- Иммунитет. Врожденный и приобретенный иммунитет

Содержание
- 2. В механизмах устойчивости организма к чужеродной информации участвуют два основных феномена: неспецифическая резистентность (врожденный, естественный иммунитет)
- 3. Неспецифическая резистентность или неспецифический иммунитет нормальная микробиологическая оболочка тела (сапрофиты кожи и слизистых) печень (система цитохрома
- 4. Специфические механизмы или приобретенный иммунитет (иммунный ответ) гуморальный или B- опосредованный иммунный ответ образование АТ: IgM,
- 5. антитоксический против экзотоксинов, ферментов - токсинов, эндотоксинов. В основе лежит гуморальный иммунный ответ антибактериальный АТ образуются
- 6. ОРГАНЫ ИММУННОЙ СИСТЕМЫ ЦЕНТРАЛЬНЫЕ ПЕРИФЕРИЧЕСКИЕ Красный костный мозг ТИМУС центральный орган, в котором осуществляется продукция ИКК
- 7. Основные функции иммунной системы: защита целостности организма от проникновения чужеродных веществ путем инактивации, разрушения и элиминации
- 8. Иммунная система — сложнейшая биологическая система. Исполнительными органами ее являются лимфоциты, общее число которых достигает 1012
- 9. Эффекторные клетки Ig Влф CD72, CD79 a/b ПК CD85 Tk CD8+ А Н CD88, CD89 ТК
- 10. Клетки памяти Тлф CD45 Влф Tk CD8+ ПК
- 11. Лф сами по себе не способны осуществлять эффективный ИО. Им помогают: АКЦЕССОРНЫЕ КЛЕТКИ Акцессорные клетки действуют
- 12. В отличие от МФ не полностью разрушают АГ, а перерабатывают его до пептидов. ДК способны в
- 13. Выделяют две ключевые функции ДК: первые маркеры АГ зрелые клетки (ч/з день или позже) стимулируют Т-лф
- 14. Строение белков МНС I и II классов S S S S S S S S α1
- 15. В тимусе происходят процессы дифференцировки Т-лф на субпопуляции (Tk, Th). HLA принадлежит центральное место в дифференцировке
- 16. 60% 30% ИФγ, ФНОα, ИЛ-2 ИЛ-4, 5, 6, 10, ТФРβ ИЛ-2, ИФγ, ФНОα Тлф CD3+ Tk
- 17. АГ ИФγ, ФНОα, ИЛ-2 ФНО, ИЛ-1, 6 и 8, MIF ИЛ-2, 4, 5, 6, 10, ТФРβ
- 18. ИММУНОЦИТОКИНЫ обусловливают развитие базовых реакций МФ и АГспецифических клеток: взаимодействие этих клеток с Лф, активацию и
- 19. Провоспалительные цитокины ИЛ-1 способен стимулировать Т- и В-лф, усиливать клеточную пролиферацию, инициировать или супрессировать экспрессию определенных
- 20. ФНОα участвует в регуляции ИО и воспаления: активирует эндотелий, способствует адгезии лейкоцитов к эндотелию за счет
- 21. Цитокины, регулирующие ГИО и КИО ИЛ-2 активирует все клоны цитотоксических клеток: Tk и НК, увеличивает продукцию
- 22. Особенности функционирования ИС Специфичность. Устойчивость к отдельным заболеваниям носит специфический характер. В основе способность ИС реагировать
- 23. Иммунный ответ ПЕРВЫЙ ЭТАП ИО комплексный процесс, переработки и представления АГ в иммуногенной форме на поверхности
- 24. С генетической точки зрения иммунологическая реакция это детерминированный процесс, в котором одновременно принимают участие различные клетки
- 25. Одновременно с распознаванием этого АГсодержащего комплекса происходит контактное взаимодействие АПК и Th с помощью ряда взаимно
- 26. Простой ответ возникает на Т-независимые АГ. Некоторые АГ способны сами активировать Т-лф. Тогда синтез IgM не
- 27. Первичный ответ как процесс накопления АТ характеризуется тремя этапами: фаза снижения - затухания ответа вплоть до
- 28. титры Ig 2 4 6 8 10 12 14 16 18 20 время после контакта с
- 29. 5 классов Ig IgA, IgG, IgM, IgD, IgE Молекула Ig состоит из тяжелых цепей одного типа,
- 30. Строение молекулы АТ (по Р.В.Петрову) --S—S-- S--------S S--------S S-----S S-----S S--------S S--------S S--------S S--------S S--------S S--------S
- 31. Легкая цепь Fab Fc Тяжелая цепь
- 32. Наиболее важные и самыми распространенными Ig сыворотки мономерного класса (80%). Это класс АТ, обладающий противовирусной и
- 33. IgА Секреторные sIgA находятся на слизистых кишечника, верхних дыхательных и мочевыводящих путей, содержатся в слезах, слюне,
- 34. IgЕ это реагины или гомоцитотропные АТ. Связываются своим Fc-фрагментом с высокоаффинными рецепторами ТК и Б IgE
- 35. IgD IgD локализуются в мембране В-лф, ПК и выполняют функции иммунноглобулиновых антигенраспознающих рецепторов. В процессе дифференцировки
- 36. IgМ IgМ – первичные АТ - выделяются на ранних стадиях развития плода и первичном ИО. У
- 37. определяются Fab и Fc-фрагментами. С Fab-фрагментом связано понятие авидности АТ. С этим свойством связано такое качество,
- 38. - иммобилизация бактерий осуществляется АТ против бактериальных ворсинок, нарушающими их подвижность - активация комплемента особенно Fc-регионом
- 39. Клеточный иммунный ответ развивается на чужеродные клетки, на клетки, пораженные вирусом, на внутриклеточные организмы Эффекторы -
- 40. МНСII АГ Тh2 B B Ig ИЛ-1, 6, 12; ФНОα ИЛ-4, 7, 13 ИЛ-5, 6, 10
- 41. Селекция тимоцитов Экспрессия ТКР на индивидуальном тимоците означает, что клетка вступает в этап позитивной селекции Позитивная
- 42. Узнавание "своего" осуществляется не всем АГраспознающим центром ТКР. Оставшаяся часть центра будет взаимодействовать встрече с чужеродными
- 43. Специальных клеток для подавления ИО, не существует. Продуцируя медиаторы с противоположным характером действия Th1 подавляют функции
- 44. Результатом ИО является формирование клеток памяти, которые появляются в результате ИО в периферических лимфоидных органах. Клетки
- 45. Эффекторные механизмы иммунитета Неспецифические эффекторные механизмы иммунитета Воспаление Фагоцитоз Н МФ ИЛ-1, ФНО, ИФγ и ИЛ-8
- 46. ИММУНОЛОГИЧЕСКАЯ ТОЛЕРАНТНОСТЬ Лф должны распознавать собственные АГ тканевой совместимости, специфичные для каждого индивидуума. Одновременно они не
- 47. Иммунологическая толерантность характеризуется: - отсутствием ответа на собственный АГ - отсутствием элиминации АГ при повторном его
- 48. В результате позитивной и негативной селекции из тимуса в кровоток и лимфоидные органы поступают только такие
- 49. РЕГУЛЯЦИЯ ИММУНОГО ОТВЕТА Эффективный ИО - результат взаимодействия между АГ и целой сетью ИКК. Кроме того,
- 50. Характер ИО зависит от присутствия цитокинов в микроокружении Т-лф в момент распознавания АГ и активации. Если
- 51. Опиоидные пептиды - связующее звено между нервной и иммунной системой. ЭНД и ЭНК стимулируют активность Т-клеток
- 52. ЩИТОВИДНАЯ ЖЕЛЕЗА Тироксин стимулируют фагоцитарную активность лейкоцитов, активирует цитотоксические функции Лф периферической крови человека НАДПОЧЕЧНИКИ Глюкокортикоиды
- 53. ДЕТСКИЕ ОСОБЕННОСТИ Развивается блок ИС матери плода: развивается толерантность к отцовским АГ трансплацентарный перенос АТ от
- 55. Скачать презентацию

