- Герпесвирусы

Содержание
- 2. нейротропні І ІІ ІІІ типи HERPESVIRUSES лімфотропні ІУ УІ УІІ УІІІ типи вісцеротропні У тип Всі
- 3. Дисемінація герпесвірусів в організмі людини: від клітини до клітини, гематогенно, транслімфатично та трансневрально (трансаксонально). Зараження плоду
- 4. Поширення герпесвірусів Більшість населення планети - у стані «здорового» носійства Віруси «ховаються» від імунної системи в
- 6. Віруси впливають на життєво важливі функції макроорганізму імунобіологічні системи його захисту (інтерферон–цитокіновий статус, клітинні та гуморальні
- 7. Онкопотенціал герпесвірусів: • Лімфома Беркітта (HHV–4). • Назофарингеальна карцинома (HHV–4). • Лімфоми, лімфосаркоми (HHV–4). • Аденокарцинома
- 8. Вірус Люке В природніх умовах вірус викликає аденокарциноми нирок леопардових жаб, які мешкають у водоймах Північної
- 9. Хвороба Марека Хвороба Марека – висококонтагіозне лімфопроліферативне захворювання курей Характерні ознаки – розвиток Т-клітинних лімфом та
- 10. Онкогенні герпесвіруси з роду Mardivirus Існують три серотипи вірусу хвороби Марека: Серотип 1 (Gallid herpesvirus-2) Уражує
- 11. Інкубаційний період – 13 – 150 днів Клінічна картина Класична (хромота, відвисання крил і хвоста, зміна
- 12. Передача і життєвий цикл Курча вдихає вірус Вірус потрапляє у макрофаги та дендритні клітини легенів фагоцитозом
- 13. Патогенез Виділяють чотири фази інфекції гостра первинна інфекція та дегенеративні зміни ранньої фази (селезінка, тимус, фабріцієва
- 14. Патанатомія Атрофія м’язів Дегенеративні ушкодження периферичних нервів Лімфоматозні утворення сало-подібної консистенції у гонадах, печінці, клітковині шкіри,
- 15. Еволюція патологічного вплву Гострота та складність протікання захворювання Хронічний поліневрит Вісцеральна лімфома Імуносупресія Тимчасовий параліч Набрякання
- 16. Загальні відомості про трансформацію Вірус здатний до трансформації СD4+ Т-клітини, в яких той перебуває у латентному
- 17. Геном
- 18. Порівняння геномів вірусу varicella–zoster virus (VZV) та вірусу хвороби Марека (MDV) VZV – найближчій родич MDV,
- 19. Регуляція МНС класу 1 та ІНФ-α Вірус здатний “вимикати” експресію білків необхідних для асоціації ендогенних пептидів
- 20. Регуляція МНС класу 1 Молекулярний механізм Ген UL49.5 кодує білок який здатний: блокувати ТАР білки, які
- 21. Латентний стан та трансформація Розрізнити Т-клітини, які несуть вірус у латентному стані, але ще не є
- 22. Загальні відомості про трансформацію Вірус здатний до трансформації СD4+ Т-клітини, в яких той перебуває у латентному
- 23. Деякі вірусні протеїни, що експресуються у трансформованих Т-лімфоцитах pp38 – 24; 38 або 41 кДа –
- 24. Регуляція МНС класу 1 та ІНФ-α Вірус здатний “вимикати” експресію білків необхідних для асоціації ендогенних пептидів
- 25. Регуляція МНС класу 1 Молекулярний механізм Ген UL49.5 кодує білок який здатний: блокувати ТАР білки, які
- 26. Основний онкоген meq (Marek’s disease EcoRI-Q) Описаний Kung та співавт. у 1992 році Поліфункціональний ДНК-з’вязуючий білок,
- 27. Основний онкоген meq. Структура 339 амінокислот, 44 кДа. Пролін- багата ділянка відповідає за трансактивацію (33 а-к-ти
- 28. Основний онкоген meq. Має здатність до асоціації із клітинними прото-онкогенами Fos та Jun cAMP-response-element-binding protein (CREB)
- 29. Докази участі Мeq у неопластичному перетворенні: постійно експресується у трансформованих CD41+ Т-клітинах, рецесивні мутантні віруси за
- 30. Загальна картина активності онкогену Взаємодія Meq з Jun веде до активанії АР-1 та 2 енхансерів, підконтрольних
- 31. Віруси герпеса приматів Із дегенеруючої первинної культури нирок білкової мавпи був виділений вірус герпеса HVSaimiri, який
- 32. Герпесоподібний вірус (HVAteles), виділений від здорових павукоподібних мавп, також виявився онкогенним для деяких видів мавп Нового
- 33. Human Herpesvirus 1 (Herpes simplex virus 1, HSV-1) and HHV2/HSV-2 HSV-1 первинна оральна інфекція. HSV-2 первинна
- 34. Структура HSV
- 35. Генетичні карти герпесвірусів
- 36. HSV-1 та HSV-2 (1) ~80 генів; половина з них необхідна для реплікації. Спайсингу піддається тільки ген
- 37. HSV-1та HSV-2 (2) Latency associated transcripts (LATs) –сплайсовані транскрипти, у яких відсутні polyA послідовності , присутні
- 38. HSV-1та HSV-2 1. HSV1 та HSV2 мають в геномі трансформуючі гені (mtr), які здатні іморталізувати компетентні
- 39. гіпотеза непрямої коканцерогенної дії HSV за механізмом “hit and run” (вдарив та втік) на основі таких
- 40. Cytomegalovirus (HHV-5) Широкий спектр клінічної патології – пантропність реплікація в клітинах життєво важливих органів людини. Шляхи
- 41. Геном ННV–5 персистує в клітинах крові (проблема трансплантології). До 2% лейкоцитів донорів містять передранні білки ННV–5.
- 42. HHV-6 Вірус герпесу людини 6–го типу Виділили з В–лімфоцитів периферичної крові хворих на лімфопроліферативними (Т–клітинна лімфома,
- 43. ННV–6 Тривала персистенція в організмі після первинного інфікування Персистенція : латентний стан (відсутність продукції інфекційного вірусу)
- 44. ННV–6А ННV–6В ННV–6А виклакає лімфогранулематоз, лімфоретикульоз, гемоцитобластоз, разсіяний склероз ННV–6В – раптову екзантему новонароджених (exanthema subitum
- 46. Скачать презентацию

