- Репликация. ДНК

Содержание
- 2. В основе репликации лежит принцип комплементарности В результате репликации образуются дочерние молекулы НК, нуклеотидные последовательности которых
- 3. Типы репликации Полуконсервативный – вновь синтезированная молекула НК состоит из одной материнской и одной дочерней полинуклеотидных
- 5. Белки и ферменты, участвующие в репликации ДНК ДНК-полимеразы ДНК-полимеразы осуществляет синтез ДНК. Субстратом ДНК-полимераз являются дезоксинуклеотид-5’-трифосфаты:
- 6. ДНК-полимераза последовательно добавляет нуклеотиды к 3’ – концу полинуклеотидной цепи
- 7. Синтез новой цепи происходит в направлении от 5’-конца к 3’-концу. ДНК-полимераза может только наращивать цепь ДНК,
- 8. . ДНК-полимераза наращивает цепь в направлении 5’→3’. Для этого ей нужны матрица и затравка
- 9. Задача ДНК-полимеразы – снять точную копию с матрицы ДНК-полимераза проверяет комплементарность каждого нуклеотида дважды: перед включением
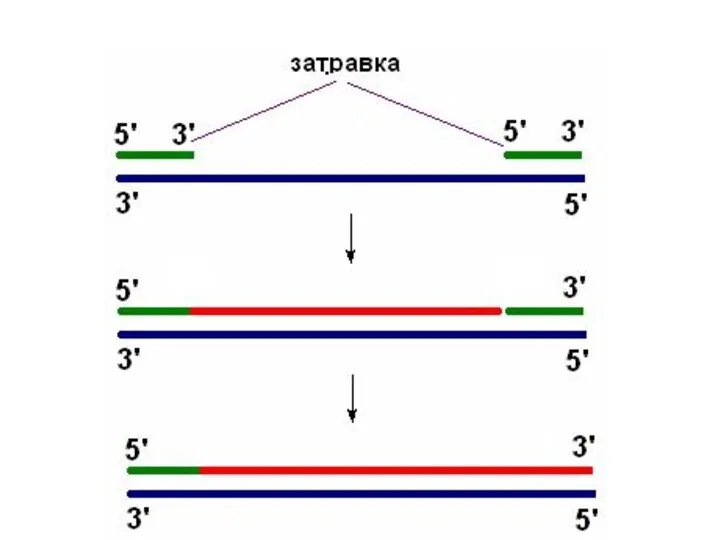
- 11. Праймаза Праймаза – синтезирует праймер (РНК-затравку)
- 12. ДНК-лигаза ДНК-лигаза соединяет 5’-фосфатную и 3’-гидроксильную группы соседних нуклеотидов, в результате образуется фосфодиэфирная связь, ликвидирующая разрыв
- 13. ДНК-хеликаза осуществляет расплетение двойной спирали ДНК, используя энергию гидролиза АТФ Белки, связывающиеся с одноцепочечной ДНК (SSB-белки),
- 14. Механизм репликации Репликация ДНК начинается в определенном месте – в точке начала репликации Репликация от точки
- 15. Бактериальная хромосома имеет одну точку начала репликации, т.е. представляет собой единицу репликации, получившее название репликон Иначе
- 16. ДНК эукариот представлена множеством репликонов
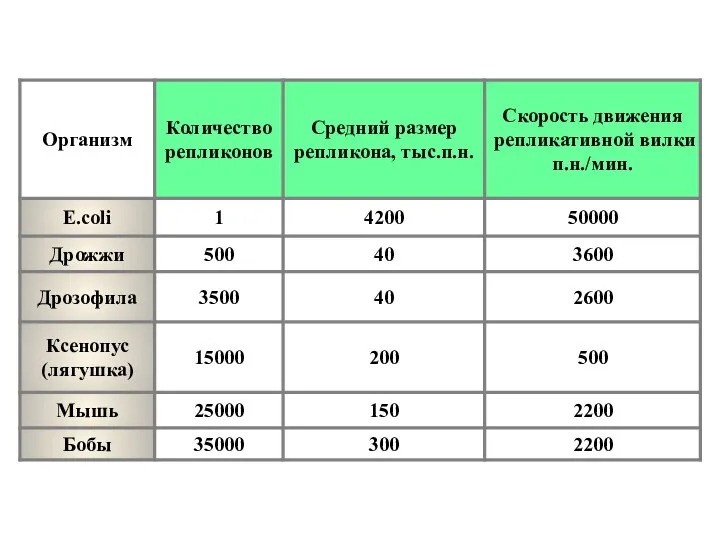
- 18. Механизм репликации у E.coli. ДНК-полимеразы E.coli ДНК-полимераза I. На одной полипептидной цепи содержится 2 активных центра:
- 20. ДНК-полимераза I (кольцеобразная структура, состоящая из нескольких одинаковых молекул белка, показанных разными цветами)
- 21. ДНК-полимераза II. Обладает полимеразной и 3’→ 5’- экзонуклеазной активностями Лучше всего работает на двухцепочечных ДНК с
- 22. ДНК-полимераза III. Обладает полимеразной и 3’→ 5’- экзонуклеазной активностями Основное назначение – репликация ДНК Скорость синтеза
- 23. Хромосома Е.coli имеет одну точку начала репликации (oriC), ее - размер 258 н. п. OriC узнает
- 24. По окончанию синтеза фрагмента Оказаки ДНК-полимераза I за счет 5’→ 3’-экзонуклеазной активности удаляет затравку и заменяет
- 27. Терминация репликации происходит после удвоения кольцевой молекулы ДНК.
- 28. Репликация хромосом у эукариот У эукариот обнаружено несколько ДНК-полимераз: ДНК-полимераза α образует прочный комплекс с праймазой.
- 29. ДНК-полимераза δ обладает полимеразной и 3' → 5'-экзонуклеазной активностями. ДНК-полимераза ε обладает полимеразной и 3' →
- 31. Размер фрагментов Оказаки у эукариот меньше, чем у прокариот и составляет около 100 нуклеотидов. Инициация репликации
- 32. Продвижение репликативной вилки прекращается при столкновении с другой вилкой или при достижении конца хромосомы. в результате
- 33. Репликация вирусов
- 34. Репликация генома аденовирусов (терминальная инициация с использованием нуклеотид-белковой затравки)
- 35. Роль затравки выполняет комплекс терминального белка и одного нуклеотида
- 36. Схема репликации SV-40
- 37. Схема репликации λ-фага (репликация по схеме вторично разматывающегося кольца) В клетке линейная ДНК λ-фага за счет
- 39. Схема репликации генома по схеме катящегося кольца (φX174)
- 40. Схема репликации парвовирусов (терминальная инициация при помощи само-затравочного механизма. Оба конца цепи ДНК способны образовывать шпильки
- 41. Схема репликации генома вируса гриппа Геном вируса гриппа представлен 8 молекулами одноцепочечных –РНК от 900 до
- 42. Схема репликации генома реовирусов Содержат 10 сегментов двухцепочечных РНК, РНК-полимеразу, способную использовать дуплекс для синтеза +РНК
- 43. Ретровирусы РНК-содержащие вирусы. Содержат ревертазу ВИЧ
- 44. Ретровирусы
- 46. Скачать презентацию