В механизмах устойчивости организма к чужеродной информации участвуют два основных феномена:
неспецифическая
В механизмах устойчивости организма к чужеродной информации участвуют два основных феномена:
неспецифическая
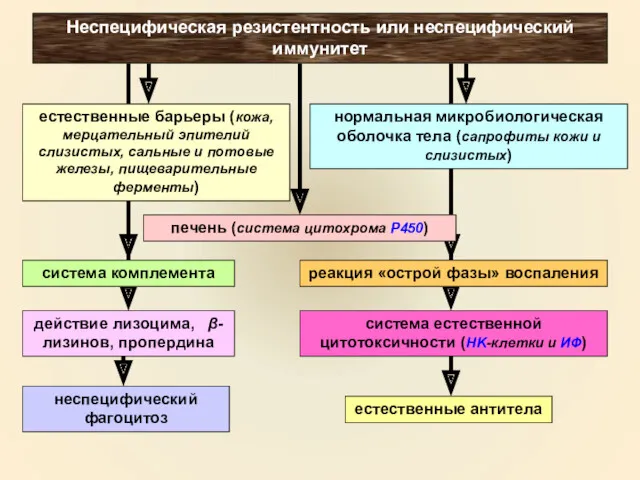
Неспецифическая резистентность или неспецифический иммунитет
нормальная микробиологическая оболочка тела (сапрофиты кожи
Неспецифическая резистентность или неспецифический иммунитет
нормальная микробиологическая оболочка тела (сапрофиты кожи
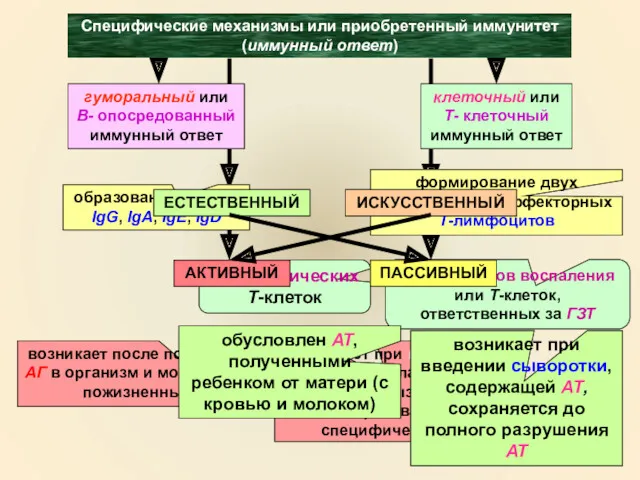
Специфические механизмы или приобретенный иммунитет (иммунный ответ)
гуморальный или B- опосредованный
Специфические механизмы или приобретенный иммунитет (иммунный ответ)
гуморальный или B- опосредованный
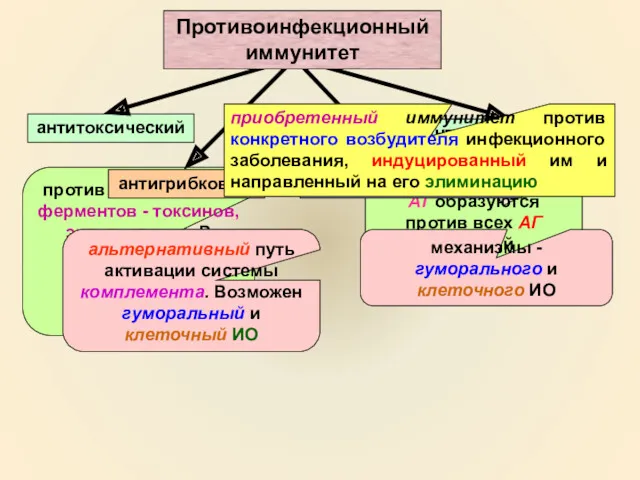
антитоксический
против экзотоксинов, ферментов - токсинов, эндотоксинов. В основе лежит гуморальный
антитоксический
против экзотоксинов, ферментов - токсинов, эндотоксинов. В основе лежит гуморальный
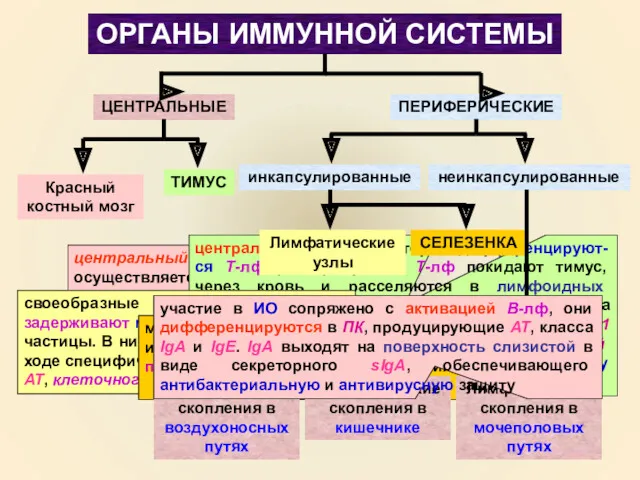
ОРГАНЫ ИММУННОЙ СИСТЕМЫ
ЦЕНТРАЛЬНЫЕ
ПЕРИФЕРИЧЕСКИЕ
Красный костный мозг
ТИМУС
центральный орган, в котором осуществляется продукция ИКК
ОРГАНЫ ИММУННОЙ СИСТЕМЫ
ЦЕНТРАЛЬНЫЕ
ПЕРИФЕРИЧЕСКИЕ
Красный костный мозг
ТИМУС
центральный орган, в котором осуществляется продукция ИКК

