- Онтогенез и жизненный цикл особей

Содержание
- 2. ОНТОГЕНЕЗ И ЖИЗНЕННЫЙ ЦИКЛ Для отбора значение имеет только динамика численности особей, оцениваемая по величине репродуктивной
- 3. Элементарной единицей, способной к консервативному самовоспроизведению, и, как следствие, селекционной эволюции является гиперцикл, где J –
- 4. Модульное воспроизведение основано на действии циклических алгоритмов морфогенеза, применяемых к реальным клеткам, или гипотетическим «морфогенам» Двойная
- 5. «МОРФОГЕНЕТИЧЕСКИЙ РОСТ» РАСТИТЕЛЬНОГО ПОБЕГА Рост апикальной меристемы побега связан с периодическими изменениями формы: избыток внутреннего давления
- 6. МОДУЛЬНОЕ ВОСПРОИЗВЕДЕНИЕ – ОСНОВА ОНТОГЕНЕЗА РАСТЕНИЙ Морфогенез всего растения – последовательная редукция начальных стадий развития его
- 7. Обратную связь такого рода подразумевает гипотеза «смешанной (слитной) наследственности»: у потомков – любое значение признака в
- 8. Платой за мозаичную наследственность (то есть, за возможность селекционной эволюции) является уникальность генотипа особи: воспроизводим только
- 9. Фенотипическое значение признака, обусловленное неаддитивным взаимодействием аллельных (доминирование) или не аллельных генов (эпистаз) не наследуется При
- 10. ОТВЕТ НА ОТБОР ПО КОЛИЧЕСТВЕННОМУ ПРИЗНАКУ X R = η2S R – ответ на отбор по
- 11. Наследуемость (η2 = 0. 71) роста у человека (линия регрессии «родителей на потомков»): количественный признак, являющийся
