЖК
жировая
ткань
ТГЛ
ТГЛ
ХМ
ЛПОНП
ЛПЛ
СЖК
СЖК
СЖК
СЖК
СЖК
+
АБ
Глицерин
+
В печень
НЭЖК
ТГЛ
липаза
ТГЛ
липаза
α-глицерофосфат
гликолиз
+
НЭЖК
ТГЛ
=
НЭЖК

ФУНКЦИОНАЛЬНАЯ РОЛЬ ХС
1. ХС является фактором краткосрочной адаптации клеток к
ФУНКЦИОНАЛЬНАЯ РОЛЬ ХС
1. ХС является фактором краткосрочной адаптации клеток к
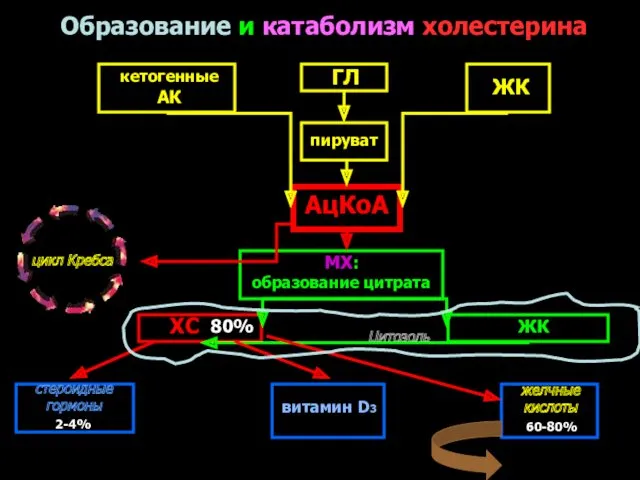
ГЛ
пируват
кетогенные
АК
ЖК
АцКоА
МХ:
образование цитрата
цикл Кребса
ЖК
ХС
Цитозоль
стероидные
гормоны
витамин D3
60-80%
Образование и катаболизм холестерина
80%
желчные
кислоты
2-4%
ГЛ
пируват
кетогенные
АК
ЖК
АцКоА
МХ:
образование цитрата
цикл Кребса
ЖК
ХС
Цитозоль
стероидные
гормоны
витамин D3
60-80%
Образование и катаболизм холестерина
80%
желчные
кислоты
2-4%
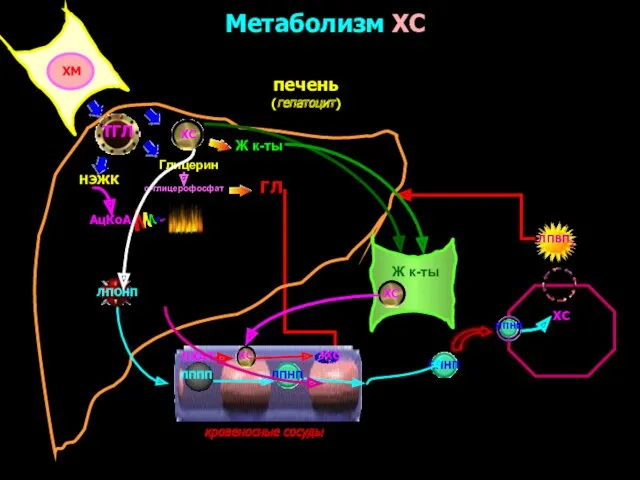
Метаболизм ХС
печень
(гепатоцит)
кровеносные сосуды
ХМ
ТГЛ
Глицерин
НЭЖК
α-глицерофосфат
АцКоА
ЛППП
ЛПНП
ЛПНП
ЛПНП
ХС
ЛПВП3
ГЛ
ХС
Ж к-ты
ЛПОНП
Ж к-ты
ХС
ХС
ЛХАТ
ЭХС
Метаболизм ХС
печень
(гепатоцит)
кровеносные сосуды
ХМ
ТГЛ
Глицерин
НЭЖК
α-глицерофосфат
АцКоА
ЛППП
ЛПНП
ЛПНП
ЛПНП
ХС
ЛПВП3
ГЛ
ХС
Ж к-ты
ЛПОНП
Ж к-ты
ХС
ХС
ЛХАТ
ЭХС

Регуляция жирового обмена
а/д, на/д
ГК, тироксин
АКТГ,
глюкагон
ПГЕ1, ИНС,
никотиновая
кислота
+
+
+
_
мембрана клетки
АЦ
АЦ
АТФ
цАМФ
5`АМФ
фосфодиэстераза
+
неактивная
ТГЛ-
липаза
активная
ТГЛ-
липаза
фосфатаза
АТФ
АДФ
киназа
+
ТГЛ
глицерин +
моноглицерид +
диглицерид +
НЭЖК
(в кровотоке)
гликолиз
+
_
Регуляция жирового обмена
а/д, на/д
ГК, тироксин
АКТГ,
глюкагон
ПГЕ1, ИНС,
никотиновая
кислота
+
+
+
_
мембрана клетки
АЦ
АЦ
АТФ
цАМФ
5`АМФ