Основные функции иммунной системы:
защита целостности организма от проникновения чужеродных веществ путем
Основные функции иммунной системы:
защита целостности организма от проникновения чужеродных веществ путем
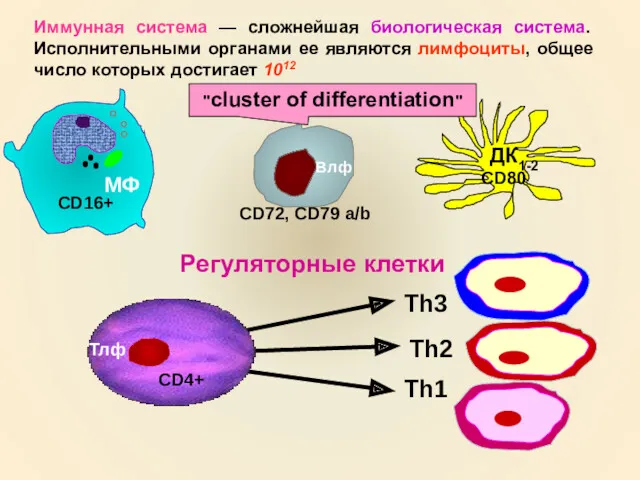
Иммунная система — сложнейшая биологическая система. Исполнительными органами ее являются лимфоциты,
Иммунная система — сложнейшая биологическая система. Исполнительными органами ее являются лимфоциты,

Эффекторные клетки
Ig
Влф
CD72, CD79 a/b
ПК
CD85
Tk
CD8+
А
Н
CD88, CD89
ТК
CD88
ФБ
CD121 a/b
Б
CD128
Э
CD88
ЭЦ
CD121 a/b
NK
CD56/CD57
Эффекторные клетки
Ig
Влф
CD72, CD79 a/b
ПК
CD85
Tk
CD8+
А
Н
CD88, CD89
ТК
CD88
ФБ
CD121 a/b
Б
CD128
Э
CD88
ЭЦ
CD121 a/b
NK
CD56/CD57
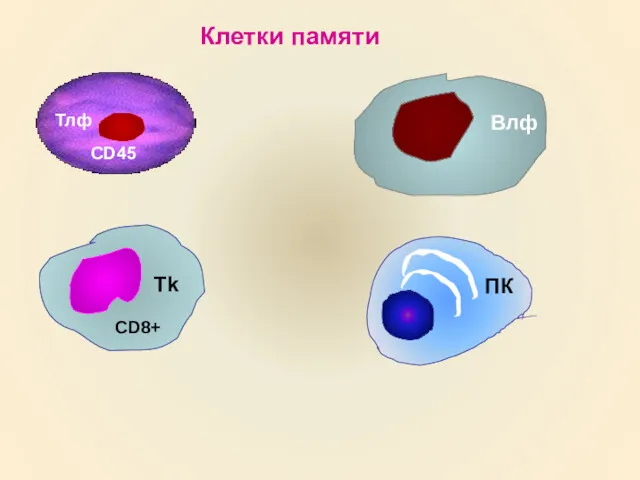
Клетки памяти
Тлф
CD45
Влф
Tk
CD8+
ПК
Клетки памяти
Тлф
CD45
Влф
Tk
CD8+
ПК