- 12. Если учесть средовую дисперсию, то VP (общая фенотипическая дисперсия) = VG + VE (средовая дисперсия) Величину
- 13. Из-за двойственности самовоспроизведения (сочетания консервативной и неконсервативной компоненты) в эволюции взаимодействуют две системы отбора (1) Дифференциальное
- 14. НОРМА РЕАКЦИИ Следуя определению нормы реакции как пределов изменения фенотипа без изменения генотипа, норма реакции генотипа
- 15. Генетическая ассимиляция (Уоддингтон, 1957) фенотипа crossveinless у дрозофилы: изменение навстречу среде («наследование благоприобретенных признаков»)
- 17. ФУНДАМЕНТАЛЬНАЯ ТЕОРЕМА ЕСТЕСТВЕННОГО ОТБОРА (ТЕОРЕМА ФИШЕРА) Отбор максимизирует среднюю приспособленность W* со скоростью, равной дисперсии W*,
- 18. ФУНДАМЕНТАЛЬНАЯ ТЕОРЕМА ОТБОРА (СЛОВЕСНАЯ ФОРМУЛИРОВКА) (1) В ходе естественного отбора средняя приспособленность популяции не может уменьшаться
- 19. Адаптивный ландшафт («горная страна») Райта Изменение самого ландшафта ~ эволюция экосистемы («борьба адаптивных пиков») Из фундаментальной
- 20. Отбор на изменение нормы реакции складывается из отбора генов с аддитивным эффектом (отвечающих за наследуемость признаков
- 21. ОТ ПЛАВНИКА РЫБ К КОНЕЧНОСТИ TETRAPODA Эволюция фиксирует гетерохронию между морфогенезом и дифференцировкой: конденсация мезенхимы смещается
- 22. У анамний наружный слой эктодермы постепенно превращается в клеточную оболочку зародыша. Из-за ее обособления ЭП теряют
- 23. Эволюция онтогенеза основана на возможности прямого и обратного перехода от «детерминированного хаоса» к множеству путей и,