фосфодиэстераза
+
неактивная
ТГЛ-
липаза
активная
ТГЛ-
липаза
фосфатаза
АТФ
АДФ
киназа
+
ТГЛ
глицерин +
моноглицерид +
диглицерид +
НЭЖК
(в кровотоке)
гликолиз
+
_
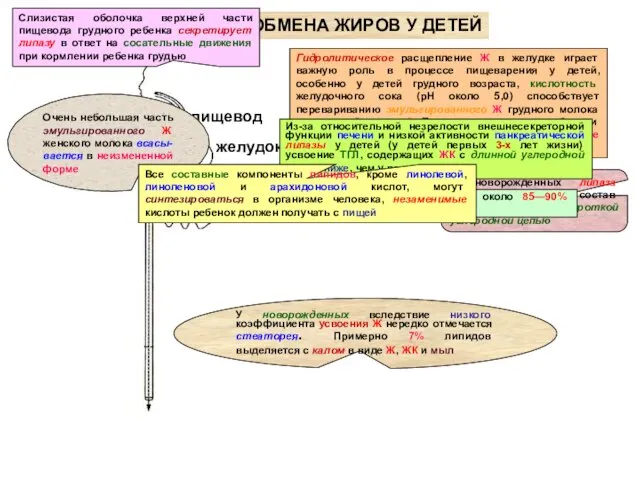
ОСОБЕННОСТИ ОБМЕНА ЖИРОВ У ДЕТЕЙ
желудок
Гидролитическое расщепление Ж в желудке играет важную
ОСОБЕННОСТИ ОБМЕНА ЖИРОВ У ДЕТЕЙ
желудок
Гидролитическое расщепление Ж в желудке играет важную

Одним из источников энергии является бурая жировая ткань. У новорожденного
Одним из источников энергии является бурая жировая ткань. У новорожденного

В детском возрасте синтез Ж из У идет наиболее интенсивно. Преимущественно
В детском возрасте синтез Ж из У идет наиболее интенсивно. Преимущественно
НАРУШЕНИЯ ОБМЕНА ЖИРОВ
нарушения гидролиза и всасывания
нарушения транспорта и распада
НАРУШЕНИЯ ОБМЕНА ЖИРОВ
нарушения гидролиза и всасывания
нарушения транспорта и распада
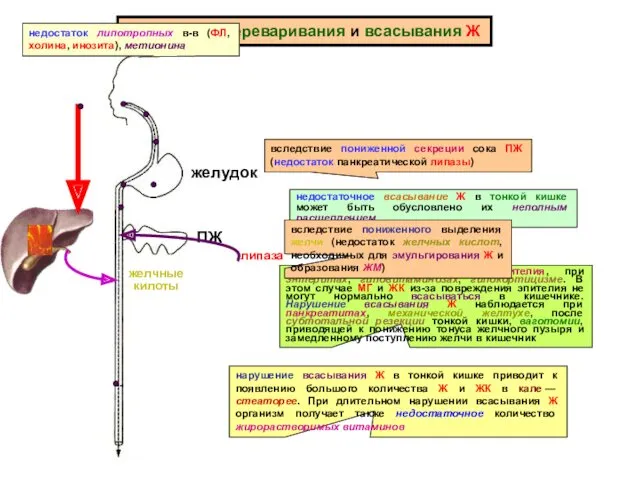
Нарушения переваривания и всасывания Ж
желудок
ПЖ
липаза
недостаточное всасывание Ж в тонкой
Нарушения переваривания и всасывания Ж
желудок
ПЖ
липаза
недостаточное всасывание Ж в тонкой
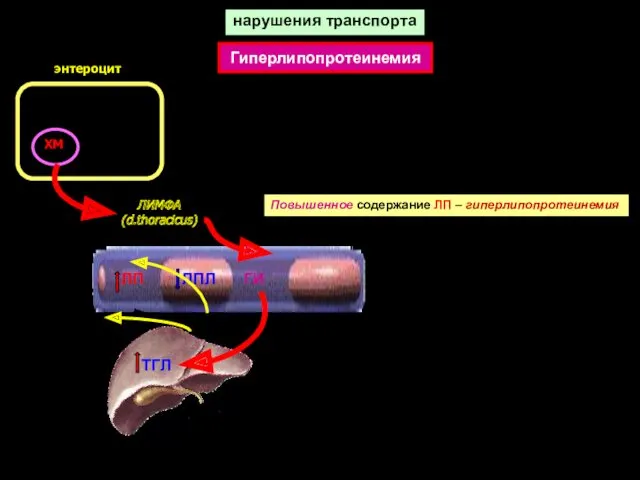
нарушения транспорта
ХМ
ЛИМФА
(d.thoracicus)
Повышенное содержание ЛП – гиперлипопротеинемия
ЛП
ЛПЛ
ГИ
Гиперлипопротеинемия
энтероцит
ТГЛ
нарушения транспорта
ХМ
ЛИМФА
(d.thoracicus)
Повышенное содержание ЛП – гиперлипопротеинемия
ЛП
ЛПЛ
ГИ
Гиперлипопротеинемия
энтероцит
ТГЛ
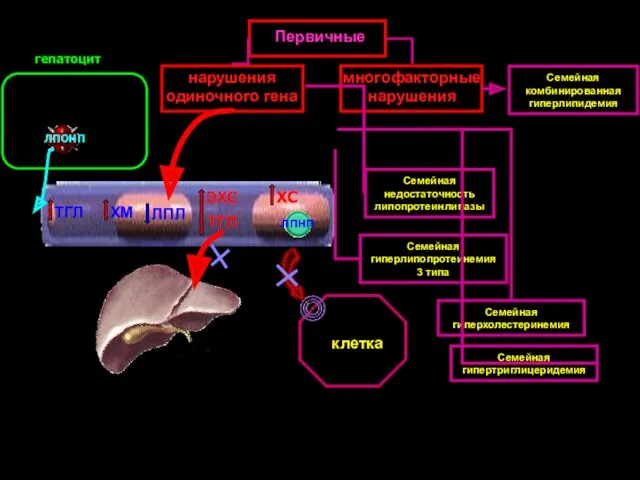
ЛПЛ
нарушения одиночного гена
многофакторные нарушения
Первичные
