Иммунопоэз: созревание Т - и В- клеточных рецепторов. Механизмы положительной и отрицательной селекции презентация
- Иммунопоэз: созревание Т - и В- клеточных рецепторов. Механизмы положительной и отрицательной селекции

Содержание
- 2. Иммунопоэз: созревание Т - и В- клеточных рецепторов. Роль факторов микроокружения. Механизмы положительной и отрицательной селекции.
- 3. Контроль исходного уровня знаний: фронтальный опрос Определение антигена. Виды антигенов. Основные свойства антигенов. Строение главного комплекса
- 4. ЦЕНТРАЛЬНЫЕ ОРГАНЫ ИММУНИТЕТА Центральные органы иммунитета - красный костный мозг и тимус. В центральных органах иммунитета
- 5. ЦЕНТРАЛЬНЫЕ ОРГАНЫ ИММУНИТЕТА В костном мозге происходит образование и дифференцировка всех типов клеток крови на основе
- 6. ЦЕНТРАЛЬНЫЕ ОРГАНЫ ИММУНИТЕТА Клетки становятся иммунокомпетентными – то есть способными различать разные классы чужеродных структур. Эта
- 8. Центральные органы иммунитета: тимус
- 9. ТИМУС Дольчатая структура с эпителиальными клетками стромы и соединительной тканью Строма обеспечивает микроокружение для развития и
- 10. Центральные органы иммунитета: тимус
- 11. Тимус - биологические часы: масса тимуса
- 12. Инволюция тимуса
- 13. Созревание Т-лимфоцитов в тимусе: 1 этап
- 14. Созревание Т-лимфоцитов в тимусе: 2 этап
- 15. ЦЕНТРАЛЬНЫЕ ОРГАНЫ ИММУНИТЕТА В центральных органах иммунитета происходят процессы селекции клонов лимфоцитов (Т-лимфоцитов - в тимусе,
- 16. Какие клетки выходят на периферию? В результате позитивной и негативной селекции в кровоток поступают только те
- 17. Интенсивность отбора тимоцитов T клетки созревают в тимусе, но гораздо больше Т- клеток погибает в тимусе
- 18. T клетки созревают в тимусе, но гораздо больше Т- клеток погибает 98% клеток погибает в тимусе
- 19. Строение Т рецепторов Т клеточный рецептор имеет α и β -цепи (есть альтернативные рецепторы, которые имеют
- 20. Т-клеточный рецептор Vα Vβ Cα Cβ карбогидраты моновалентность для сравнения:BCR - Ig Fab фрагмент Нет альтернативных
- 21. Строение Т рецепторного комплекса На клеточной поверхности αβ -Т клеточный рецептор (или γδ) расположен в непосредственной
- 23. Строение корецепторов Корецепторы расположены на мембране Т лимфоцита рядом с комплексом TCR/ CD3. Корецепторы «узнают» молекулы
- 25. Процесс созревания тимоцитов: этапы формирования корецепторов Корецепторы: CD4 – распознает молекулы МНС II CD8 – распознает
- 26. Процесс созревания тимоцитов: этапы формирования корецепторов При переходе в мозговой слой клетки теряют либо CD4, либо
- 27. Сигнал с CD4 отменяет экспрессию CD8 и наоборот ДВОЙНЫЕ ПОЗИТИВНЫЕ ТИМОЦИТЫ Эпителий тимуса Переход от двойных
- 28. Реаранжировка генов, кодирующих β -цепь Т рецептора Изначальная конфигурация генов, кодирующих β цепь
- 29. Реаранжировка генов, кодирующих Т рецептор, путем соматической рекомбинации 1 этап – слияние генов D-J 2 этап
- 30. Реаранжировка генов, кодирующих α -цепь Т рецептора, сборка Т рецептора Те же этапы, что и при
- 31. Каким образом развивается аутотолерантность к антигенам, отсутствующим в тимусе? T клетки, несущие TcR , и вступающие
- 32. Механизмы формирования периферической толерантность или анергии роль АПК: антигенпрезентирующие клетки не выполняют функцию костимуляции, т.е. не
- 33. Костимуляция Процесс презентации антигенов сопровождается костимуляцией в результате взаимодействия молекул и их лигандов, экспрессирующихся на поверхностных
- 34. Ко- стимулирующие молекулы
- 35. Гипотезы повреждения и костимуляции Полная экспрессия функций Т лимфоцитов зависит от того, когда и где экспрессируются
- 36. Гипотеза угрозы АПК , определившие сигналы угрозы, экспрессируют костимулирующие молекулы, активируют Т – клетки и иммунный
- 37. Зрелые Т-лимфоциты Зрелые Т-лимфоциты до встречи с антигеном называются «наивными» и составляют большую часть общего пула
- 38. ОТКРЫТИЕ В - КЛЕТОЧНОГО ИММУНИТЕТА 1954 - Bruce Glick, США Изучение функции Фабрициевой сумки (bursa Fabricius),
- 39. Происхождение В клеток и орган, в котором созревают В-клетки После рождения их развитие продолжается в костном
- 40. Костный мозг
- 41. Развитие В клеток в костном мозге Костный мозг обеспечивает МИКРООКРУЖЕНИЕ ДЛЯ СОЗРЕВАНИЯ , ДИФФЕРЕНЦИРОВКИ И РАЗВИТИЯ
- 42. X X X Схема развития В – клеток в костном мозге
- 43. Развитие В клеток в костном мозге Выжившие клетки продолжают созревать и достигают центрального венозного синуса. На
- 44. Клетки стромы Созревающие В клетки
- 45. Развитие В клеток в костном мозге Более 75% созревающих в костном мозге В-клеток не попадает в
- 46. B B Клетка стромы
- 47. Постулаты теории клональной селекции Каждый В лимфоцит имеет рецептор уникальной специфичности. Высокоаффинное (прочное) взаимодействие рецептора с
- 48. Y Y Y Y Y Y Зрелый В-лимфоцит выходит на периферию Y Y В - клеточная
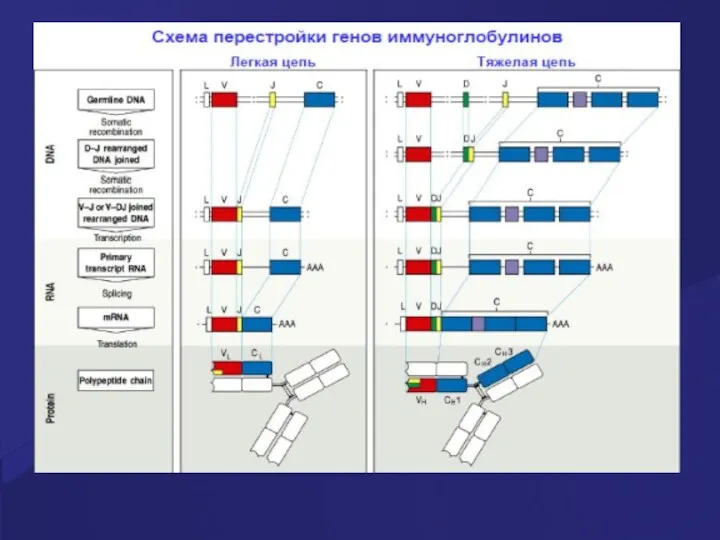
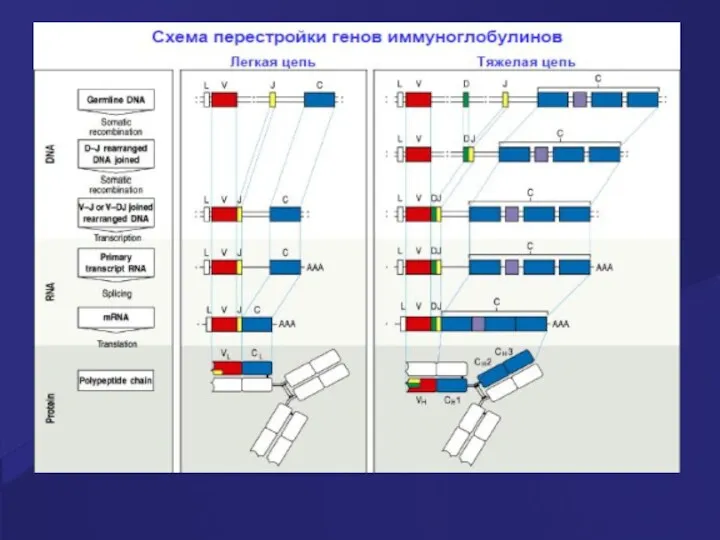
- 49. Стадии развития В клеток На каждой стадии развития происходит реаранжировка генов тяжелых и легких цепей Ig,
- 51. Гены иммуноглобулинов (Ig) Каждая молекула Ig состоит из 2 тяжелых (Н) и двух легких (L) цепей,
- 52. Гены Ig молекул Для вариабельных областей существует множество генов (V1-Vn), а для константой части молекулы Ig
- 53. Гены Ig молекул Рекомбинации ограниченного числа генных сегментов V, D и J создают бесконечное число вариабельных
- 54. Развитие В-лимфоцитов Из костного мозга уже отобранные В-лимфоциты попадают с током крови в первичные фолликулы селезенки.
- 56. Стадии дифференцировки определяются реаранжировкой генов Ig Стадии созревания конфигурация генов IgH Стволовая клетка Ранний про-В Поздний
- 57. Реаранжировка генов, кодирующих легкие цепи молекулы Ig После завершения перестройки (реаранжировки) генов, кодирующих тяжелые цепи молекулы
- 60. 5 источников разнообразия V-областей Н- и L -цепей молекул Ig 1. Многочисленность гаметных генов. Имеется большое
- 61. 5 источников разнообразия V-областей Н- и L -цепей молекул Ig 3.Соматическая рекомбинация В онтогенезе В-клеток происходит
- 62. 5 источников разнообразия V-областей Н- и L -цепей молекул Ig 4.Генная конверсия. Отрезки ДНК, принадлежащие ряду
- 63. 5 источников разнообразия V-областей Н- и L -цепей молекул Ig 5. Вставка добавочных нуклеотидов. При рекомбинации
- 64. Положительная и отрицательная селекция В лимфоцитов в костном мозге + селекция происходит при взаимодействии В -
- 65. Выход зрелых В клеток на периферию Из костного мозга выходят только те В –лимфоциты, у которых
- 66. Каким образом В клетки могут экспрессировать IgM и IgD одновременно? Два типа РНК могут образоваться одновременно
- 67. Рециркулирующие В – клетки встречаются со «своим» антигеном в лимфоузле
- 68. Распознал неауто-АГ на периферии Ig - секретирующая плазматическая клетка Дифференцировка В – клеток на периферии Зрелый
- 70. Субпопуляции В лимфоцитов: В1 и В2 В 2 лимфоциты связывают белковые антигены, им нужна помощь Т-хелперов,
- 71. Субпопуляция В1 лимфоцитов После активации В1 клетки секретируют анти-полисахаридные антитела класса М (IgM), которые присоединяются к
- 72. В1 лимфоциты
- 73. В 1 лимфоциты – клетки врожденного иммунитета Распознают эпитопы АГ с повторяющимися структурами - фосфотидилхолин, липополисахариды
- 74. Взаимодействие АПК, Т- и В- лимфоцитов в ходе иммунного ответа на АГ
- 75. Вопросы к занятию № 4 Какова роль тимуса в процессе дифференцировки Т-лимфоцитов? Каков биологический смысл положительной
- 76. Тестовые задания к занятию №4 1. Антигеннезависимая дифференцировка Т лимфоцитов происходит в: щитовидной железе тимусе лимфатических
- 77. Тестовые задания к занятию №4 3. Общим маркером Т-лимфоцитов является молекула: CD 3 CD 4 CD
- 78. Тестовые задания к занятию №3 5. Для цитотоксических Т-лимфоцитов характерны маркеры: CD 3 ; CD 4
- 79. Тестовые задания к занятию № 4 7. Зародышевые гены тяжелых цепей иммуноглобулинов включают регионы: B DP
- 80. Тестовые задания к занятию № 4 9. На поверхности зрелых В-лимфоцитов присутствуют в качестве В-рецепторов: Ig
- 82. Скачать презентацию