- 24. СТАБИЛИЗИРУЮЩИЙ ОТБОР Кроме того, стабилизирующему отбору часто приписывают канализирующий эффект (картинка), то есть, переход от нормального
- 25. Скутеллярные щетинки дрозофилы Дикий тип (мода) – 4 щетинки по углам скутеллума. Сильный положительный эксцесс в
- 26. Увеличение дозы генов scute Макрохеты конкурируют за материал с кутикулой скутеллума – при избытке его материала
- 27. ПОДТВЕРЖДАЮЩИЙ ПРИМЕР СО СТЕРНОПЛЕВРАЬНЫМИ ЩЕТИНКАМИ Для стерноплевральных щетинок (микрохеты) их конкуренция друг с другом – единственный
- 28. РЕКАПИТУЛЯЦИЯ ФИЛОГЕНЕЗА ИЛИ ЭВОЛЮЦИЯ ОНТОГЕНЕЗА? «Онтогения есть краткое повторение филогении» (Геккель) «Отклонения онтогении от филогении вторичны
- 30. Скачать презентацию

ОНТОГЕНЕЗ И ЖИЗНЕННЫЙ ЦИКЛ
Для отбора значение имеет только динамика численности особей,
ОНТОГЕНЕЗ И ЖИЗНЕННЫЙ ЦИКЛ
Для отбора значение имеет только динамика численности особей,
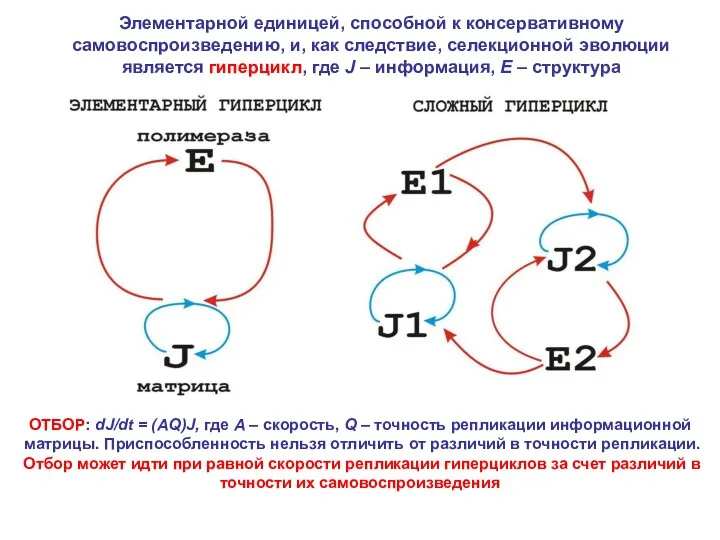
Элементарной единицей, способной к консервативному самовоспроизведению, и, как следствие, селекционной эволюции
Элементарной единицей, способной к консервативному самовоспроизведению, и, как следствие, селекционной эволюции
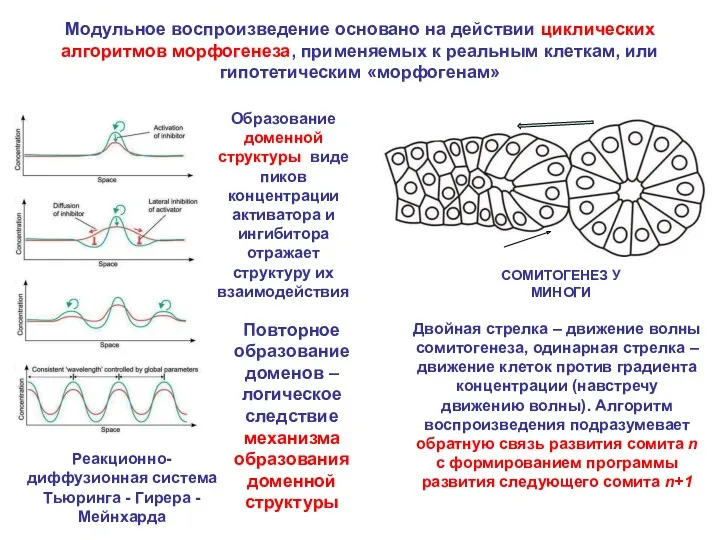
Модульное воспроизведение основано на действии циклических алгоритмов морфогенеза, применяемых к реальным
Модульное воспроизведение основано на действии циклических алгоритмов морфогенеза, применяемых к реальным
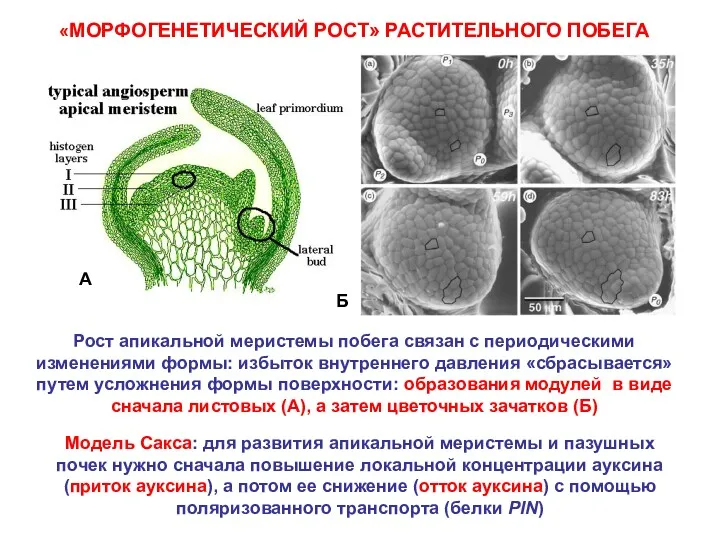