нейротропні І ІІ ІІІ типи
HERPESVIRUSES
лімфотропні ІУ УІ УІІ УІІІ
нейротропні І ІІ ІІІ типи
HERPESVIRUSES
лімфотропні ІУ УІ УІІ УІІІ

Дисемінація герпесвірусів в організмі людини:
від клітини до клітини,
гематогенно,
Дисемінація герпесвірусів в організмі людини:
від клітини до клітини,
гематогенно,

Поширення герпесвірусів
Більшість населення планети - у стані «здорового» носійства
Віруси «ховаються» від
Поширення герпесвірусів
Більшість населення планети - у стані «здорового» носійства
Віруси «ховаються» від
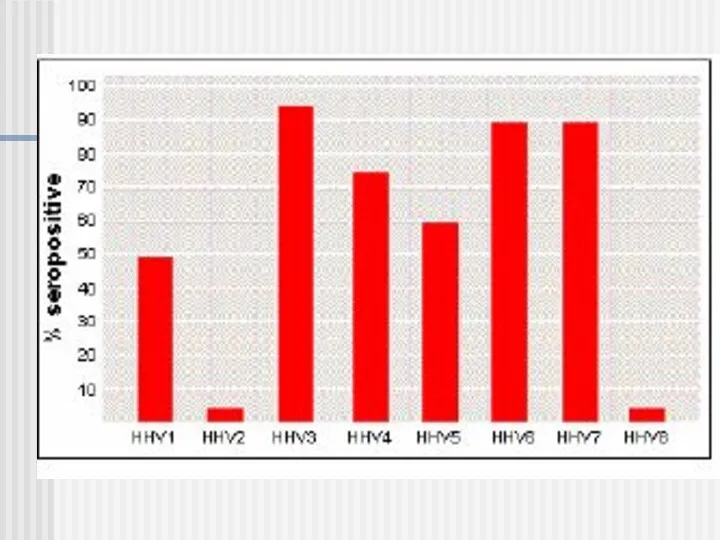

