- Процессы торможения. Доминанта. Методы исследования ЦНС. Спинной мозг

Содержание
- 2. Торможение – местный нервный процесс, приводящий к угнетению или устранению возбуждения. В отличие от возбуждения не
- 3. Торможение в ЦНС выполняет две основные функции: Во-первых, оно координирует функции, т.е. оно направляет возбуждение по
- 4. Классификация центрального торможения По различным признакам: По электрическому состоянию мембраны –деполяризационное и гиперполяризационное; По отношению к
- 5. Торможение в ЦНС (опыт И.М. Сеченова)
- 6. ТРИ СОСТОЯНИЯ МЕМБРАНЫ
- 7. Механизм торможения Е0 Ек Ек Ек Е0 Е0 гиперполяризация Длительная деполяризация
- 8. Постсинаптическое торможение Развивается в условиях, когда медиатор, выделяемый нервным окончанием, изменяет свойства постсинаптической мембраны таким образом,
- 9. Постсинаптическое торможение В нервных окончаниях тормозящих нейронов под влиянием приходящего по аксону импульса выделяется медиатор, гиперполяризующий
- 10. Эффективность торможения клетки зависит от соотношения между величинами возбуждающего и тормозного потенциалов и от числа возбуждающих
- 11. По нейрональной организации выделяют: поступательное, латеральное (боковое), возвратное, реципрокное.
- 12. Поступательное торможение обусловлено включением тормозных нейронов на пути следования возбуждения.
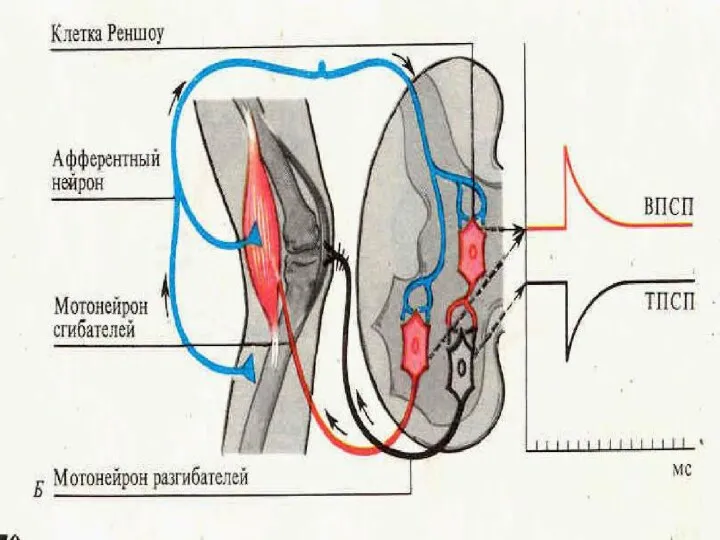
- 13. Торможение возвратное - процесс регуляции нервными клетками интенсивности поступающих к ним сигналов по способу обратной связи.
- 14. Коллатерали аксона мотонейрона (1) контактируют с телом клетки Реншоу (2), короткий аксон которой, разветвляясь, образует тормозные
- 15. Латеральное (боковое) торможение Вставочные клетки формируют тормозные синапсы на соседних нейронах, блокируя боковые пути распространения возбуждения.
- 16. Латеральное торможение в формировании контрастного изображения Возбуждающий нейрон обозначен светлым цветом, тормозящий – черным, серым –
- 17. Реципрокное торможение Примером реципрокного торможения является торможение центров мышц-антагонистов. Суть этого вида торможения заключается в том,
- 19. ТОРМОЖЕНИЕ В ЦНС а) Возвратное торможение по Реншоу б) Латеральное торможение В - возбуждение Т -
- 20. Разновидности постсинаптического торможения: 1 – параллельное; 2 – возвратное; 3 – латеральное; 4 – прямое. Нейроны
- 21. Пессимальное торможение • В этом случае торможение развивается в возбуждающих синапсах в результате сильной деполяризации постсинаптической
- 22. Нервные импульсы, возникающее при возбуждении тормозных нейронов, не отличаются от потенциалов действия обычных возбуждающих нейронов. Однако
- 23. Посттетаническое торможение Возникает в случае, если после окончания возбуждения в клетке возникает сильная гиперполяризация мембраны. Причина
- 24. Пресинаптическое торможение Пресинаптическое торможение локализуется в пресинаптических терминалях перед синаптической бляшкой. На пресинаптических терминалях располагаются окончания
- 26. ПРЕСИНАПТИЧЕСКОЕ ТОРМОЖЕНИЕ 1 - аксон тормозного нейрона 2 - аксон возбуждающего нейрона 3 - постсинаптическая мембрана
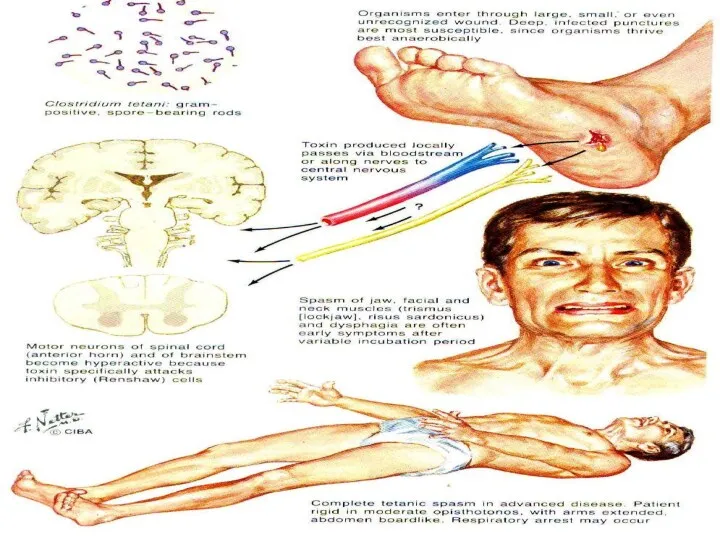
- 27. Примеры нарушения торможения в ЦНС НАРУШЕНИЕ ПОСТСИНАПТИЧЕСКОГО ТОРМОЖЕНИЯ: СТРИХНИН →БЛОКАДА РЕЦЕПТОРОВ ТОРМОЗНЫХ СИНАПСОВ; СТОЛБНЯЧНЫЙ ТОКСИН →НАРУШЕНИЕ
- 29. Поза больного в состоянии опистотонуса
- 30. ПРИНЦИПЫ КООРДИНАЦИИ РЕФЛЕКТОРНОЙ ДЕЯТЕЛЬНОСТИ Иррадиация и концентрация возбуждения и торможения, их взаимная индукция. -возбуждение и торможение
- 31. ПРИНЦИПЫ КООРДИНАЦИИ РЕФЛЕКТОРНОЙ ДЕЯТЕЛЬНОСТИ 1. РЕЦИПРОКНОСТИ; 2. ОБЩЕГО КОНЕЧНОГО ПУТИ; 3. ДОМИНАНТЫ; 4. СУБОРДИНАЦИИ; 5. ОБРАТНОЙ
- 32. Реципрокная иннервация Для центров безусловно-рефлекторной деятельности взаимная индукция наиболее рельефно выступает в центрах сопряженно работающих сгибателей
- 33. ПРИНЦИП РЕЦИПРОКНОСТИ (СОПРЯЖЕННОГО ТОРМОЖЕНИЯ) + + - - +
- 34. Принцип общего поля Если иметь в виду только чувствующие нейроны, несущие импульсы к спинному мозгу, то
- 35. ЦНС можно представить в качестве "воронки", с широким входным отверстием, куда поступают раздражения с различных рецепторов,
- 36. Принцип «конечного пути» основан на анатомических отношениях в НС. Открыт в 1904 г. Чарльзом С. Шеррингтоном.
- 37. Принцип общего конечного пути Актуальным становится самый сильный раздражитель. Принцип общего поля обеспечивает использование одних и
- 38. ПРИНЦИП ОБЩЕГО КОНЕЧНОГО ПУТИ КОРА ПОДКОРКА СТВОЛ СПИННОЙ МОЗГ
- 39. Принцип общего конечного пути в спинном мозге
- 40. Афферентные импульсы, рождающиеся в организме в результате деятельности органов и тканей, получили название вторичных афферентных импульсов
- 41. Принцип обратной связи В осуществлении рефлекторных реакций и их координации огромное значение принадлежит обратной связи, которая
- 42. ПРИНЦИП ОБРАТНОЙ АФФЕРЕНТАЦИИ ДВИГАТЕЛЬНАЯ КОМАНДА ОБРАТНАЯ АФФЕРЕНТАЦИЯ
- 43. Благодаря существованию обратной связи между НЦ и рабочими аппаратами интенсивность возбуждения различных групп нейронов в НЦ
- 44. ОПРЕДЕЛЕНИЕ ДОМИНАНТЫ ( ПО А.А. Ухтомскому, 1931) В 1904-1911 г. А.А. Ухтомский провел серию экспериментов, которые
- 45. ОПРЕДЕЛЕНИЕ ДОМИНАНТЫ ( ПО А.А.Ухтомскому, 1931) • Доминанта - временно господствующий рефлекс или поведенческий акт, которым
- 46. Основные признаки доминанты ( по А.А.Ухтомскому) 1. Повышенная возбудимость доминантного центра; 2. Стойкость возбуждения в доминантном
- 47. Пример проявления активности доминантного очага → обнимательный рефлекс у весенних лягушек-самцов. За счет сокращения сгибателей передних
- 48. МЕТОДЫ ИССЛЕДОВАНИЯ ЦНС Анатомо-клинический Наблюдения Раздражения Экстирпации Регистрации электропотенциалов (ЭЭГ) Компьютерная томография (рентгеновская, ЯМР, магнитная) Исследование
- 49. Спинной мозг Это наиболее простой, примитивный по строению и физиологическим функциям отдел ЦНС. Спинной мозг представляет
- 50. Закон Белла - Мажанди Вентральные (передние) корешки содержат эфферентные двигательные (выходящие) волокна, дорсальные (задние) корешки содержат
- 51. Спинной мозг (medullaspinalis) - наиболее древний нижний отдел центральной нервной системы, расположенный в позвоночном канале и
- 52. Обозначения цветом: оранжевым- шейные сегменты и шейные позвонки фиолетовым—грудные зеленым —поясничные розовым—крестцовые голубым—копчиковые. Обозначения цифрами: римскими
- 53. Сегменты спинного мозга 8 шейных (C1 - C8) 12 грудных (Th1 - Th12) 5 поясничных (L1
- 55. В спинном мозге расположены: - центр диафрагмального нерва (3-4 шейный сегмент), - центры мускулатуры верхних конечностей
- 56. Схема сегментарной чувствительности кожи человека Примечание. Буквы и цифры указывают сегменты спинного мозга, афферентные нервы которых
- 57. Нейроны спинного мозга Вставочные или интернейроны (97%); Двигательные или мотонейроны (3%): альфа-мотонейроны фазические (быстрые) тонические (медленные)
- 58. ФУНКЦИИ СПИННОГО МОЗГА РЕФЛЕКТОРНАЯ ПРОВОДНИКОВАЯ ТОНИЧЕСКАЯ
- 59. Проводниковые функции спинного мозга Проводниковые функции спинного мозга основаны на деятельности восходящих и нисходящих путей. Они
- 60. Проводниковые функции мозга Системы Афферентная Эфферентная Разрезы промежуточного мозга среднего мозга Моста продолговатого мозга спинного мозга
- 61. ПРОВОДЯЩИЕ СИСТЕМЫ СПИННОГО МОЗГА ВОСХОДЯЩИЕ ПУТИ (ЭКСТЕРО- ПРОПРИО-ИНТЕРОЦЕПТИВНАЯ ЧУВСТВИТЕЛЬНОСТЬ) НИСХОДЯЩИЕ ПУТИ (ЭФФЕКТОРНЫЕ, ДВИГАТЕЛЬНЫЕ) СОБСТВЕННЫЕ (ПРОПРИОСПИНАЛЬНЫЕ) ПУТИ
- 62. ВОСХОДЯЩИЕ ПУТИ СПИННОГО МОЗГА Тонкий пучок Голля (fasciculus gracilis) - от нижней части тела – проприоцепторы
- 63. Нисходящие пути спинного мозга Латеральный кортикоспинальный пирамидный тракт - двигательные зоны коры - перекрест в продолговатом
- 64. Корково-спинномозговой (пирамидный) путь (лат. tractus corticospinalis)
- 65. Основные рефлексы спинного мозга Рефлекторные функции спинного мозга основываются на объединении афферентных входов через задние корешки
- 66. ТОНИЧЕСКАЯ ФУНКЦИЯ СПИННОГО МОЗГА. ГАММА-МОТОРНАЯ ПЕТЛЯ
- 67. Рефлексы спинного мозга Классификация рефлексов Специальные: мочеиспускания, дефекации, эрекции, эякуляции Сухожильные Миотатические Тонические Ритмические
- 69. Особенности компенсаторных процессов при нарушениях функций спинного мозга При интенсивном воздействии на спинной мозг (травма, нарушение
- 70. Длительность восстановления нарушенных спинальных функций зависит: от обширности и характера травмы; от уровня повреждения спинного мозга,
- 71. СИНДРОМ БРОУН-СЕКАРА - (Brown-Sequard syndrome) - неврологическое заболевание, связанное с нарушением функции ряда сегментов спинного мозга.
- 72. Зоны поражения у человека при повреждении спинного мозга на уровне грудных сегментов (паралич Броун-Секара)
- 73. Ведущими клиническими признаками синдрома Броун-Секара являются: 1. Спастический (центральный) паралич (парез) на ипсилатеральной стороне (стороне поражения)
- 74. 4. Утрата болевой и температурной чувствительности по проводниковому типу на контрлатеральной стороне вследствие поражения нео-спино-таламического тракта,
- 75. 5. Расстройство всех видов чувствительности по сегментарному типу на стороне поражения, если повреждено два сегмента и
- 76. Схема расстройства поверхностной чувствительности у больного с синдромом Броун–Секара с верхней границей DXI (на уровне поясничного
- 78. Скачать презентацию

