- Антигены. Антитела

Содержание
- 2. Антигены Это генетически чужеродные вещества, вызывающие в организме разные формы иммунного ответа. Белки (8-10а/к; от 10
- 3. Антигены: чужеродность Это свойство АГ, рассматриваемое по отношению к тому организму, в который АГ попадает КсеноАГ
- 4. Антигены: специфичность Определяется структурными особенностями АГ, придающими ему уникальность и отличающими его от других Заключена в
- 5. Антигены: специфичность
- 6. Антигены: специфичность Валентность антигена – количество идентичных эпитопов Классификация эпитопов: 1. Линейные (секвенциальные) - аминокислотная последовательность
- 7. Антигены: антигенность Определяет способность вызывать иммунный ответ в организме. Различные АГ вызывают разный по силе и
- 8. Антигены: иммуногенность Способность АГ формировать иммунитет, или иммунологическую память Важна для АГ патогенных микроорганизмов, будет определять
- 9. Виды антигенов 1. Полноценные (иммуногены) Вызывают иммунный ответ 2. Неполноценные (гаптены и толерогены) Не распознаются иммунокомпетентными
- 10. Виды антигенов Аллергены – АГ, вызывающие наработку IgE, опосредующего развитие аллергической реакции Тимусзависимые АГ – вызывают
- 11. Виды антигенов Суперантигены – способны непосредственно и без предварительной «переработки» АПК взаимодействовать с молекулами МНС II
- 12. Структуры, активирующие клетки врожденного иммунитета Патоген-ассоциированные молекулярные паттерны (образы) — РАМР (англ. pathogen-associated molecular patterns). Главными
- 13. Структуры, активирующие клетки врожденного иммунитета Эндогенные сигналы биологической опасности — DAMP (англ. danger associated molecular patterns),
- 14. АНТИТЕЛА АТ – гликопротеиновые молекулы, относящиеся к семейству Ig, способные специфически связываться с АГ. Являются основными
- 15. Расшифрована Р. Портером и Д. Эдельманом в 1958 г VL VL VH CL CH1 CH2 CH3
- 16. Fab-фрагменты F(ab)2 Fc-фрагмент Активные центры Структура иммуноглобулинов
- 17. Структура Ig две легкие L-цепи (от англ. Light — легкий) и две тяжелые Н-цепи (от англ.
- 18. Функции доменов VL и VH – формируют активный центр АТ за счет гипервариабельных последовательностей аминокислот; т.е.
- 19. Функции доменов СL и СH1 – участвуют в формировании шарнирной области иммуноглобулина; СH2 – участвует в
- 20. Взаимодействие АГ и АТ Пространственная комплементарность между паратопом АТ (антигенсвязывающим участком, образованным V доменами H- и
- 21. Взаимодействие АГ и АТ Аффинность – сила связывания одного эпитопа с оним активным центром Ig Авидность
- 22. Функции АТ Специфическое связывание АГ за счет комплементарных вариабельных участков Fab-фрагмента Эффекторные (вторичные) функции, реализуемые через
- 23. Функции АТ
- 24. Функции АТ: активация фагоцитоза
- 25. Функции АТ: АЗКЦ
- 26. Функции АТ: дегрануляция тучных клеток
- 27. Классы иммуноглобулинов IgG - отвечает за общий инфекционный иммунитет IgM - отвечает за общий инфекционный иммунитет
- 28. Иммуноглобулин G
- 29. Иммуноглобулин G Строение Н-цепи – γ, состоит из 4 доменов; строение L-цепи – κ или λ;
- 30. Иммуноглобулин М J - цепь
- 31. Иммуноглобулин М Строение Н-цепи – μ (состоит из 5 доменов); строение легкой L-цепи – κ или
- 32. Иммуноглобулин М Активация комплемента по классическому пути (IgM >> IgG) Не проходит через плаценту Участвует в
- 33. Иммуноглобулин А Секреторный IgA (sIgA) Секреторный компонент J-цепь Сывороточный IgA
- 34. Иммуноглобулин А Строение Н-цепи – α; строение L-цепи – κили λ; существует в сыворотке в виде
- 35. Иммуноглобулин А Концентрация в крови - 0,5- 3 г/л; Период полувыведения из крови – 6 дней;
- 36. Иммуноглобулин D
- 37. Иммуноглобулин D Строение Н-цепи – δ (состоит из 5 доменов); строение L-цепи –κили λ (состоит из
- 38. Иммуноглобулин Е
- 39. Иммуноглобулин Е Строение Н-цепи – ε (состоит из 5 доменов); строение легкой L-цепи – κ или
- 40. Возрастные изменения концентрации иммуноглобулинов в крови человека Содержание IgM в сыворотке крови ребенка достигает нормы взрослого
- 42. Скачать презентацию