Віруси впливають на життєво важливі функції макроорганізму імунобіологічні системи його захисту

Онкопотенціал герпесвірусів:
• Лімфома Беркітта (HHV–4).
• Назофарингеальна карцинома (HHV–4).
• Лімфоми, лімфосаркоми (HHV–4).
•
Онкопотенціал герпесвірусів:
• Лімфома Беркітта (HHV–4). • Назофарингеальна карцинома (HHV–4). • Лімфоми, лімфосаркоми (HHV–4). •

Вірус Люке
В природніх умовах вірус викликає аденокарциноми нирок леопардових жаб, які
Вірус Люке
В природніх умовах вірус викликає аденокарциноми нирок леопардових жаб, які

Хвороба Марека
Хвороба Марека – висококонтагіозне лімфопроліферативне захворювання курей
Характерні ознаки – розвиток
Хвороба Марека
Хвороба Марека – висококонтагіозне лімфопроліферативне захворювання курей
Характерні ознаки – розвиток
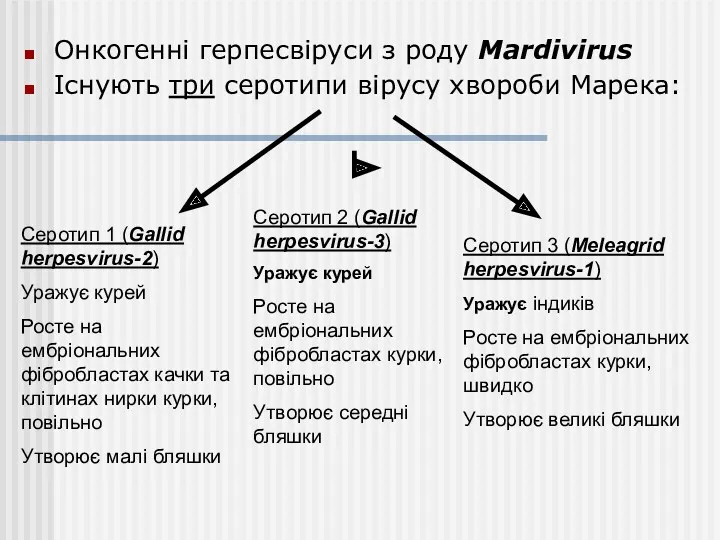
Онкогенні герпесвіруси з роду Mardivirus
Існують три серотипи вірусу хвороби Марека:
Існують три серотипи вірусу хвороби Марека:
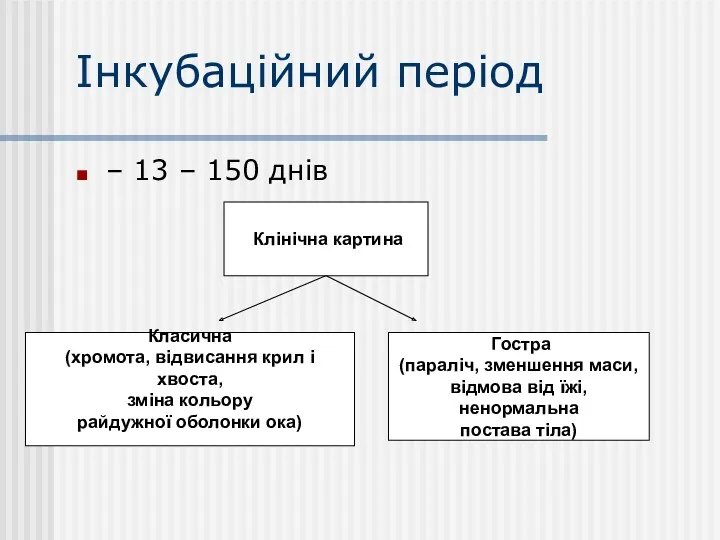
Інкубаційний період
– 13 – 150 днів
Клінічна картина
Класична
(хромота, відвисання крил і
Інкубаційний період
– 13 – 150 днів
Клінічна картина
Класична
(хромота, відвисання крил і
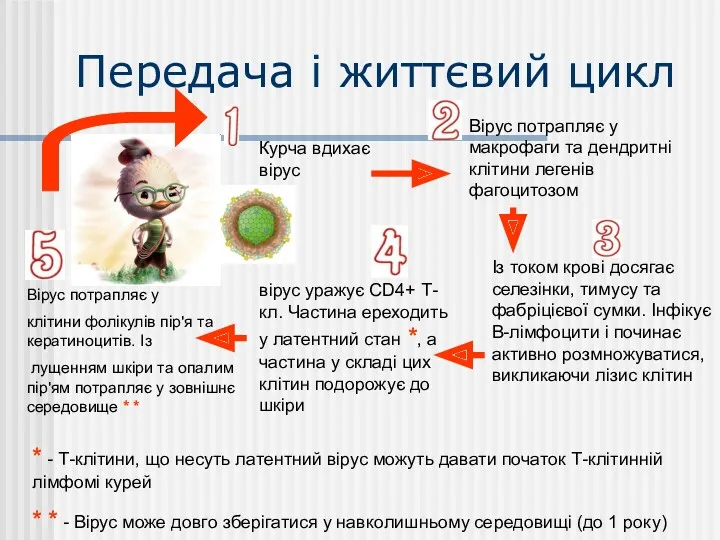
Передача і життєвий цикл
Курча вдихає вірус
Вірус потрапляє у макрофаги та дендритні
Передача і життєвий цикл
Курча вдихає вірус
Вірус потрапляє у макрофаги та дендритні