Торможение – местный нервный процесс, приводящий к угнетению или устранению
Торможение – местный нервный процесс, приводящий к угнетению или устранению

Торможение в ЦНС выполняет две основные функции:
Во-первых, оно координирует функции, т.е.
Торможение в ЦНС выполняет две основные функции:
Во-первых, оно координирует функции, т.е.

Классификация центрального торможения
По различным признакам:
По электрическому состоянию мембраны –деполяризационное и
Классификация центрального торможения
По различным признакам:
По электрическому состоянию мембраны –деполяризационное и
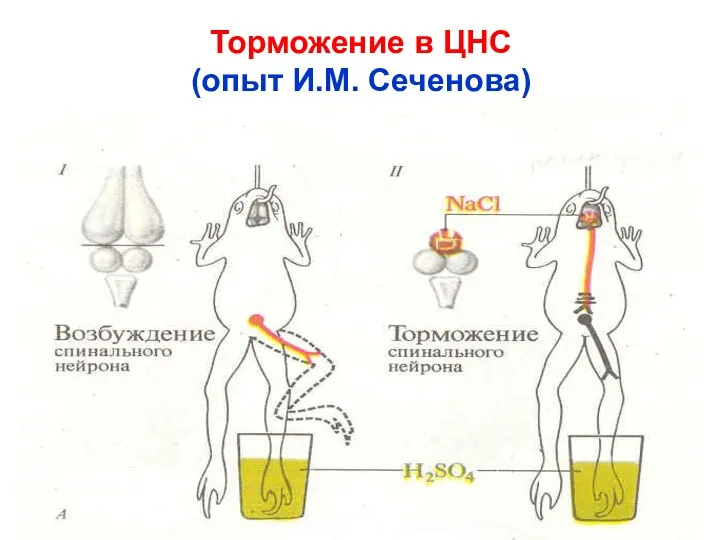
Торможение в ЦНС
(опыт И.М. Сеченова)
Торможение в ЦНС
(опыт И.М. Сеченова)
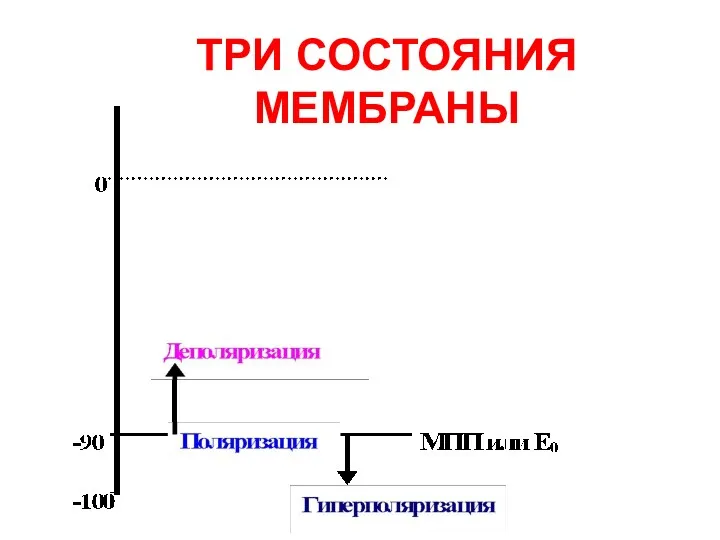
ТРИ СОСТОЯНИЯ МЕМБРАНЫ
ТРИ СОСТОЯНИЯ МЕМБРАНЫ
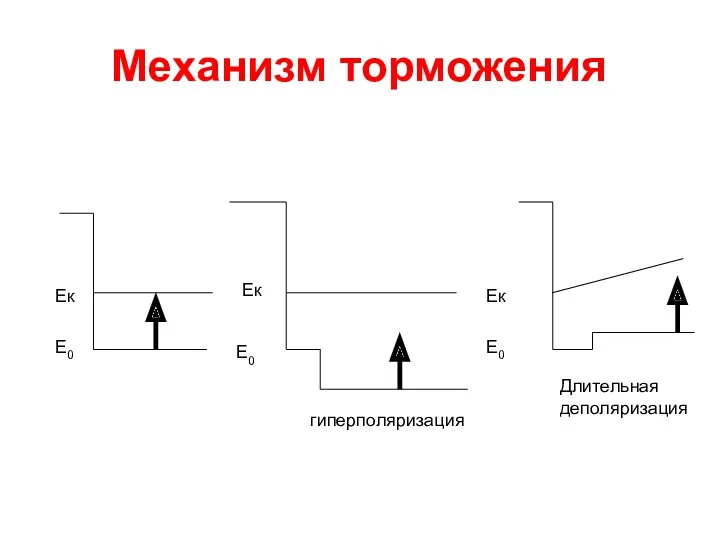
Механизм торможения
Е0
Ек
Ек
Ек
Е0
Е0
гиперполяризация
Длительная деполяризация
Механизм торможения
Е0
Ек
Ек
Ек
Е0
Е0
гиперполяризация
Длительная деполяризация