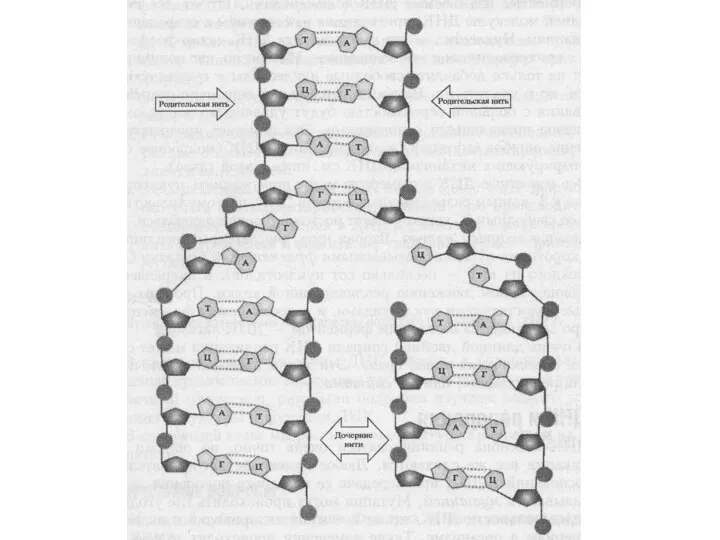
В основе репликации лежит принцип комплементарности
В результате репликации образуются дочерние
В результате репликации образуются дочерние

Типы репликации
Полуконсервативный – вновь синтезированная молекула НК состоит из одной материнской
Типы репликации
Полуконсервативный – вновь синтезированная молекула НК состоит из одной материнской

Белки и ферменты, участвующие в репликации ДНК
ДНК-полимеразы
ДНК-полимеразы осуществляет синтез ДНК.
Субстратом
Белки и ферменты, участвующие в репликации ДНК
ДНК-полимеразы
ДНК-полимеразы осуществляет синтез ДНК.
Субстратом
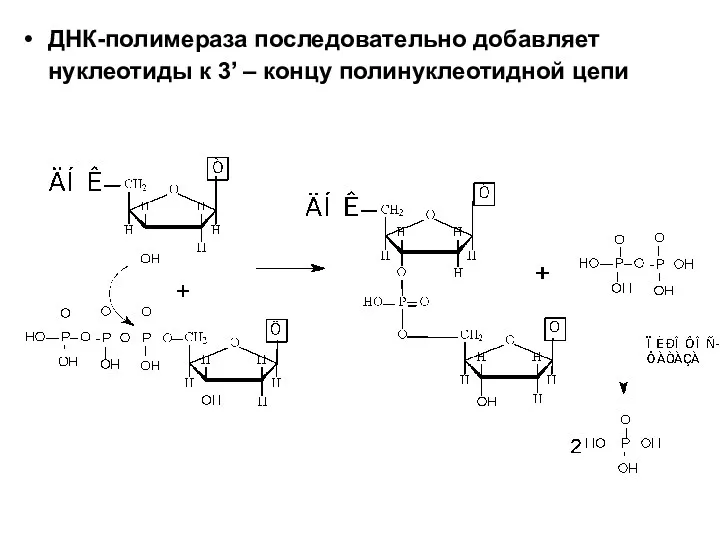
ДНК-полимераза последовательно добавляет нуклеотиды к 3’ – концу полинуклеотидной цепи
ДНК-полимераза последовательно добавляет нуклеотиды к 3’ – концу полинуклеотидной цепи

Синтез новой цепи происходит в направлении от 5’-конца к 3’-концу.
ДНК-полимераза
Синтез новой цепи происходит в направлении от 5’-конца к 3’-концу.
ДНК-полимераза
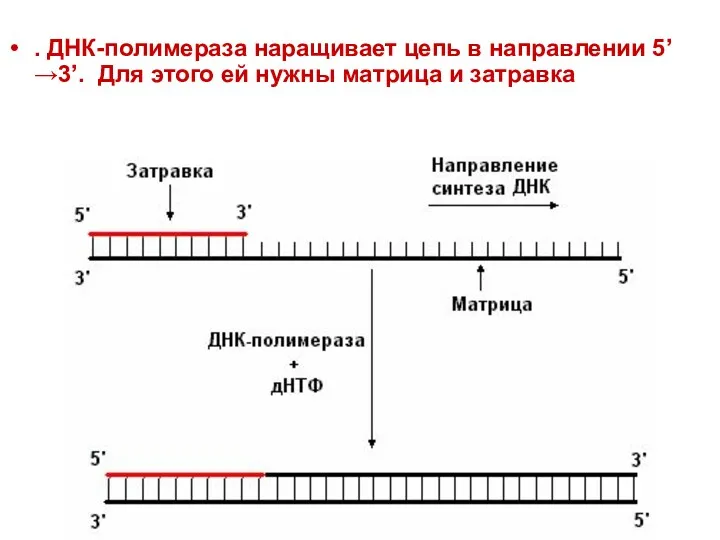
. ДНК-полимераза наращивает цепь в направлении 5’→3’. Для этого ей нужны
. ДНК-полимераза наращивает цепь в направлении 5’→3’. Для этого ей нужны