«МОРФОГЕНЕТИЧЕСКИЙ РОСТ» РАСТИТЕЛЬНОГО ПОБЕГА
Рост апикальной меристемы побега связан с периодическими изменениями
«МОРФОГЕНЕТИЧЕСКИЙ РОСТ» РАСТИТЕЛЬНОГО ПОБЕГА
Рост апикальной меристемы побега связан с периодическими изменениями

МОДУЛЬНОЕ ВОСПРОИЗВЕДЕНИЕ – ОСНОВА ОНТОГЕНЕЗА РАСТЕНИЙ
Морфогенез всего растения – последовательная редукция
МОДУЛЬНОЕ ВОСПРОИЗВЕДЕНИЕ – ОСНОВА ОНТОГЕНЕЗА РАСТЕНИЙ
Морфогенез всего растения – последовательная редукция
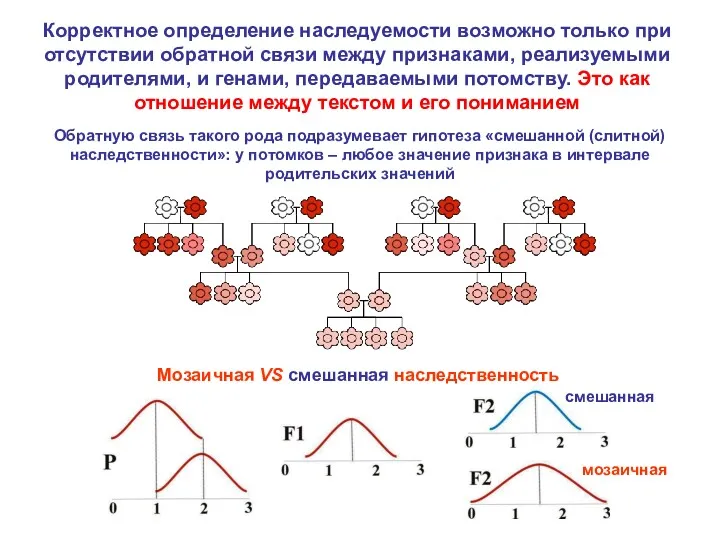
Обратную связь такого рода подразумевает гипотеза «смешанной (слитной) наследственности»: у потомков
Обратную связь такого рода подразумевает гипотеза «смешанной (слитной) наследственности»: у потомков

Платой за мозаичную наследственность (то есть, за возможность селекционной эволюции) является
Платой за мозаичную наследственность (то есть, за возможность селекционной эволюции) является
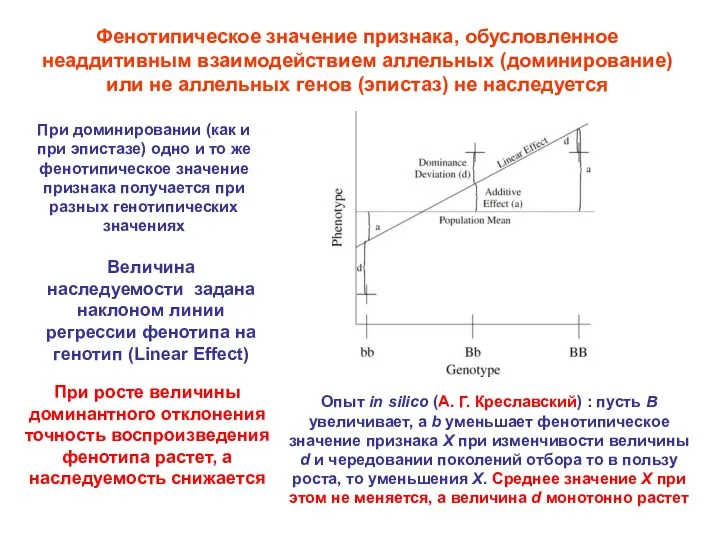
Фенотипическое значение признака, обусловленное неаддитивным взаимодействием аллельных (доминирование) или не аллельных
Фенотипическое значение признака, обусловленное неаддитивным взаимодействием аллельных (доминирование) или не аллельных
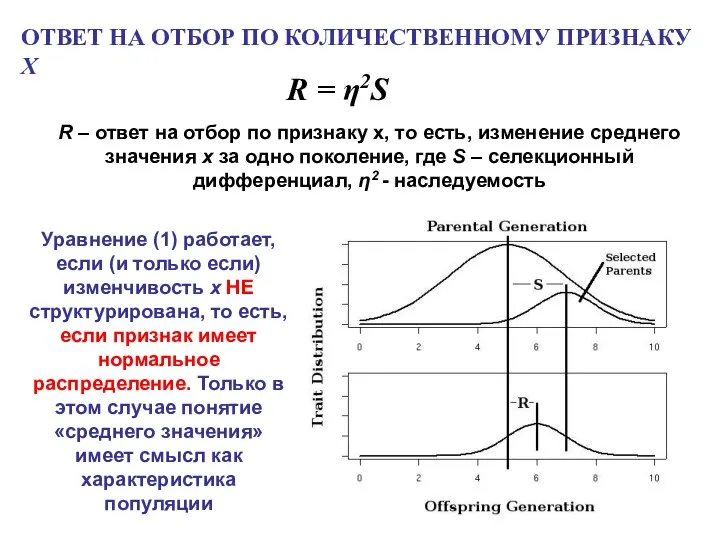
ОТВЕТ НА ОТБОР ПО КОЛИЧЕСТВЕННОМУ ПРИЗНАКУ X
R = η2S
R –
ОТВЕТ НА ОТБОР ПО КОЛИЧЕСТВЕННОМУ ПРИЗНАКУ X
R = η2S
R –

Наследуемость (η2 = 0. 71) роста у человека (линия регрессии «родителей
Наследуемость (η2 = 0. 71) роста у человека (линия регрессии «родителей