Лф сами по себе не способны осуществлять эффективный ИО. Им помогают:
АКЦЕССОРНЫЕ
Лф сами по себе не способны осуществлять эффективный ИО. Им помогают:
АКЦЕССОРНЫЕ
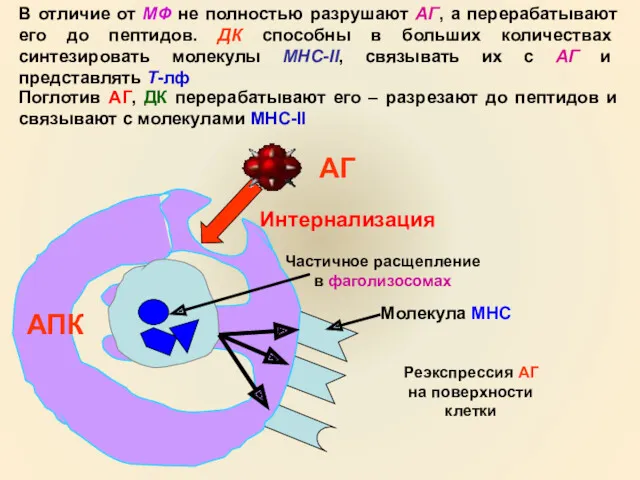
В отличие от МФ не полностью разрушают АГ, а перерабатывают его
В отличие от МФ не полностью разрушают АГ, а перерабатывают его

Выделяют две ключевые функции ДК:
первые маркеры АГ
зрелые клетки (ч/з день или
Выделяют две ключевые функции ДК:
первые маркеры АГ
зрелые клетки (ч/з день или
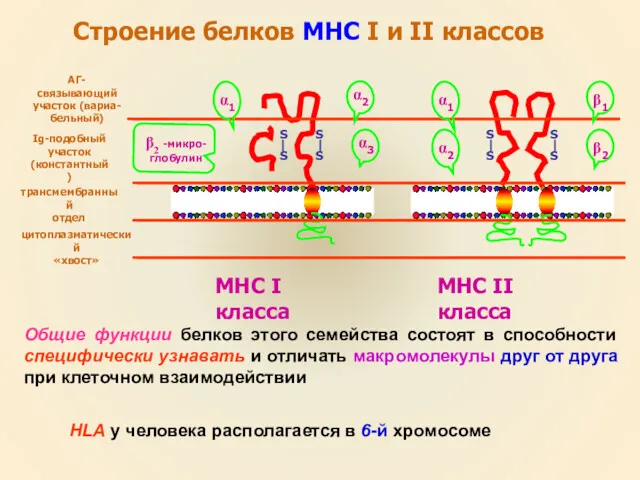
Строение белков МНС I и II классов
S
S
S
S
S
S
S
S
α1
α1
α2
α3
α2
β1
β2
β2
Строение белков МНС I и II классов
S
S
S
S
S
S
S
S
α1
α1
α2
α3
α2
β1
β2
β2

В тимусе происходят процессы дифференцировки Т-лф на субпопуляции (Tk, Th). HLA
В тимусе происходят процессы дифференцировки Т-лф на субпопуляции (Tk, Th). HLA
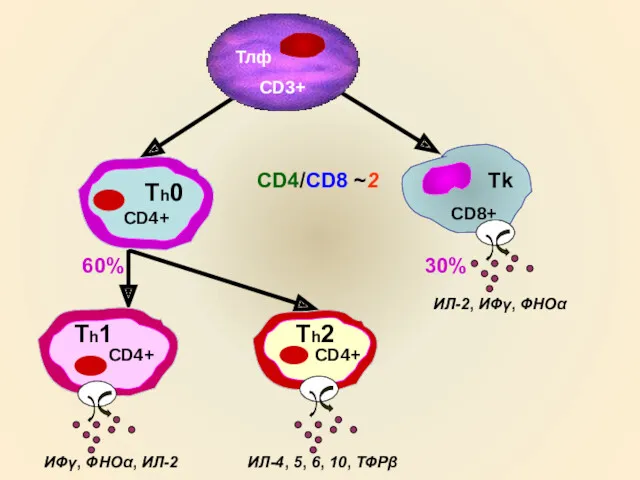
60%
30%
ИФγ, ФНОα, ИЛ-2
ИЛ-4, 5, 6, 10, ТФРβ
ИЛ-2, ИФγ, ФНОα
60%
30%
ИФγ, ФНОα, ИЛ-2
ИЛ-4, 5, 6, 10, ТФРβ
ИЛ-2, ИФγ, ФНОα
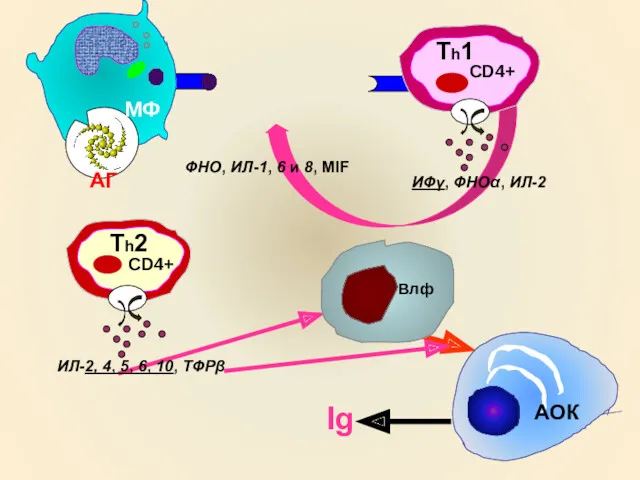
АГ
ИФγ, ФНОα, ИЛ-2
ФНО, ИЛ-1, 6 и 8, MIF
ИЛ-2, 4, 5,
АГ
ИФγ, ФНОα, ИЛ-2
ФНО, ИЛ-1, 6 и 8, MIF
ИЛ-2, 4, 5,