Антигены
Это генетически чужеродные вещества, вызывающие в организме разные формы иммунного ответа.
Белки
Антигены
Это генетически чужеродные вещества, вызывающие в организме разные формы иммунного ответа.
Белки

Антигены: чужеродность
Это свойство АГ, рассматриваемое по отношению к тому организму, в
Антигены: чужеродность
Это свойство АГ, рассматриваемое по отношению к тому организму, в

Антигены: специфичность
Определяется структурными особенностями АГ, придающими ему уникальность и отличающими его
Антигены: специфичность
Определяется структурными особенностями АГ, придающими ему уникальность и отличающими его
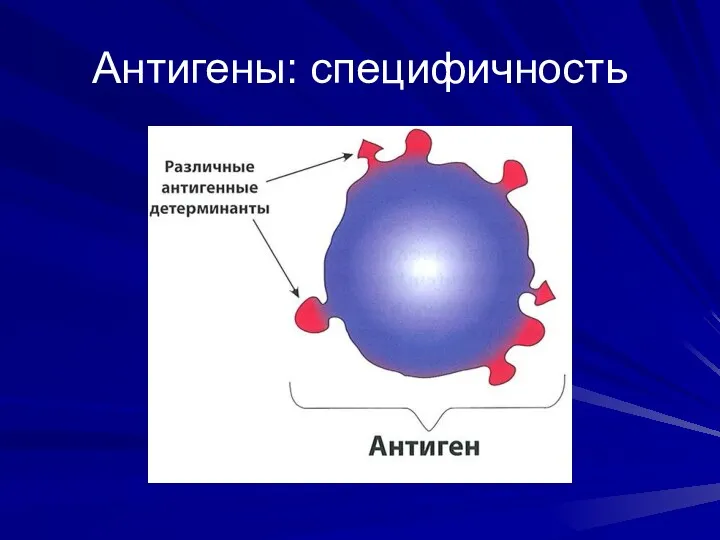
Антигены: специфичность
Антигены: специфичность

Антигены: специфичность
Валентность антигена – количество идентичных эпитопов
Классификация эпитопов:
1. Линейные (секвенциальные) -
Антигены: специфичность
Валентность антигена – количество идентичных эпитопов
Классификация эпитопов:
1. Линейные (секвенциальные) -

Антигены: антигенность
Определяет способность вызывать иммунный ответ в организме.
Различные АГ вызывают разный
Антигены: антигенность
Определяет способность вызывать иммунный ответ в организме.
Различные АГ вызывают разный

Антигены: иммуногенность
Способность АГ формировать иммунитет, или иммунологическую память
Важна для АГ патогенных
Антигены: иммуногенность
Способность АГ формировать иммунитет, или иммунологическую память
Важна для АГ патогенных

Виды антигенов
1. Полноценные (иммуногены)
Вызывают иммунный ответ
2. Неполноценные (гаптены и толерогены)
Не распознаются
Виды антигенов
1. Полноценные (иммуногены)
Вызывают иммунный ответ
2. Неполноценные (гаптены и толерогены)
Не распознаются