Если учесть средовую дисперсию, то VP (общая фенотипическая дисперсия) = VG
Если учесть средовую дисперсию, то VP (общая фенотипическая дисперсия) = VG

Из-за двойственности самовоспроизведения (сочетания консервативной и неконсервативной компоненты) в эволюции взаимодействуют
Из-за двойственности самовоспроизведения (сочетания консервативной и неконсервативной компоненты) в эволюции взаимодействуют
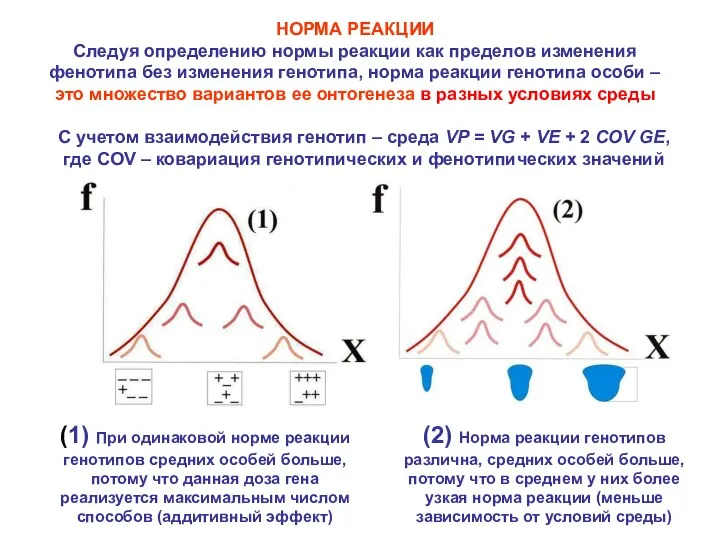
НОРМА РЕАКЦИИ
Следуя определению нормы реакции как пределов изменения фенотипа без изменения
НОРМА РЕАКЦИИ
Следуя определению нормы реакции как пределов изменения фенотипа без изменения
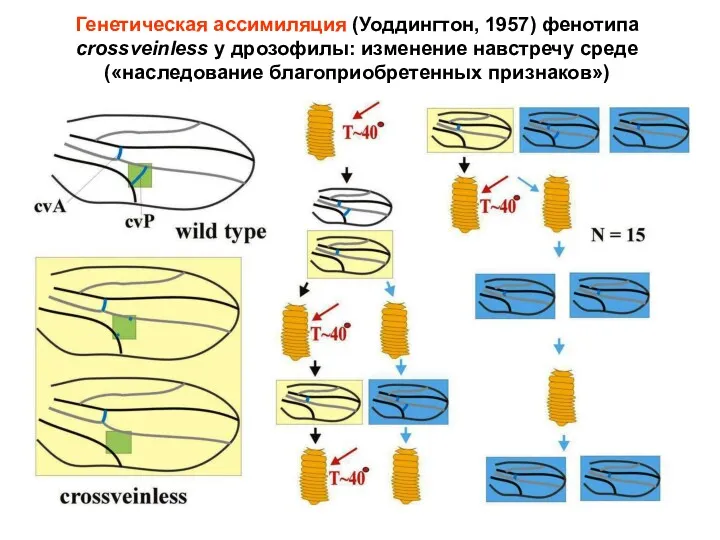
Генетическая ассимиляция (Уоддингтон, 1957) фенотипа crossveinless у дрозофилы: изменение навстречу среде
Генетическая ассимиляция (Уоддингтон, 1957) фенотипа crossveinless у дрозофилы: изменение навстречу среде

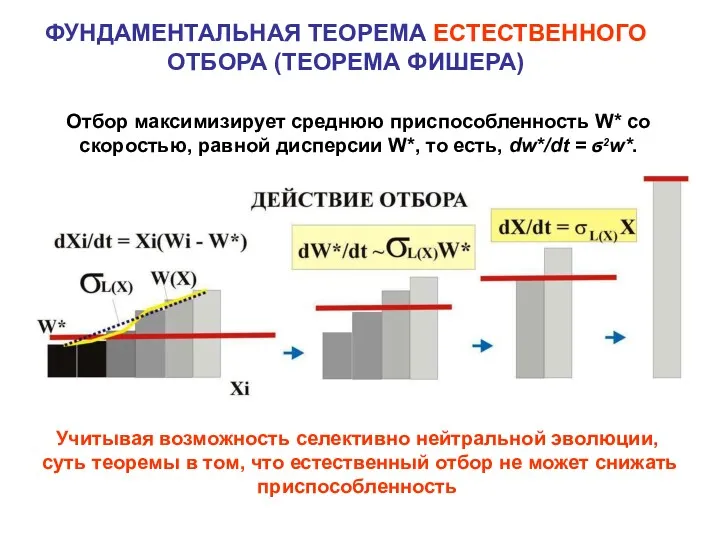
ФУНДАМЕНТАЛЬНАЯ ТЕОРЕМА ЕСТЕСТВЕННОГО ОТБОРА (ТЕОРЕМА ФИШЕРА)
Отбор максимизирует среднюю приспособленность W* со
ФУНДАМЕНТАЛЬНАЯ ТЕОРЕМА ЕСТЕСТВЕННОГО ОТБОРА (ТЕОРЕМА ФИШЕРА)
Отбор максимизирует среднюю приспособленность W* со

ФУНДАМЕНТАЛЬНАЯ ТЕОРЕМА ОТБОРА (СЛОВЕСНАЯ ФОРМУЛИРОВКА)
(1) В ходе естественного отбора средняя приспособленность
ФУНДАМЕНТАЛЬНАЯ ТЕОРЕМА ОТБОРА (СЛОВЕСНАЯ ФОРМУЛИРОВКА)
(1) В ходе естественного отбора средняя приспособленность
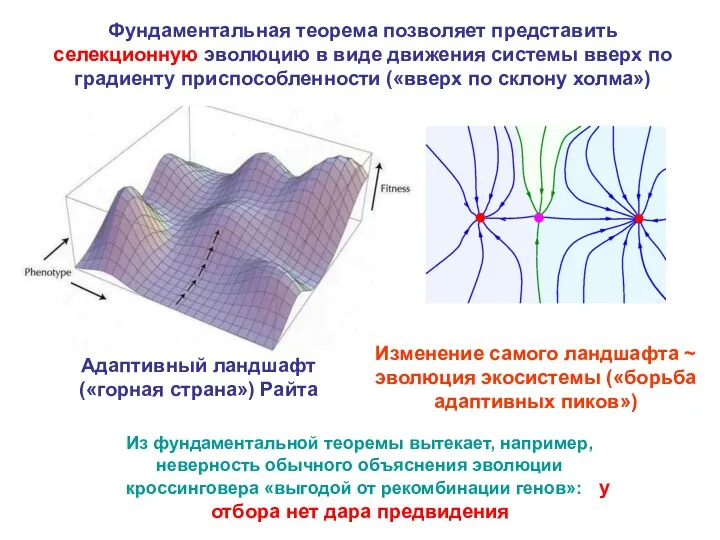
Адаптивный ландшафт («горная страна») Райта