Иммунопоэз: созревание Т - и В- клеточных рецепторов.
Роль факторов микроокружения.
Иммунопоэз: созревание Т - и В- клеточных рецепторов. Роль факторов микроокружения.

Контроль исходного уровня знаний: фронтальный опрос
Определение антигена.
Виды антигенов.
Основные свойства антигенов.
Строение
Контроль исходного уровня знаний: фронтальный опрос
Определение антигена.
Виды антигенов.
Основные свойства антигенов.
Строение

ЦЕНТРАЛЬНЫЕ ОРГАНЫ ИММУНИТЕТА
Центральные органы иммунитета - красный костный мозг и
ЦЕНТРАЛЬНЫЕ ОРГАНЫ ИММУНИТЕТА
Центральные органы иммунитета - красный костный мозг и

ЦЕНТРАЛЬНЫЕ ОРГАНЫ ИММУНИТЕТА
В костном мозге происходит образование и дифференцировка всех типов
ЦЕНТРАЛЬНЫЕ ОРГАНЫ ИММУНИТЕТА
В костном мозге происходит образование и дифференцировка всех типов

ЦЕНТРАЛЬНЫЕ ОРГАНЫ ИММУНИТЕТА
Клетки становятся иммунокомпетентными – то есть способными различать разные
ЦЕНТРАЛЬНЫЕ ОРГАНЫ ИММУНИТЕТА
Клетки становятся иммунокомпетентными – то есть способными различать разные
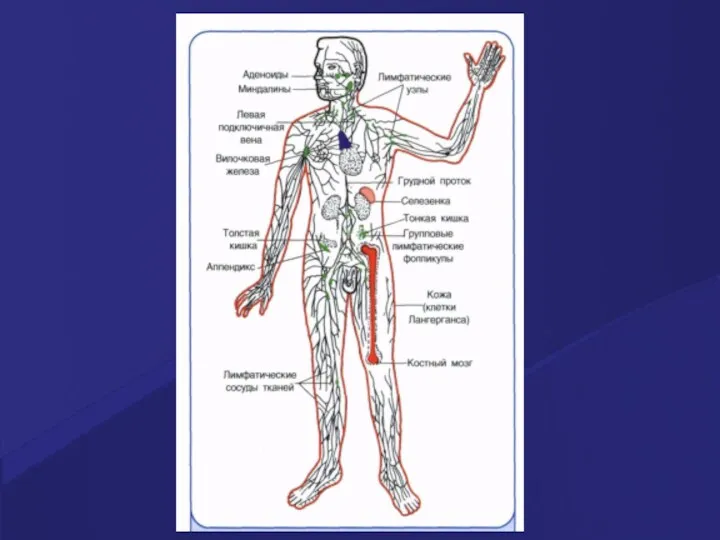
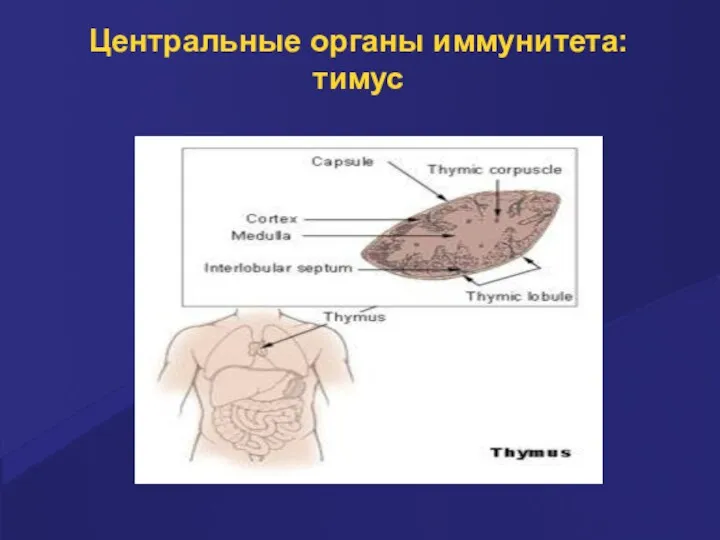
Центральные органы иммунитета: тимус
Центральные органы иммунитета: тимус
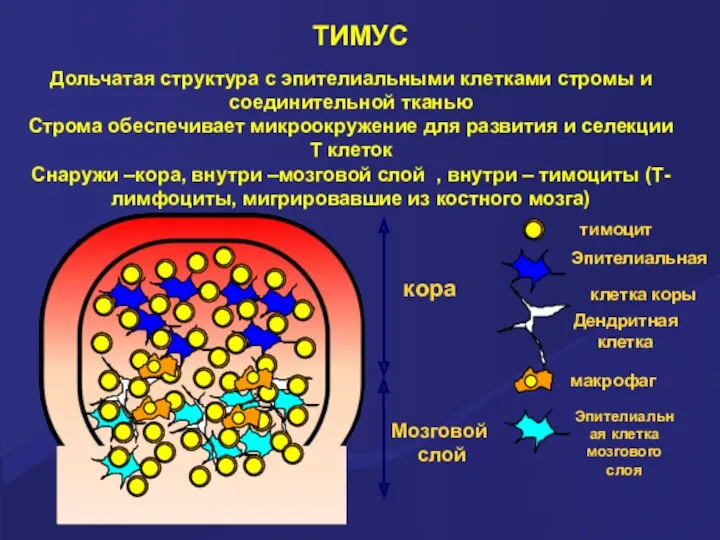
ТИМУС
Дольчатая структура с эпителиальными клетками стромы и соединительной тканью
Строма обеспечивает микроокружение
ТИМУС
Дольчатая структура с эпителиальными клетками стромы и соединительной тканью
Строма обеспечивает микроокружение
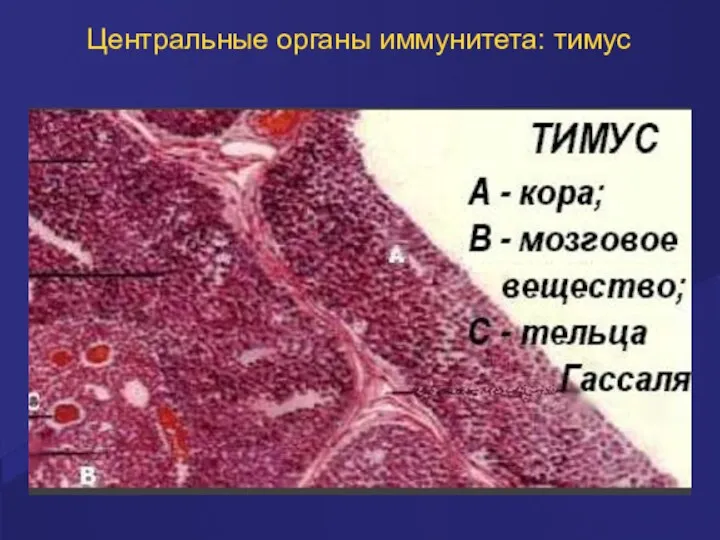
Центральные органы иммунитета: тимус
Центральные органы иммунитета: тимус
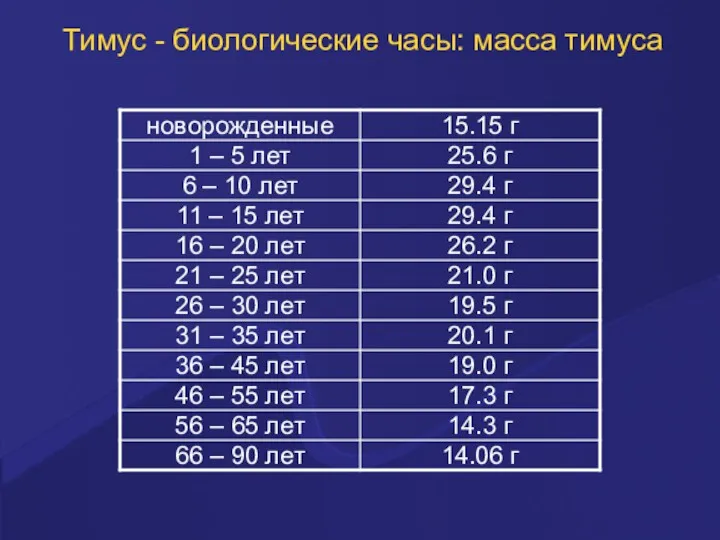
Тимус - биологические часы: масса тимуса
Тимус - биологические часы: масса тимуса
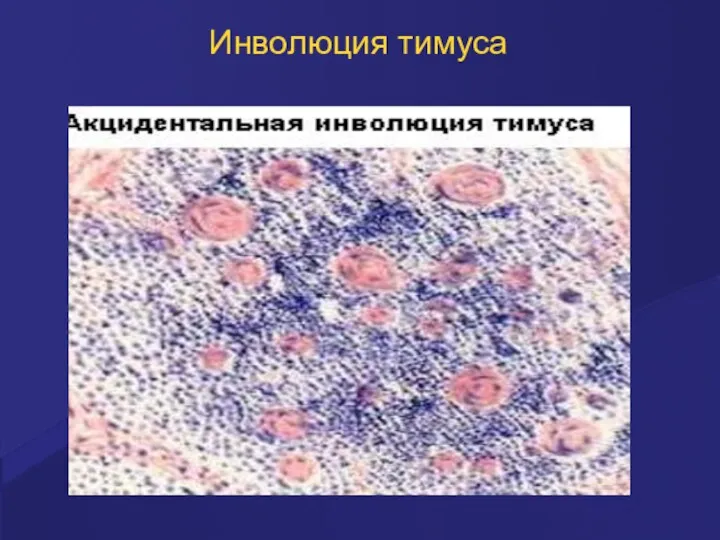
Инволюция тимуса
Инволюция тимуса