ИММУНОЦИТОКИНЫ
обусловливают развитие базовых реакций МФ и АГспецифических клеток: взаимодействие этих клеток
ИММУНОЦИТОКИНЫ
обусловливают развитие базовых реакций МФ и АГспецифических клеток: взаимодействие этих клеток

Провоспалительные цитокины
ИЛ-1
способен стимулировать Т- и В-лф, усиливать клеточную пролиферацию, инициировать
Провоспалительные цитокины
ИЛ-1
способен стимулировать Т- и В-лф, усиливать клеточную пролиферацию, инициировать

ФНОα
участвует в регуляции ИО и воспаления: активирует эндотелий, способствует адгезии
ФНОα
участвует в регуляции ИО и воспаления: активирует эндотелий, способствует адгезии

Цитокины, регулирующие ГИО и КИО
ИЛ-2
активирует все клоны цитотоксических клеток: Tk
Цитокины, регулирующие ГИО и КИО
ИЛ-2
активирует все клоны цитотоксических клеток: Tk

Особенности функционирования ИС
Специфичность. Устойчивость к отдельным заболеваниям носит специфический характер. В
Особенности функционирования ИС
Специфичность. Устойчивость к отдельным заболеваниям носит специфический характер. В

Иммунный ответ
ПЕРВЫЙ ЭТАП ИО
комплексный процесс, переработки и представления АГ в иммуногенной
Иммунный ответ
ПЕРВЫЙ ЭТАП ИО
комплексный процесс, переработки и представления АГ в иммуногенной

С генетической точки зрения иммунологическая реакция это детерминированный процесс, в котором
С генетической точки зрения иммунологическая реакция это детерминированный процесс, в котором

Одновременно с распознаванием этого АГсодержащего комплекса происходит контактное взаимодействие АПК и
Одновременно с распознаванием этого АГсодержащего комплекса происходит контактное взаимодействие АПК и

Простой ответ возникает на Т-независимые АГ. Некоторые АГ способны сами активировать
Простой ответ возникает на Т-независимые АГ. Некоторые АГ способны сами активировать
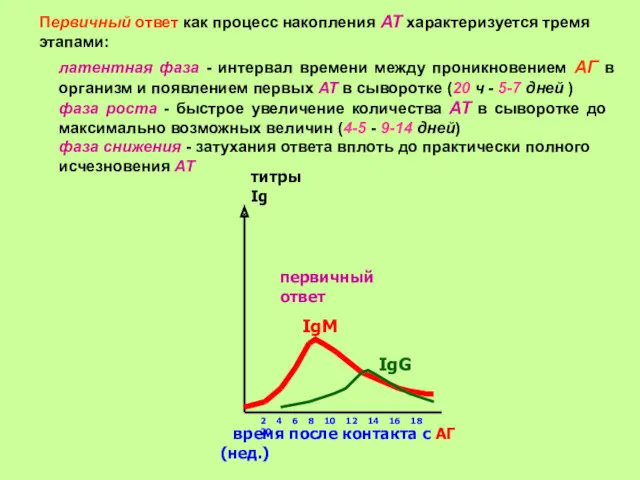
Первичный ответ как процесс накопления АТ характеризуется тремя этапами:
фаза снижения -
Первичный ответ как процесс накопления АТ характеризуется тремя этапами:
фаза снижения -
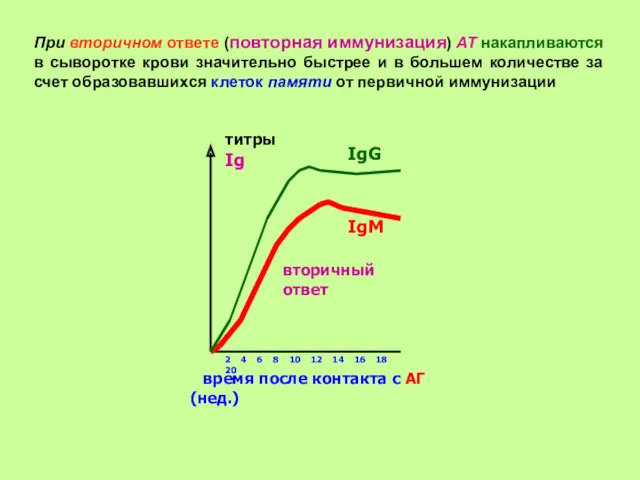
титры Ig
2 4 6 8 10 12 14 16 18 20
титры Ig
2 4 6 8 10 12 14 16 18 20