Изменение самого ландшафта ~ эволюция экосистемы («борьба
Адаптивный ландшафт («горная страна») Райта
Изменение самого ландшафта ~ эволюция экосистемы («борьба

Отбор на изменение нормы реакции складывается из отбора генов с аддитивным
Отбор на изменение нормы реакции складывается из отбора генов с аддитивным
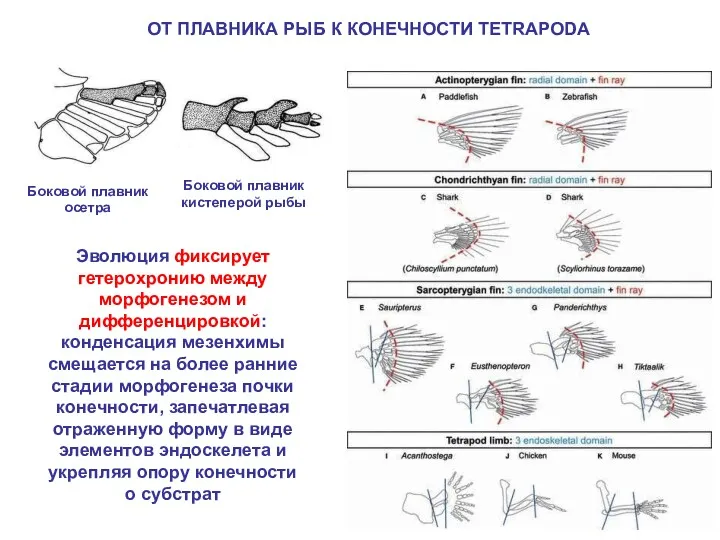
ОТ ПЛАВНИКА РЫБ К КОНЕЧНОСТИ TETRAPODA
Эволюция фиксирует гетерохронию между морфогенезом и
ОТ ПЛАВНИКА РЫБ К КОНЕЧНОСТИ TETRAPODA
Эволюция фиксирует гетерохронию между морфогенезом и
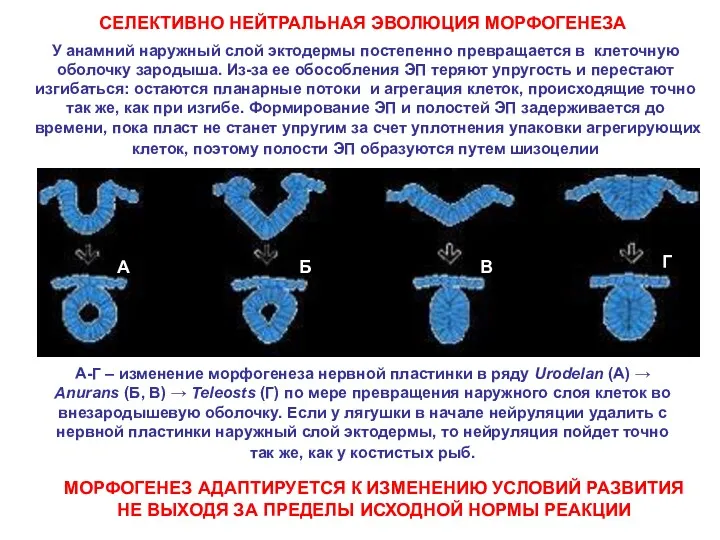
У анамний наружный слой эктодермы постепенно превращается в клеточную оболочку зародыша.
У анамний наружный слой эктодермы постепенно превращается в клеточную оболочку зародыша.
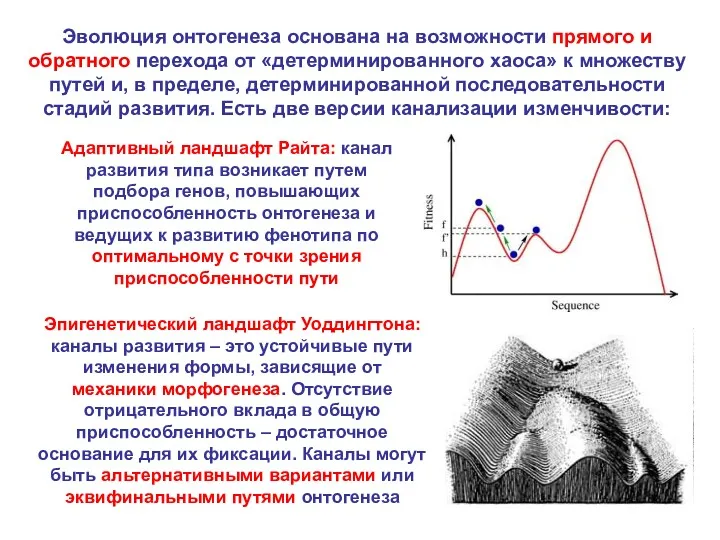
Эволюция онтогенеза основана на возможности прямого и обратного перехода от «детерминированного
Эволюция онтогенеза основана на возможности прямого и обратного перехода от «детерминированного
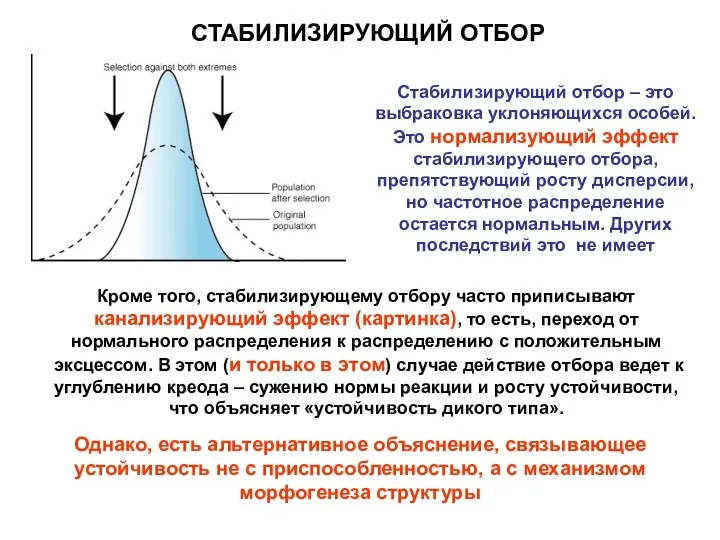
СТАБИЛИЗИРУЮЩИЙ ОТБОР
Кроме того, стабилизирующему отбору часто приписывают канализирующий эффект (картинка), то
СТАБИЛИЗИРУЮЩИЙ ОТБОР
Кроме того, стабилизирующему отбору часто приписывают канализирующий эффект (картинка), то
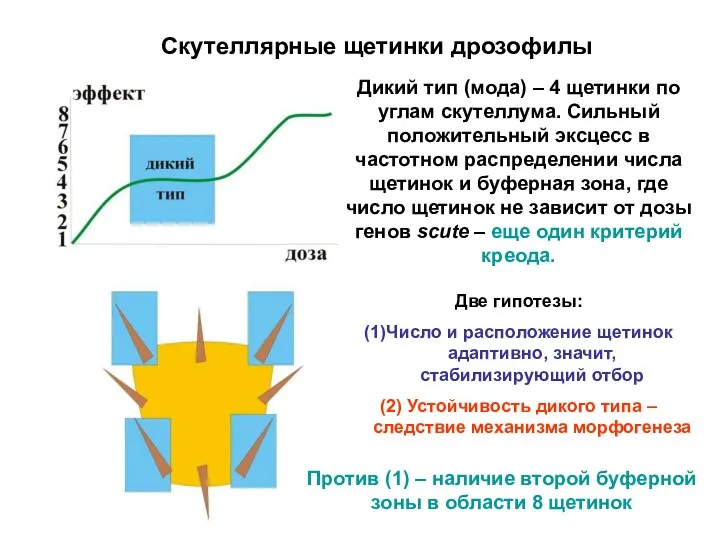
Скутеллярные щетинки дрозофилы
Дикий тип (мода) – 4 щетинки по углам скутеллума.
Скутеллярные щетинки дрозофилы
Дикий тип (мода) – 4 щетинки по углам скутеллума.

Увеличение дозы генов scute