Созревание Т-лимфоцитов в тимусе: 1 этап
Созревание Т-лимфоцитов в тимусе: 1 этап

Созревание Т-лимфоцитов в тимусе: 2 этап
Созревание Т-лимфоцитов в тимусе: 2 этап

ЦЕНТРАЛЬНЫЕ ОРГАНЫ ИММУНИТЕТА
В центральных органах иммунитета происходят процессы селекции клонов лимфоцитов
ЦЕНТРАЛЬНЫЕ ОРГАНЫ ИММУНИТЕТА
В центральных органах иммунитета происходят процессы селекции клонов лимфоцитов

Какие клетки выходят на периферию?
В результате позитивной и негативной селекции в
Какие клетки выходят на периферию?
В результате позитивной и негативной селекции в

Интенсивность отбора тимоцитов
T клетки созревают в тимусе,
но гораздо больше
Интенсивность отбора тимоцитов
T клетки созревают в тимусе,
но гораздо больше
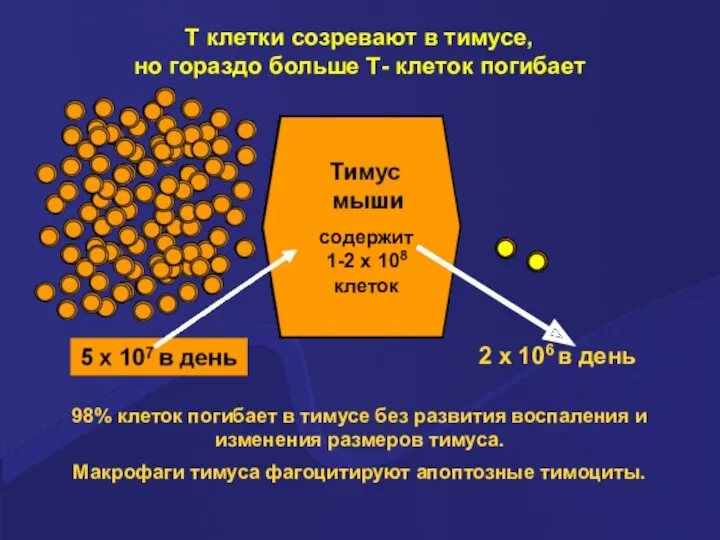
T клетки созревают в тимусе,
но гораздо больше Т- клеток погибает
98%
T клетки созревают в тимусе,
но гораздо больше Т- клеток погибает
98%

Строение Т рецепторов
Т клеточный рецептор имеет α и β -цепи (есть
Строение Т рецепторов
Т клеточный рецептор имеет α и β -цепи (есть
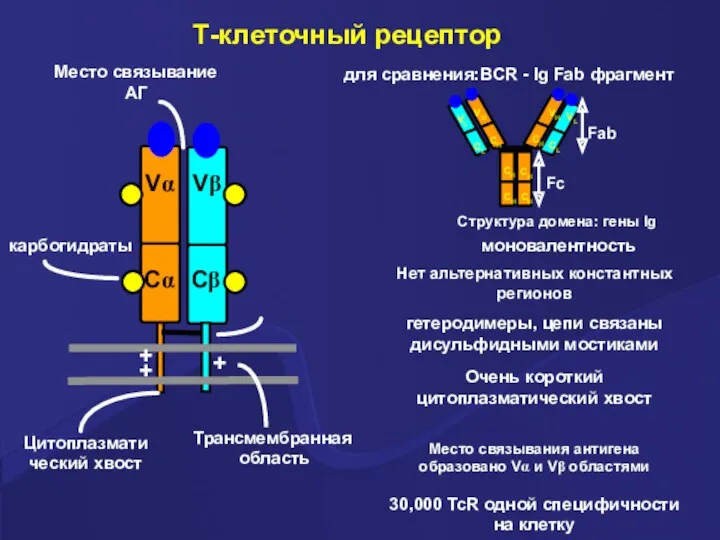
Т-клеточный рецептор
Vα
Vβ
Cα
Cβ
карбогидраты
моновалентность
для сравнения:BCR - Ig Fab фрагмент
Нет альтернативных константных регионов
Структура домена:
Т-клеточный рецептор
Vα
Vβ
Cα
Cβ
карбогидраты
моновалентность
для сравнения:BCR - Ig Fab фрагмент
Нет альтернативных константных регионов
Структура домена:

Строение Т рецепторного комплекса
На клеточной поверхности αβ -Т клеточный рецептор (или
Строение Т рецепторного комплекса
На клеточной поверхности αβ -Т клеточный рецептор (или
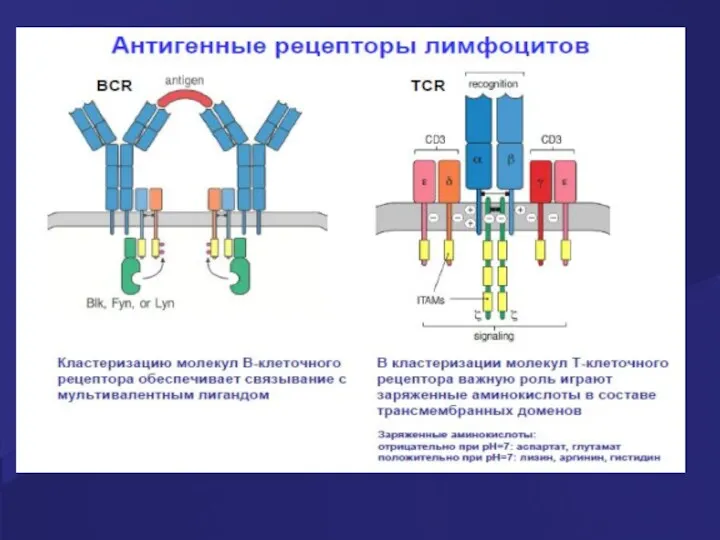

Строение корецепторов
Корецепторы расположены на мембране Т лимфоцита рядом с комплексом TCR/
Строение корецепторов
Корецепторы расположены на мембране Т лимфоцита рядом с комплексом TCR/
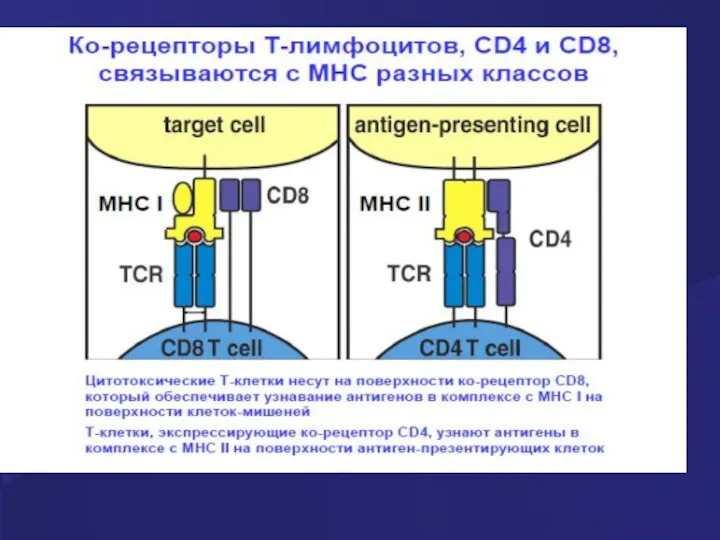

Процесс созревания тимоцитов:
этапы формирования корецепторов
Корецепторы:
CD4 – распознает молекулы МНС II
Процесс созревания тимоцитов:
этапы формирования корецепторов
Корецепторы:
CD4 – распознает молекулы МНС II

Процесс созревания тимоцитов:
этапы формирования корецепторов
При переходе в мозговой слой клетки теряют
Процесс созревания тимоцитов:
этапы формирования корецепторов
При переходе в мозговой слой клетки теряют
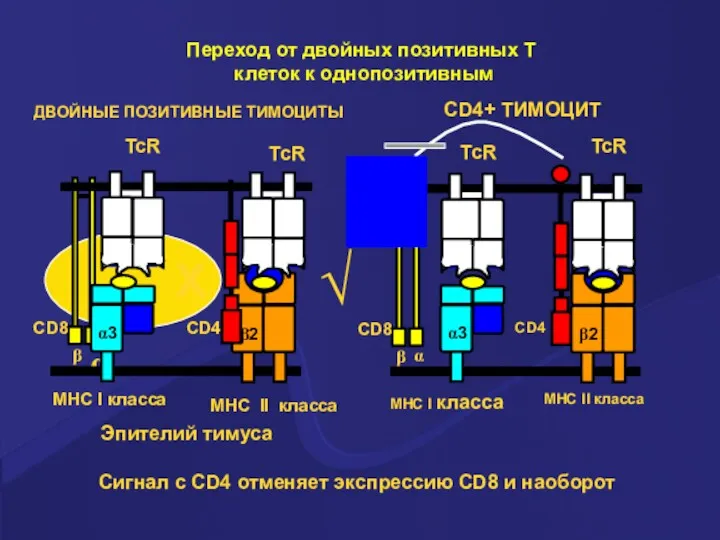
Сигнал с CD4 отменяет экспрессию CD8 и наоборот
ДВОЙНЫЕ ПОЗИТИВНЫЕ ТИМОЦИТЫ
Эпителий тимуса
Переход
Сигнал с CD4 отменяет экспрессию CD8 и наоборот
ДВОЙНЫЕ ПОЗИТИВНЫЕ ТИМОЦИТЫ
Эпителий тимуса
Переход
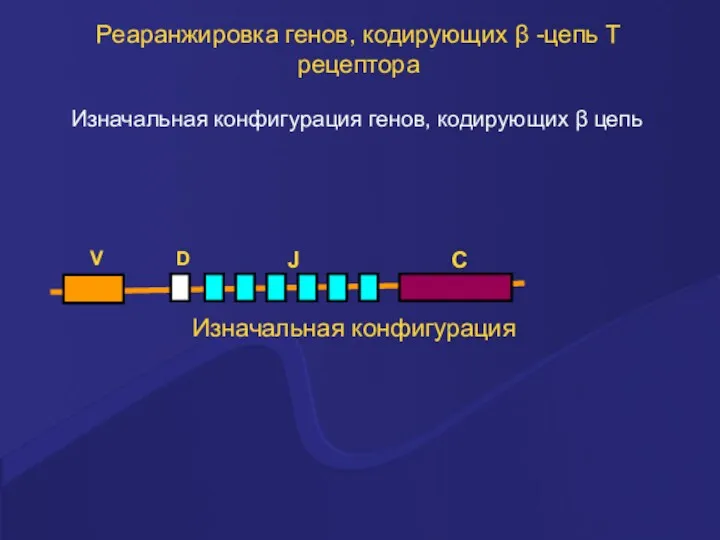
Реаранжировка генов, кодирующих β -цепь Т рецептора
Изначальная конфигурация генов, кодирующих β
Реаранжировка генов, кодирующих β -цепь Т рецептора
Изначальная конфигурация генов, кодирующих β
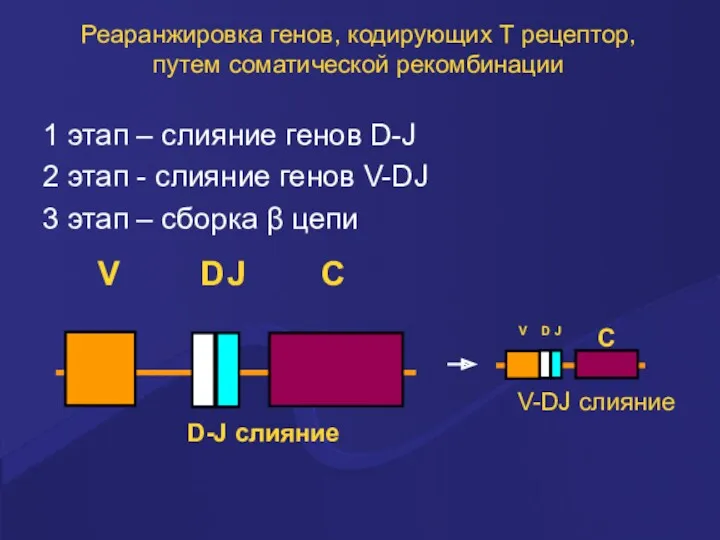
Реаранжировка генов, кодирующих Т рецептор, путем соматической рекомбинации
1 этап – слияние
Реаранжировка генов, кодирующих Т рецептор, путем соматической рекомбинации
1 этап – слияние

Реаранжировка генов, кодирующих α -цепь Т рецептора, сборка Т рецептора
Те же
Реаранжировка генов, кодирующих α -цепь Т рецептора, сборка Т рецептора
Те же

Каким образом развивается
аутотолерантность к антигенам,
отсутствующим в тимусе?
T клетки, несущие
Каким образом развивается
аутотолерантность к антигенам,
отсутствующим в тимусе?
T клетки, несущие

Механизмы формирования периферической толерантность или анергии
роль АПК:
антигенпрезентирующие клетки не
Механизмы формирования периферической толерантность или анергии
роль АПК:
антигенпрезентирующие клетки не