Задача ДНК-полимеразы – снять точную копию с матрицы
ДНК-полимераза проверяет комплементарность каждого
Задача ДНК-полимеразы – снять точную копию с матрицы
ДНК-полимераза проверяет комплементарность каждого
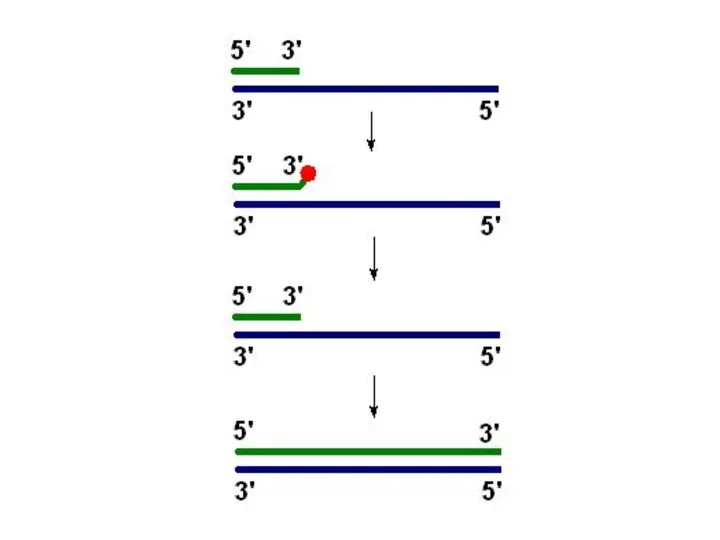

Праймаза
Праймаза – синтезирует праймер (РНК-затравку)
Праймаза
Праймаза – синтезирует праймер (РНК-затравку)
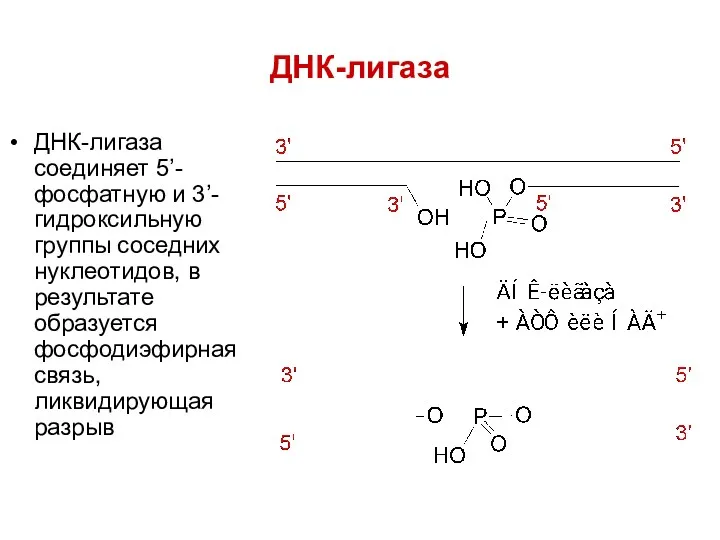
ДНК-лигаза
ДНК-лигаза соединяет 5’-фосфатную и 3’-гидроксильную группы соседних нуклеотидов, в результате образуется
ДНК-лигаза
ДНК-лигаза соединяет 5’-фосфатную и 3’-гидроксильную группы соседних нуклеотидов, в результате образуется
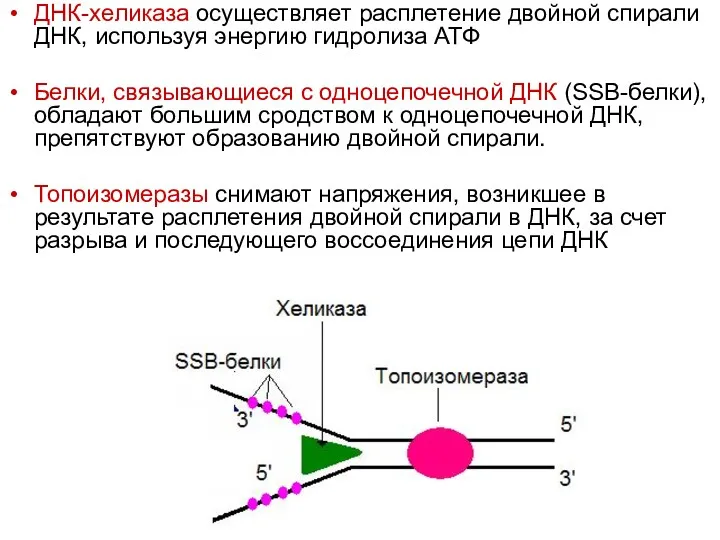
ДНК-хеликаза осуществляет расплетение двойной спирали ДНК, используя энергию гидролиза АТФ
Белки, связывающиеся
ДНК-хеликаза осуществляет расплетение двойной спирали ДНК, используя энергию гидролиза АТФ
Белки, связывающиеся
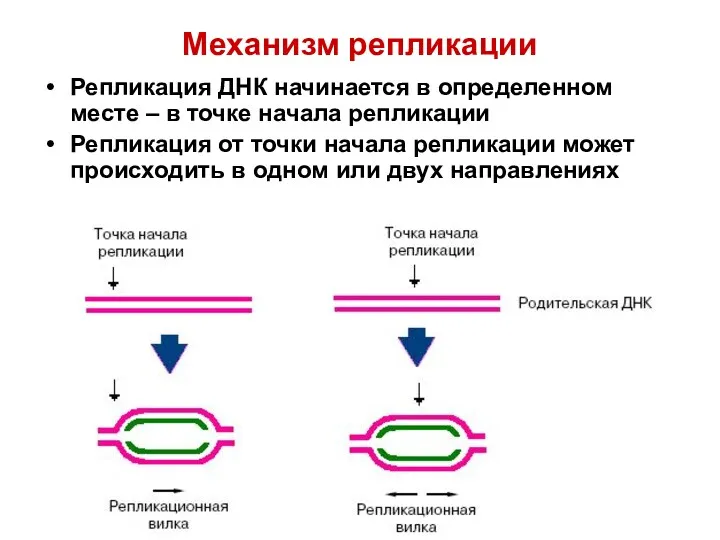
Механизм репликации
Репликация ДНК начинается в определенном месте – в точке начала
Механизм репликации
Репликация ДНК начинается в определенном месте – в точке начала
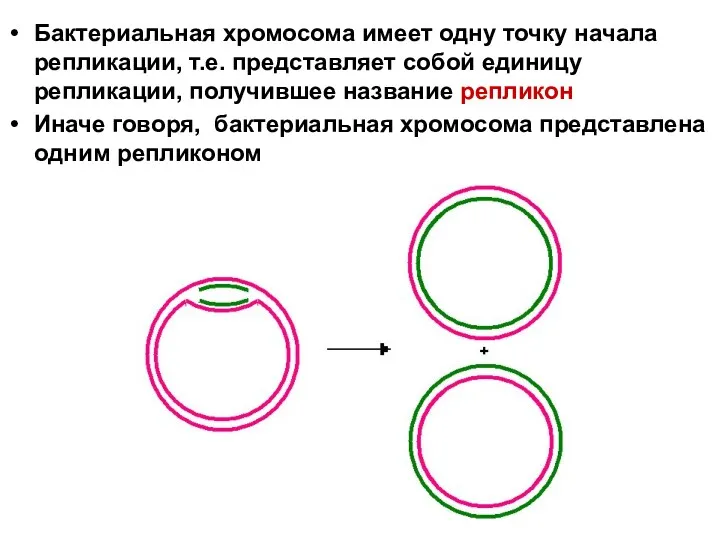
Бактериальная хромосома имеет одну точку начала репликации, т.е. представляет собой единицу
Бактериальная хромосома имеет одну точку начала репликации, т.е. представляет собой единицу
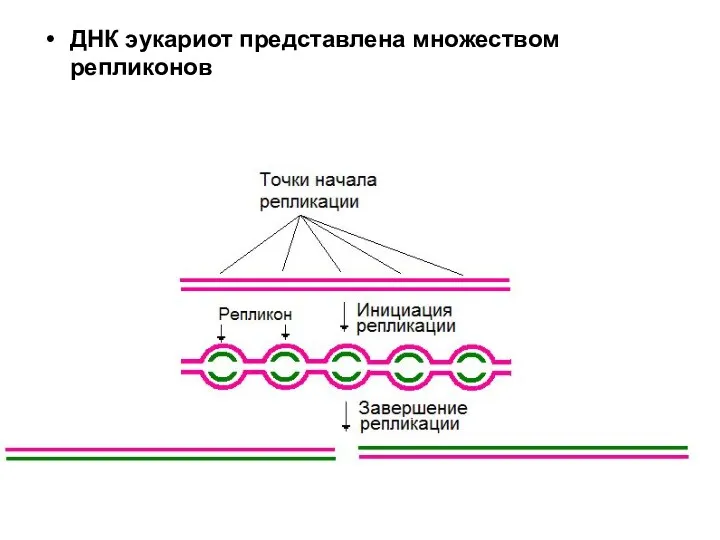
ДНК эукариот представлена множеством репликонов
ДНК эукариот представлена множеством репликонов