Виды антигенов
Аллергены – АГ, вызывающие наработку IgE, опосредующего развитие аллергической реакции
Тимусзависимые
Виды антигенов
Аллергены – АГ, вызывающие наработку IgE, опосредующего развитие аллергической реакции
Тимусзависимые

Виды антигенов
Суперантигены – способны непосредственно и без предварительной «переработки» АПК
Виды антигенов
Суперантигены – способны непосредственно и без предварительной «переработки» АПК

Структуры, активирующие клетки врожденного иммунитета
Патоген-ассоциированные
молекулярные паттерны (образы) — РАМР
(англ. pathogen-associated molecular
Структуры, активирующие клетки врожденного иммунитета
Патоген-ассоциированные молекулярные паттерны (образы) — РАМР (англ. pathogen-associated molecular

Структуры, активирующие клетки врожденного иммунитета
Эндогенные сигналы биологической опасности — DAMP (англ.
Структуры, активирующие клетки врожденного иммунитета
Эндогенные сигналы биологической опасности — DAMP (англ.

АНТИТЕЛА
АТ – гликопротеиновые молекулы, относящиеся к семейству Ig, способные специфически связываться
АНТИТЕЛА
АТ – гликопротеиновые молекулы, относящиеся к семейству Ig, способные специфически связываться
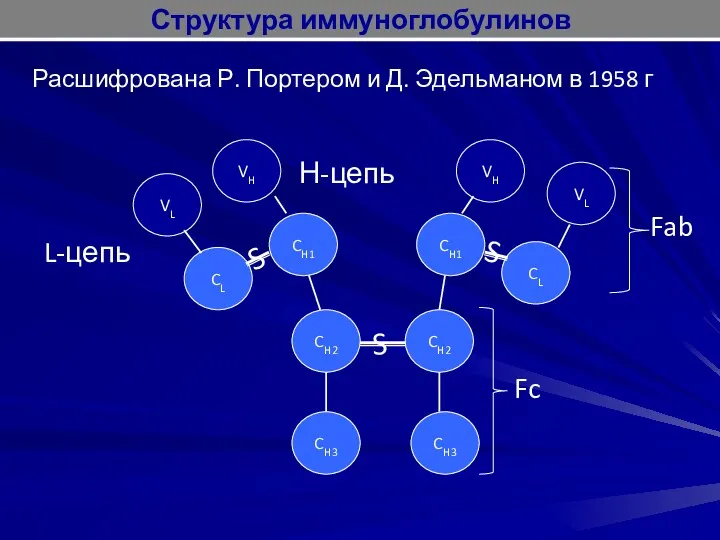
Расшифрована Р. Портером и Д. Эдельманом в 1958 г
VL
VL
VH
CL
CH1
CH2
CH3
CL
VH
CH1
CH2
CH3
Fc
Fab
L-цепь
Н-цепь
S
S
S
Структура иммуноглобулинов
Расшифрована Р. Портером и Д. Эдельманом в 1958 г
VL
VL
VH
CL
CH1
CH2
CH3
CL
VH
CH1
CH2
CH3
Fc
Fab
L-цепь
Н-цепь
S
S
S
Структура иммуноглобулинов
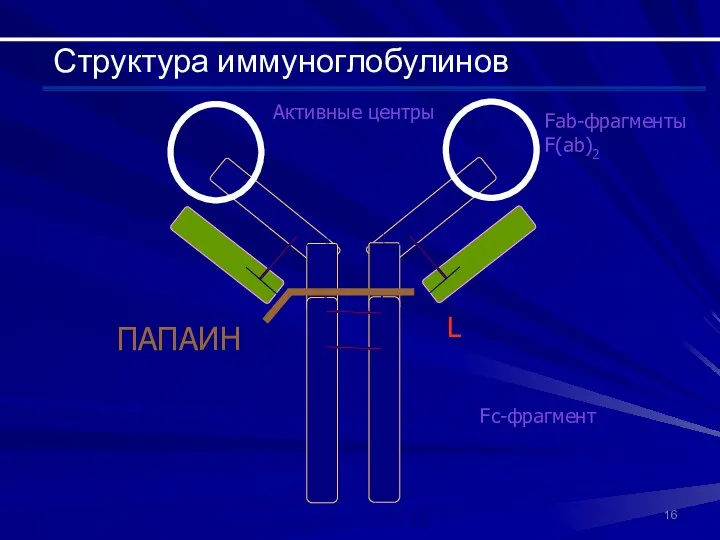
Fab-фрагменты F(ab)2
Fc-фрагмент
Активные центры
Структура иммуноглобулинов
Fab-фрагменты F(ab)2
Fc-фрагмент
Активные центры
Структура иммуноглобулинов