Макрохеты конкурируют за материал с кутикулой скутеллума –
Увеличение дозы генов scute
Макрохеты конкурируют за материал с кутикулой скутеллума –
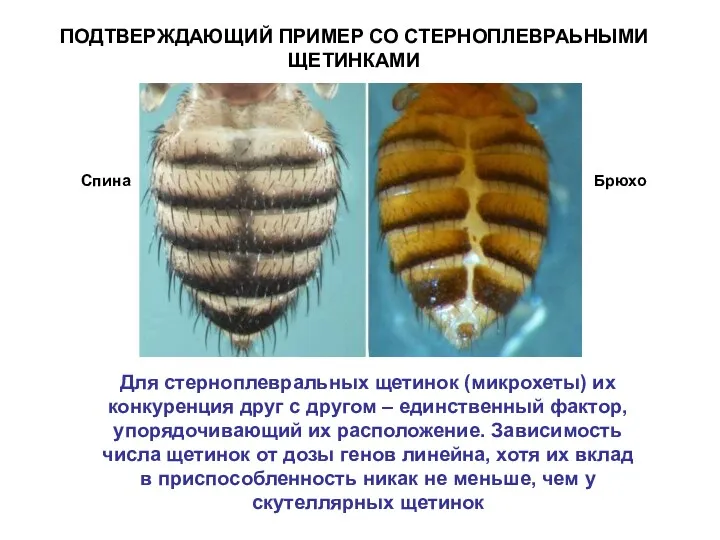
ПОДТВЕРЖДАЮЩИЙ ПРИМЕР СО СТЕРНОПЛЕВРАЬНЫМИ ЩЕТИНКАМИ
Для стерноплевральных щетинок (микрохеты) их конкуренция друг
ПОДТВЕРЖДАЮЩИЙ ПРИМЕР СО СТЕРНОПЛЕВРАЬНЫМИ ЩЕТИНКАМИ
Для стерноплевральных щетинок (микрохеты) их конкуренция друг
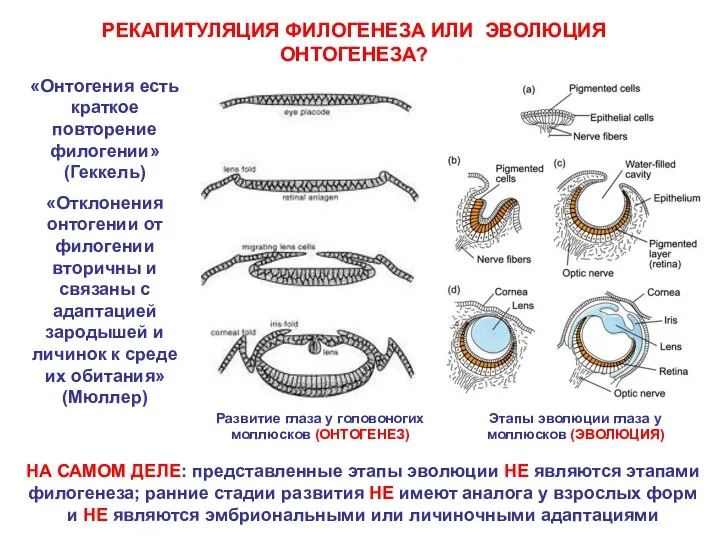
РЕКАПИТУЛЯЦИЯ ФИЛОГЕНЕЗА ИЛИ ЭВОЛЮЦИЯ ОНТОГЕНЕЗА?
«Онтогения есть краткое повторение филогении» (Геккель)
«Отклонения онтогении
РЕКАПИТУЛЯЦИЯ ФИЛОГЕНЕЗА ИЛИ ЭВОЛЮЦИЯ ОНТОГЕНЕЗА?
«Онтогения есть краткое повторение филогении» (Геккель)
«Отклонения онтогении
 Строение стебля
Строение стебля Анализаторы. Зрительный анализатор
Анализаторы. Зрительный анализатор Анатомия человека. Средний мозг. Промежуточный мозг
Анатомия человека. Средний мозг. Промежуточный мозг Гниение аминокислот в кишечнике
Гниение аминокислот в кишечнике Лишайники. Значение лишайников
Лишайники. Значение лишайников Променеве дослідження нирок та сечовидільної системи
Променеве дослідження нирок та сечовидільної системи Моногибридное скрещивание. Задача
Моногибридное скрещивание. Задача Микробные технологии – основа биотехнологии
Микробные технологии – основа биотехнологии Гормональная регуляция метаболизма: основы
Гормональная регуляция метаболизма: основы Пищеварение животных
Пищеварение животных Эволюция низших растений
Эволюция низших растений Composting (part 1)
