- Кровь и кроветворение

Содержание
- 2. Слайд 1. Цели и задачи занятия На этом занятии мы рассмотрим следующие вопросы: - локализацию кроветворения
- 3. Слайд 2. Кровь — жидкая подвижная соединительная ткань внутренней среды организма, которая состоит из жидкой среды
- 4. Слайд 5 Происхождение форменных элементов крови Имеется теория, которая предполагает существование так называемые стволовой клетки, функционирующей
- 5. Слайд 6
- 6. Слайд 7 Упрощенная схема кроветворения
- 7. Слайд 8 ПОНЯТИЕ ГОМОПОЭЗА Под кроветворением, или гемоп о э з о м (от греч. haima—кровь
- 8. Слайд 9
- 9. Слайд 11
- 10. Слайд 14 Эмбриональный период кроветворения У зародыша кроветворение начинается в желточном мешке. Со 2-го месяца эту
- 11. Слайд 15 Эмбриональное кроветворение В эмбриональном периоде кроветворение происходит вначале в желточном мешке, а затем последовательно
- 12. Слайд 16 Желточный этап кроветворения Желточный этап осуществляется в мезенхиме желточного мешка, начиная со 2—3-ей недели
- 13. Слайд 17 Желточный этап кроветворения По данным некоторых исследователей желточный мешок является источником полипотентных стволовых кроветворных
- 14. Слайд 19 Гепато-тимусо-лиенальный этап гемопоэза Этот этап осуществляется в начале в печени, несколько позже в тимусе
- 15. Слайд 21
- 16. Слайд 22
- 17. Слайд 25
- 18. Слайд 25 (а)
- 19. Слайд 26 Роль лимфатических узлов в кроветворении Закладка лимфатических узлов у эмбриона человека происходит не одновременно.
- 20. Слайд 27
- 21. Слайд 28 Медулло-тимусо-лимфоидный этап гемопоэза Закладка красного костного мозга начинается со 2-го месяца, кроветворение в нем
- 22. Слайд 29 Т-лимфоцитопоэз В отличие от миелопоэза, лимфоцитопоэз в эмбриональном и постэмбриональном периодах осуществляется поэтапно, сменяя
- 23. Слайд 30 Этапы Т-лимфоцитопоэза Первый этап Т-лимфоцитопоэза осуществляется в лимфоидной ткани красного костного мозга, где образуются
- 24. Слайд 32 Обеспечение клеточного иммунитета Т-лимфацитами В обеспечении клеточного иммунитета рассматривают два механизма уничтожения киллерами антигенных
- 25. Слайд 33 В-лимфоцитопоэз Первый этап В-лимфоцитопоэза осуществляется в красном костном мозге, где образуются следующие классы клеток:
- 26. Слайд 34 В-лимфоцитопоэз Влияние антигенного стимула на В-лимфоцит недостаточно для его бласттрансформации. Это происходит только после
- 27. Слайд 35 ВНЕЭМБРИОНАЛЬНОЕ КРОВЕТВОРЕНИЕ В неэмбриональное кроветворение начинается в самый ранний период развития зародыша, одновременно с
- 28. Слайд 36 ПОСТЭМБРИОНАЛЬНОЕ КРОВЕТВОРЕНИЕ В постэмбриональном периоде кроветворение осуществляется главным образом в костном мозге, селезенке, лимфатических
- 29. Слайд 37 Схема – кроветворные органы у человека
- 30. Слайд 39
- 31. Слайд 40
- 32. Слайд 41 Схема кроветворения Согласно современным представлениям, в кроветворной ткани помимо морфологически распознаваемых клеток есть клетки-предшественницы
- 33. Слайд 42
- 34. Слайд 43
- 35. Слайд 44 выделяют два вида кроветворения: миелопоэз - образование всех форменных элементов крови, кроме лимфоцитов, т.е.
- 36. Слайд 48 Фрагмент общей схемы
- 37. Слайд 49 Общие свойства клеток классов I-III Локализация: Данные клетки находятся, в основном, в красном костном
- 38. Слайд 50 Особенности клеток классов I, II и III Класс I: стволовые клетки крови. Эти клетки
- 39. Слайд 51 Виды полустволовых клеток Из вышеприведённых схем следует, что к полустволовым клеткам относятся предшественники миелопоэза
- 40. Слайд 52 Потенции развития полустволовых КоЕ В обозначениях полустволовых КоЕ буквы после чёрточки показывают, в какие
- 41. Слайд 53 Регуляторы миелопоэза Превращение предшественников миелопоэза в тот или иной из трёх перечисленных видов КоЕ
- 42. Слайд 55 Гемопоэтические клетки класса IV 20.2.3.1. Фрагмент общей схемы
- 43. Слайд 56 Свойства клеток Деления и созревание 8 видов клеток класса III приводят к образованию бластов
- 44. Слайд 57 Завершающие стадии миелопоэза Общая характеристика Множественность промежуточных форм. Класс V гемопоэтических клеток почти в
- 45. Слайд 58 Стволовые клетки в кроветворных органах Стволове, клетки способными к самоподдержанию, и к дифференцировкам по
- 46. Слайд 60
- 47. Слайд 61 поэтиночувствительные клетки-предшественницы После того как было доказано существование стволовых клеток с помощью метода культуры
- 48. Слайд 62 Лейкопоэз Лейкопоэз (лат. leucopoesis, leucopoiesis); лейко- + греч. poiesis выработка, образование; синонимум - лейкогенез,
- 49. Слайд 63
- 50. Слайд 64
- 51. Слайд 66 Гранулоцитопоэз. Гранулоцитопоэз – это образование гранулоцитов Важнейшие функции гранулоцитов - способность к фагоцитозу и
- 52. Слайд 67
- 53. Слайд 68
- 54. Слайд 69 Гранулоцитопоэз. На стадии поздних миелобластов и промиелоцитов происходит образование первичных гранул (азурофильной зернистости), специфическим
- 55. Слайд 70 Гранулоцитопоэз.
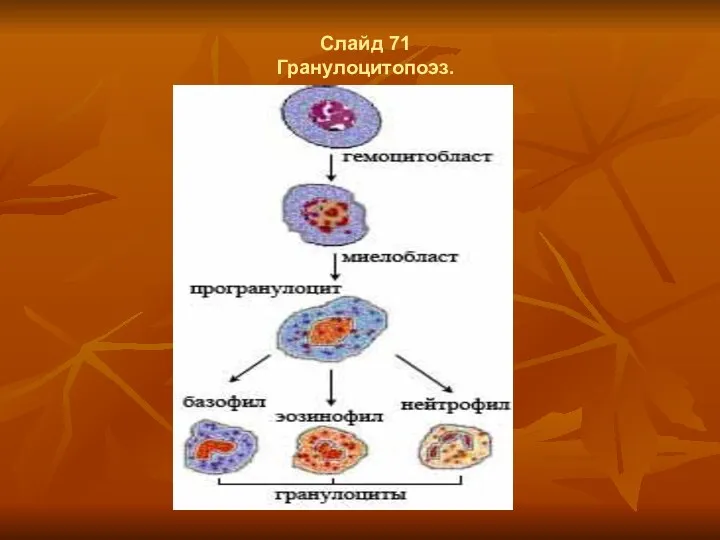
- 56. Слайд 71 Гранулоцитопоэз.