Структура Ig
две легкие L-цепи (от англ. Light — легкий) и две
Структура Ig
две легкие L-цепи (от англ. Light — легкий) и две

Функции доменов
VL и VH – формируют активный центр АТ за счет
Функции доменов
VL и VH – формируют активный центр АТ за счет

Функции доменов
СL и СH1 – участвуют в формировании шарнирной области иммуноглобулина;
СH2
Функции доменов
СL и СH1 – участвуют в формировании шарнирной области иммуноглобулина;
СH2

Взаимодействие АГ и АТ
Пространственная комплементарность между паратопом АТ (антигенсвязывающим участком, образованным
Взаимодействие АГ и АТ
Пространственная комплементарность между паратопом АТ (антигенсвязывающим участком, образованным

Взаимодействие АГ и АТ
Аффинность – сила связывания одного эпитопа с оним
Взаимодействие АГ и АТ
Аффинность – сила связывания одного эпитопа с оним

Функции АТ
Специфическое связывание АГ за счет комплементарных вариабельных участков Fab-фрагмента
Эффекторные (вторичные)
Функции АТ
Специфическое связывание АГ за счет комплементарных вариабельных участков Fab-фрагмента
Эффекторные (вторичные)
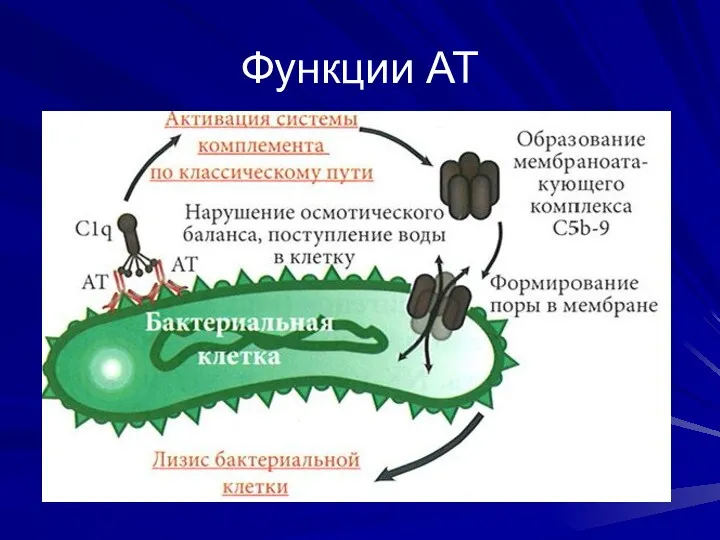
Функции АТ
Функции АТ
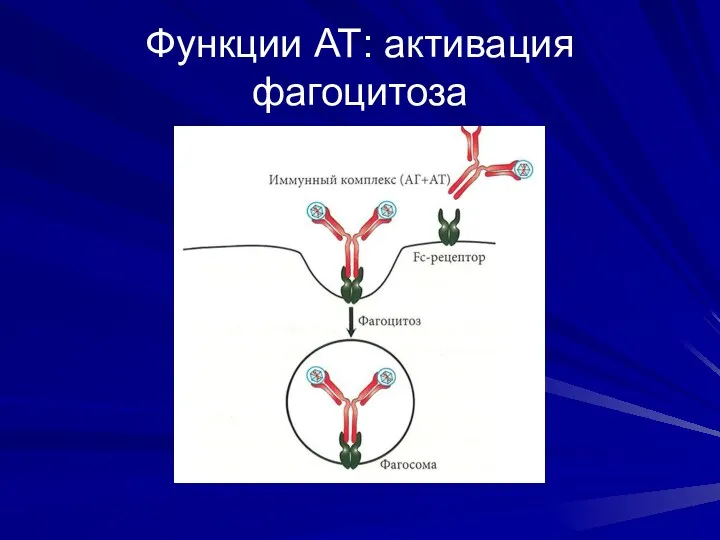
Функции АТ: активация фагоцитоза
Функции АТ: активация фагоцитоза
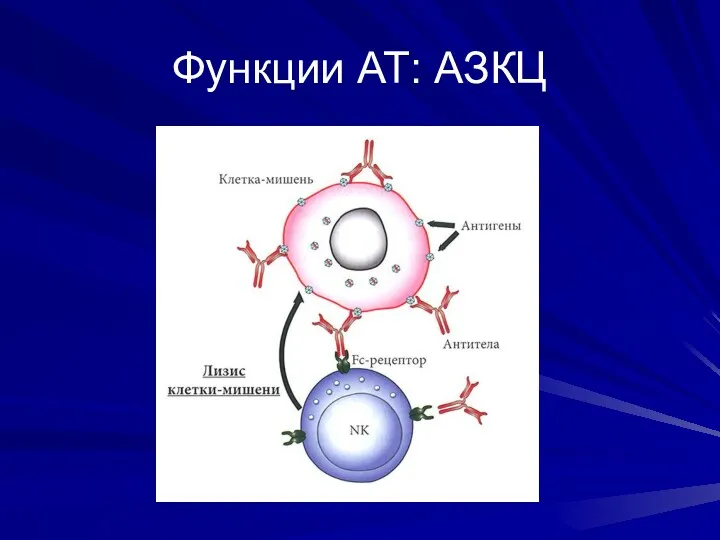
Функции АТ: АЗКЦ
Функции АТ: АЗКЦ
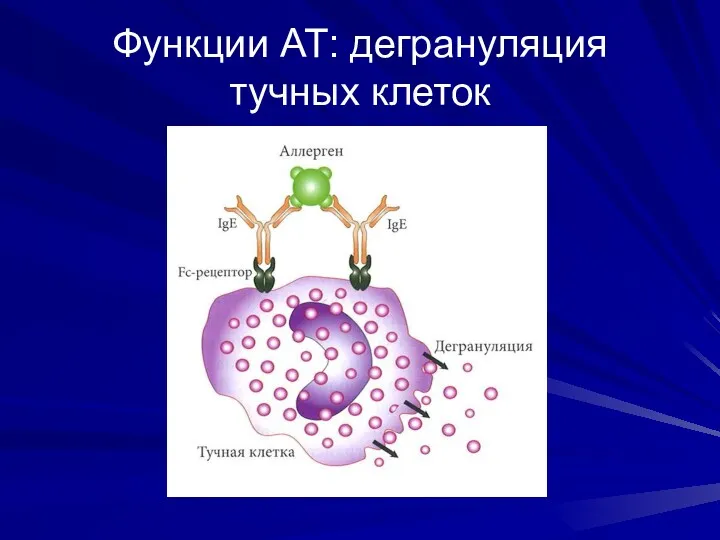
Функции АТ: дегрануляция тучных клеток
Функции АТ: дегрануляция тучных клеток

Классы иммуноглобулинов
IgG - отвечает за общий инфекционный иммунитет
IgM - отвечает за
Классы иммуноглобулинов
IgG - отвечает за общий инфекционный иммунитет
IgM - отвечает за
Иммуноглобулин G
Иммуноглобулин G