Патогенез
Виділяють чотири фази інфекції
гостра первинна інфекція та дегенеративні зміни ранньої
Патогенез
Виділяють чотири фази інфекції
гостра первинна інфекція та дегенеративні зміни ранньої

Патанатомія
Атрофія м’язів
Дегенеративні ушкодження периферичних нервів
Лімфоматозні утворення сало-подібної консистенції у гонадах, печінці,
Патанатомія
Атрофія м’язів
Дегенеративні ушкодження периферичних нервів
Лімфоматозні утворення сало-подібної консистенції у гонадах, печінці,
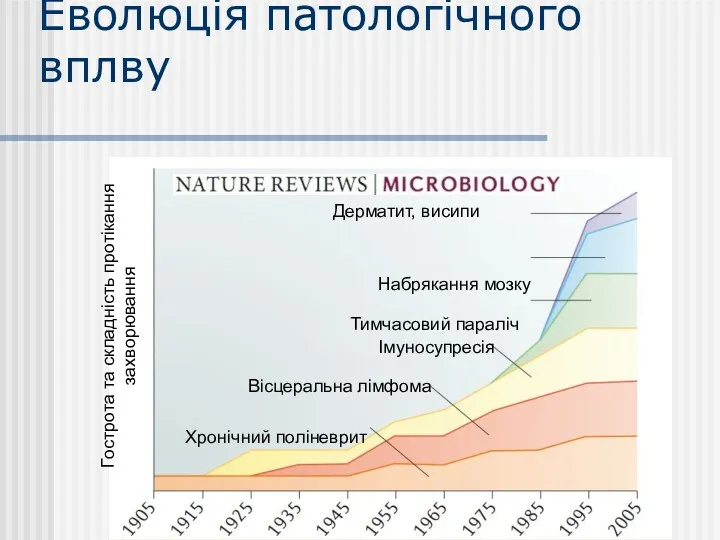
Еволюція патологічного вплву
Гострота та складність протікання захворювання
Хронічний поліневрит
Вісцеральна лімфома
Імуносупресія
Тимчасовий параліч
Набрякання мозку
Дерматит,
Еволюція патологічного вплву
Гострота та складність протікання захворювання
Хронічний поліневрит
Вісцеральна лімфома
Імуносупресія
Тимчасовий параліч
Набрякання мозку
Дерматит,