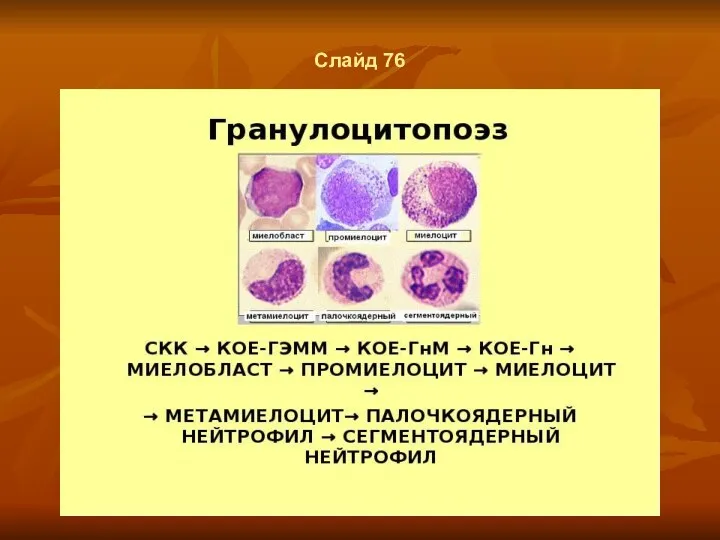
- 57. Слайд 76
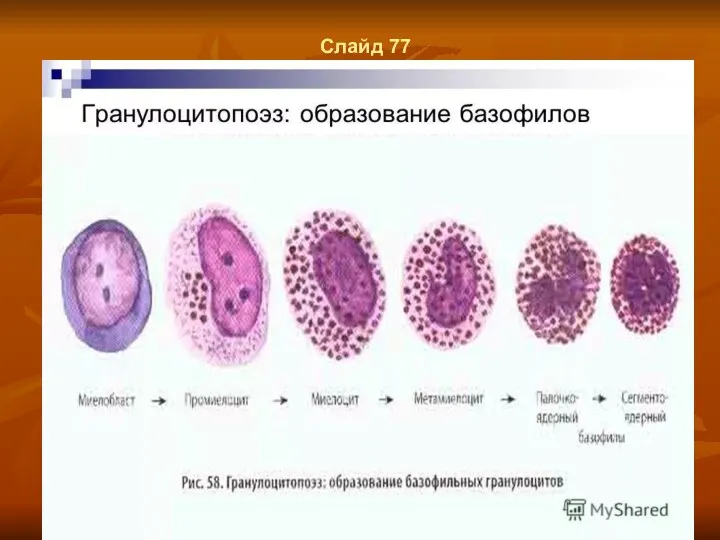
- 58. Слайд 77
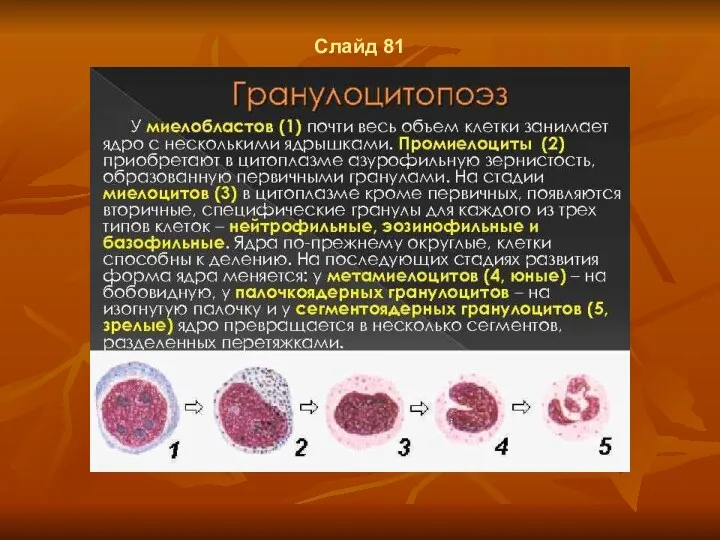
- 59. Слайд 81
- 60. Слайд 82
- 61. Слайд 83
- 62. Слайд 85 Нейтрофилы Нейтрофилы составляют 60-70% общего числа лейкоцитов крови. После выхода нейтрофильных гранулоцитов из костного
- 63. Слайд 86
- 64. Слайд 88
- 65. Слайд 90 Эозинофилы Эозинофилы составляют 0.5-5% от всех лейкоцитов крови, циркулируют в течение 6-12 часов, после
- 66. Слайд 91 Базофилы Базофилы и тучные клетки имеют костномозговое происхождение. Предполагают, что предшественники тучных клеток покидают
- 67. Слайд 92 Моноциты и макрофаги Моноциты и макрофаги являются основными клетками системы мононуклеарных фагоцитов (СМФ) или
- 68. Слайд 93
- 69. Слайд 95 МАКРОФАГИ Зрелые макрофаги имеют ряд общих морфологических признаков: значительные размеры (диаметр от 20-25 до
- 70. Слайд 96 Лейкопоэз Лейкопоэз начинается в костном мозге со стволовой клетки (I класс), которая способна к
- 71. Слайд 97 Лейкопоэз Клетки I, II и III класса морфологически недифференцируемы, выглядят как малые темные лимфоциты
- 72. Слайд 98 Лимфоцитопоэз Лимфоцитопоэз или лимфопоэз (lymphopoesis, lymptiopoiesis, лимфо- + греч. poiesis — выработка, образование) или
- 73. Слайд 99 Лимфоцитопоэз Существенным в представлении о лимфоцитопоэзе явилось открытие двух типов лимфоцитов — В- и
- 74. Слайд 100 Лейкоцитопоэз Лейкоцитопоэз (лейкопоэз) включает гранулоцитопоэз (гранулопоэз), лимфоцитопоэз (лимфопоэз) и моноцитопоэз (монопоэз). В гранулоцитарном ряду
- 75. Слайд 102
- 76. Слайд 103
- 77. Слайд 104 Лимфоцитопоэз Поскольку переход от промиелоцита к следующей стадии созревания клеток — миелоциту — не
- 78. Слайд 105
- 79. Слайд 106
- 80. Слайд 107
- 81. Слайд 108
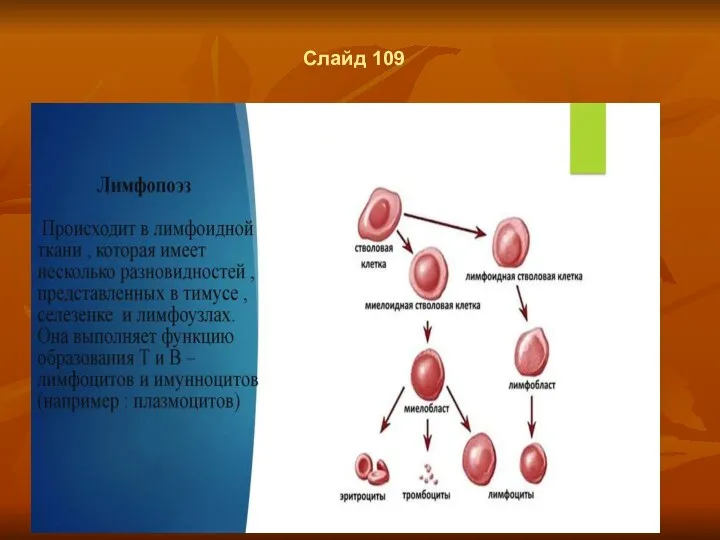
- 82. Слайд 109
- 83. Слайд 110
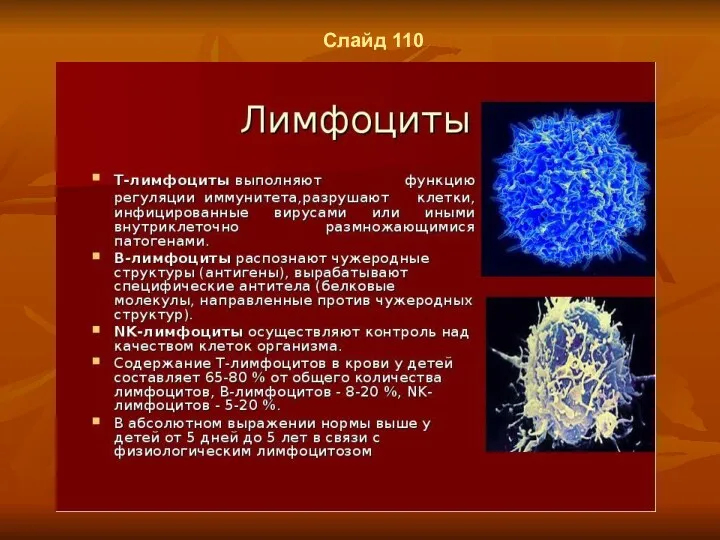
- 84. Слайд 111
- 85. Слайд 112
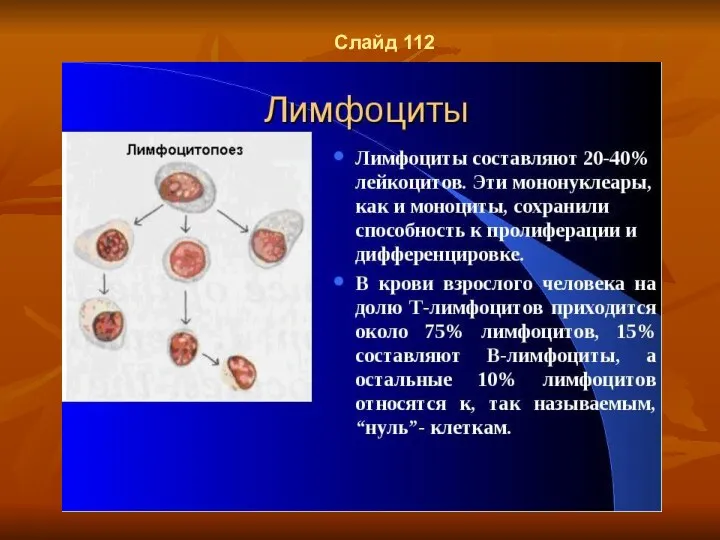
- 86. Слайд 113
- 87. Слайд 114
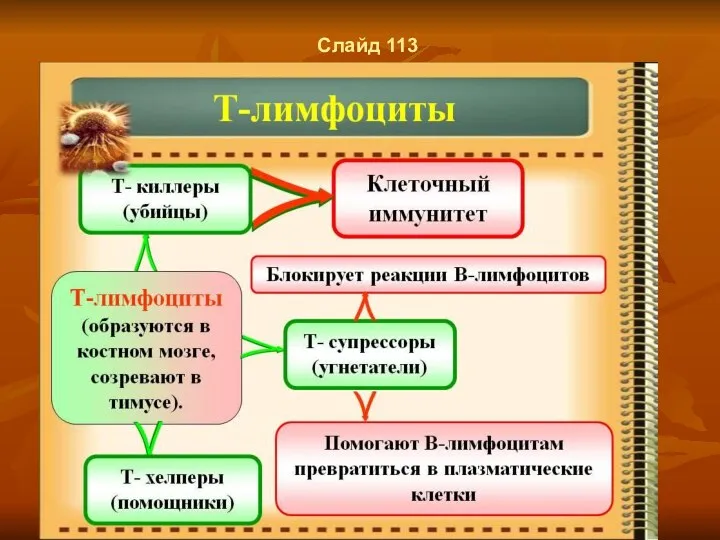
- 88. Эритропоэз Слайд 116 Клетки класса V (созревающие клетки) (Эритробласт, IV) а) Проэритробласт. 1. На стадии проэритробласта
- 89. Слайд 117 Клетки класса VI (зрелые клетки) Ретикулоциты. 1. На стадии ретикулоцита клетка у млекопитающих уже
- 90. Слайд 118
- 91. Слайд 121
- 92. Слайд 123
- 93. Слайд 124
- 94. Слайд 125
- 95. Слайд 128 Эритропоэз В норме эритробласты развиваются в костном мозгу преимущественно гомопластическим способом, т. е. путем
- 96. Слайд 129 Эритропоэз В эритроцитопоэзе (эритропоэзе) самой молодой клеткой является эритробласт (ее называют также проэритробластом), который
- 97. Слайд 134 Норма эритроцитов в крови норме содержание красных телец в крови у взрослых мужчин составляет
- 98. Слайд 135 Гранулоцитопоэз I. Фрагмент общей схемы
- 99. Слайд 136 Промиелоциты - первые клетки класса V 1. Промиелоциты (нейтрофильные, эозинофильные и базофильные) ещё друг
- 100. Слайд 137 Ядра, их последующие изменения и способность к делениям. 1. а) Ядра у миелоцитов по-прежнему
- 101. Слайд 138 Клетки завершающих стадий развития Метамиелоциты: нейтрофильные, эозинофильные, базофильные. Палочкоядерные гранулоциты: нейтрофильные эозинофильные, базофильные. Сегментоядерные
- 102. Слайд 139 Моноцитопоэз Х а р а к т е р и с т и к
- 103. Слайд 140 Тромбоцитопоэз Фрагмент общей схемы: Мегакариобласты, Промегакариоцит, Мегакариоцит. Характеристика клеток. 1. При переходе от мегакариобласта
- 104. Слайд 141 Зрелые форменные элементы класса VI 1. а) Мегакариоцит "проталкивает" часть своей цитоплазмы(в виде отростков)
- 105. Слайд 144
- 106. Слайд 146
- 107. Слайд 148 Норма тромбоцитов Количество кровяных пластинок измеряется в тысячах на 1 микролитр крови. Для мужчин
- 108. Слайд 149 Отношения между кроветворными органами и кровью Морфологически состав крови зависит от двух факторов: от
- 109. Слайд 150 Отношения между кроветворными органами и кровью При лейкозах явления гиперплазии кроветворных органов в большинстве
- 110. Слайд 164 Лейкоциты Лейкоциты, или «белые кровяные тельца» (Л, White Blood Cells, WBC) — это гетерогенная
- 111. Слайд 165 Лейкоциты Функции: 1)защитная(фагоцитоз микробов, бактерицидное и антитоксическое действие, участие в иммунных реакциях, в процессе
- 112. Слайд 166 Отличие гранулоцитов от агранулоцитов. Агранулоциты - это одноядерные лейкоциты, один из двух типов лейкоцитов.
- 113. Слайд 167 Общие свойства для всех лецкоциов, связанные с функцией: Способность к хемотаксису (сомат.движение) в тканях
- 114. Слайд 168 Защитная функция лейкоцитов Главная функция лейкоцитов - защита. Они играют главную роль в специфической
- 115. Слайд 169 Очистка. Регенерация лейкоцитов Макрофаги пожирая остатки погибших клеток, очищают место битвы. Таким образом, создаются
- 116. Слайд 191 Адгезия Вовлечение тромбоцитов в процесс гемостаза невозможно без их адгезии к поверхности. Повреждение сосуда
- 117. Слайд 195
- 119. Скачать презентацию