5 классов Ig
IgA, IgG, IgM, IgD, IgE
Молекула Ig состоит из тяжелых
5 классов Ig
IgA, IgG, IgM, IgD, IgE
Молекула Ig состоит из тяжелых
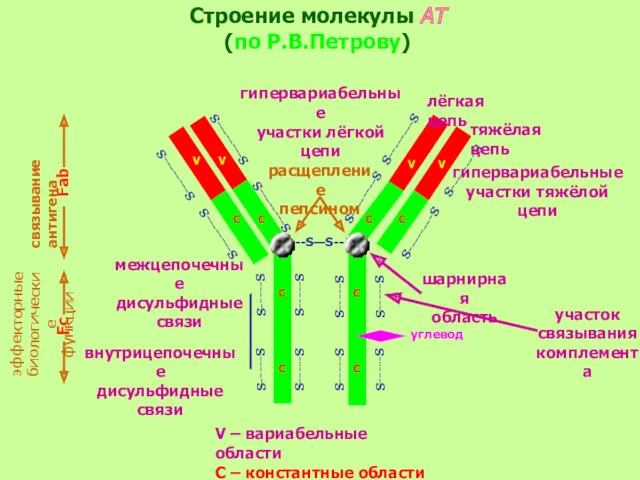
Строение молекулы АТ
(по Р.В.Петрову)
--S—S--
S--------S
S--------S
S-----S
S-----S
S--------S
S--------S
S--------S
S--------S
S--------S
S--------S
S-----S
S-----S
S-----S
S-----S
S-----S
S-----S
углевод
гипервариабельные
участки лёгкой цепи
шарнирная
область
межцепочечные
дисульфидные
связи
внутрицепочечные
дисульфидные
связи
участок
связывания
комплемента
лёгкая цепь
тяжёлая цепь
расщепление
пепсином
гипервариабельные
участки тяжёлой
Строение молекулы АТ
(по Р.В.Петрову)
--S—S--
S--------S
S--------S
S-----S
S-----S
S--------S
S--------S
S--------S
S--------S
S--------S
S--------S
S-----S
S-----S
S-----S
S-----S
S-----S
S-----S
углевод
гипервариабельные
участки лёгкой цепи
шарнирная
область
межцепочечные
дисульфидные
связи
внутрицепочечные
дисульфидные
связи
участок
связывания
комплемента
лёгкая цепь
тяжёлая цепь
расщепление
пепсином
гипервариабельные
участки тяжёлой

Легкая цепь
Fab
Fc
Тяжелая цепь
Легкая цепь
Fab
Fc
Тяжелая цепь

Наиболее важные и самыми распространенными Ig сыворотки мономерного класса (80%). Это
Наиболее важные и самыми распространенными Ig сыворотки мономерного класса (80%). Это
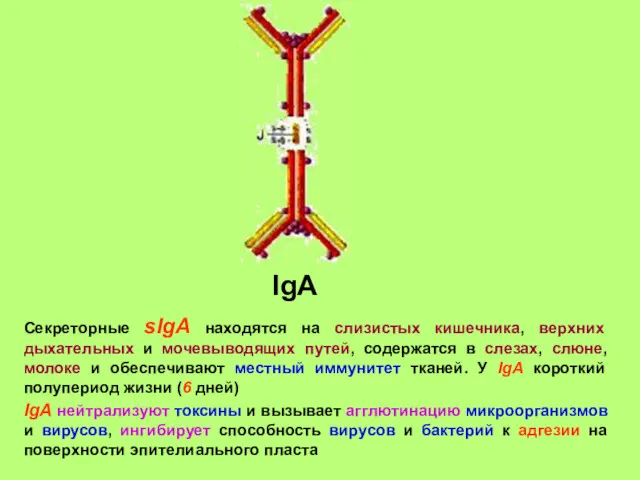
IgА
Секреторные sIgA находятся на слизистых кишечника, верхних дыхательных и мочевыводящих путей,
IgА
Секреторные sIgA находятся на слизистых кишечника, верхних дыхательных и мочевыводящих путей,

IgЕ
это реагины или гомоцитотропные АТ. Связываются своим Fc-фрагментом с высокоаффинными рецепторами
IgЕ
это реагины или гомоцитотропные АТ. Связываются своим Fc-фрагментом с высокоаффинными рецепторами

IgD
IgD локализуются в мембране В-лф, ПК и выполняют функции иммунноглобулиновых антигенраспознающих
IgD
IgD локализуются в мембране В-лф, ПК и выполняют функции иммунноглобулиновых антигенраспознающих
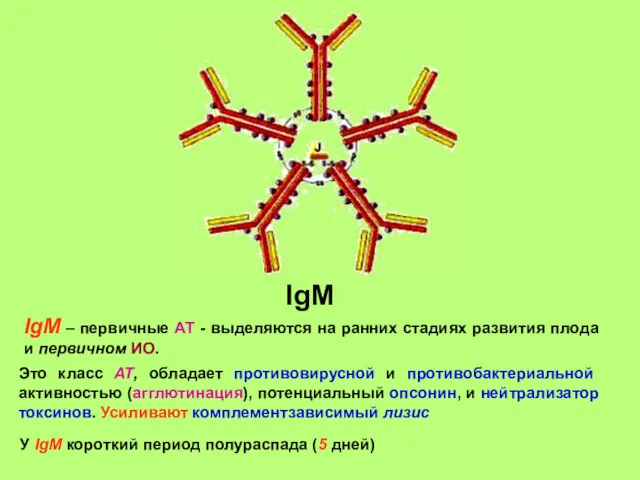
IgМ
IgМ – первичные АТ - выделяются на ранних стадиях развития плода
IgМ
IgМ – первичные АТ - выделяются на ранних стадиях развития плода