Загальні відомості про трансформацію
Вірус здатний до трансформації СD4+ Т-клітини, в яких
Загальні відомості про трансформацію
Вірус здатний до трансформації СD4+ Т-клітини, в яких
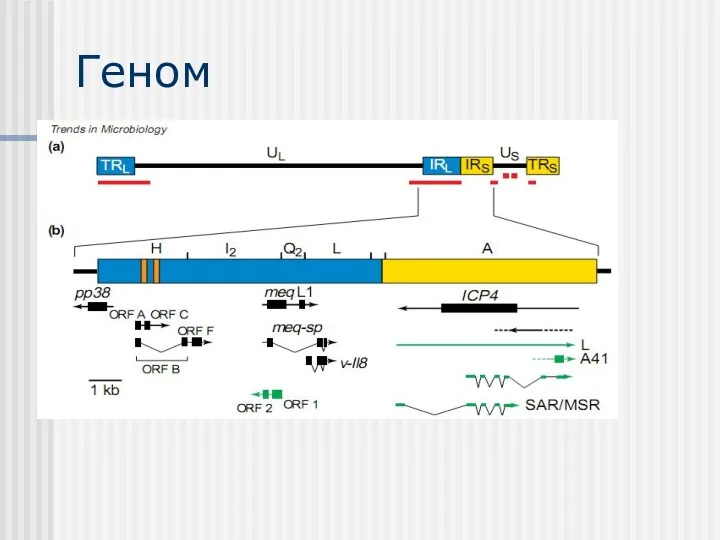
Геном
Геном

Порівняння геномів вірусу varicella–zoster virus (VZV) та вірусу хвороби Марека (MDV)
Порівняння геномів вірусу varicella–zoster virus (VZV) та вірусу хвороби Марека (MDV)

Регуляція МНС класу 1 та ІНФ-α
Вірус здатний “вимикати” експресію білків необхідних
Регуляція МНС класу 1 та ІНФ-α
Вірус здатний “вимикати” експресію білків необхідних
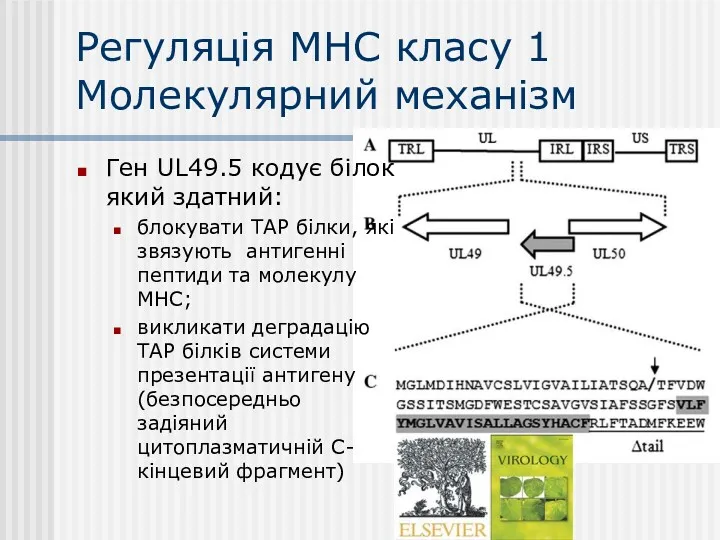
Регуляція МНС класу 1
Молекулярний механізм
Ген UL49.5 кодує білок який здатний:
блокувати ТАР
Регуляція МНС класу 1
Молекулярний механізм
Ген UL49.5 кодує білок який здатний:
блокувати ТАР

Латентний стан та трансформація
Розрізнити Т-клітини, які несуть вірус у латентному
Латентний стан та трансформація
Розрізнити Т-клітини, які несуть вірус у латентному

Загальні відомості про трансформацію
Вірус здатний до трансформації СD4+ Т-клітини, в яких
Загальні відомості про трансформацію
Вірус здатний до трансформації СD4+ Т-клітини, в яких

Деякі вірусні протеїни, що експресуються у трансформованих Т-лімфоцитах
pp38 – 24; 38
Деякі вірусні протеїни, що експресуються у трансформованих Т-лімфоцитах
pp38 – 24; 38