Постсинаптическое торможение
Развивается в условиях, когда медиатор, выделяемый нервным окончанием, изменяет свойства
Постсинаптическое торможение
Развивается в условиях, когда медиатор, выделяемый нервным окончанием, изменяет свойства

Постсинаптическое торможение
В нервных окончаниях тормозящих нейронов под влиянием приходящего по аксону
Постсинаптическое торможение
В нервных окончаниях тормозящих нейронов под влиянием приходящего по аксону

Эффективность торможения клетки зависит от соотношения между величинами возбуждающего и тормозного
Эффективность торможения клетки зависит от соотношения между величинами возбуждающего и тормозного

По нейрональной организации выделяют:
поступательное,
латеральное (боковое),
возвратное,
реципрокное.
По нейрональной организации выделяют:
поступательное,
латеральное (боковое),
возвратное,
реципрокное.
Поступательное торможение
обусловлено включением тормозных нейронов на пути следования возбуждения.
Поступательное торможение
обусловлено включением тормозных нейронов на пути следования возбуждения.

Торможение возвратное - процесс регуляции нервными клетками интенсивности поступающих к ним
Торможение возвратное - процесс регуляции нервными клетками интенсивности поступающих к ним
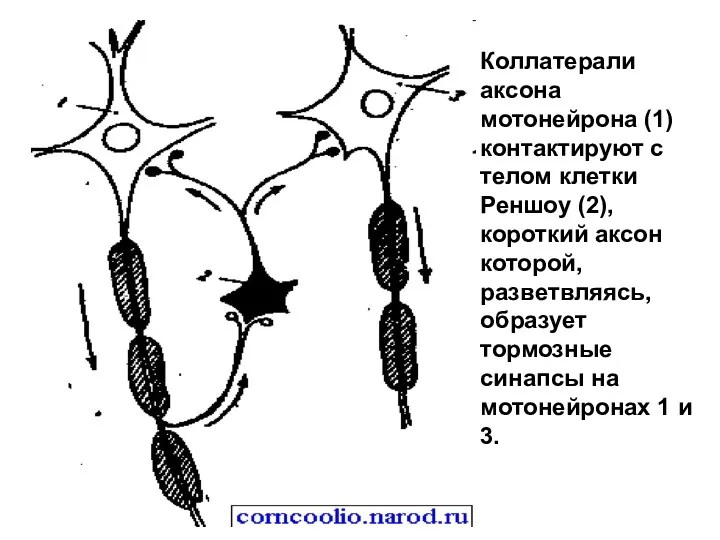
Коллатерали аксона мотонейрона (1) контактируют с телом клетки Реншоу (2), короткий
Коллатерали аксона мотонейрона (1) контактируют с телом клетки Реншоу (2), короткий

Латеральное (боковое) торможение
Вставочные клетки формируют тормозные синапсы на соседних нейронах, блокируя
Латеральное (боковое) торможение
Вставочные клетки формируют тормозные синапсы на соседних нейронах, блокируя
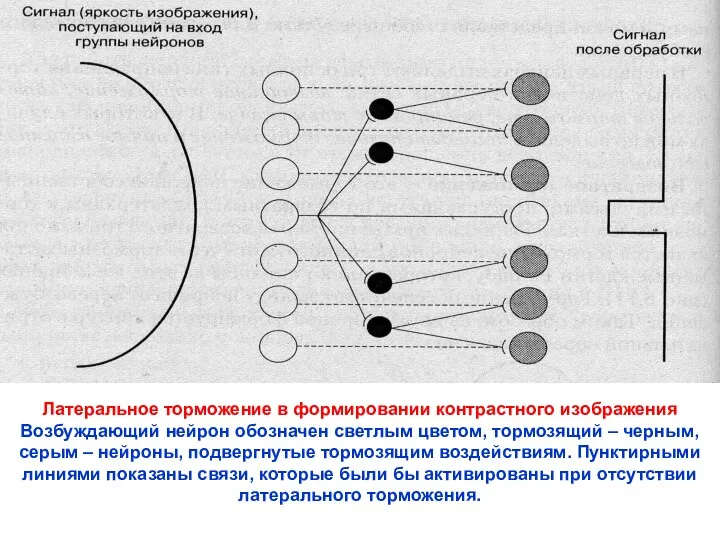
Латеральное торможение в формировании контрастного изображения
Возбуждающий нейрон обозначен светлым цветом, тормозящий
Латеральное торможение в формировании контрастного изображения
Возбуждающий нейрон обозначен светлым цветом, тормозящий

Реципрокное торможение
Примером реципрокного торможения является торможение центров мышц-антагонистов.
Суть этого вида
Реципрокное торможение
Примером реципрокного торможения является торможение центров мышц-антагонистов.
Суть этого вида
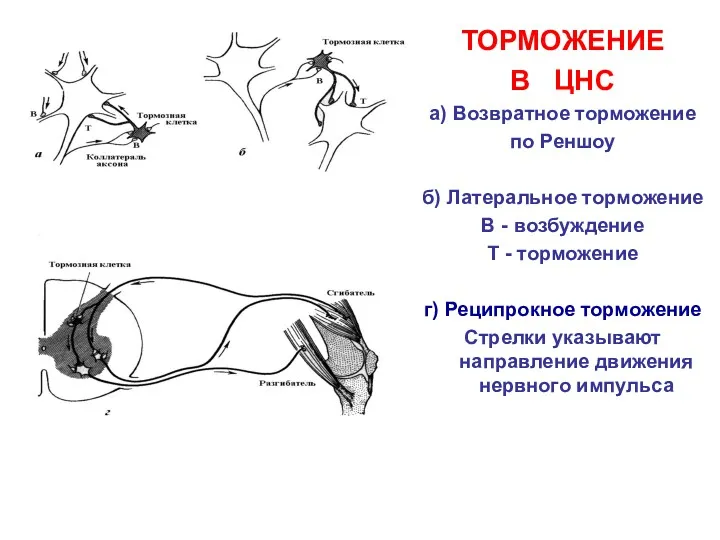
ТОРМОЖЕНИЕ
В ЦНС
а) Возвратное торможение
по Реншоу
б) Латеральное торможение
В - возбуждение
Т -
ТОРМОЖЕНИЕ
В ЦНС
а) Возвратное торможение
по Реншоу
б) Латеральное торможение
В - возбуждение
Т -
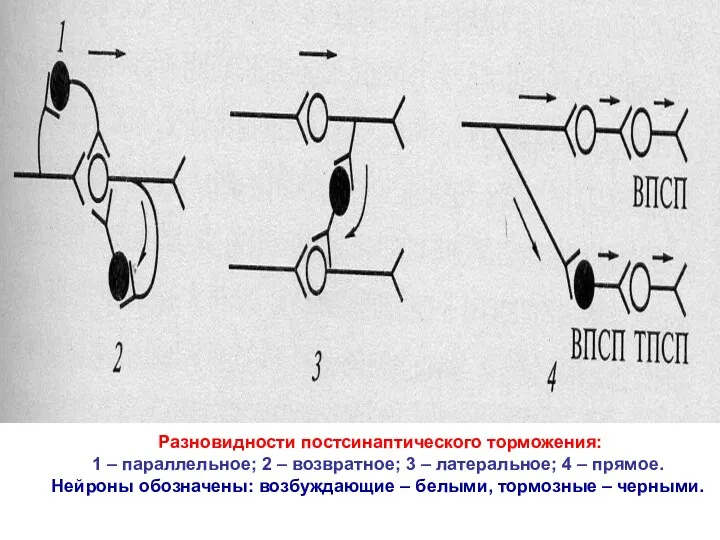
Разновидности постсинаптического торможения:
1 – параллельное; 2 – возвратное; 3 –
Разновидности постсинаптического торможения:
1 – параллельное; 2 – возвратное; 3 –

Пессимальное торможение
• В этом случае торможение развивается в
возбуждающих синапсах
Пессимальное торможение
• В этом случае торможение развивается в
возбуждающих синапсах