определяются Fab и Fc-фрагментами. С Fab-фрагментом связано понятие авидности АТ. С
определяются Fab и Fc-фрагментами. С Fab-фрагментом связано понятие авидности АТ. С

- иммобилизация бактерий осуществляется АТ против бактериальных ворсинок, нарушающими их подвижность
- иммобилизация бактерий осуществляется АТ против бактериальных ворсинок, нарушающими их подвижность

Клеточный иммунный ответ
развивается на чужеродные клетки, на клетки, пораженные вирусом, на
Клеточный иммунный ответ
развивается на чужеродные клетки, на клетки, пораженные вирусом, на
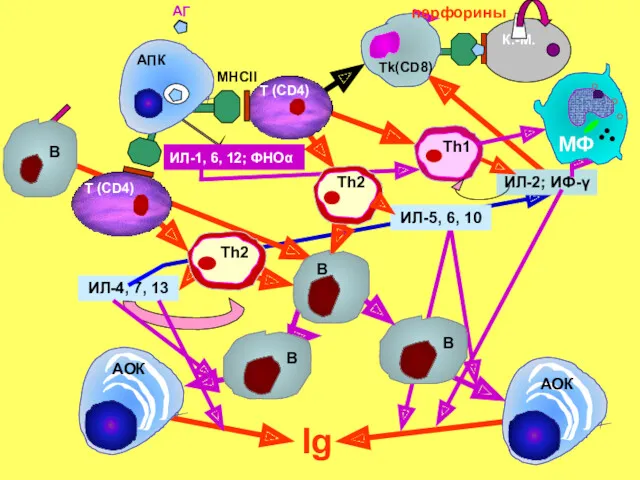
МНСII
АГ
Тh2
B
B
Ig
ИЛ-1, 6, 12; ФНОα
ИЛ-4, 7, 13
ИЛ-5, 6, 10
B
ИЛ-2;
МНСII
АГ
Тh2
B
B
Ig
ИЛ-1, 6, 12; ФНОα
ИЛ-4, 7, 13
ИЛ-5, 6, 10
B
ИЛ-2;

Селекция тимоцитов
Экспрессия ТКР на индивидуальном тимоците означает, что клетка вступает в
Селекция тимоцитов
Экспрессия ТКР на индивидуальном тимоците означает, что клетка вступает в

Узнавание "своего" осуществляется не всем АГраспознающим центром ТКР. Оставшаяся часть центра
Узнавание "своего" осуществляется не всем АГраспознающим центром ТКР. Оставшаяся часть центра

Специальных клеток для подавления ИО, не существует. Продуцируя медиаторы с противоположным
Специальных клеток для подавления ИО, не существует. Продуцируя медиаторы с противоположным

Результатом ИО является формирование клеток памяти, которые появляются в результате ИО
Результатом ИО является формирование клеток памяти, которые появляются в результате ИО
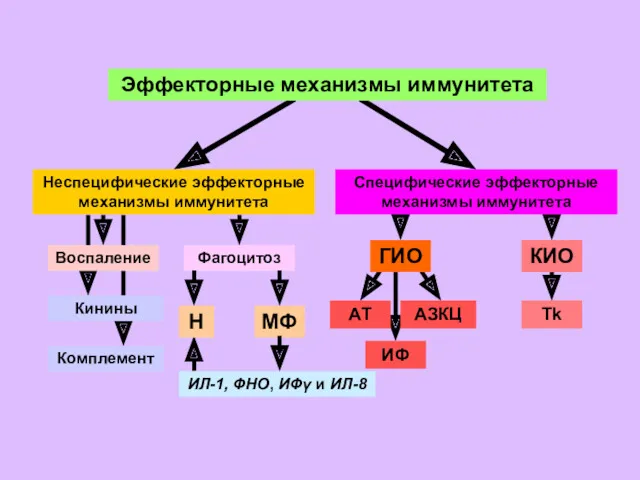
Эффекторные механизмы иммунитета
Неспецифические эффекторные механизмы иммунитета
Воспаление
Фагоцитоз
Н
МФ
ИЛ-1, ФНО, ИФγ
Эффекторные механизмы иммунитета
Неспецифические эффекторные механизмы иммунитета
Воспаление
Фагоцитоз
Н
МФ
ИЛ-1, ФНО, ИФγ

ИММУНОЛОГИЧЕСКАЯ ТОЛЕРАНТНОСТЬ
Лф должны распознавать собственные АГ тканевой совместимости, специфичные для
ИММУНОЛОГИЧЕСКАЯ ТОЛЕРАНТНОСТЬ
Лф должны распознавать собственные АГ тканевой совместимости, специфичные для

Иммунологическая толерантность характеризуется:
- отсутствием ответа на собственный АГ
- отсутствием элиминации АГ
Иммунологическая толерантность характеризуется:
- отсутствием ответа на собственный АГ
- отсутствием элиминации АГ

В результате позитивной и негативной селекции из тимуса в кровоток и
В результате позитивной и негативной селекции из тимуса в кровоток и