Слайд 1.
Цели и задачи занятия
На этом занятии мы рассмотрим следующие вопросы:
-
Слайд 1.
Цели и задачи занятия
На этом занятии мы рассмотрим следующие вопросы:
-

Слайд 2.
Кровь — жидкая подвижная соединительная ткань внутренней среды организма, которая
Слайд 2.
Кровь — жидкая подвижная соединительная ткань внутренней среды организма, которая

Слайд 5
Происхождение форменных элементов крови
Имеется теория, которая предполагает существование так называемые
Слайд 5
Происхождение форменных элементов крови
Имеется теория, которая предполагает существование так называемые
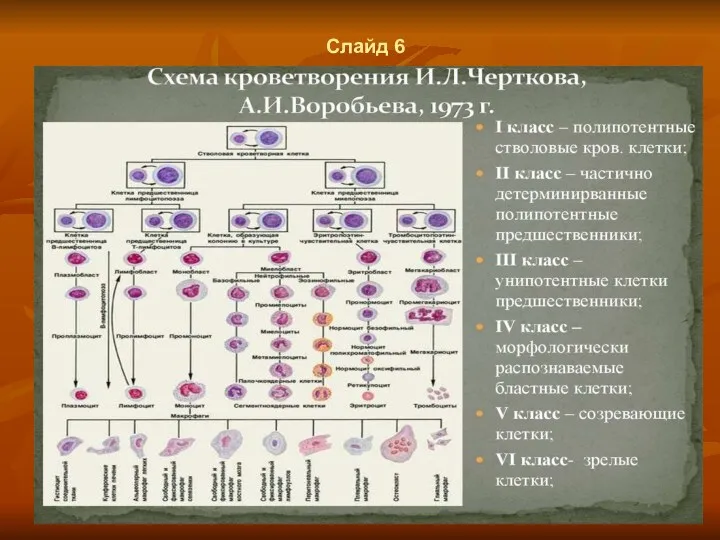
Слайд 6
Слайд 6

Слайд 7
Упрощенная схема кроветворения
Слайд 7
Упрощенная схема кроветворения

Слайд 8
ПОНЯТИЕ ГОМОПОЭЗА
Под кроветворением, или гемоп о э з о м
Слайд 8
ПОНЯТИЕ ГОМОПОЭЗА
Под кроветворением, или гемоп о э з о м
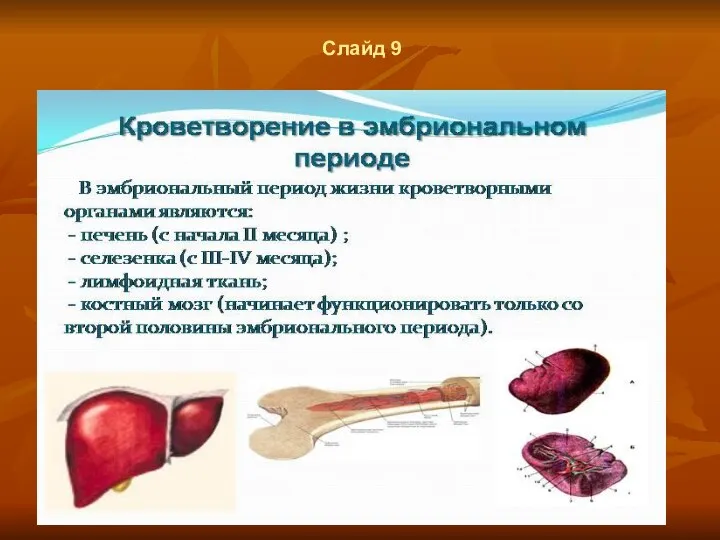
Слайд 9
Слайд 9

Слайд 11
Слайд 11

Слайд 14
Эмбриональный период кроветворения
У зародыша кроветворение начинается в желточном мешке.
Слайд 14
Эмбриональный период кроветворения
У зародыша кроветворение начинается в желточном мешке.