Регуляція МНС класу 1 та ІНФ-α
Вірус здатний “вимикати” експресію білків необхідних
Регуляція МНС класу 1 та ІНФ-α
Вірус здатний “вимикати” експресію білків необхідних
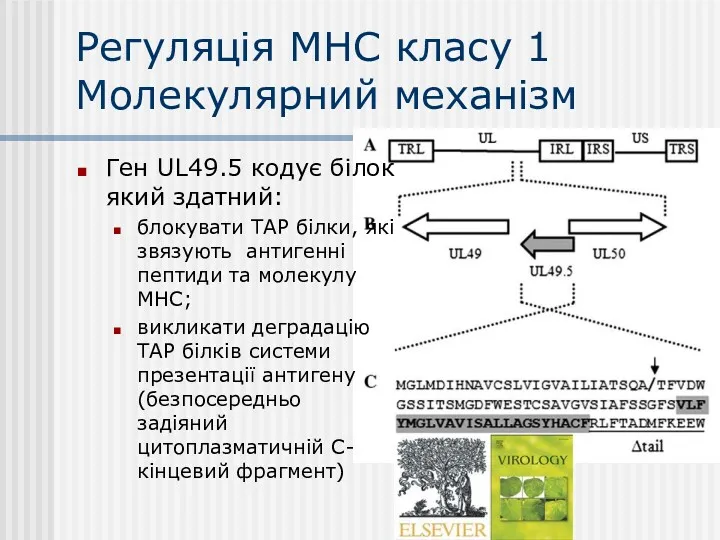
Регуляція МНС класу 1
Молекулярний механізм
Ген UL49.5 кодує білок який здатний:
блокувати ТАР
Регуляція МНС класу 1
Молекулярний механізм
Ген UL49.5 кодує білок який здатний:
блокувати ТАР

Основний онкоген meq
(Marek’s disease EcoRI-Q)
Описаний Kung та співавт. у 1992
Основний онкоген meq
(Marek’s disease EcoRI-Q)
Описаний Kung та співавт. у 1992

Основний онкоген meq. Структура
339 амінокислот, 44 кДа. Пролін- багата ділянка відповідає
Основний онкоген meq. Структура
339 амінокислот, 44 кДа. Пролін- багата ділянка відповідає

Основний онкоген meq.
Має здатність до асоціації із клітинними прото-онкогенами
Fos
Основний онкоген meq.
Має здатність до асоціації із клітинними прото-онкогенами
Fos

Докази участі Мeq у неопластичному перетворенні:
постійно експресується у трансформованих CD41+ Т-клітинах,
Докази участі Мeq у неопластичному перетворенні:
постійно експресується у трансформованих CD41+ Т-клітинах,
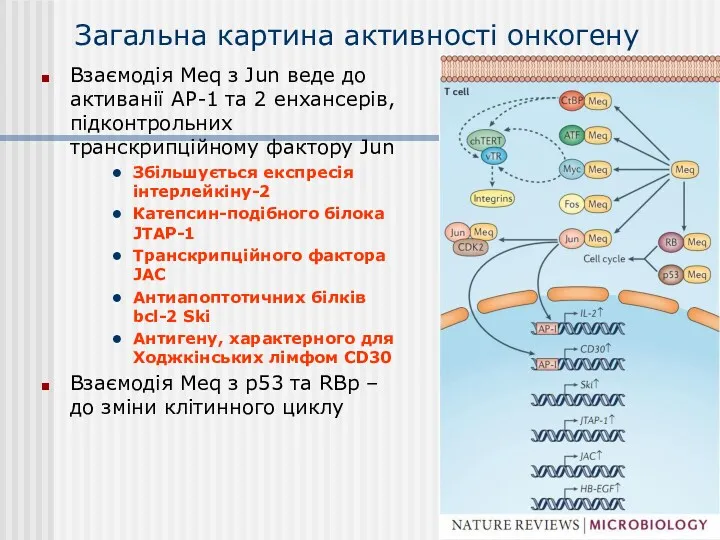
Загальна картина активності онкогену
Взаємодія Meq з Jun веде до активанії АР-1
Загальна картина активності онкогену
Взаємодія Meq з Jun веде до активанії АР-1