Иммуноглобулин G
Строение Н-цепи – γ, состоит из 4 доменов; строение L-цепи
Иммуноглобулин G
Строение Н-цепи – γ, состоит из 4 доменов; строение L-цепи
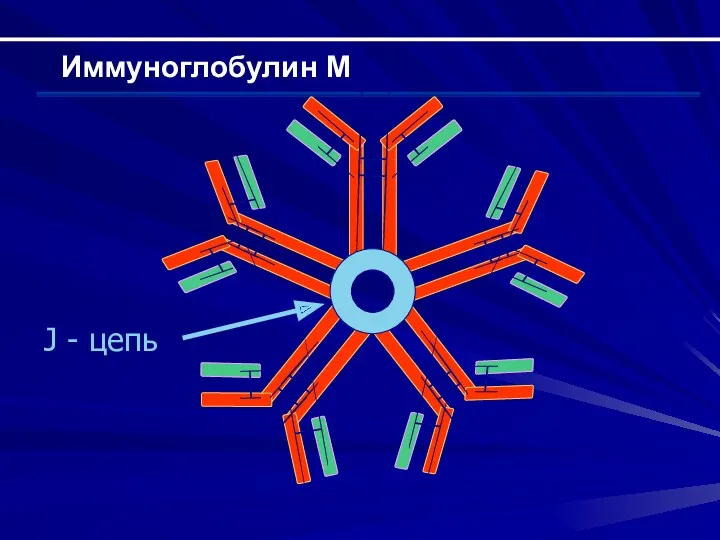
Иммуноглобулин М
J - цепь
Иммуноглобулин М
J - цепь

Иммуноглобулин М
Строение Н-цепи – μ (состоит из 5 доменов); строение легкой
Иммуноглобулин М
Строение Н-цепи – μ (состоит из 5 доменов); строение легкой

Иммуноглобулин М
Активация комплемента по классическому пути (IgM >> IgG)
Не проходит через
Иммуноглобулин М
Активация комплемента по классическому пути (IgM >> IgG)
Не проходит через
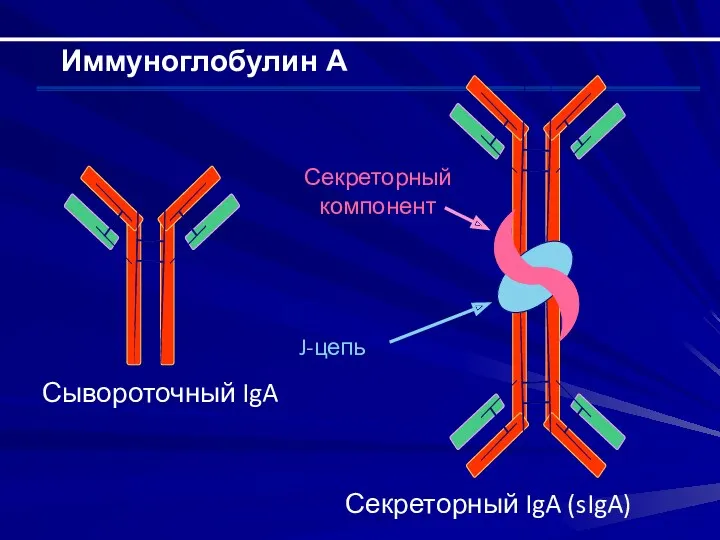
Иммуноглобулин А
Секреторный IgA (sIgA)
Секреторный компонент
J-цепь
Сывороточный IgA
Иммуноглобулин А
Секреторный IgA (sIgA)
Секреторный компонент
J-цепь
Сывороточный IgA

Иммуноглобулин А
Строение Н-цепи – α; строение L-цепи – κили λ;
существует
Иммуноглобулин А
Строение Н-цепи – α; строение L-цепи – κили λ;
существует