Слайд 15
Эмбриональное кроветворение
В эмбриональном периоде кроветворение происходит вначале в желточном мешке,
Слайд 15
Эмбриональное кроветворение
В эмбриональном периоде кроветворение происходит вначале в желточном мешке,

Слайд 16
Желточный этап кроветворения
Желточный этап осуществляется в мезенхиме желточного мешка, начиная
Слайд 16
Желточный этап кроветворения
Желточный этап осуществляется в мезенхиме желточного мешка, начиная

Слайд 17
Желточный этап кроветворения
По данным некоторых исследователей желточный мешок является источником
Слайд 17
Желточный этап кроветворения
По данным некоторых исследователей желточный мешок является источником

Слайд 19
Гепато-тимусо-лиенальный этап гемопоэза
Этот этап осуществляется в начале в печени, несколько
Слайд 19
Гепато-тимусо-лиенальный этап гемопоэза
Этот этап осуществляется в начале в печени, несколько

Слайд 21
Слайд 21

Слайд 22
Слайд 22

Слайд 25
Слайд 25

Слайд 25 (а)
Слайд 25 (а)

Слайд 26
Роль лимфатических узлов в кроветворении
Закладка лимфатических узлов у эмбриона человека
Слайд 26
Роль лимфатических узлов в кроветворении
Закладка лимфатических узлов у эмбриона человека

Слайд 27
Слайд 27

Слайд 28
Медулло-тимусо-лимфоидный этап гемопоэза
Закладка красного костного мозга начинается со 2-го месяца,
Слайд 28
Медулло-тимусо-лимфоидный этап гемопоэза
Закладка красного костного мозга начинается со 2-го месяца,

Слайд 29
Т-лимфоцитопоэз
В отличие от миелопоэза, лимфоцитопоэз в эмбриональном и постэмбриональном
Слайд 29
Т-лимфоцитопоэз
В отличие от миелопоэза, лимфоцитопоэз в эмбриональном и постэмбриональном

Слайд 30
Этапы Т-лимфоцитопоэза
Первый этап Т-лимфоцитопоэза осуществляется в лимфоидной ткани красного костного
Слайд 30
Этапы Т-лимфоцитопоэза
Первый этап Т-лимфоцитопоэза осуществляется в лимфоидной ткани красного костного

Слайд 32
Обеспечение клеточного иммунитета Т-лимфацитами
В обеспечении клеточного иммунитета рассматривают два механизма
Слайд 32
Обеспечение клеточного иммунитета Т-лимфацитами
В обеспечении клеточного иммунитета рассматривают два механизма

Слайд 33
В-лимфоцитопоэз
Первый этап В-лимфоцитопоэза осуществляется в красном костном мозге, где
Слайд 33
В-лимфоцитопоэз
Первый этап В-лимфоцитопоэза осуществляется в красном костном мозге, где

Слайд 34
В-лимфоцитопоэз
Влияние антигенного стимула на В-лимфоцит недостаточно для его бласттрансформации. Это
Слайд 34
В-лимфоцитопоэз
Влияние антигенного стимула на В-лимфоцит недостаточно для его бласттрансформации. Это

Слайд 35
ВНЕЭМБРИОНАЛЬНОЕ КРОВЕТВОРЕНИЕ
В неэмбриональное кроветворение начинается в самый ранний период развития
Слайд 35
ВНЕЭМБРИОНАЛЬНОЕ КРОВЕТВОРЕНИЕ
В неэмбриональное кроветворение начинается в самый ранний период развития

Слайд 36
ПОСТЭМБРИОНАЛЬНОЕ КРОВЕТВОРЕНИЕ
В постэмбриональном периоде кроветворение осуществляется главным образом в костном
Слайд 36
ПОСТЭМБРИОНАЛЬНОЕ КРОВЕТВОРЕНИЕ
В постэмбриональном периоде кроветворение осуществляется главным образом в костном
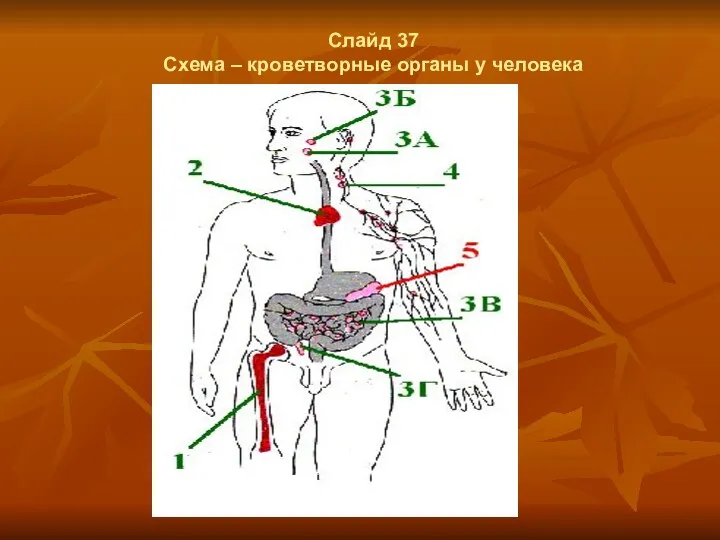
Слайд 37
Схема – кроветворные органы у человека
Слайд 37
Схема – кроветворные органы у человека
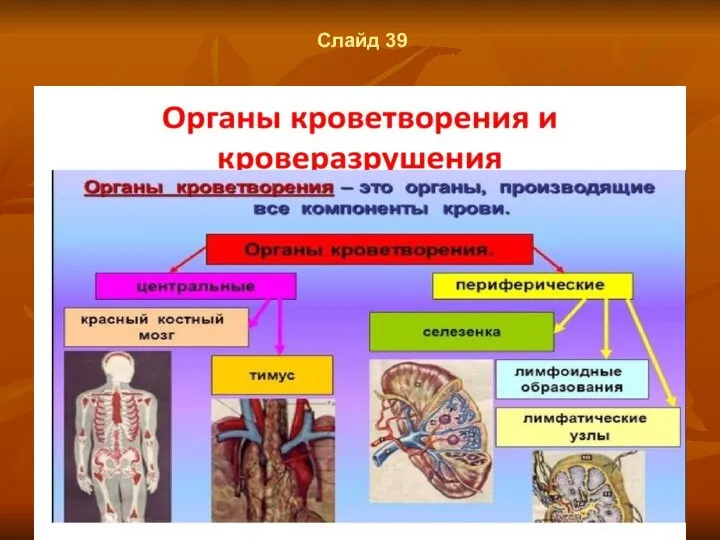
Слайд 39
Слайд 39
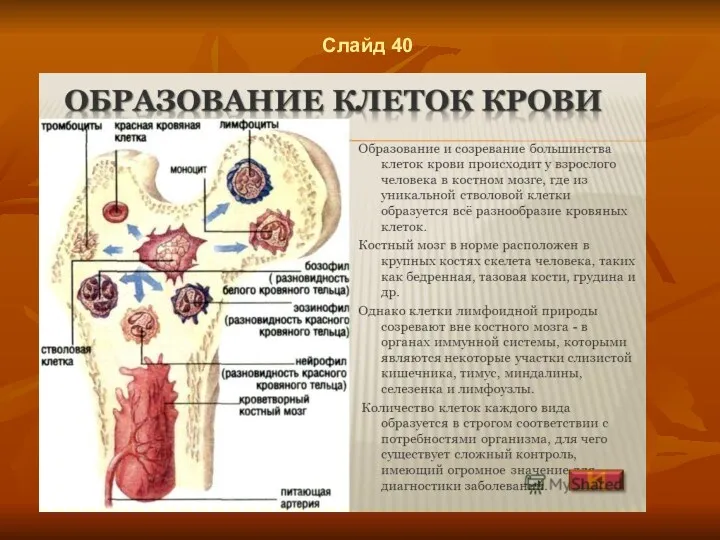
Слайд 40
Слайд 40

Слайд 41
Схема кроветворения
Согласно современным представлениям, в кроветворной ткани помимо морфологически распознаваемых
Слайд 41
Схема кроветворения
Согласно современным представлениям, в кроветворной ткани помимо морфологически распознаваемых
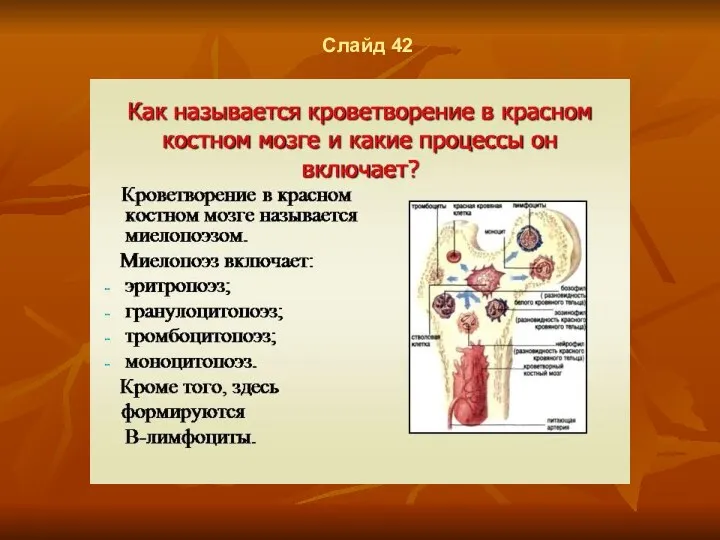
Слайд 42
Слайд 42

Слайд 43
Слайд 43

Слайд 44
выделяют два вида кроветворения:
миелопоэз - образование всех форменных элементов крови, кроме
Слайд 44
выделяют два вида кроветворения:
миелопоэз - образование всех форменных элементов крови, кроме
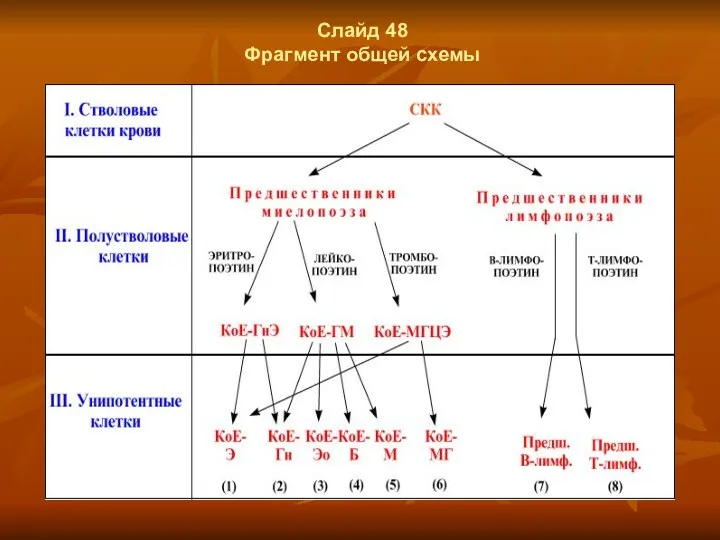
Слайд 48
Фрагмент общей схемы
Слайд 48
Фрагмент общей схемы