Механизм репликации у E.coli.
ДНК-полимеразы E.coli
ДНК-полимераза I.
На одной полипептидной цепи содержится
Механизм репликации у E.coli.
ДНК-полимеразы E.coli
ДНК-полимераза I.
На одной полипептидной цепи содержится
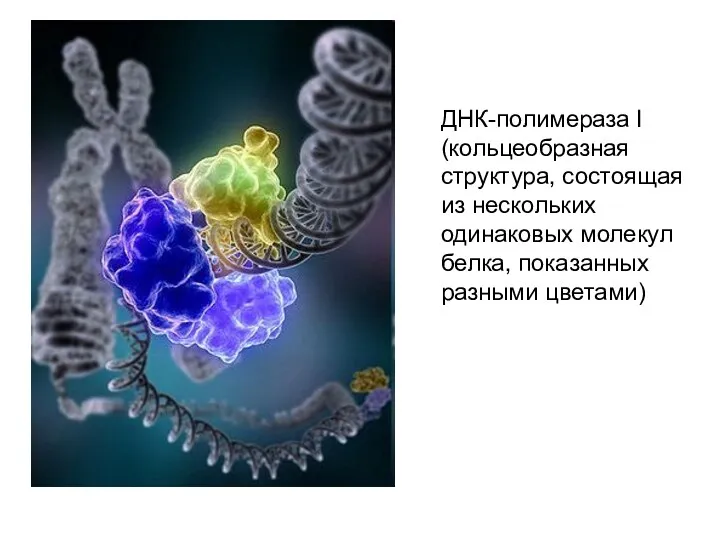
ДНК-полимераза I (кольцеобразная структура, состоящая из нескольких одинаковых молекул белка, показанных
ДНК-полимераза I (кольцеобразная структура, состоящая из нескольких одинаковых молекул белка, показанных

ДНК-полимераза II.
Обладает полимеразной и 3’→ 5’- экзонуклеазной активностями
Лучше всего работает на
ДНК-полимераза II.
Обладает полимеразной и 3’→ 5’- экзонуклеазной активностями
Лучше всего работает на

ДНК-полимераза III.
Обладает полимеразной и 3’→ 5’- экзонуклеазной активностями
Основное назначение – репликация
ДНК-полимераза III.
Обладает полимеразной и 3’→ 5’- экзонуклеазной активностями
Основное назначение – репликация