Нервные импульсы, возникающее при возбуждении тормозных нейронов, не отличаются от потенциалов
Нервные импульсы, возникающее при возбуждении тормозных нейронов, не отличаются от потенциалов
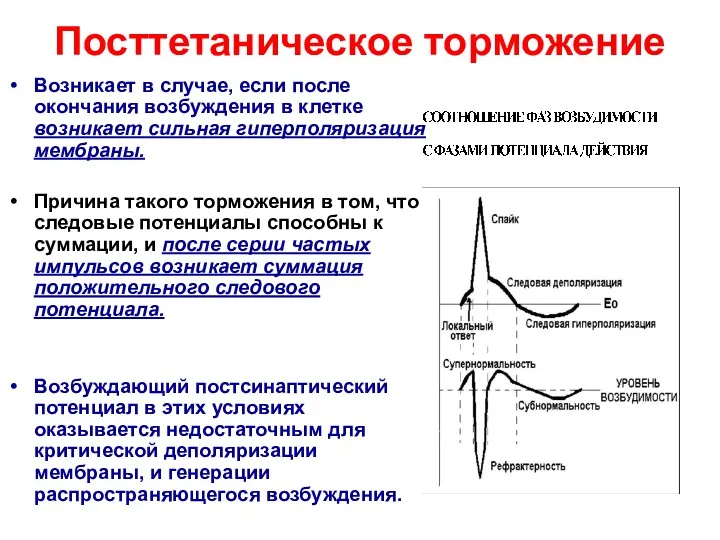
Посттетаническое торможение
Возникает в случае, если после окончания возбуждения в клетке возникает
Посттетаническое торможение
Возникает в случае, если после окончания возбуждения в клетке возникает

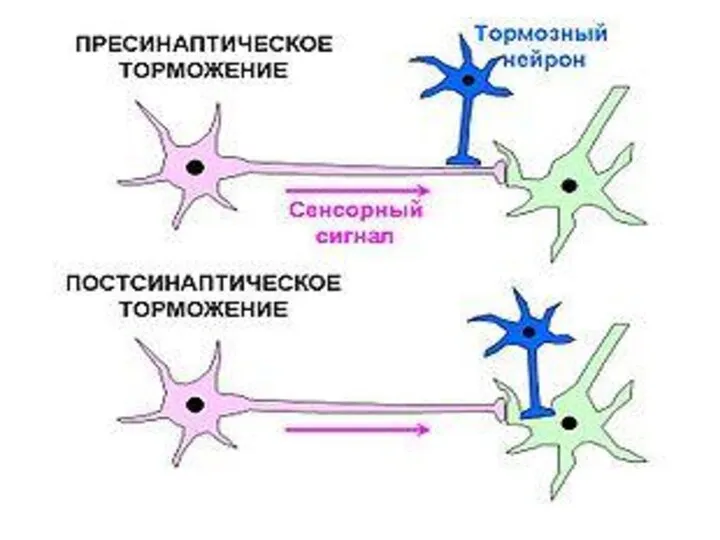
Пресинаптическое торможение
Пресинаптическое торможение локализуется в пресинаптических терминалях перед синаптической бляшкой.
Пресинаптическое торможение
Пресинаптическое торможение локализуется в пресинаптических терминалях перед синаптической бляшкой.
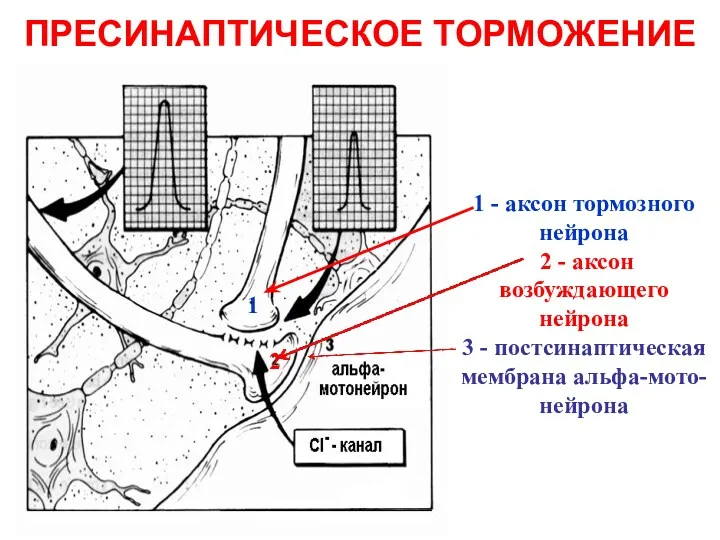
ПРЕСИНАПТИЧЕСКОЕ ТОРМОЖЕНИЕ
1 - аксон тормозного нейрона
2 - аксон возбуждающего нейрона
3
ПРЕСИНАПТИЧЕСКОЕ ТОРМОЖЕНИЕ
1 - аксон тормозного нейрона
2 - аксон возбуждающего нейрона
3

Примеры нарушения торможения в ЦНС
НАРУШЕНИЕ ПОСТСИНАПТИЧЕСКОГО ТОРМОЖЕНИЯ:
СТРИХНИН →БЛОКАДА РЕЦЕПТОРОВ ТОРМОЗНЫХ
Примеры нарушения торможения в ЦНС
НАРУШЕНИЕ ПОСТСИНАПТИЧЕСКОГО ТОРМОЖЕНИЯ:
СТРИХНИН →БЛОКАДА РЕЦЕПТОРОВ ТОРМОЗНЫХ
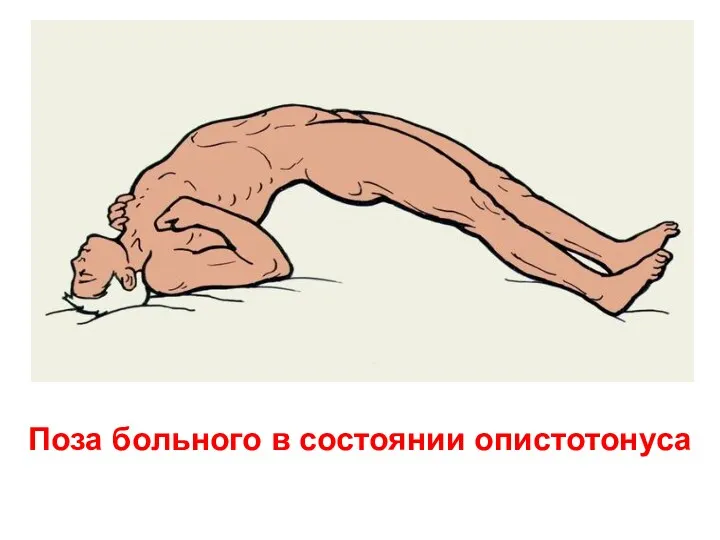
Поза больного в состоянии опистотонуса
Поза больного в состоянии опистотонуса

ПРИНЦИПЫ КООРДИНАЦИИ РЕФЛЕКТОРНОЙ ДЕЯТЕЛЬНОСТИ
Иррадиация и концентрация возбуждения и торможения, их взаимная
ПРИНЦИПЫ КООРДИНАЦИИ РЕФЛЕКТОРНОЙ ДЕЯТЕЛЬНОСТИ
Иррадиация и концентрация возбуждения и торможения, их взаимная

ПРИНЦИПЫ КООРДИНАЦИИ РЕФЛЕКТОРНОЙ ДЕЯТЕЛЬНОСТИ
1. РЕЦИПРОКНОСТИ;
2. ОБЩЕГО КОНЕЧНОГО ПУТИ;
3. ДОМИНАНТЫ;
4. СУБОРДИНАЦИИ;
5. ОБРАТНОЙ
ПРИНЦИПЫ КООРДИНАЦИИ РЕФЛЕКТОРНОЙ ДЕЯТЕЛЬНОСТИ
1. РЕЦИПРОКНОСТИ;
2. ОБЩЕГО КОНЕЧНОГО ПУТИ;
3. ДОМИНАНТЫ;
4. СУБОРДИНАЦИИ;
5. ОБРАТНОЙ

Реципрокная иннервация
Для центров безусловно-рефлекторной деятельности взаимная индукция наиболее рельефно выступает
Реципрокная иннервация
Для центров безусловно-рефлекторной деятельности взаимная индукция наиболее рельефно выступает
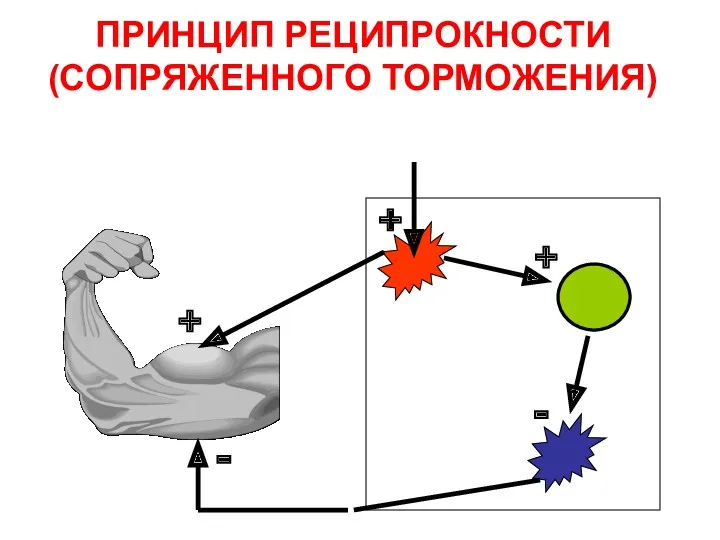
ПРИНЦИП РЕЦИПРОКНОСТИ
(СОПРЯЖЕННОГО ТОРМОЖЕНИЯ)
+
+
-
-
+
ПРИНЦИП РЕЦИПРОКНОСТИ
(СОПРЯЖЕННОГО ТОРМОЖЕНИЯ)
+
+
-
-
+