Семейная недостаточность липопротеинлипазы
ХМ
ЭХС
ТГЛ
Семейная гиперлипопротеинемия 3
ЛПЛ
нарушения одиночного гена
многофакторные нарушения
Первичные
Семейная недостаточность липопротеинлипазы
ХМ
ЭХС
ТГЛ
Семейная гиперлипопротеинемия 3
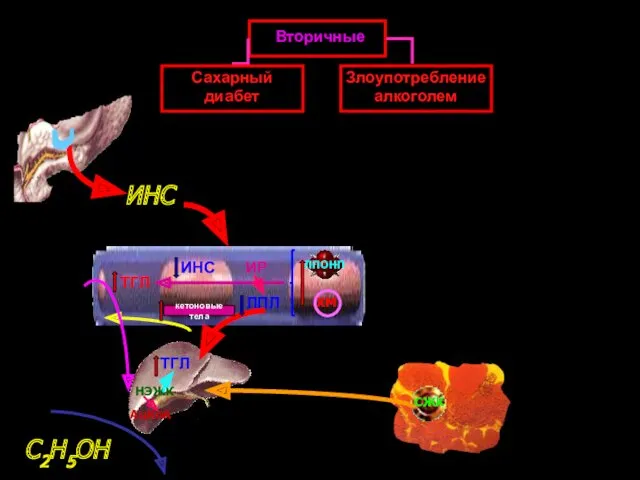
ТГЛ
ИНС
ИР
ТГЛ
Вторичные
Сахарный диабет
Злоупотребление алкоголем
ИНС
ЛПОНП
ХМ
кетоновые тела
СЖК
НЭЖК
АцКоА
С2Н5ОН
ЛПЛ
ТГЛ
ИНС
ИР
ТГЛ
Вторичные
Сахарный диабет
Злоупотребление алкоголем
ИНС
ЛПОНП
ХМ
кетоновые тела
СЖК
НЭЖК
АцКоА
С2Н5ОН
ЛПЛ
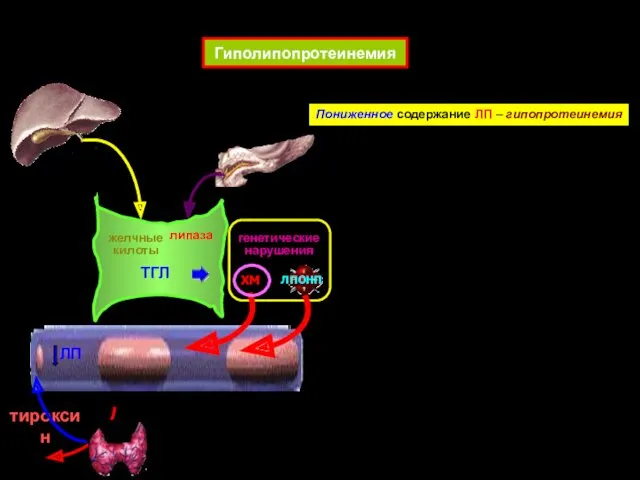
Гиполипопротеинемия
желчные килоты
липаза
ЛП
ХМ
ЛПОНП
ТГЛ
Пониженное содержание ЛП – гипопротеинемия
тироксин
генетические нарушения
Гиполипопротеинемия
желчные килоты
липаза
ЛП
ХМ
ЛПОНП
ТГЛ
Пониженное содержание ЛП – гипопротеинемия
тироксин
генетические нарушения
 Исследование утомляемости и работоспособности школьников. Исследовательская работа школьников
Исследование утомляемости и работоспособности школьников. Исследовательская работа школьников Травы и цветы полей и лугов
Травы и цветы полей и лугов Биологическая эффективность гербицида Miuris 125 в борьбе с однодольными сорными растениями в посевах сой
Биологическая эффективность гербицида Miuris 125 в борьбе с однодольными сорными растениями в посевах сой Лишайники 6 класс
Лишайники 6 класс Побег и почки
Побег и почки Презентация к уроку Гигиена дыхания
Презентация к уроку Гигиена дыхания Человек. Природное и общественное в человеке
Человек. Природное и общественное в человеке Надорганизменный уровень биомониторинга