Віруси герпеса приматів
Із дегенеруючої первинної культури нирок білкової мавпи був виділений
Віруси герпеса приматів
Із дегенеруючої первинної культури нирок білкової мавпи був виділений

Герпесоподібний вірус (HVAteles), виділений від здорових павукоподібних мавп, також виявився онкогенним
Герпесоподібний вірус (HVAteles), виділений від здорових павукоподібних мавп, також виявився онкогенним

Human Herpesvirus 1 (Herpes simplex virus 1, HSV-1) and HHV2/HSV-2
HSV-1 первинна
Human Herpesvirus 1 (Herpes simplex virus 1, HSV-1) and HHV2/HSV-2
HSV-1 первинна
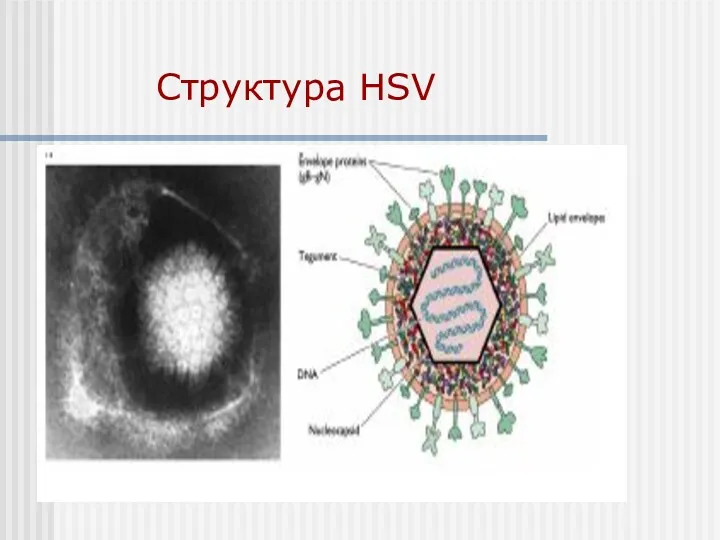
Структура HSV
Структура HSV
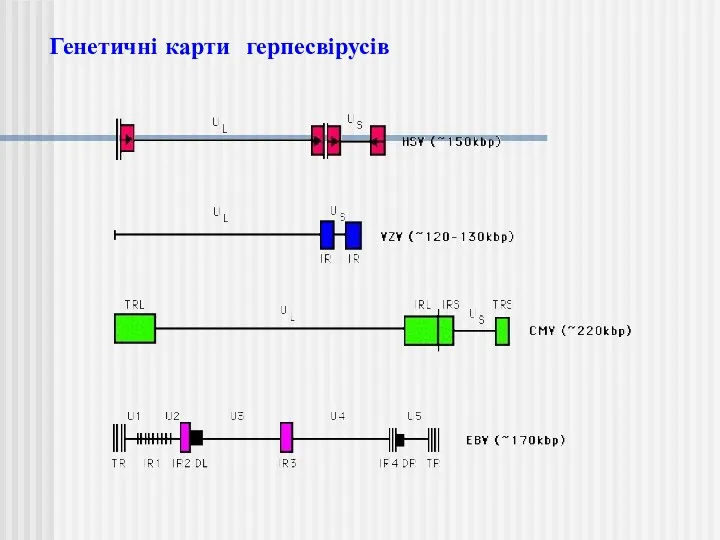
Генетичні карти герпесвірусів
Генетичні карти герпесвірусів

HSV-1 та HSV-2
(1)
~80 генів; половина з них необхідна
HSV-1 та HSV-2
(1)
~80 генів; половина з них необхідна

HSV-1та HSV-2 (2)
Latency associated transcripts (LATs) –сплайсовані транскрипти, у яких
HSV-1та HSV-2 (2)
Latency associated transcripts (LATs) –сплайсовані транскрипти, у яких

HSV-1та HSV-2
1. HSV1 та HSV2 мають в геномі трансформуючі гені (mtr),
HSV-1та HSV-2
1. HSV1 та HSV2 мають в геномі трансформуючі гені (mtr),