Хромосома Е.coli имеет одну точку начала репликации (oriC), ее - размер
Хромосома Е.coli имеет одну точку начала репликации (oriC), ее - размер

По окончанию синтеза фрагмента Оказаки ДНК-полимераза I за счет 5’→ 3’-экзонуклеазной
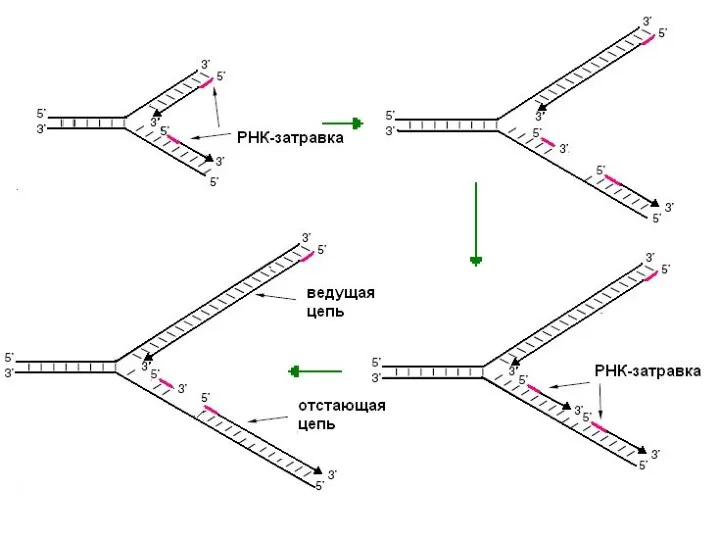
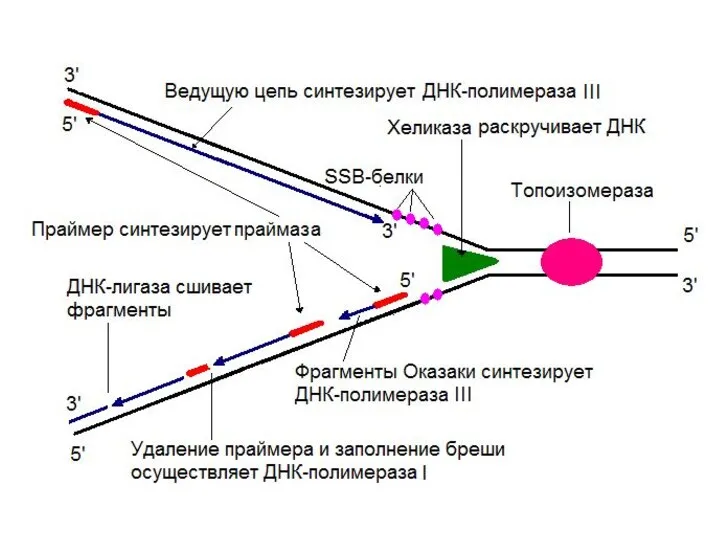

Терминация репликации происходит после удвоения кольцевой молекулы ДНК.

Репликация хромосом у эукариот
У эукариот обнаружено несколько ДНК-полимераз:
ДНК-полимераза α образует
Репликация хромосом у эукариот
У эукариот обнаружено несколько ДНК-полимераз:
ДНК-полимераза α образует

ДНК-полимераза δ обладает полимеразной и 3' → 5'-экзонуклеазной активностями.
ДНК-полимераза ε
ДНК-полимераза ε
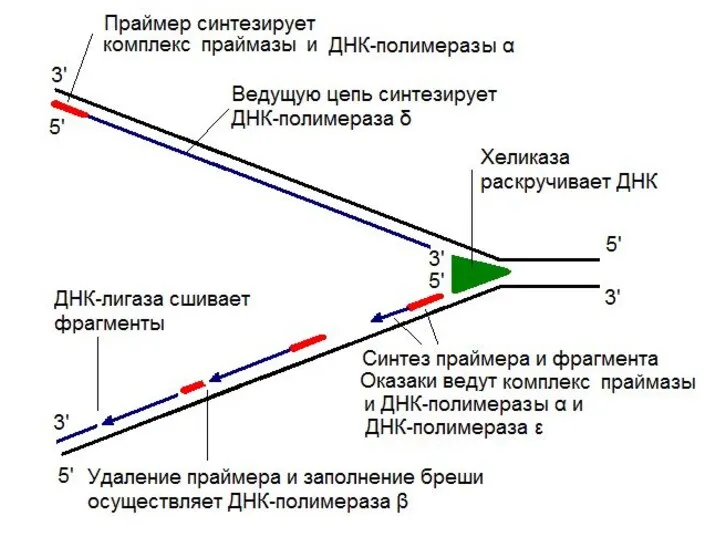
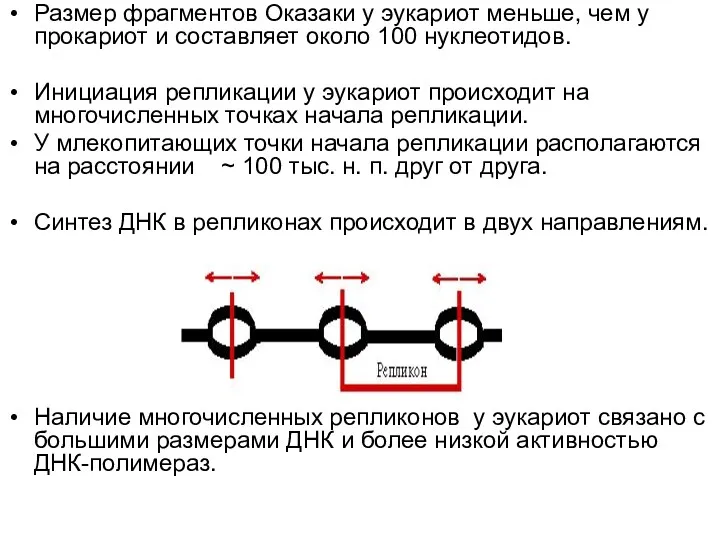
Размер фрагментов Оказаки у эукариот меньше, чем у прокариот и составляет
Размер фрагментов Оказаки у эукариот меньше, чем у прокариот и составляет