Надорганизменный уровень биомониторинга Анатомо-физиологические особенности (АФО)
Анатомо-физиологические особенности (АФО) Самые красивые и необычные грибы мира
Самые красивые и необычные грибы мира Строение тела человека. (2 класс)
Строение тела человека. (2 класс) Разнообразие живой природы. Царства живых организмов. Отличительные признаки живого
Разнообразие живой природы. Царства живых организмов. Отличительные признаки живого Пищеварительная система человека
Пищеварительная система человека Класс Паукообразные
Класс Паукообразные Биомеханика двигательной деятельности
Биомеханика двигательной деятельности Нуклеиновые кислоты. ДНК и РНК
Нуклеиновые кислоты. ДНК и РНК Исследовательская деятельность школьников
Исследовательская деятельность школьников Готовимся к ЕГЭ по биологии
Готовимся к ЕГЭ по биологии Биологические и социально-биологические основы адаптации организма человека к физической и умственной деятельности
Биологические и социально-биологические основы адаптации организма человека к физической и умственной деятельности Грибы. Где и чем живут
Грибы. Где и чем живут Ингибиторы ферментов как лекарственные средства
Ингибиторы ферментов как лекарственные средства Такая неизвестная известная гидра
Такая неизвестная известная гидра Пищеварительная система
Пищеварительная система История открытия клетки. Клеточная теория
История открытия клетки. Клеточная теория Дыхание. Этапы дыхания
Дыхание. Этапы дыхания Отруйні гриби
Отруйні гриби Митохондрии и хлоропласты
Митохондрии и хлоропласты Эндемики Африки
Эндемики Африки