Иммуноглобулин А
Концентрация в крови - 0,5- 3 г/л;
Период полувыведения из крови
Иммуноглобулин А
Концентрация в крови - 0,5- 3 г/л;
Период полувыведения из крови
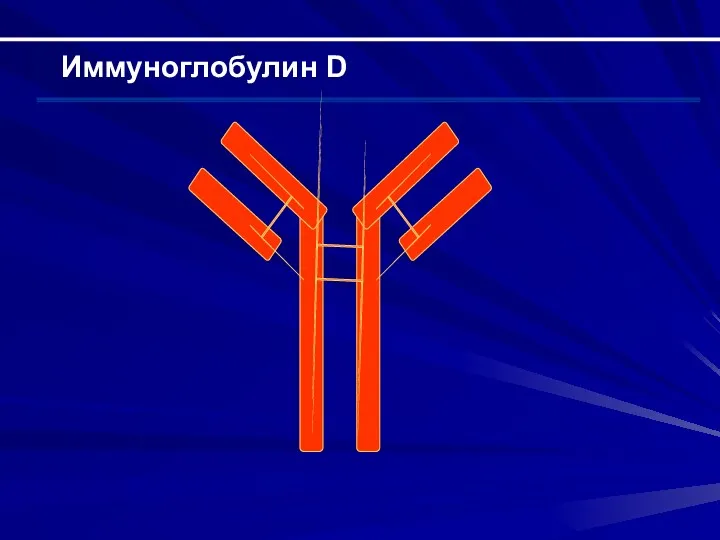
Иммуноглобулин D
Иммуноглобулин D

Иммуноглобулин D
Строение Н-цепи – δ (состоит из 5 доменов); строение L-цепи
Иммуноглобулин D
Строение Н-цепи – δ (состоит из 5 доменов); строение L-цепи
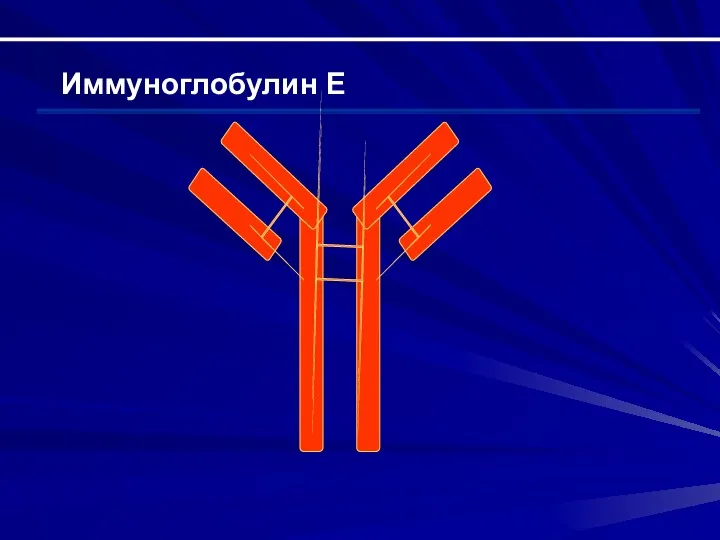
Иммуноглобулин Е
Иммуноглобулин Е