РЕГУЛЯЦИЯ ИММУНОГО ОТВЕТА
Эффективный ИО - результат взаимодействия между АГ и
РЕГУЛЯЦИЯ ИММУНОГО ОТВЕТА
Эффективный ИО - результат взаимодействия между АГ и

Характер ИО зависит от присутствия цитокинов в микроокружении Т-лф в момент
Характер ИО зависит от присутствия цитокинов в микроокружении Т-лф в момент

Опиоидные пептиды - связующее звено между нервной и иммунной системой. ЭНД
Опиоидные пептиды - связующее звено между нервной и иммунной системой. ЭНД
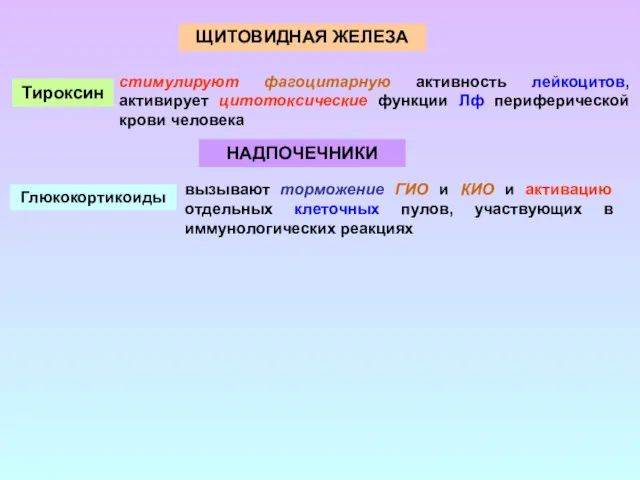
ЩИТОВИДНАЯ ЖЕЛЕЗА
Тироксин
стимулируют фагоцитарную активность лейкоцитов, активирует цитотоксические функции Лф периферической
ЩИТОВИДНАЯ ЖЕЛЕЗА
Тироксин
стимулируют фагоцитарную активность лейкоцитов, активирует цитотоксические функции Лф периферической

ДЕТСКИЕ ОСОБЕННОСТИ
Развивается блок ИС матери плода:
развивается толерантность к отцовским АГ
трансплацентарный перенос
ДЕТСКИЕ ОСОБЕННОСТИ
Развивается блок ИС матери плода:
развивается толерантность к отцовским АГ
трансплацентарный перенос
 Синдром разбитого сердца
Синдром разбитого сердца Классификация медицинских отходов. Работа по обращению с медицинскими отходами
Классификация медицинских отходов. Работа по обращению с медицинскими отходами Струс мозку
Струс мозку Технические регламенты и национальные стандарты по обеспечению оздоровительных услуг гостиничными организациями. (Лекция 6.2)
Технические регламенты и национальные стандарты по обеспечению оздоровительных услуг гостиничными организациями. (Лекция 6.2) Препараты выпускаемые фирмой ООО Альтфарм. Нестероидные противовоспалительные средства
Препараты выпускаемые фирмой ООО Альтфарм. Нестероидные противовоспалительные средства Жатыр мойны адам папилломавирусының инфекциясы
Жатыр мойны адам папилломавирусының инфекциясы Рак молочной железы. Скрининг и диагностика
Рак молочной железы. Скрининг и диагностика Исследование сосудов. Дополнительные методы исследования сердечно-сосудистой системы
Исследование сосудов. Дополнительные методы исследования сердечно-сосудистой системы Алалияның симптоматикасы мен механизмі
Алалияның симптоматикасы мен механизмі Алгоритм действий сотрудника аптеки. Часть 4
Алгоритм действий сотрудника аптеки. Часть 4 Острые респираторные заболевания у детей раннего и дошкольного возраста и их профилактика. (Лекция 7)
Острые респираторные заболевания у детей раннего и дошкольного возраста и их профилактика. (Лекция 7) Высшая нервная деятельность
Высшая нервная деятельность Bronchial asthma in children (treatment)
Bronchial asthma in children (treatment) Предметы гигиены полости рта. Методы чистки зубов
Предметы гигиены полости рта. Методы чистки зубов Дифференциальный диагноз заболеваний аноректальной области
Дифференциальный диагноз заболеваний аноректальной области Болезни оперированного желудка
Болезни оперированного желудка Итранатальные повреждения ЦНС (родовая травма)
Итранатальные повреждения ЦНС (родовая травма) Вскармливание детей 1 года жизни . Естественное вскармливание. Оценка питания ребенка при естественном вскармливании
Вскармливание детей 1 года жизни . Естественное вскармливание. Оценка питания ребенка при естественном вскармливании Остеомиелиттің емі
Остеомиелиттің емі Патологиялық анатомияның басты әдісі
Патологиялық анатомияның басты әдісі Классный час Профилактика ОРВИ
Классный час Профилактика ОРВИ Ультразвуковое исследование тазобедренного сустава
Ультразвуковое исследование тазобедренного сустава Болезнь Гоше
Болезнь Гоше Structura morfo-funcţională a intestinului
Structura morfo-funcţională a intestinului Роль воды и минеральных веществ в формировании потребительских свойств продовольственных товаров
Роль воды и минеральных веществ в формировании потребительских свойств продовольственных товаров Наука нозология
Наука нозология Оспа натуральная и ветряная
Оспа натуральная и ветряная Классический массаж. Приемы
Классический массаж. Приемы