Принцип общего поля
Если иметь в виду только чувствующие нейроны, несущие
Принцип общего поля
Если иметь в виду только чувствующие нейроны, несущие

ЦНС можно представить в качестве "воронки", с широким входным отверстием, куда
ЦНС можно представить в качестве "воронки", с широким входным отверстием, куда

Принцип «конечного пути» основан на анатомических отношениях в НС. Открыт в
Принцип «конечного пути» основан на анатомических отношениях в НС. Открыт в

Принцип общего конечного пути
Актуальным становится самый сильный раздражитель.
Принцип общего поля
Принцип общего конечного пути
Актуальным становится самый сильный раздражитель.
Принцип общего поля
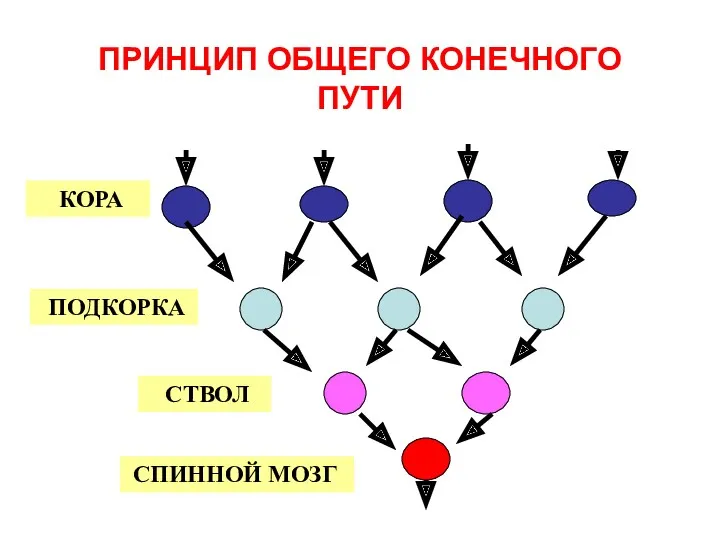
ПРИНЦИП ОБЩЕГО КОНЕЧНОГО ПУТИ
КОРА
ПОДКОРКА
СТВОЛ
СПИННОЙ МОЗГ
ПРИНЦИП ОБЩЕГО КОНЕЧНОГО ПУТИ
КОРА
ПОДКОРКА
СТВОЛ
СПИННОЙ МОЗГ
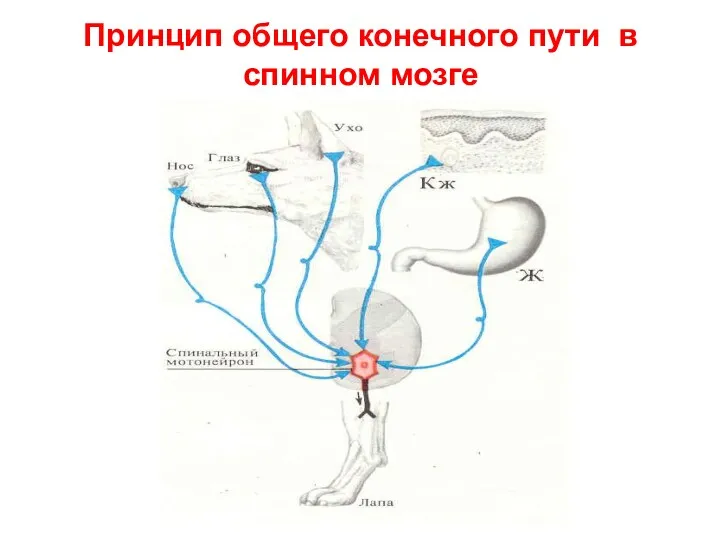
Принцип общего конечного пути в спинном мозге
Принцип общего конечного пути в спинном мозге

Афферентные импульсы, рождающиеся в организме в результате деятельности органов и тканей,
Афферентные импульсы, рождающиеся в организме в результате деятельности органов и тканей,

Принцип обратной связи
В осуществлении рефлекторных реакций и их координации огромное
Принцип обратной связи
В осуществлении рефлекторных реакций и их координации огромное
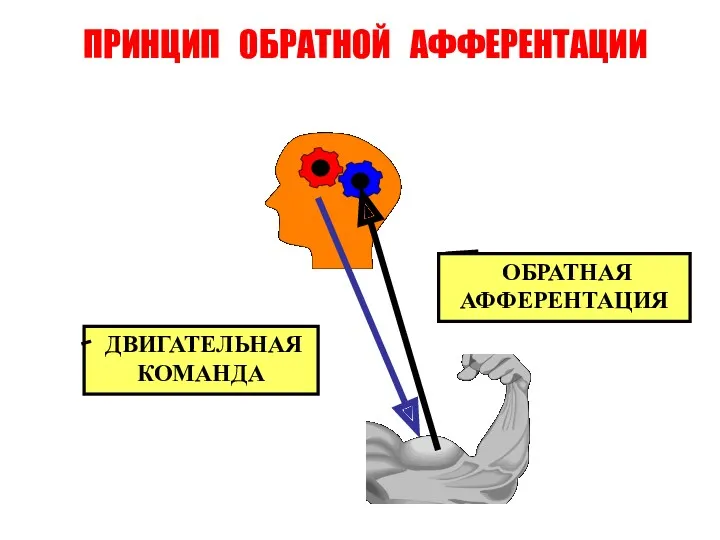
ПРИНЦИП ОБРАТНОЙ АФФЕРЕНТАЦИИ
ДВИГАТЕЛЬНАЯ КОМАНДА
ОБРАТНАЯ
АФФЕРЕНТАЦИЯ
ПРИНЦИП ОБРАТНОЙ АФФЕРЕНТАЦИИ
ДВИГАТЕЛЬНАЯ КОМАНДА
ОБРАТНАЯ
АФФЕРЕНТАЦИЯ

Благодаря существованию обратной связи между НЦ и рабочими аппаратами интенсивность возбуждения
Благодаря существованию обратной связи между НЦ и рабочими аппаратами интенсивность возбуждения

ОПРЕДЕЛЕНИЕ ДОМИНАНТЫ
( ПО А.А. Ухтомскому, 1931)
В 1904-1911 г. А.А. Ухтомский провел
ОПРЕДЕЛЕНИЕ ДОМИНАНТЫ
( ПО А.А. Ухтомскому, 1931)
В 1904-1911 г. А.А. Ухтомский провел

ОПРЕДЕЛЕНИЕ ДОМИНАНТЫ
( ПО А.А.Ухтомскому, 1931)
• Доминанта - временно господствующий рефлекс или
ОПРЕДЕЛЕНИЕ ДОМИНАНТЫ
( ПО А.А.Ухтомскому, 1931)
• Доминанта - временно господствующий рефлекс или

Основные признаки доминанты
( по А.А.Ухтомскому)
1. Повышенная возбудимость доминантного центра;
2. Стойкость возбуждения
Основные признаки доминанты
( по А.А.Ухтомскому)
1. Повышенная возбудимость доминантного центра;
2. Стойкость возбуждения

Пример проявления активности доминантного очага → обнимательный рефлекс у весенних лягушек-самцов.
Пример проявления активности доминантного очага → обнимательный рефлекс у весенних лягушек-самцов.