Иммуноглобулин Е
Строение Н-цепи – ε (состоит из 5 доменов); строение легкой
Иммуноглобулин Е
Строение Н-цепи – ε (состоит из 5 доменов); строение легкой
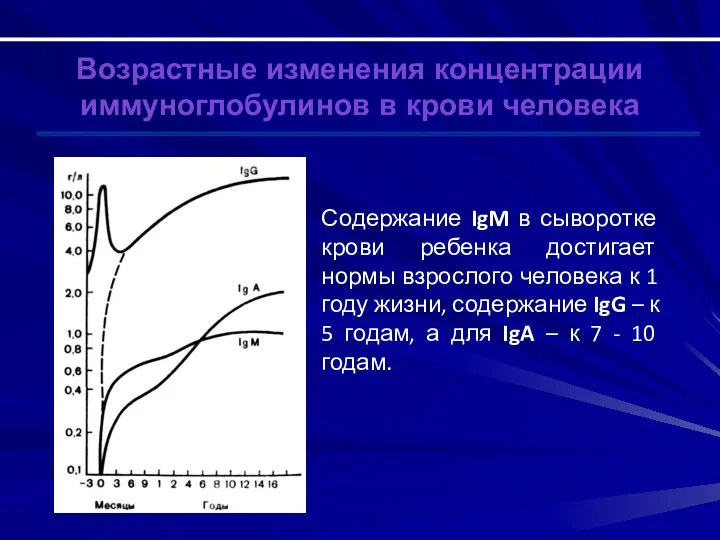
Возрастные изменения концентрации иммуноглобулинов в крови человека
Содержание IgM в сыворотке крови
Возрастные изменения концентрации иммуноглобулинов в крови человека
Содержание IgM в сыворотке крови
 Чувствительная система. Анатомофизиологические особенности чувствительного анализатора. Виды нарушений чувствительности
Чувствительная система. Анатомофизиологические особенности чувствительного анализатора. Виды нарушений чувствительности Иммунная система и клеточные взаимодействия в иммунных реакциях
Иммунная система и клеточные взаимодействия в иммунных реакциях Остеохондропатии
Остеохондропатии Трансплантация печени
Трансплантация печени Ошибки и осложнения, возникающие при лечении осложненного кариеса
Ошибки и осложнения, возникающие при лечении осложненного кариеса Особо опасные инфекции
Особо опасные инфекции Шок
Шок Глюкокортикоиды. Клиническая фармакология
Глюкокортикоиды. Клиническая фармакология Самойлович Данила Самойлович (1744-1805)
Самойлович Данила Самойлович (1744-1805) Врождённые пороки развития конечностей. Врождённые деформации шеи и грудной клетки
Врождённые пороки развития конечностей. Врождённые деформации шеи и грудной клетки Эволюция хирургического лечения рака молочной железы
Эволюция хирургического лечения рака молочной железы Методика аускультации сердца
Методика аускультации сердца Филогенез бранхиогенной группы желез
Филогенез бранхиогенной группы желез Причины привычного невынашивания беременности
Причины привычного невынашивания беременности Нормативно-правовые основы профессиональной деятельности младшего медицинского персонала
Нормативно-правовые основы профессиональной деятельности младшего медицинского персонала АФО. Органов пищеварения у детей. Семиотика поражений. Методы диагностики при заболеваниях органов пищеварения
АФО. Органов пищеварения у детей. Семиотика поражений. Методы диагностики при заболеваниях органов пищеварения Интервенционный ультразвук в лечении абсцессов брюшной полости и полости малого таза
Интервенционный ультразвук в лечении абсцессов брюшной полости и полости малого таза Аномалии конституции у детей. Диатезы
Аномалии конституции у детей. Диатезы Общая анестезия. Подготовка больного. Операционно-анестезиологический риск
Общая анестезия. Подготовка больного. Операционно-анестезиологический риск Үшкіл жүйке жүйесінің невралгиясын емдеудегі тиімді әдістерге талдау жасау
Үшкіл жүйке жүйесінің невралгиясын емдеудегі тиімді әдістерге талдау жасау Диагностика отосклероза по данным МСКТ
Диагностика отосклероза по данным МСКТ Диссеминированный туберкулез легких
Диссеминированный туберкулез легких Протей экологиясы, лабораторлы диагностикасы, профилактикасы мен емі
Протей экологиясы, лабораторлы диагностикасы, профилактикасы мен емі Жатырдан тыс жүктіліктің хирургиялық емнен кейінгі науқастардың гинекологиялық реабилитациясы. Бедеу неке
Жатырдан тыс жүктіліктің хирургиялық емнен кейінгі науқастардың гинекологиялық реабилитациясы. Бедеу неке Сестринский уход при заболеваниях опорнодвигательного аппарата
Сестринский уход при заболеваниях опорнодвигательного аппарата Фосфат- диабет
Фосфат- диабет Расстройства личности
Расстройства личности Вегетативные дисфункции у детей
Вегетативные дисфункции у детей