Костимуляция
Процесс презентации антигенов сопровождается костимуляцией в результате взаимодействия молекул и их
Костимуляция
Процесс презентации антигенов сопровождается костимуляцией в результате взаимодействия молекул и их
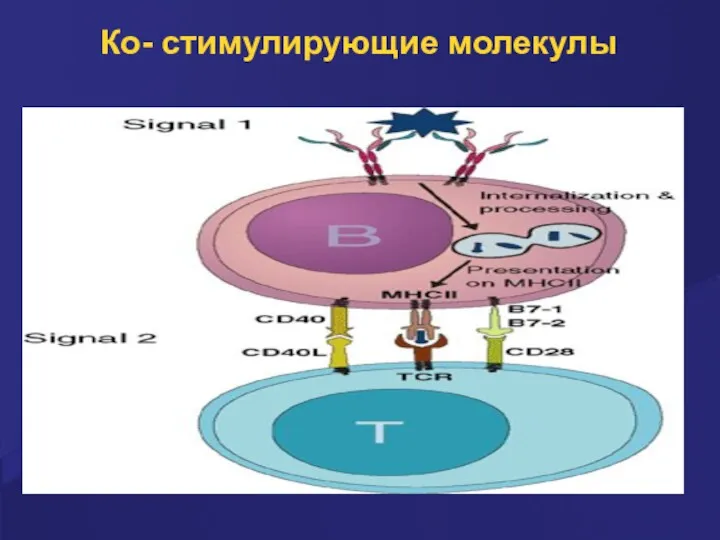
Ко- стимулирующие молекулы
Ко- стимулирующие молекулы
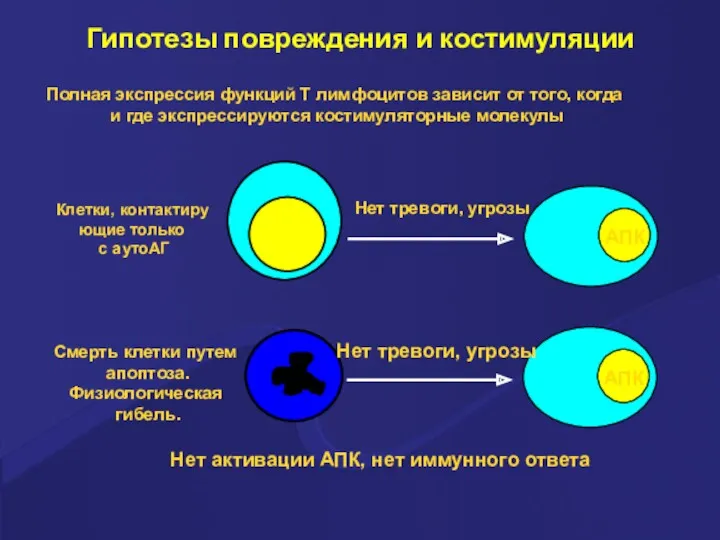
Гипотезы повреждения и костимуляции
Полная экспрессия функций Т лимфоцитов зависит от того,
Гипотезы повреждения и костимуляции
Полная экспрессия функций Т лимфоцитов зависит от того,
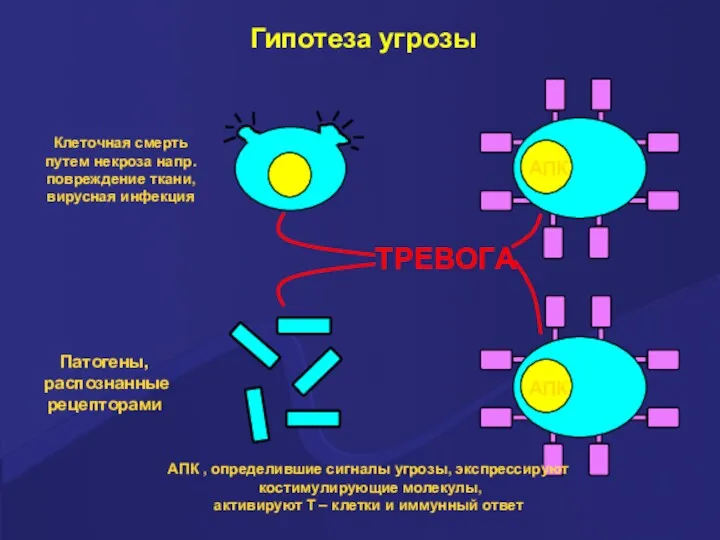
Гипотеза угрозы
АПК , определившие сигналы угрозы, экспрессируют
костимулирующие молекулы,
активируют Т
Гипотеза угрозы
АПК , определившие сигналы угрозы, экспрессируют
костимулирующие молекулы,
активируют Т

Зрелые Т-лимфоциты
Зрелые Т-лимфоциты до встречи с антигеном называются «наивными» и составляют
Зрелые Т-лимфоциты
Зрелые Т-лимфоциты до встречи с антигеном называются «наивными» и составляют
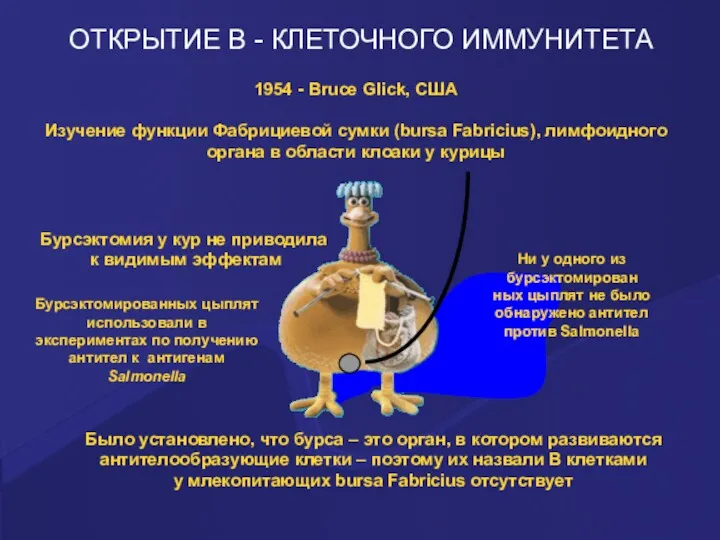
ОТКРЫТИЕ В - КЛЕТОЧНОГО ИММУНИТЕТА
1954 - Bruce Glick, США
Изучение функции Фабрициевой
ОТКРЫТИЕ В - КЛЕТОЧНОГО ИММУНИТЕТА
1954 - Bruce Glick, США
Изучение функции Фабрициевой
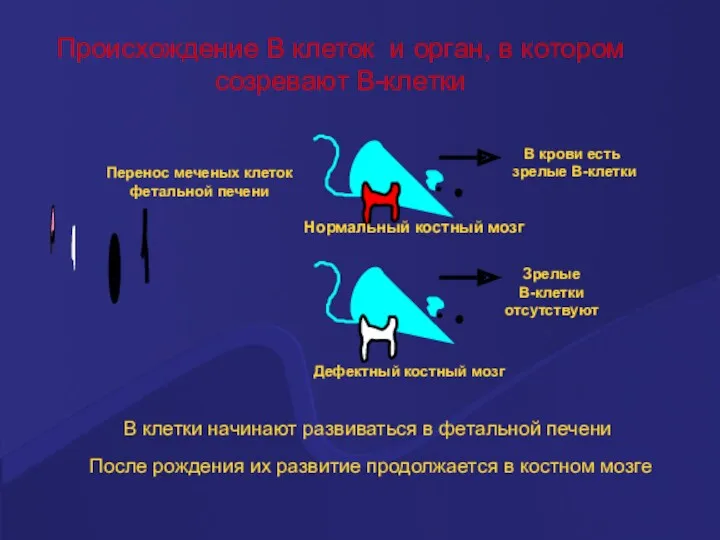
Происхождение В клеток и орган, в котором
созревают В-клетки
После рождения их
Происхождение В клеток и орган, в котором
созревают В-клетки
После рождения их
Костный мозг
Костный мозг
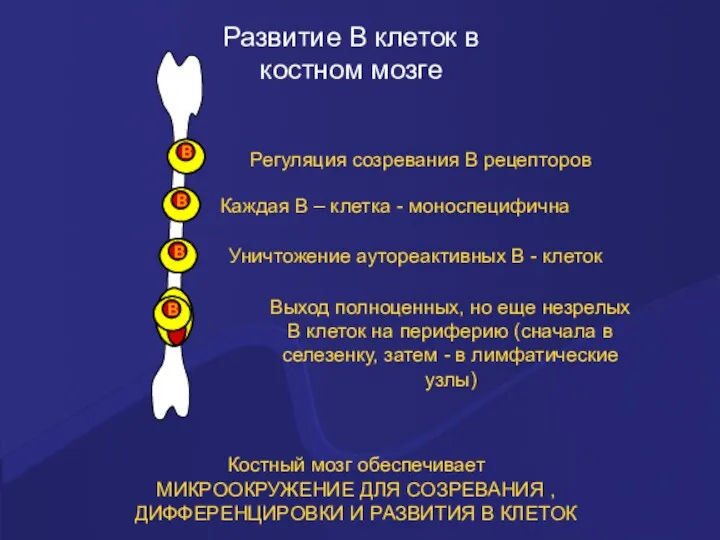
Развитие В клеток в
костном мозге
Костный мозг обеспечивает
МИКРООКРУЖЕНИЕ ДЛЯ СОЗРЕВАНИЯ , ДИФФЕРЕНЦИРОВКИ
Развитие В клеток в
костном мозге
Костный мозг обеспечивает
МИКРООКРУЖЕНИЕ ДЛЯ СОЗРЕВАНИЯ , ДИФФЕРЕНЦИРОВКИ
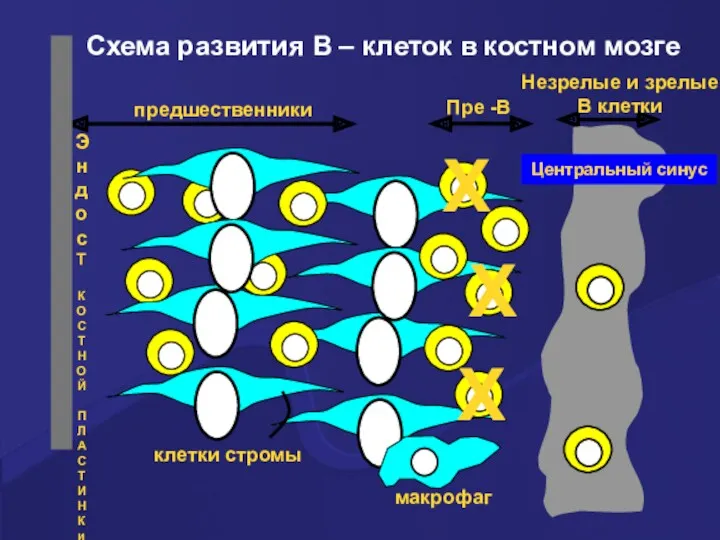
X
X
X
Схема развития В – клеток в костном мозге
X
X
X
Схема развития В – клеток в костном мозге