Слайд 49
Общие свойства клеток классов I-III
Локализация: Данные клетки находятся, в основном, в
Слайд 49
Общие свойства клеток классов I-III
Локализация: Данные клетки находятся, в основном, в

Слайд 50
Особенности клеток классов I, II и III
Класс I: стволовые клетки
Слайд 50
Особенности клеток классов I, II и III
Класс I: стволовые клетки

Слайд 51
Виды полустволовых клеток
Из вышеприведённых схем следует, что к полустволовым клеткам
Слайд 51
Виды полустволовых клеток
Из вышеприведённых схем следует, что к полустволовым клеткам

Слайд 52
Потенции развития полустволовых КоЕ
В обозначениях полустволовых КоЕ буквы после чёрточки
Слайд 52
Потенции развития полустволовых КоЕ
В обозначениях полустволовых КоЕ буквы после чёрточки

Слайд 53
Регуляторы миелопоэза
Превращение предшественников миелопоэза в тот или иной из трёх
Слайд 53
Регуляторы миелопоэза
Превращение предшественников миелопоэза в тот или иной из трёх
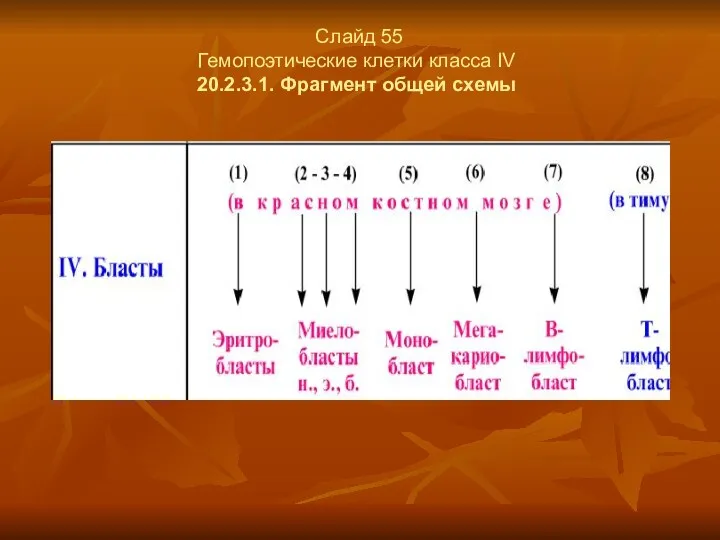
Слайд 55
Гемопоэтические клетки класса IV
20.2.3.1. Фрагмент общей схемы
Слайд 55
Гемопоэтические клетки класса IV
20.2.3.1. Фрагмент общей схемы

Слайд 56
Свойства клеток
Деления и созревание 8 видов клеток класса III приводят
Слайд 56
Свойства клеток
Деления и созревание 8 видов клеток класса III приводят

Слайд 57
Завершающие стадии миелопоэза
Общая характеристика
Множественность промежуточных форм. Класс V гемопоэтических клеток
Слайд 57
Завершающие стадии миелопоэза
Общая характеристика
Множественность промежуточных форм. Класс V гемопоэтических клеток

Слайд 58
Стволовые клетки в кроветворных органах
Стволове, клетки способными к самоподдержанию, и
Слайд 58
Стволовые клетки в кроветворных органах
Стволове, клетки способными к самоподдержанию, и
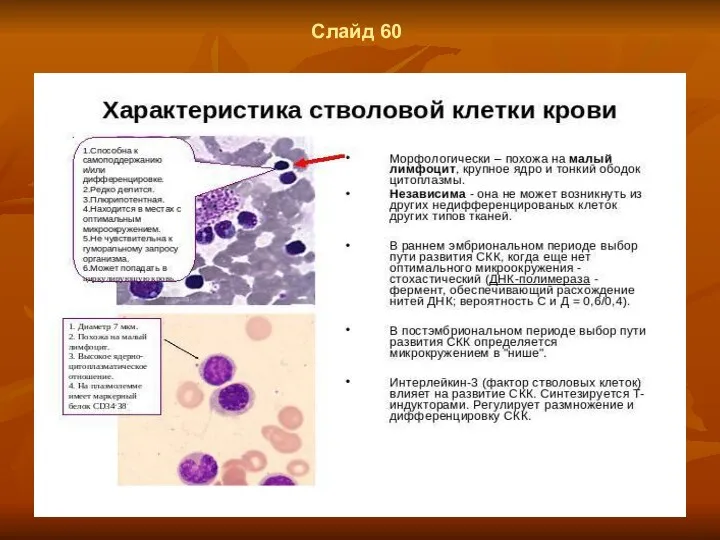
Слайд 60
Слайд 60

Слайд 61
поэтиночувствительные клетки-предшественницы
После того как было доказано существование стволовых клеток с
Слайд 61
поэтиночувствительные клетки-предшественницы
После того как было доказано существование стволовых клеток с

Слайд 62
Лейкопоэз
Лейкопоэз (лат. leucopoesis, leucopoiesis); лейко- + греч. poiesis выработка, образование;
Слайд 62
Лейкопоэз
Лейкопоэз (лат. leucopoesis, leucopoiesis); лейко- + греч. poiesis выработка, образование;
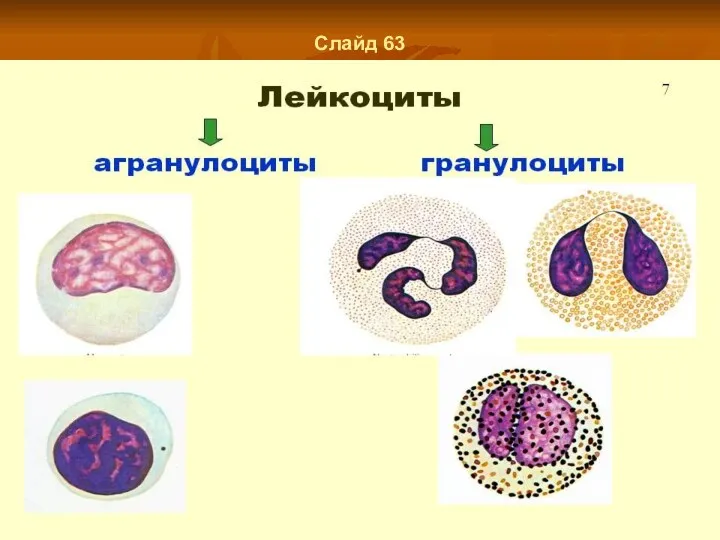
Слайд 63
Слайд 63

Слайд 64
Слайд 64

Слайд 66
Гранулоцитопоэз.
Гранулоцитопоэз – это образование гранулоцитов
Важнейшие функции гранулоцитов - способность
Слайд 66
Гранулоцитопоэз.
Гранулоцитопоэз – это образование гранулоцитов
Важнейшие функции гранулоцитов - способность