Composting (part 1) Презентация к уроку биологии на тему:Индивидуальное развитие организмов. Биогенетический закон. 9 класс
Презентация к уроку биологии на тему:Индивидуальное развитие организмов. Биогенетический закон. 9 класс Разнообразие животных
Разнообразие животных Гетеротрофное питание. 6 класс
Гетеротрофное питание. 6 класс Неорганические вещества клетки (10 класс)
Неорганические вещества клетки (10 класс) презентация к уроку Пищеварение в кишечнике
презентация к уроку Пищеварение в кишечнике Хемосинтез и его экологическая роль
Хемосинтез и его экологическая роль Презентація. Заповідники України.
Презентація. Заповідники України. fМуравьи - какие они?
fМуравьи - какие они? Собака - лучший друг человека
Собака - лучший друг человека Антропогенез. Происхождение человека
Антропогенез. Происхождение человека Витаминные препараты
Витаминные препараты Бактериялар және актиномициттер туғызатын ауыл шаруашылық дақылдарының ауруы
Бактериялар және актиномициттер туғызатын ауыл шаруашылық дақылдарының ауруы Гербарии растений, содержащих простые фенолы, фенолгликозиды и антраценпроизводные
Гербарии растений, содержащих простые фенолы, фенолгликозиды и антраценпроизводные Микробиологические методы определения активности антибиотиков
Микробиологические методы определения активности антибиотиков Технология рекомбинантных ДНК. (Лекция 3)
Технология рекомбинантных ДНК. (Лекция 3) Презентация к урокуДеление клетки. Митоз
Презентация к урокуДеление клетки. Митоз