Развитие В клеток в костном мозге
Выжившие клетки продолжают созревать и достигают
Развитие В клеток в костном мозге
Выжившие клетки продолжают созревать и достигают
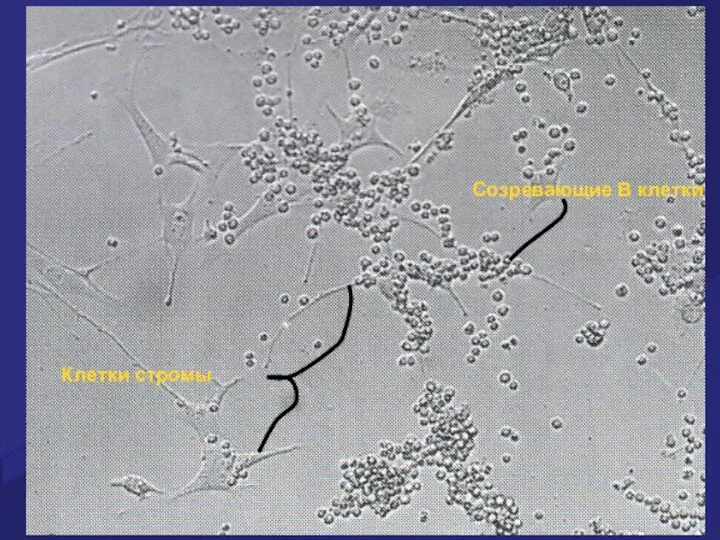
Клетки стромы
Созревающие В клетки
Клетки стромы
Созревающие В клетки

Развитие В клеток в костном мозге
Более 75% созревающих в костном мозге
Развитие В клеток в костном мозге
Более 75% созревающих в костном мозге
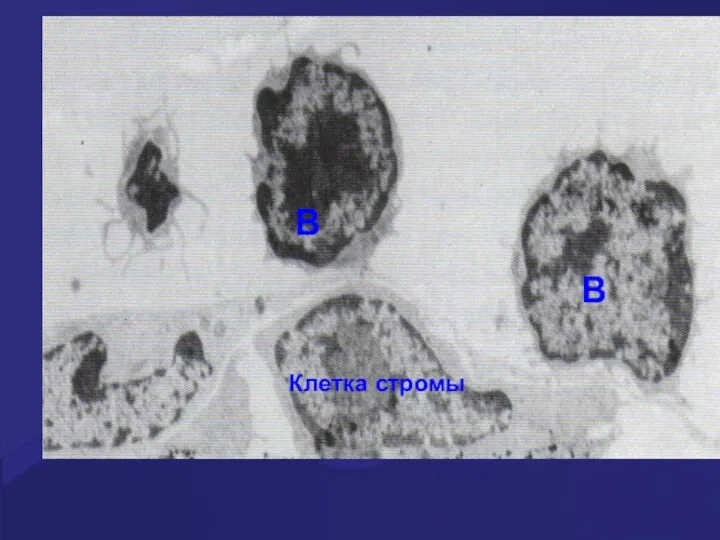
B
B
Клетка стромы
B
B
Клетка стромы

Постулаты теории клональной селекции
Каждый В лимфоцит имеет рецептор уникальной специфичности.
Высокоаффинное (прочное)
Постулаты теории клональной селекции
Каждый В лимфоцит имеет рецептор уникальной специфичности.
Высокоаффинное (прочное)
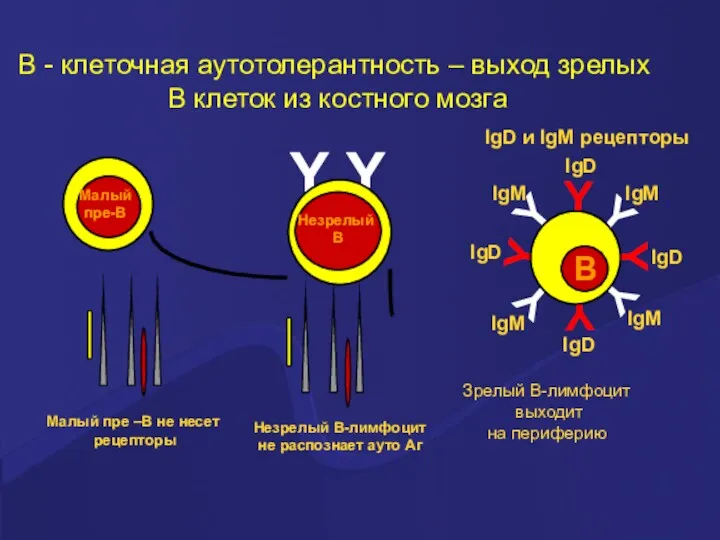
Y
Y
Y
Y
Y
Y
Зрелый В-лимфоцит
выходит
на периферию
Y
Y
В - клеточная аутотолерантность – выход зрелых
Y
Y
Y
Y
Y
Y
Зрелый В-лимфоцит
выходит
на периферию
Y
Y
В - клеточная аутотолерантность – выход зрелых
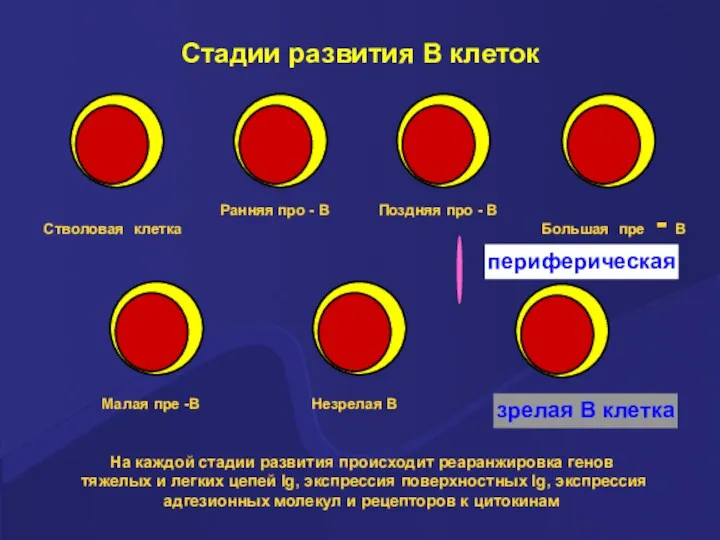
Стадии развития В клеток
На каждой стадии развития происходит реаранжировка генов тяжелых
Стадии развития В клеток
На каждой стадии развития происходит реаранжировка генов тяжелых
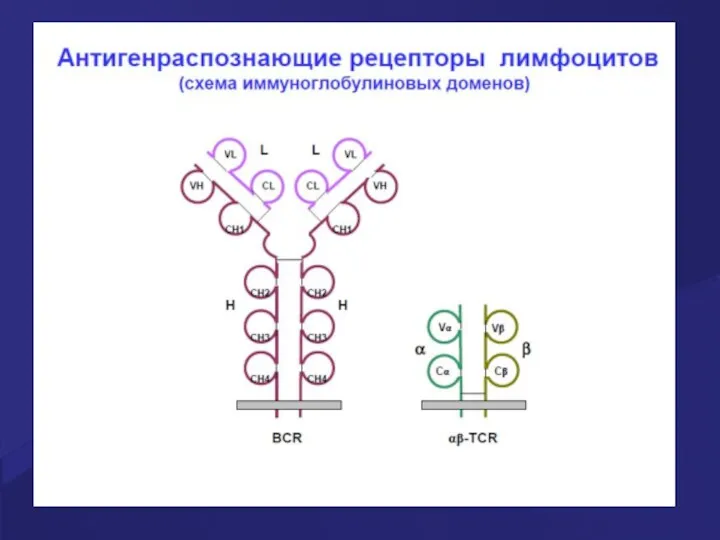