Продвижение репликативной вилки прекращается при столкновении с другой вилкой или при
Продвижение репликативной вилки прекращается при столкновении с другой вилкой или при

Репликация вирусов
Репликация вирусов
Репликация генома аденовирусов (терминальная инициация с использованием нуклеотид-белковой затравки)
Репликация генома аденовирусов (терминальная инициация с использованием нуклеотид-белковой затравки)
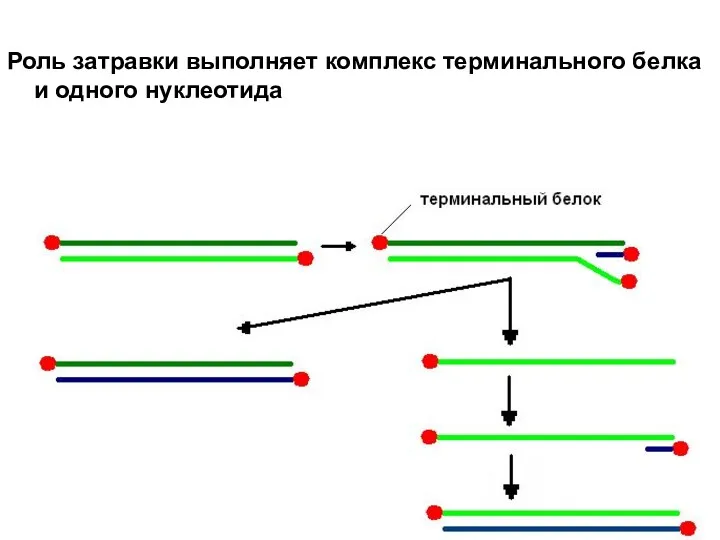
Роль затравки выполняет комплекс терминального белка и одного нуклеотида
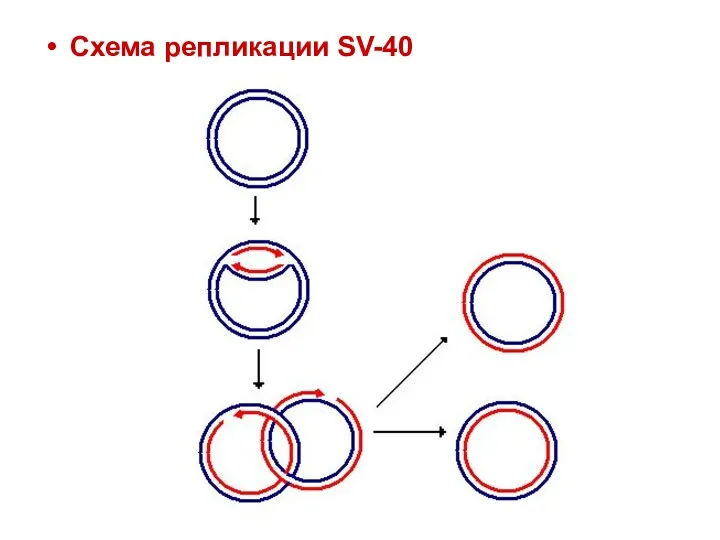
Схема репликации SV-40
Схема репликации SV-40
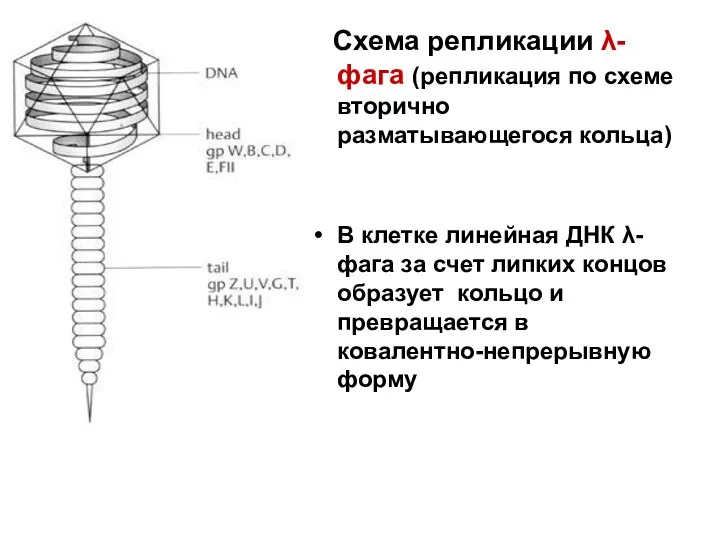
Схема репликации λ-фага (репликация по схеме вторично разматывающегося кольца)
В клетке
Схема репликации λ-фага (репликация по схеме вторично разматывающегося кольца)
В клетке
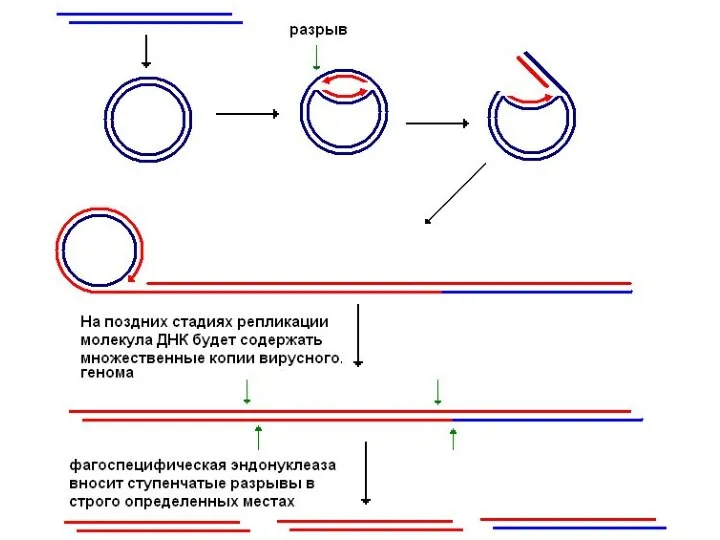
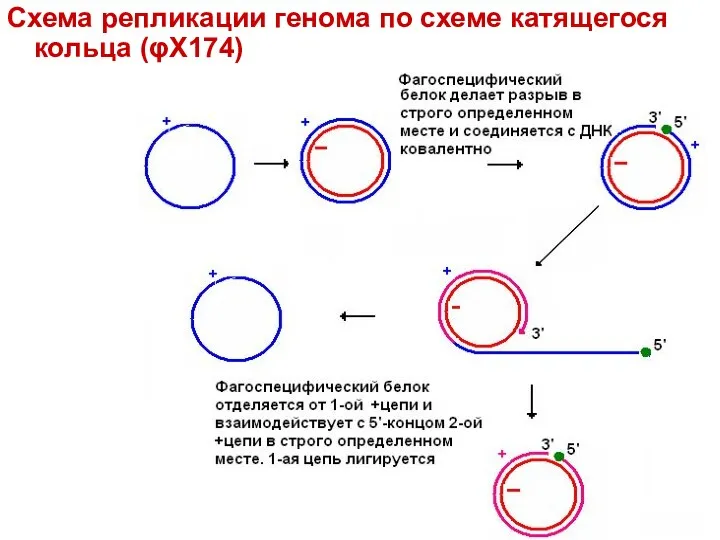
Схема репликации генома по схеме катящегося кольца (φX174)
Схема репликации генома по схеме катящегося кольца (φX174)
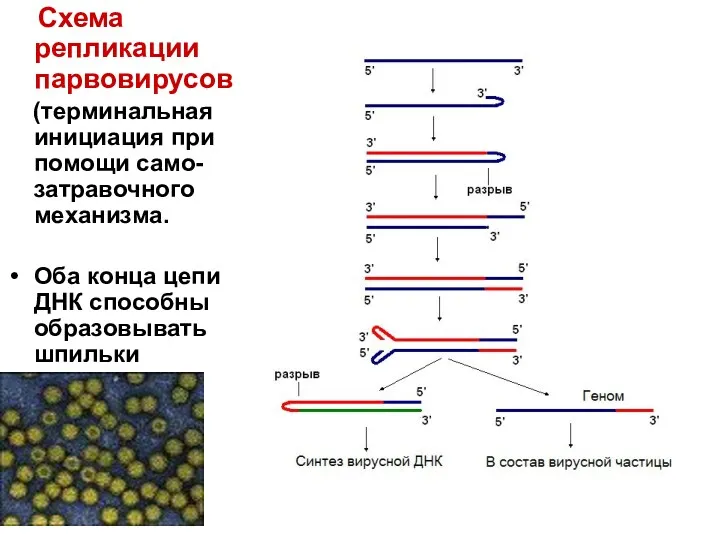
Схема репликации парвовирусов
(терминальная инициация при помощи само-затравочного механизма.
Оба
Схема репликации парвовирусов
(терминальная инициация при помощи само-затравочного механизма.
Оба
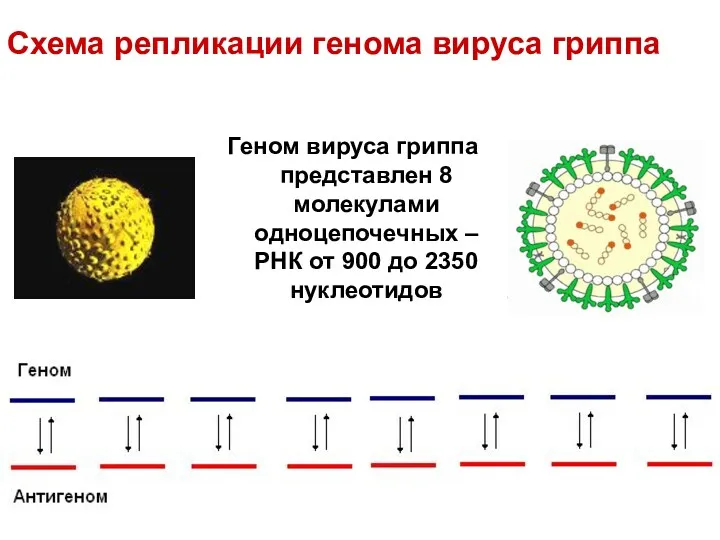
Схема репликации генома вируса гриппа
Геном вируса гриппа представлен 8 молекулами
Схема репликации генома вируса гриппа
Геном вируса гриппа представлен 8 молекулами
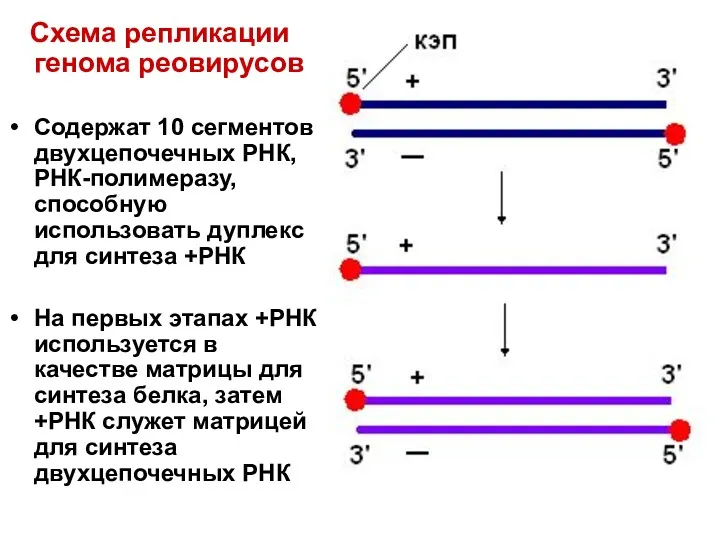