МЕТОДЫ ИССЛЕДОВАНИЯ ЦНС
Анатомо-клинический
Наблюдения
Раздражения
Экстирпации
Регистрации электропотенциалов (ЭЭГ)
Компьютерная томография (рентгеновская, ЯМР, магнитная)
Исследование навязанного и
МЕТОДЫ ИССЛЕДОВАНИЯ ЦНС
Анатомо-клинический
Наблюдения
Раздражения
Экстирпации
Регистрации электропотенциалов (ЭЭГ)
Компьютерная томография (рентгеновская, ЯМР, магнитная)
Исследование навязанного и

Спинной мозг
Это наиболее простой, примитивный по строению и физиологическим функциям отдел
Спинной мозг
Это наиболее простой, примитивный по строению и физиологическим функциям отдел
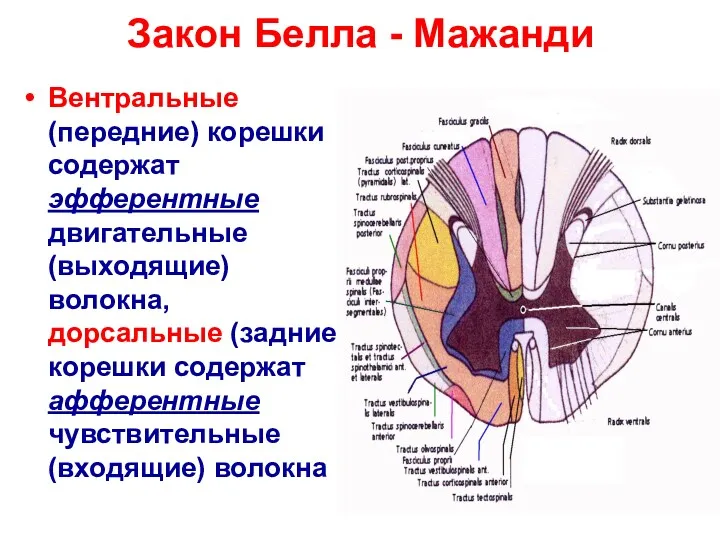
Закон Белла - Мажанди
Вентральные (передние) корешки содержат эфферентные двигательные (выходящие) волокна,
Закон Белла - Мажанди
Вентральные (передние) корешки содержат эфферентные двигательные (выходящие) волокна,
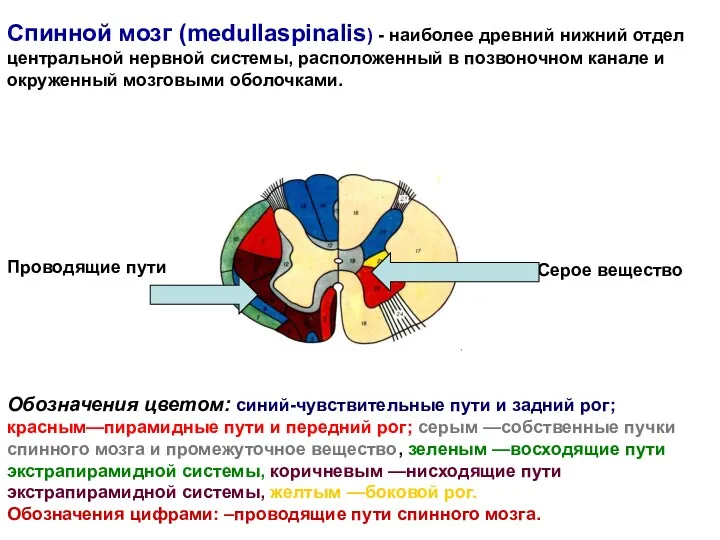
Спинной мозг (medullaspinalis) - наиболее древний нижний отдел центральной нервной системы,
Спинной мозг (medullaspinalis) - наиболее древний нижний отдел центральной нервной системы,
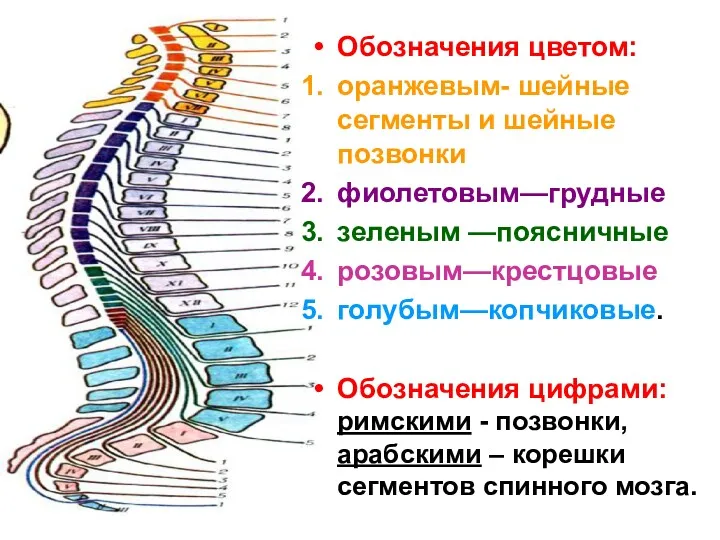
Обозначения цветом:
оранжевым- шейные сегменты и шейные позвонки
фиолетовым—грудные
зеленым —поясничные
розовым—крестцовые
голубым—копчиковые.
Обозначения цифрами: римскими -
Обозначения цветом:
оранжевым- шейные сегменты и шейные позвонки
фиолетовым—грудные
зеленым —поясничные
розовым—крестцовые
голубым—копчиковые.
Обозначения цифрами: римскими -
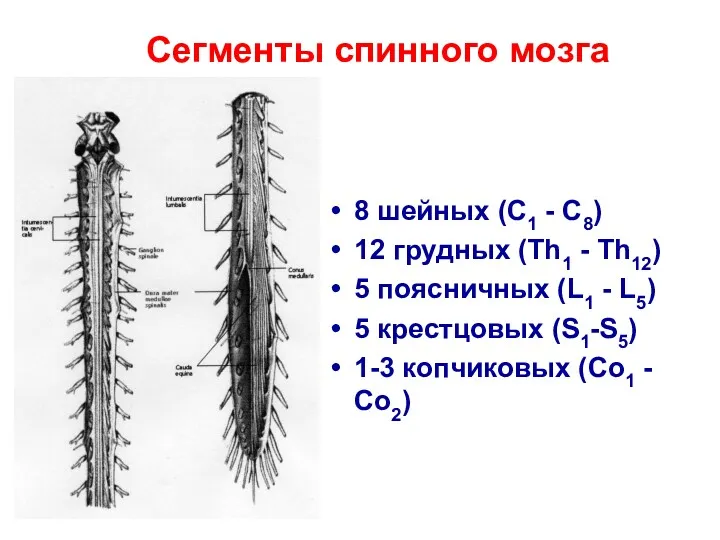
Сегменты спинного мозга
8 шейных (C1 - C8)
12 грудных (Th1 - Th12)
5
Сегменты спинного мозга
8 шейных (C1 - C8)
12 грудных (Th1 - Th12)
5
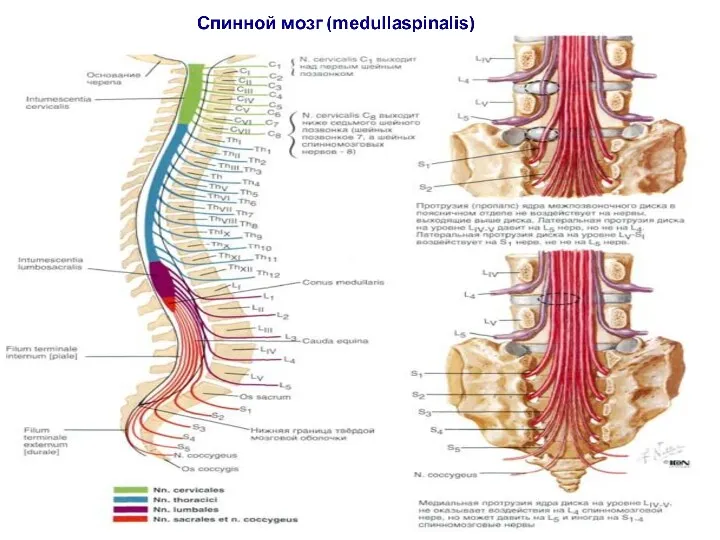

В спинном мозге расположены:
- центр диафрагмального нерва (3-4 шейный
В спинном мозге расположены:
- центр диафрагмального нерва (3-4 шейный
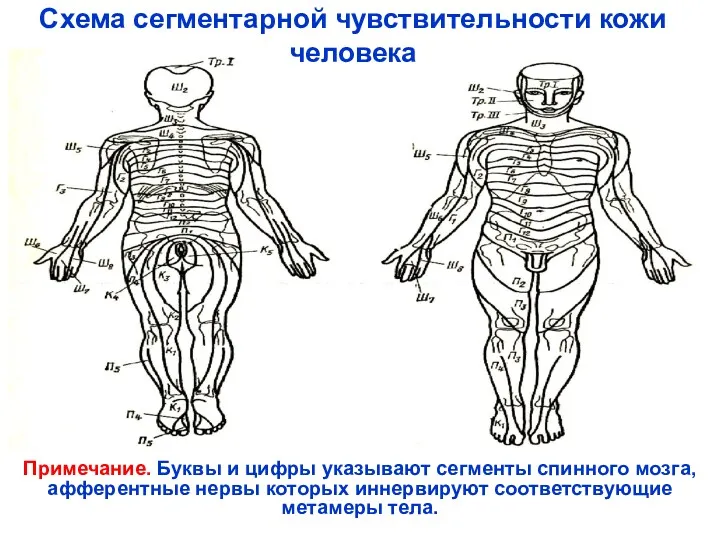
Схема сегментарной чувствительности кожи человека
Примечание. Буквы и цифры указывают сегменты спинного
Схема сегментарной чувствительности кожи человека
Примечание. Буквы и цифры указывают сегменты спинного