Слайд 67
Слайд 67
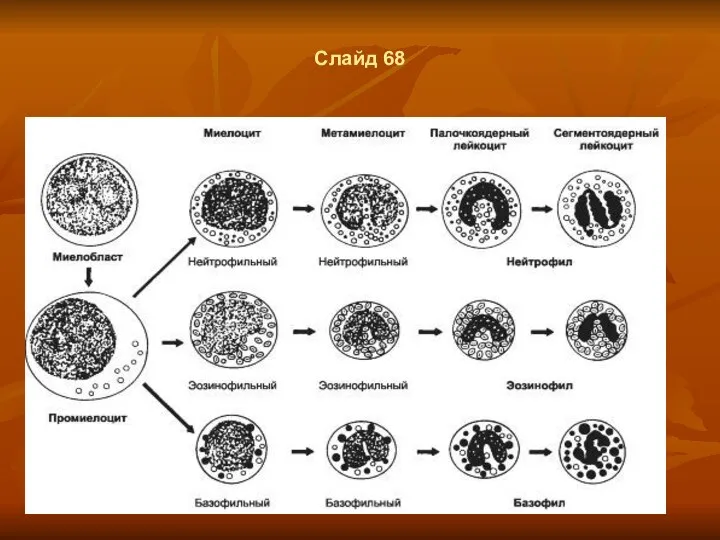
Слайд 68
Слайд 68

Слайд 69
Гранулоцитопоэз.
На стадии поздних миелобластов и промиелоцитов происходит образование первичных
Слайд 69
Гранулоцитопоэз.
На стадии поздних миелобластов и промиелоцитов происходит образование первичных
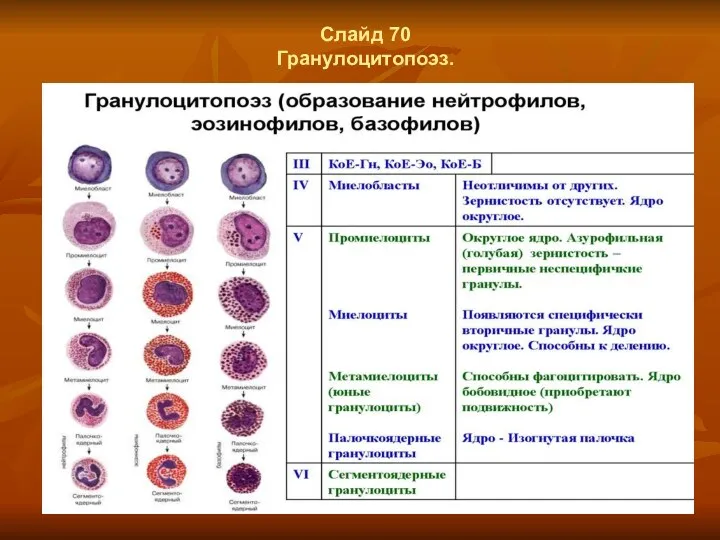
Слайд 70
Гранулоцитопоэз.
Слайд 70
Гранулоцитопоэз.
Слайд 71
Гранулоцитопоэз.
Слайд 71
Гранулоцитопоэз.
Слайд 76
Слайд 76
Слайд 77
Слайд 77
Слайд 81
Слайд 81
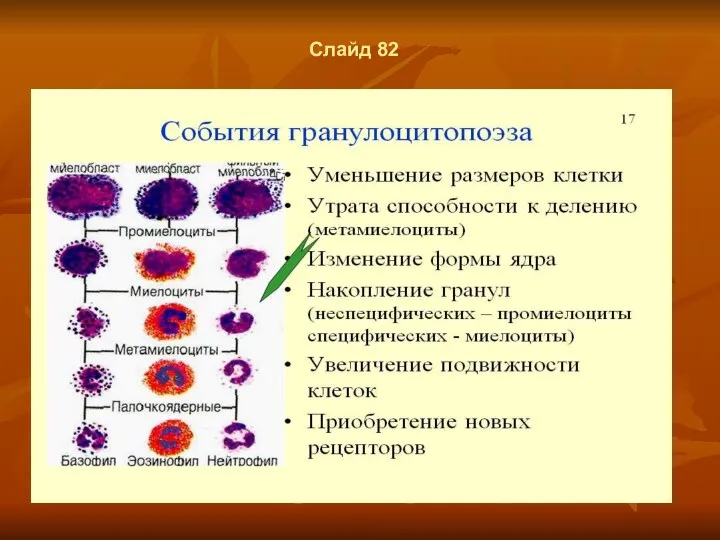
Слайд 82
Слайд 82
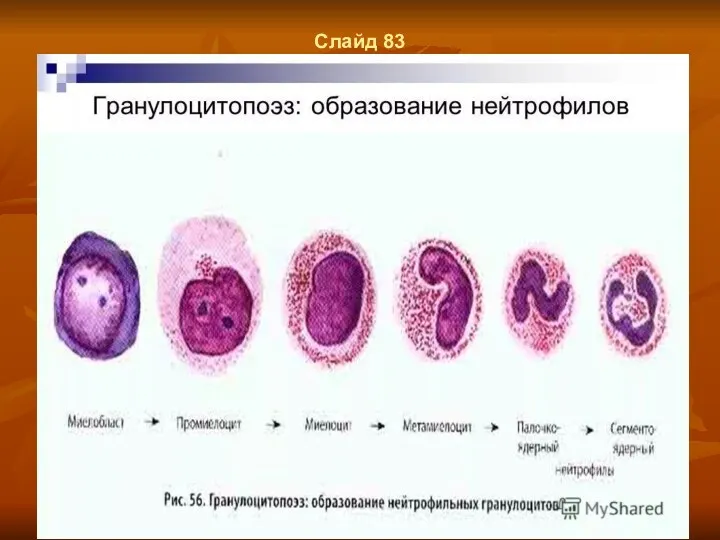
Слайд 83
Слайд 83

Слайд 85
Нейтрофилы
Нейтрофилы составляют 60-70% общего числа лейкоцитов крови. После выхода
Слайд 85
Нейтрофилы
Нейтрофилы составляют 60-70% общего числа лейкоцитов крови. После выхода
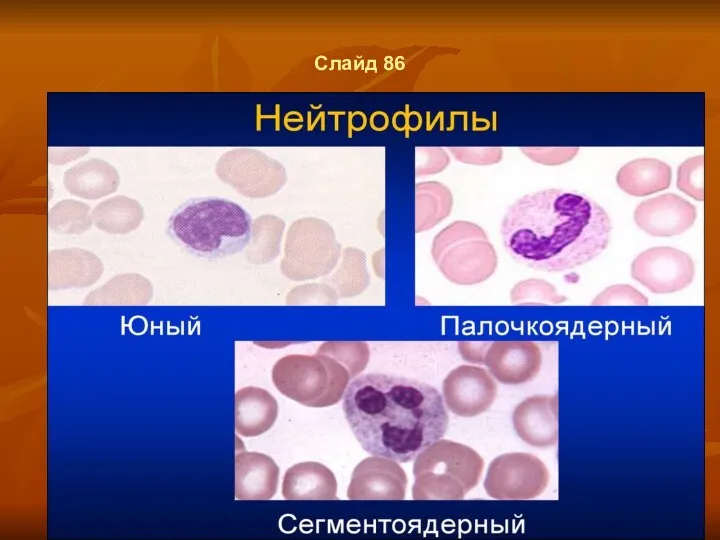
Слайд 86
Слайд 86
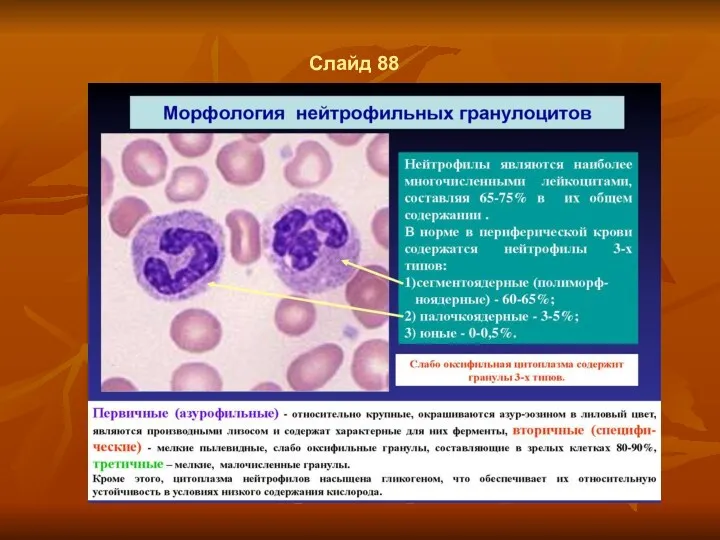
Слайд 88
Слайд 88

Слайд 90
Эозинофилы
Эозинофилы составляют 0.5-5% от всех лейкоцитов крови, циркулируют в течение
Слайд 90
Эозинофилы
Эозинофилы составляют 0.5-5% от всех лейкоцитов крови, циркулируют в течение

Слайд 91
Базофилы
Базофилы и тучные клетки имеют костномозговое происхождение. Предполагают, что предшественники
Слайд 91
Базофилы
Базофилы и тучные клетки имеют костномозговое происхождение. Предполагают, что предшественники

Слайд 92
Моноциты и макрофаги
Моноциты и макрофаги являются основными клетками системы мононуклеарных
Слайд 92
Моноциты и макрофаги
Моноциты и макрофаги являются основными клетками системы мононуклеарных
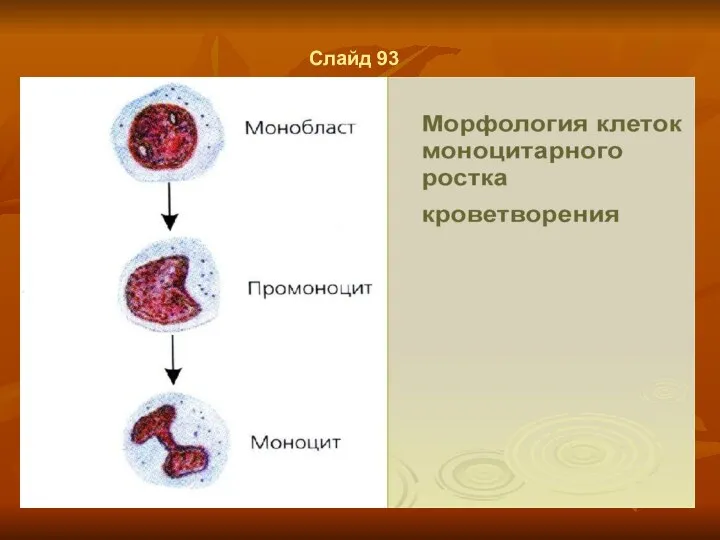
Слайд 93
Слайд 93