Гены иммуноглобулинов (Ig)
Каждая молекула Ig состоит из 2 тяжелых (Н) и
Гены иммуноглобулинов (Ig)
Каждая молекула Ig состоит из 2 тяжелых (Н) и

Гены Ig молекул
Для вариабельных областей существует множество генов (V1-Vn), а для
Гены Ig молекул
Для вариабельных областей существует множество генов (V1-Vn), а для

Гены Ig молекул
Рекомбинации ограниченного числа генных сегментов V, D и J
Гены Ig молекул
Рекомбинации ограниченного числа генных сегментов V, D и J

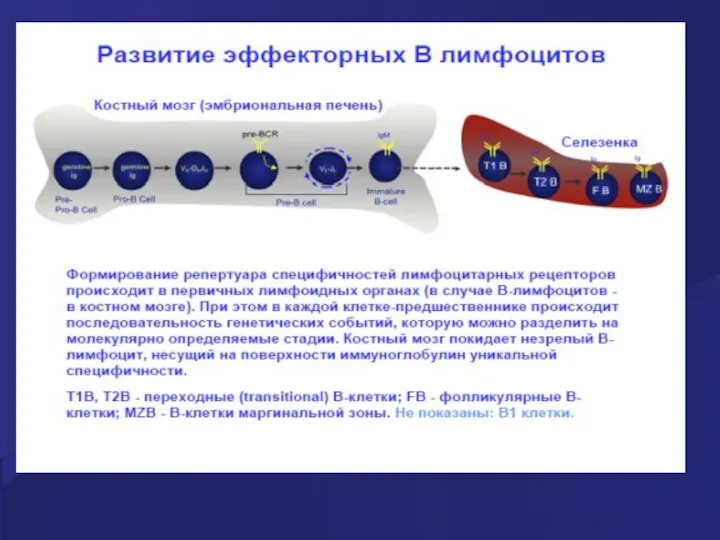
Развитие В-лимфоцитов
Из костного мозга уже отобранные В-лимфоциты попадают с током крови
Развитие В-лимфоцитов
Из костного мозга уже отобранные В-лимфоциты попадают с током крови
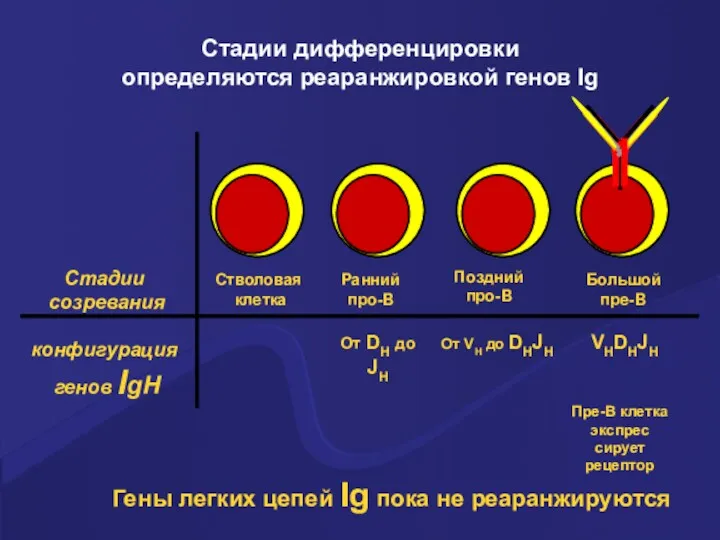
Стадии дифференцировки
определяются реаранжировкой генов Ig
Стадии
созревания
конфигурация
генов IgH
Стволовая
клетка
Ранний
про-В
Поздний
Стадии дифференцировки
определяются реаранжировкой генов Ig
Стадии
созревания
конфигурация
генов IgH
Стволовая
клетка
Ранний
про-В
Поздний

Реаранжировка генов, кодирующих легкие цепи молекулы Ig
После завершения перестройки (реаранжировки) генов,
Реаранжировка генов, кодирующих легкие цепи молекулы Ig
После завершения перестройки (реаранжировки) генов,

5 источников разнообразия V-областей Н- и L -цепей молекул Ig
1. Многочисленность
5 источников разнообразия V-областей Н- и L -цепей молекул Ig
1. Многочисленность

5 источников разнообразия V-областей Н- и L -цепей молекул Ig
3.Соматическая рекомбинация
В
5 источников разнообразия V-областей Н- и L -цепей молекул Ig
3.Соматическая рекомбинация
В

5 источников разнообразия V-областей Н- и L -цепей молекул Ig
4.Генная
5 источников разнообразия V-областей Н- и L -цепей молекул Ig
4.Генная

5 источников разнообразия V-областей Н- и L -цепей молекул Ig
5. Вставка
5 источников разнообразия V-областей Н- и L -цепей молекул Ig
5. Вставка

Положительная и отрицательная селекция В лимфоцитов в костном мозге
+ селекция происходит
Положительная и отрицательная селекция В лимфоцитов в костном мозге
+ селекция происходит

Выход зрелых В клеток на периферию
Из костного мозга выходят только те
Выход зрелых В клеток на периферию
Из костного мозга выходят только те
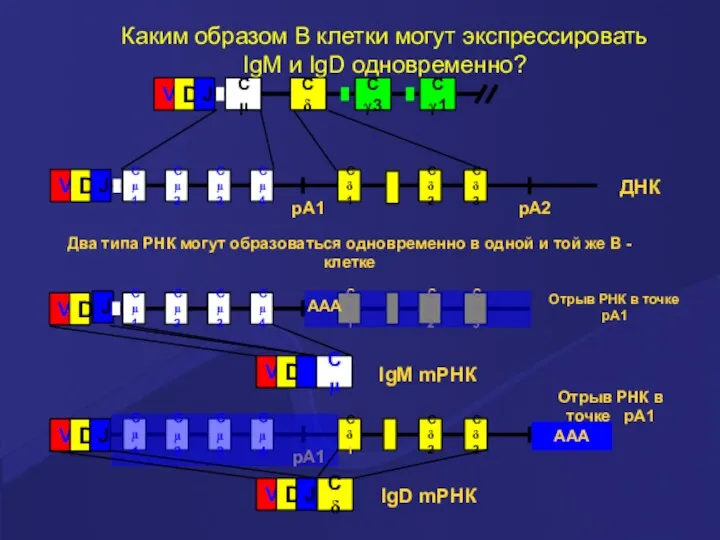
Каким образом В клетки могут экспрессировать
IgM и IgD одновременно?
Два типа РНК
Каким образом В клетки могут экспрессировать
IgM и IgD одновременно?
Два типа РНК
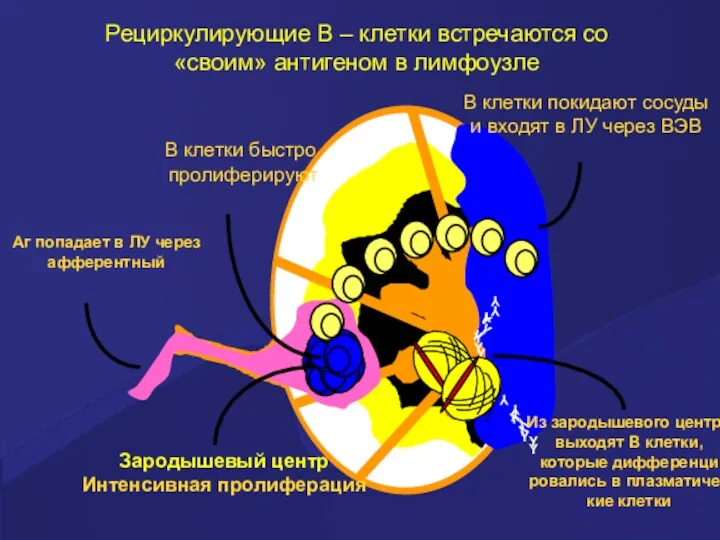
Рециркулирующие В – клетки встречаются со «своим» антигеном в лимфоузле
Рециркулирующие В – клетки встречаются со «своим» антигеном в лимфоузле
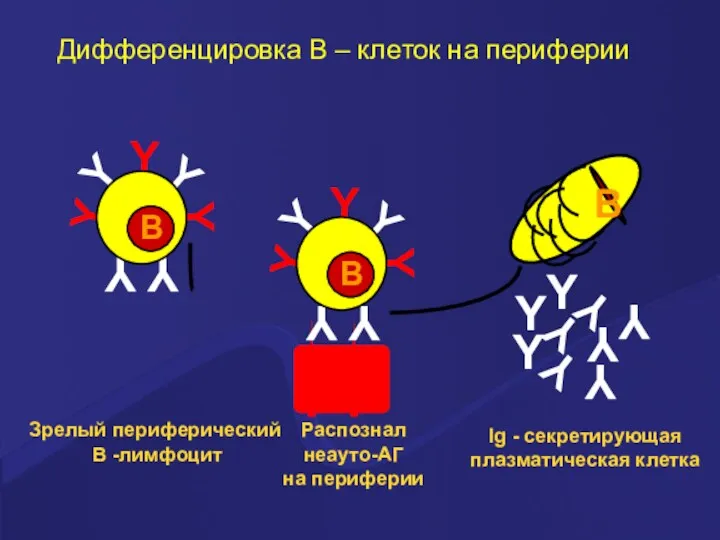
Распознал
неауто-АГ
на периферии
Ig - секретирующая
плазматическая клетка
Дифференцировка В –
Распознал
неауто-АГ
на периферии
Ig - секретирующая
плазматическая клетка
Дифференцировка В –