Схема репликации генома реовирусов
Содержат 10 сегментов двухцепочечных РНК, РНК-полимеразу, способную
Схема репликации генома реовирусов
Содержат 10 сегментов двухцепочечных РНК, РНК-полимеразу, способную
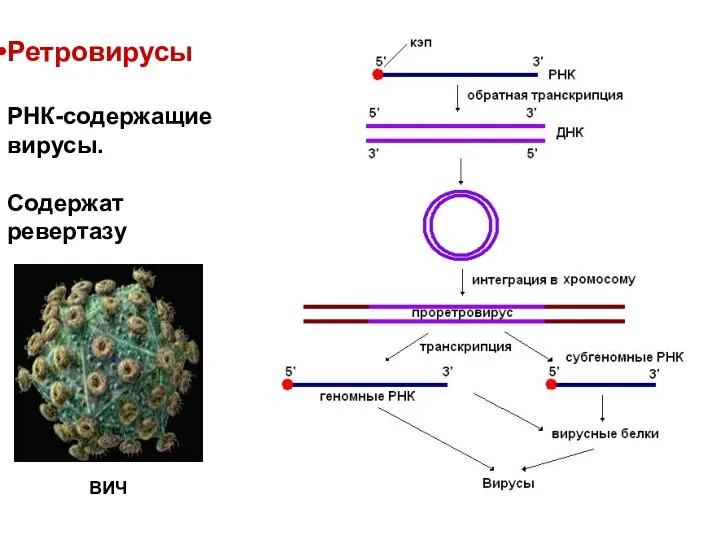
Ретровирусы
РНК-содержащие вирусы.
Содержат ревертазу
ВИЧ
Ретровирусы
РНК-содержащие вирусы.
Содержат ревертазу
ВИЧ
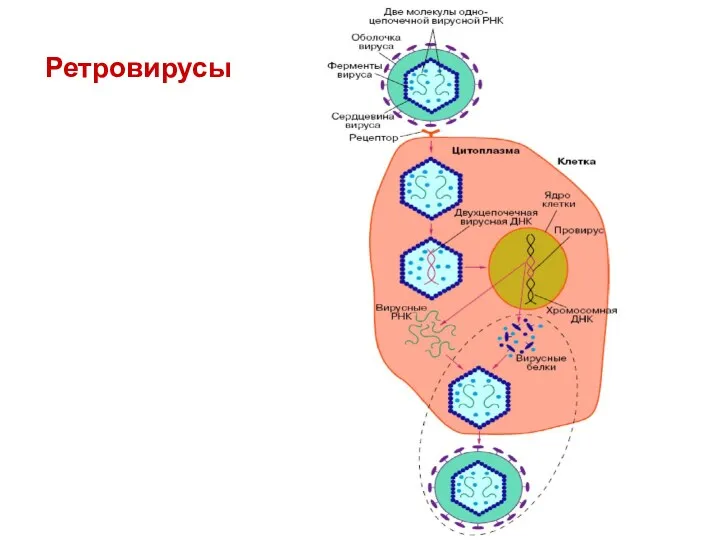
Ретровирусы
Ретровирусы
 Микроэволюция. Вид, его критерии. Видообразование
Микроэволюция. Вид, его критерии. Видообразование Первая и вторая сигнальные системы человека
Первая и вторая сигнальные системы человека Ферменты – 1
Ферменты – 1 Разработка и обоснование системы удобрения в севооборотах ООО Журавушка
Разработка и обоснование системы удобрения в севооборотах ООО Журавушка Биологическая роль деревьев и кустарников в городе
Биологическая роль деревьев и кустарников в городе Использование ЭОР, компьютерных и сетевых технологий на уроках. Опыт работы учителя биологии
Использование ЭОР, компьютерных и сетевых технологий на уроках. Опыт работы учителя биологии Органы мочевыделительной системы
Органы мочевыделительной системы ТН ВЭД: Группа 08. Съедобные фрукты и орехи; кожура цитрусовых плодов или корки дынь
ТН ВЭД: Группа 08. Съедобные фрукты и орехи; кожура цитрусовых плодов или корки дынь 20231016_prezentatsiya
20231016_prezentatsiya Гомеостаз және гормональды реттелу (Координация және реттелу биологиядағы басқару жүйесін сипаттау)