Слайд 95
МАКРОФАГИ
Зрелые макрофаги имеют ряд общих морфологических признаков: значительные размеры (диаметр
Слайд 95
МАКРОФАГИ
Зрелые макрофаги имеют ряд общих морфологических признаков: значительные размеры (диаметр

Слайд 96
Лейкопоэз
Лейкопоэз начинается в костном мозге со стволовой клетки (I класс),
Слайд 96
Лейкопоэз
Лейкопоэз начинается в костном мозге со стволовой клетки (I класс),

Слайд 97
Лейкопоэз
Клетки I, II и III класса морфологически недифференцируемы, выглядят как
Слайд 97
Лейкопоэз
Клетки I, II и III класса морфологически недифференцируемы, выглядят как

Слайд 98
Лимфоцитопоэз
Лимфоцитопоэз или лимфопоэз (lymphopoesis, lymptiopoiesis, лимфо- + греч. poiesis —
Слайд 98
Лимфоцитопоэз
Лимфоцитопоэз или лимфопоэз (lymphopoesis, lymptiopoiesis, лимфо- + греч. poiesis —

Слайд 99
Лимфоцитопоэз
Существенным в представлении о лимфоцитопоэзе явилось открытие двух типов лимфоцитов
Слайд 99
Лимфоцитопоэз
Существенным в представлении о лимфоцитопоэзе явилось открытие двух типов лимфоцитов

Слайд 100
Лейкоцитопоэз
Лейкоцитопоэз (лейкопоэз) включает гранулоцитопоэз (гранулопоэз), лимфоцитопоэз (лимфопоэз) и моноцитопоэз (монопоэз).
В
Слайд 100
Лейкоцитопоэз
Лейкоцитопоэз (лейкопоэз) включает гранулоцитопоэз (гранулопоэз), лимфоцитопоэз (лимфопоэз) и моноцитопоэз (монопоэз).
В
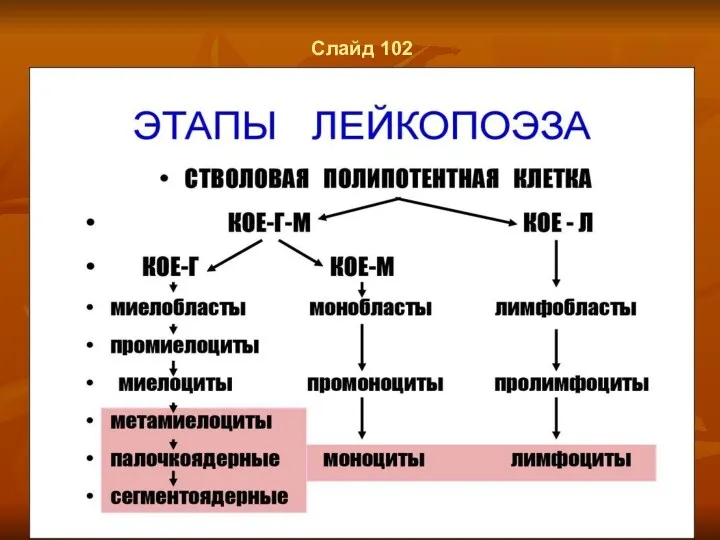
Слайд 102
Слайд 102
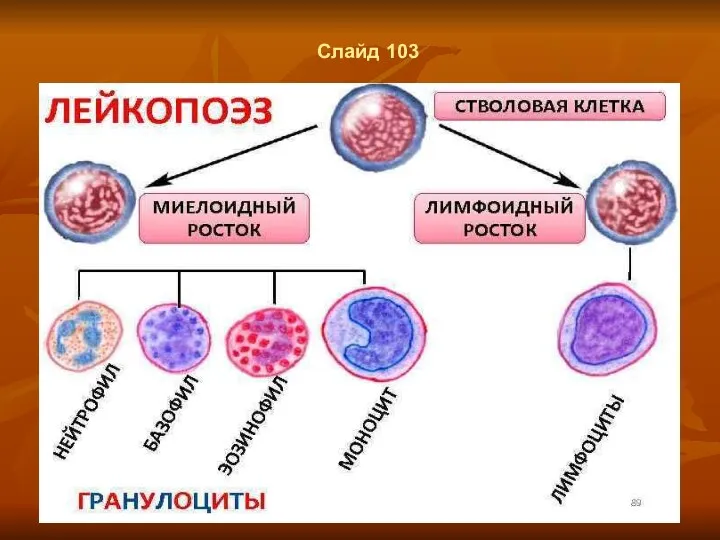
Слайд 103
Слайд 103

Слайд 104
Лимфоцитопоэз
Поскольку переход от промиелоцита к следующей стадии созревания клеток —
Слайд 104
Лимфоцитопоэз
Поскольку переход от промиелоцита к следующей стадии созревания клеток —

Слайд 105
Слайд 105
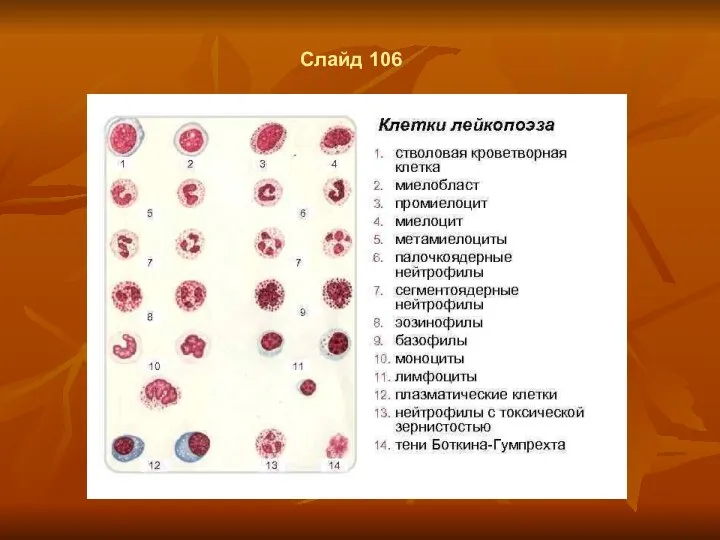
Слайд 106
Слайд 106

Слайд 107
Слайд 107

Слайд 108
Слайд 108
Слайд 109
Слайд 109
Слайд 110
Слайд 110

Слайд 111
Слайд 111
Слайд 112
Слайд 112
Слайд 113
Слайд 113
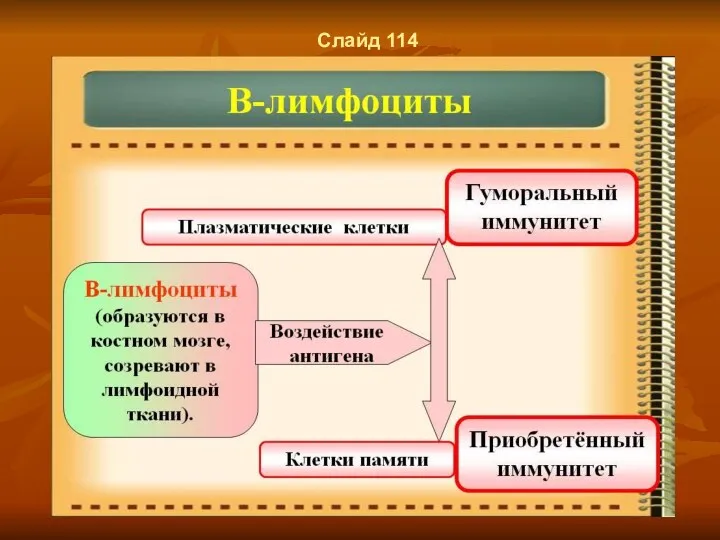
Слайд 114
Слайд 114

Эритропоэз
Слайд 116
Клетки класса V (созревающие клетки)
(Эритробласт, IV) а) Проэритробласт.
1.
Эритропоэз
Слайд 116
Клетки класса V (созревающие клетки)
(Эритробласт, IV) а) Проэритробласт.
1.