Субпопуляции В лимфоцитов: В1 и В2
В 2 лимфоциты связывают белковые антигены,
Субпопуляции В лимфоцитов: В1 и В2
В 2 лимфоциты связывают белковые антигены,

Субпопуляция В1 лимфоцитов
После активации В1 клетки секретируют анти-полисахаридные антитела класса М
Субпопуляция В1 лимфоцитов
После активации В1 клетки секретируют анти-полисахаридные антитела класса М
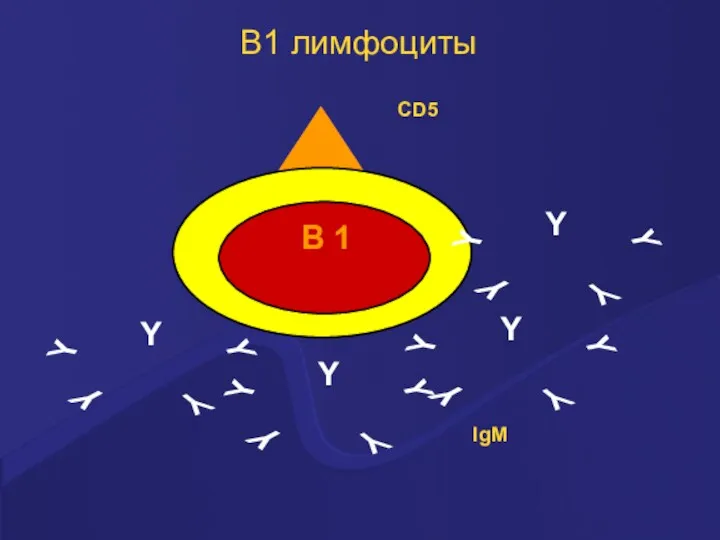
В1 лимфоциты
В1 лимфоциты

В 1 лимфоциты – клетки врожденного иммунитета
Распознают эпитопы АГ с
В 1 лимфоциты – клетки врожденного иммунитета
Распознают эпитопы АГ с
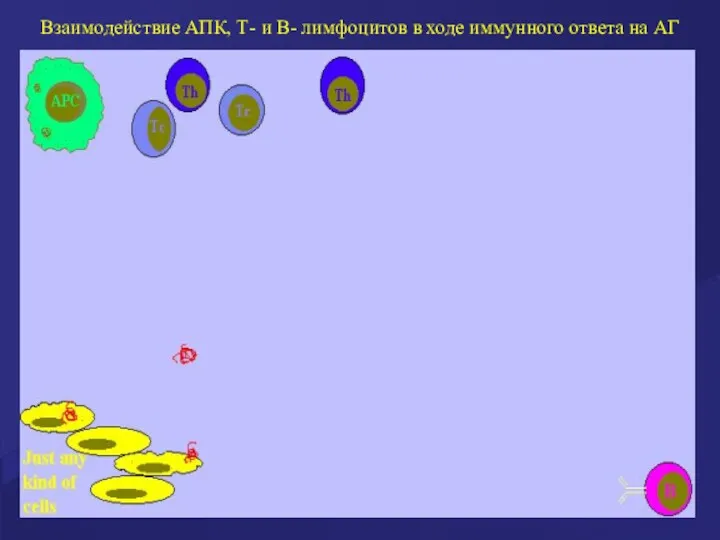
Взаимодействие АПК, Т- и В- лимфоцитов в ходе иммунного ответа на
Взаимодействие АПК, Т- и В- лимфоцитов в ходе иммунного ответа на

Вопросы к занятию № 4
Какова роль тимуса в процессе дифференцировки Т-лимфоцитов?
Каков
Вопросы к занятию № 4
Какова роль тимуса в процессе дифференцировки Т-лимфоцитов?
Каков

Тестовые задания к занятию №4
1. Антигеннезависимая дифференцировка Т лимфоцитов происходит в:
щитовидной
Тестовые задания к занятию №4
1. Антигеннезависимая дифференцировка Т лимфоцитов происходит в:
щитовидной

Тестовые задания к занятию №4
3. Общим маркером Т-лимфоцитов является молекула:
CD 3
CD
Тестовые задания к занятию №4
3. Общим маркером Т-лимфоцитов является молекула:
CD 3
CD

Тестовые задания к занятию №3
5. Для цитотоксических Т-лимфоцитов характерны маркеры:
CD 3
Тестовые задания к занятию №3
5. Для цитотоксических Т-лимфоцитов характерны маркеры:
CD 3

Тестовые задания к занятию № 4
7. Зародышевые гены тяжелых цепей иммуноглобулинов
Тестовые задания к занятию № 4
7. Зародышевые гены тяжелых цепей иммуноглобулинов

Тестовые задания к занятию № 4
9. На поверхности зрелых В-лимфоцитов присутствуют
Тестовые задания к занятию № 4
9. На поверхности зрелых В-лимфоцитов присутствуют
 Гигиеническое обучение и воспитание населения
Гигиеническое обучение и воспитание населения Неопухолевые заболевания толстой кишки
Неопухолевые заболевания толстой кишки Профилактика туберкулеза
Профилактика туберкулеза Внутренняя среда организма. Кровь
Внутренняя среда организма. Кровь Лечебные прокладки
Лечебные прокладки Абдоминальная хирургия
Абдоминальная хирургия Методы контрацепции
Методы контрацепции Соціальний проект Здорове харчування
Соціальний проект Здорове харчування Клиникалық зерттеулерді жүргізу және ұйымдастыру. КЗ этикалық аспектілері
Клиникалық зерттеулерді жүргізу және ұйымдастыру. КЗ этикалық аспектілері Классификация психических расстройств. Подходы к терапии психических заболеваний с точки зрения доказательной медицины
Классификация психических расстройств. Подходы к терапии психических заболеваний с точки зрения доказательной медицины Заболевания и травмы при нерациональных занятиях спортом
Заболевания и травмы при нерациональных занятиях спортом Классический массаж и самомассаж – средство восстановления организма после физической нагрузки
Классический массаж и самомассаж – средство восстановления организма после физической нагрузки Медико-соціальне обґрунтування організації функціонування територіальної ланки державної служби медицини катастроф України
Медико-соціальне обґрунтування організації функціонування територіальної ланки державної служби медицини катастроф України Профилактика осложнений гипертонической болезни
Профилактика осложнений гипертонической болезни Современные принципы лечения и профилактики гриппа и ОРИ
Современные принципы лечения и профилактики гриппа и ОРИ Гормональные средства
Гормональные средства Вирусные дерматозы
Вирусные дерматозы Ультразвуковая диагностика узловых образований щитовидной железы
Ультразвуковая диагностика узловых образований щитовидной железы Индивидуальное здоровье человека, его физическая, духовная и социальная сущность
Индивидуальное здоровье человека, его физическая, духовная и социальная сущность Атипичные нейролептики нового поколения (антипсихотики)
Атипичные нейролептики нового поколения (антипсихотики) Химические реакции с участием макромолекул
Химические реакции с участием макромолекул Сестринский уход за пациентами с пупочными грыжами
Сестринский уход за пациентами с пупочными грыжами Эндогенные и экзогенные канцерогенные факторы. Лекция 9
Эндогенные и экзогенные канцерогенные факторы. Лекция 9 Иммунитет. Вакцинация
Иммунитет. Вакцинация Дифференциальная диагностика первичных злокачественных опухолей костей
Дифференциальная диагностика первичных злокачественных опухолей костей ენურეზის-მართვა-პირველად-ჯანდაცვაში
ენურეზის-მართვა-პირველად-ჯანდაცვაში Изменения в системе кроветворения во время беременности
Изменения в системе кроветворения во время беременности Урологические заболевания
Урологические заболевания