Нейроны спинного мозга
Вставочные или интернейроны (97%);
Двигательные или мотонейроны (3%):
альфа-мотонейроны
фазические (быстрые)
тонические
Нейроны спинного мозга
Вставочные или интернейроны (97%);
Двигательные или мотонейроны (3%):
альфа-мотонейроны
фазические (быстрые)
тонические
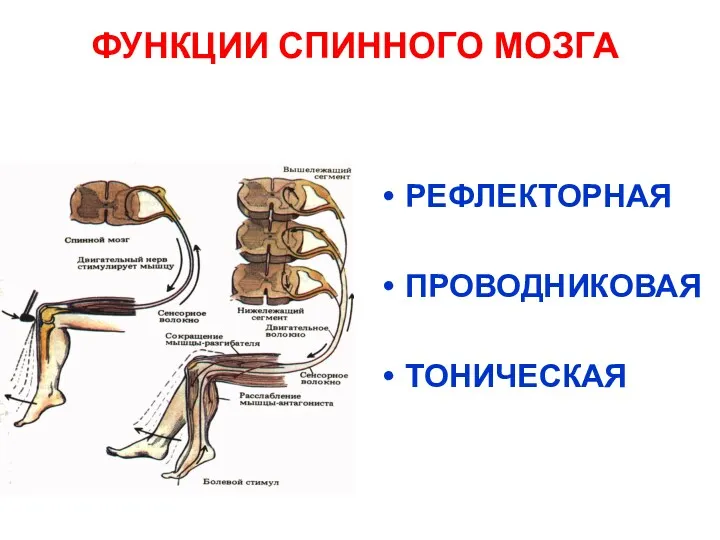
ФУНКЦИИ СПИННОГО МОЗГА
РЕФЛЕКТОРНАЯ
ПРОВОДНИКОВАЯ
ТОНИЧЕСКАЯ
ФУНКЦИИ СПИННОГО МОЗГА
РЕФЛЕКТОРНАЯ
ПРОВОДНИКОВАЯ
ТОНИЧЕСКАЯ
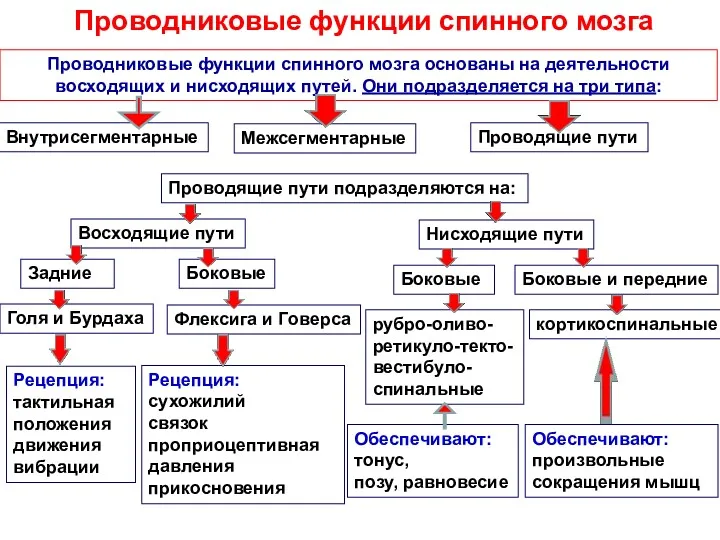
Проводниковые функции спинного мозга
Проводниковые функции спинного мозга основаны на деятельности
восходящих и
Проводниковые функции спинного мозга
Проводниковые функции спинного мозга основаны на деятельности
восходящих и
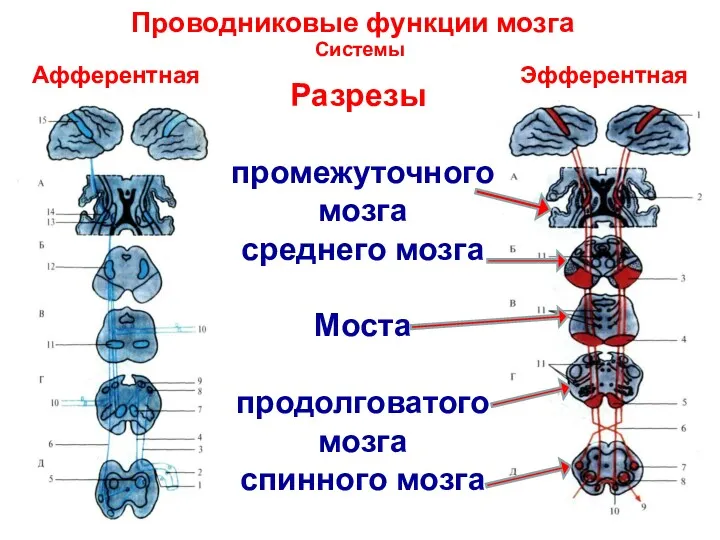
Проводниковые функции мозга
Системы
Афферентная Эфферентная
Разрезы
промежуточного мозга
среднего мозга
Моста
продолговатого
мозга
спинного мозга
Проводниковые функции мозга
Системы
Афферентная Эфферентная
Разрезы
промежуточного мозга
среднего мозга
Моста
продолговатого
мозга
спинного мозга

ПРОВОДЯЩИЕ СИСТЕМЫ
СПИННОГО МОЗГА
ВОСХОДЯЩИЕ ПУТИ (ЭКСТЕРО- ПРОПРИО-ИНТЕРОЦЕПТИВНАЯ ЧУВСТВИТЕЛЬНОСТЬ)
НИСХОДЯЩИЕ ПУТИ (ЭФФЕКТОРНЫЕ, ДВИГАТЕЛЬНЫЕ)
ПРОВОДЯЩИЕ СИСТЕМЫ
СПИННОГО МОЗГА
ВОСХОДЯЩИЕ ПУТИ (ЭКСТЕРО- ПРОПРИО-ИНТЕРОЦЕПТИВНАЯ ЧУВСТВИТЕЛЬНОСТЬ)
НИСХОДЯЩИЕ ПУТИ (ЭФФЕКТОРНЫЕ, ДВИГАТЕЛЬНЫЕ)

ВОСХОДЯЩИЕ ПУТИ СПИННОГО МОЗГА
Тонкий пучок Голля (fasciculus gracilis) - от нижней
ВОСХОДЯЩИЕ ПУТИ СПИННОГО МОЗГА
Тонкий пучок Голля (fasciculus gracilis) - от нижней

Нисходящие пути спинного мозга
Латеральный кортикоспинальный пирамидный тракт - двигательные зоны коры
Нисходящие пути спинного мозга
Латеральный кортикоспинальный пирамидный тракт - двигательные зоны коры
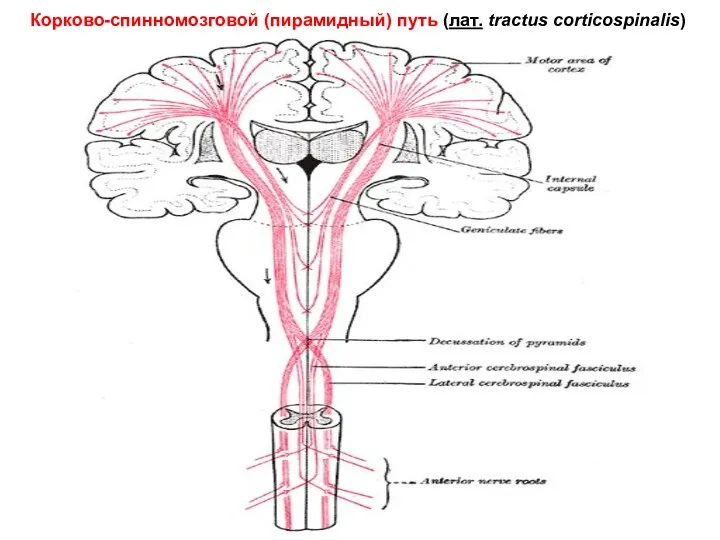
Корково-спинномозговой (пирамидный) путь (лат. tractus corticospinalis)
Корково-спинномозговой (пирамидный) путь (лат. tractus corticospinalis)