гіпотеза
непрямої коканцерогенної дії HSV за механізмом “hit and run” (вдарив та
гіпотеза
непрямої коканцерогенної дії HSV за механізмом “hit and run” (вдарив та

Cytomegalovirus (HHV-5)
Широкий спектр клінічної патології – пантропність реплікація в клітинах
Cytomegalovirus (HHV-5)
Широкий спектр клінічної патології – пантропність реплікація в клітинах

Геном ННV–5 персистує в клітинах крові (проблема трансплантології).
До 2% лейкоцитів
Геном ННV–5 персистує в клітинах крові (проблема трансплантології).
До 2% лейкоцитів

HHV-6
Вірус герпесу людини 6–го типу
Виділили з В–лімфоцитів периферичної крові хворих
HHV-6
Вірус герпесу людини 6–го типу
Виділили з В–лімфоцитів периферичної крові хворих

ННV–6
Тривала персистенція в організмі після первинного інфікування
Персистенція : латентний стан
ННV–6
Тривала персистенція в організмі після первинного інфікування
Персистенція : латентний стан

ННV–6А ННV–6В
ННV–6А виклакає лімфогранулематоз, лімфоретикульоз, гемоцитобластоз, разсіяний склероз
ННV–6В – раптову
ННV–6А ННV–6В
ННV–6А виклакає лімфогранулематоз, лімфоретикульоз, гемоцитобластоз, разсіяний склероз
ННV–6В – раптову
 Синтетические противомикробные, противогрибковые и противовирусные средства
Синтетические противомикробные, противогрибковые и противовирусные средства Pathomorphology of systemic and local violation of blood circulation
Pathomorphology of systemic and local violation of blood circulation Врожденные пороки развития
Врожденные пороки развития Клинический случай пациента
Клинический случай пациента Натуральная оспа
Натуральная оспа Внутриматочные спирали
Внутриматочные спирали Творчество в шизофрении
Творчество в шизофрении Teri gigiyenasi: teri shikastlanganda 1-yordam berish
Teri gigiyenasi: teri shikastlanganda 1-yordam berish Пищевые токсикоинфекции. Ботулизм
Пищевые токсикоинфекции. Ботулизм Тактика ведения больных в постинсультном периоде
Тактика ведения больных в постинсультном периоде Методы анализа лекарственных средств. Поляриметрия
Методы анализа лекарственных средств. Поляриметрия Стационар мен емханалардың жұмысын ұйымдастыру
Стационар мен емханалардың жұмысын ұйымдастыру Коклюш. Паракоклюш
Коклюш. Паракоклюш Қанның қан тамырларымен қозғалысының гемодинамикалық заңдылықтары. (Дәріс 11)
Қанның қан тамырларымен қозғалысының гемодинамикалық заңдылықтары. (Дәріс 11) Дезагрегантная терапия острого коронарного синдрома на реанимационном этапе
Дезагрегантная терапия острого коронарного синдрома на реанимационном этапе Косметические средства ухода за кожей детей при аномалиях конституции
Косметические средства ухода за кожей детей при аномалиях конституции German Medical Association
German Medical Association Общий уход за больными в предоперационный и послеоперационный период
Общий уход за больными в предоперационный и послеоперационный период Глаукома
Глаукома Лекарственная болезнь
Лекарственная болезнь Дисфункциональные маточные кровотечения. Ювенильные маточные кровотечения
Дисфункциональные маточные кровотечения. Ювенильные маточные кровотечения Arenaviruses: unique virology. Diseases of the Old World and New World
Arenaviruses: unique virology. Diseases of the Old World and New World Витамин Д3
Витамин Д3 Питание в футболе
Питание в футболе Магнитные наночастицы в современной медицине
Магнитные наночастицы в современной медицине Киборг. Искусственный прототип человека
Киборг. Искусственный прототип человека Введение в сексологию
Введение в сексологию Онкологиялық пациенттерге үй жағдайында мейірбикелік күтім көрсету
Онкологиялық пациенттерге үй жағдайында мейірбикелік күтім көрсету