Гомеостаз және гормональды реттелу (Координация және реттелу биологиядағы басқару жүйесін сипаттау) :Сомалық гибридизацияны практикада қолдану
:Сомалық гибридизацияны практикада қолдану ПРЕЗЕНТАЦИИ 5 КЛАСС 1 ЧЕТВЕРТЬ
ПРЕЗЕНТАЦИИ 5 КЛАСС 1 ЧЕТВЕРТЬ Бактериальная клеточная стенка 2 часть
Бактериальная клеточная стенка 2 часть Міні-проєкт Яку воду ми п‘ємо
Міні-проєкт Яку воду ми п‘ємо Особенности дыхания при физических нагрузках
Особенности дыхания при физических нагрузках Пут.нат.6в 12.11.2021, Дикие жив.леса
Пут.нат.6в 12.11.2021, Дикие жив.леса Ріст та розвиток вегетативних органів рослин
Ріст та розвиток вегетативних органів рослин Опорно-двигательный аппарат
Опорно-двигательный аппарат Зачем мы спим ночью. Мир вокруг нас
Зачем мы спим ночью. Мир вокруг нас Выведение пород животных (декоративные или охранные породы)
Выведение пород животных (декоративные или охранные породы) Редуценты и их экологическая роль
Редуценты и их экологическая роль Генетика: основные термины и понятия. 11 тема
Генетика: основные термины и понятия. 11 тема Отдел голосеменные. ОГЭ
Отдел голосеменные. ОГЭ Генетическая безопастность трансгенных растений
Генетическая безопастность трансгенных растений Развитие агрохимии как науки о взаимодействия почвы, растений и удобрений
Развитие агрохимии как науки о взаимодействия почвы, растений и удобрений Аэробное окисление углеводов
Аэробное окисление углеводов Презентация к уроку биологии в 6 классе
Презентация к уроку биологии в 6 классе Электрические свойства нейронов
Электрические свойства нейронов