Основные рефлексы спинного мозга
Рефлекторные функции спинного мозга основываются на объединении афферентных
Основные рефлексы спинного мозга
Рефлекторные функции спинного мозга основываются на объединении афферентных
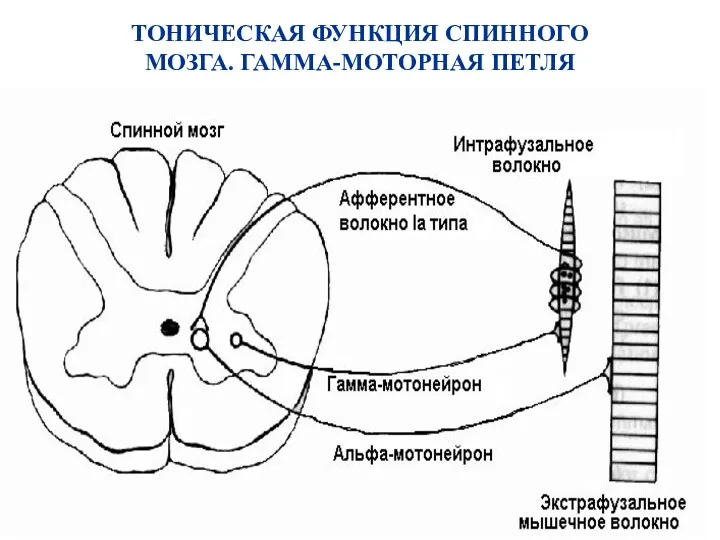
ТОНИЧЕСКАЯ ФУНКЦИЯ СПИННОГО МОЗГА. ГАММА-МОТОРНАЯ ПЕТЛЯ
ТОНИЧЕСКАЯ ФУНКЦИЯ СПИННОГО МОЗГА. ГАММА-МОТОРНАЯ ПЕТЛЯ
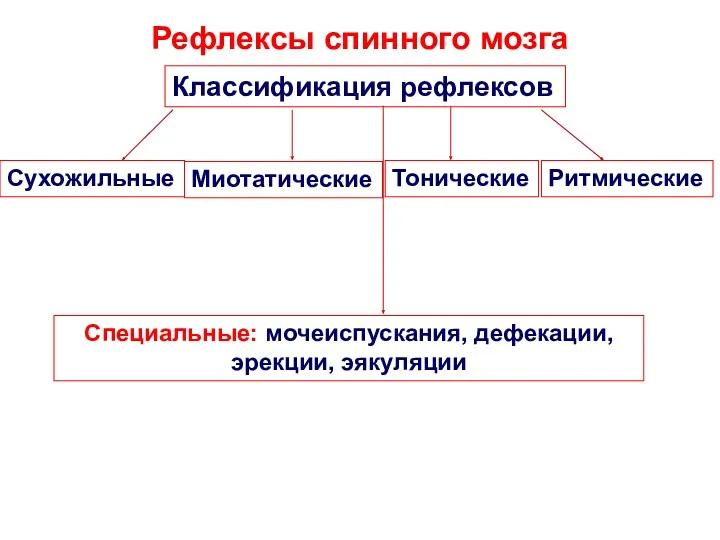
Рефлексы спинного мозга
Классификация рефлексов
Специальные: мочеиспускания, дефекации, эрекции, эякуляции
Сухожильные
Миотатические
Тонические
Ритмические
Рефлексы спинного мозга
Классификация рефлексов
Специальные: мочеиспускания, дефекации, эрекции, эякуляции
Сухожильные
Миотатические
Тонические
Ритмические

Особенности компенсаторных процессов при нарушениях функций спинного мозга
При интенсивном воздействии на
Особенности компенсаторных процессов при нарушениях функций спинного мозга
При интенсивном воздействии на

Длительность восстановления нарушенных спинальных функций зависит:
от обширности и характера травмы;
Длительность восстановления нарушенных спинальных функций зависит:
от обширности и характера травмы;

СИНДРОМ БРОУН-СЕКАРА - (Brown-Sequard syndrome) - неврологическое заболевание, связанное с нарушением
СИНДРОМ БРОУН-СЕКАРА - (Brown-Sequard syndrome) - неврологическое заболевание, связанное с нарушением
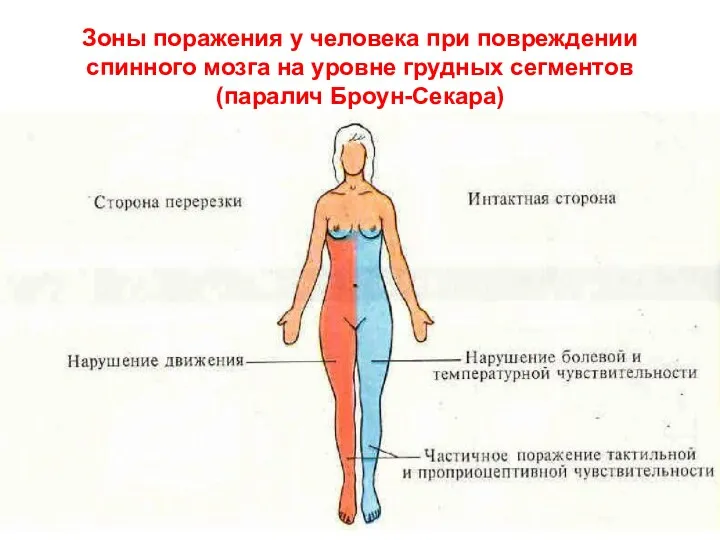
Зоны поражения у человека при повреждении спинного мозга на уровне грудных
Зоны поражения у человека при повреждении спинного мозга на уровне грудных

Ведущими клиническими признаками синдрома Броун-Секара являются:
1. Спастический (центральный) паралич (парез) на
Ведущими клиническими признаками синдрома Броун-Секара являются:
1. Спастический (центральный) паралич (парез) на

4. Утрата болевой и температурной чувствительности по проводниковому типу на контрлатеральной
4. Утрата болевой и температурной чувствительности по проводниковому типу на контрлатеральной

5. Расстройство всех видов чувствительности по сегментарному типу на стороне поражения,
5. Расстройство всех видов чувствительности по сегментарному типу на стороне поражения,
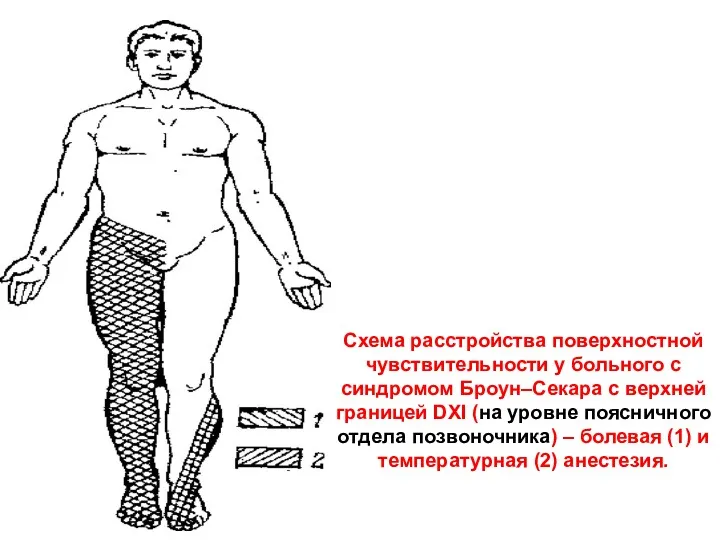
Схема расстройства поверхностной чувствительности у больного с синдромом Броун–Секара с верхней
Схема расстройства поверхностной чувствительности у больного с синдромом Броун–Секара с верхней
 Энергетический обмен. Взаимосвязь обмена веществ и энергии. Лекция №3
Энергетический обмен. Взаимосвязь обмена веществ и энергии. Лекция №3 Влияние условий хранения хлеба на образование плесени на нём
Влияние условий хранения хлеба на образование плесени на нём Урок биологии по теме Бактерии
Урок биологии по теме Бактерии Обмен веществ и энергии в организме
Обмен веществ и энергии в организме Ковыль. Многолетние травы
Ковыль. Многолетние травы Редактирование генома с CRISPR/Cas9
Редактирование генома с CRISPR/Cas9 Энергетика мышечного сокращения. Ферменты в толстых нитях поперечно-полосатых мышц позвоночных
Энергетика мышечного сокращения. Ферменты в толстых нитях поперечно-полосатых мышц позвоночных 9- мінливість
9- мінливість Составление контейнерных композиций из комнатных цветов
Составление контейнерных композиций из комнатных цветов Нормальная микрофлора тела человека
Нормальная микрофлора тела человека Биология бурундука азиатского
Биология бурундука азиатского Биологическое и социальное в человеке
Биологическое и социальное в человеке Асимметрия функциональная головного мозга
Асимметрия функциональная головного мозга Нерв тіні
Нерв тіні Пищеварение. Обмен веществ у человека. (ОГЭ. Тест 4)
Пищеварение. Обмен веществ у человека. (ОГЭ. Тест 4) Понятие о стволовых клетках
Понятие о стволовых клетках Перелетные птицы
Перелетные птицы Биологический уровень организации материи
Биологический уровень организации материи Презентация про ЗОЖ для 5классов.
Презентация про ЗОЖ для 5классов. Мозг и личность
Мозг и личность Энергетикалық алмасудың маңызы
Энергетикалық алмасудың маңызы Скелет. Строение скелета
Скелет. Строение скелета Видозміни пагона та його частин
Видозміни пагона та його частин Строение и жизнедеятельность бактерий
Строение и жизнедеятельность бактерий презентация 10 класс тема Белки-ферменты
презентация 10 класс тема Белки-ферменты Дополнительная презентация Основы аранжировки цветовк элективному курсу Основы фитодизайна, 9 класс
Дополнительная презентация Основы аранжировки цветовк элективному курсу Основы фитодизайна, 9 класс Неспецифічні фактори захисту організму. Імунна система організму
Неспецифічні фактори захисту організму. Імунна система організму Жизнь муравьев
Жизнь муравьев