Слайд 117
Клетки класса VI (зрелые клетки)
Ретикулоциты. 1. На стадии ретикулоцита клетка
Слайд 117
Клетки класса VI (зрелые клетки)
Ретикулоциты. 1. На стадии ретикулоцита клетка

Слайд 118
Слайд 118

Слайд 121
Слайд 121
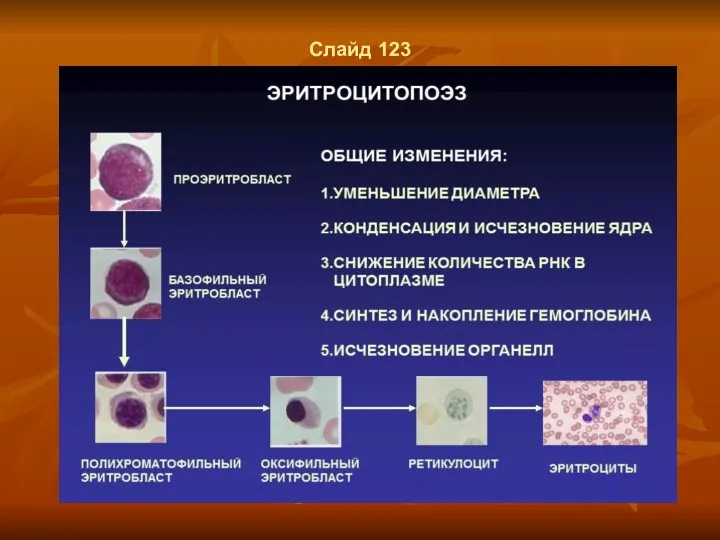
Слайд 123
Слайд 123
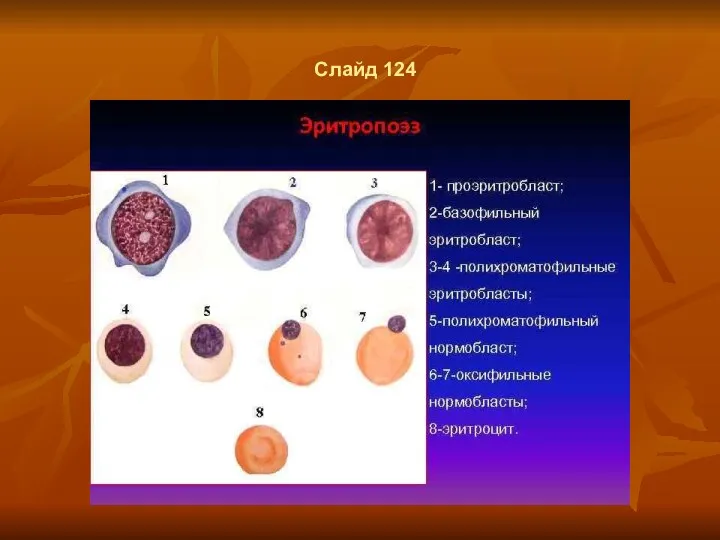
Слайд 124
Слайд 124
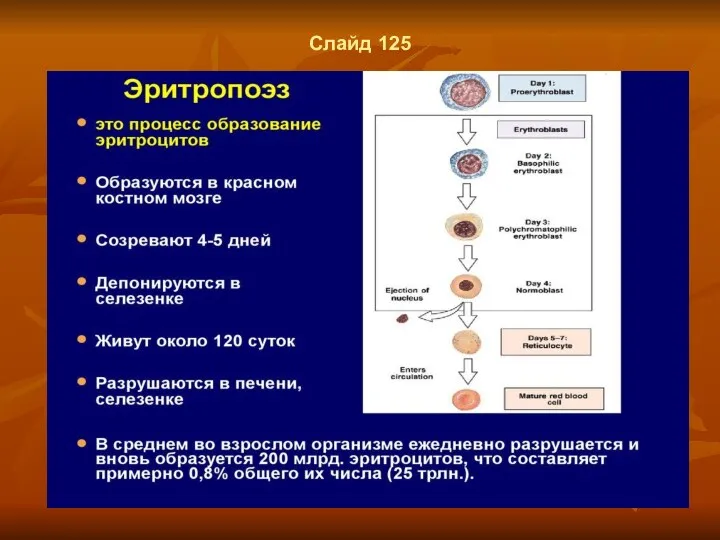
Слайд 125
Слайд 125

Слайд 128
Эритропоэз
В норме эритробласты развиваются в костном мозгу преимущественно
Слайд 128
Эритропоэз
В норме эритробласты развиваются в костном мозгу преимущественно

Слайд 129
Эритропоэз
В эритроцитопоэзе (эритропоэзе) самой молодой клеткой является эритробласт (ее
Слайд 129
Эритропоэз
В эритроцитопоэзе (эритропоэзе) самой молодой клеткой является эритробласт (ее

Слайд 134
Норма эритроцитов в крови
норме содержание красных телец в крови у
Слайд 134
Норма эритроцитов в крови
норме содержание красных телец в крови у
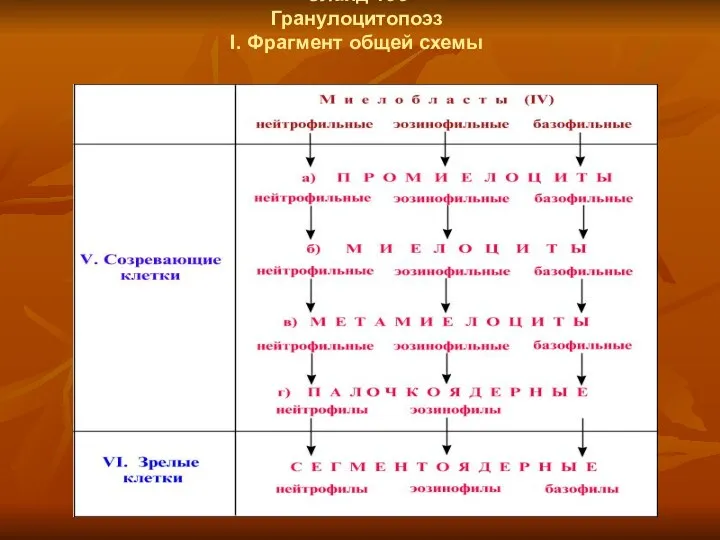
Слайд 135
Гранулоцитопоэз
I. Фрагмент общей схемы
Слайд 135
Гранулоцитопоэз
I. Фрагмент общей схемы

Слайд 136
Промиелоциты - первые клетки класса V
1. Промиелоциты (нейтрофильные, эозинофильные и базофильные) ещё друг
Слайд 136
Промиелоциты - первые клетки класса V
1. Промиелоциты (нейтрофильные, эозинофильные и базофильные) ещё друг

Слайд 137
Ядра, их последующие изменения и способность к делениям.
1. а) Ядра у миелоцитов по-прежнему округлые.
б)
Слайд 137
Ядра, их последующие изменения и способность к делениям.
1. а) Ядра у миелоцитов по-прежнему округлые.
б)

Слайд 138
Клетки завершающих стадий развития
Метамиелоциты: нейтрофильные, эозинофильные, базофильные.
Палочкоядерные гранулоциты: нейтрофильные эозинофильные,
Слайд 138
Клетки завершающих стадий развития
Метамиелоциты: нейтрофильные, эозинофильные, базофильные.
Палочкоядерные гранулоциты: нейтрофильные эозинофильные,

Слайд 139
Моноцитопоэз
Х а р а к т е р и с
Слайд 139
Моноцитопоэз
Х а р а к т е р и с

Слайд 140
Тромбоцитопоэз
Фрагмент общей схемы: Мегакариобласты, Промегакариоцит, Мегакариоцит.
Характеристика клеток. 1. При переходе от
Слайд 140
Тромбоцитопоэз
Фрагмент общей схемы: Мегакариобласты, Промегакариоцит, Мегакариоцит.
Характеристика клеток. 1. При переходе от

Слайд 141
Зрелые форменные элементы класса VI
1. а) Мегакариоцит "проталкивает" часть своей цитоплазмы(в виде отростков)
Слайд 141
Зрелые форменные элементы класса VI
1. а) Мегакариоцит "проталкивает" часть своей цитоплазмы(в виде отростков)

Слайд 144
Слайд 144

Слайд 146
Слайд 146

Слайд 148
Норма тромбоцитов
Количество кровяных пластинок измеряется в тысячах на 1 микролитр
Слайд 148
Норма тромбоцитов
Количество кровяных пластинок измеряется в тысячах на 1 микролитр

Слайд 149
Отношения между кроветворными органами и кровью
Морфологически состав крови зависит от
Слайд 149
Отношения между кроветворными органами и кровью
Морфологически состав крови зависит от

Слайд 150
Отношения между кроветворными органами и кровью
При лейкозах явления гиперплазии кроветворных
Слайд 150
Отношения между кроветворными органами и кровью
При лейкозах явления гиперплазии кроветворных

Слайд 164
Лейкоциты
Лейкоциты, или «белые кровяные тельца» (Л, White Blood Cells, WBC)
Слайд 164
Лейкоциты
Лейкоциты, или «белые кровяные тельца» (Л, White Blood Cells, WBC)

Слайд 165
Лейкоциты
Функции:
1)защитная(фагоцитоз микробов, бактерицидное и антитоксическое действие, участие в иммунных реакциях,
Слайд 165
Лейкоциты
Функции:
1)защитная(фагоцитоз микробов, бактерицидное и антитоксическое действие, участие в иммунных реакциях,

Слайд 166
Отличие гранулоцитов от агранулоцитов.
Агранулоциты - это одноядерные лейкоциты, один из
Слайд 166
Отличие гранулоцитов от агранулоцитов.
Агранулоциты - это одноядерные лейкоциты, один из

Слайд 167
Общие свойства для всех лецкоциов, связанные с функцией:
Способность к хемотаксису
Слайд 167
Общие свойства для всех лецкоциов, связанные с функцией:
Способность к хемотаксису

Слайд 168
Защитная функция лейкоцитов
Главная функция лейкоцитов - защита. Они играют главную
Слайд 168
Защитная функция лейкоцитов
Главная функция лейкоцитов - защита. Они играют главную

Слайд 169
Очистка. Регенерация лейкоцитов
Макрофаги пожирая остатки погибших клеток, очищают место битвы.
Слайд 169
Очистка. Регенерация лейкоцитов
Макрофаги пожирая остатки погибших клеток, очищают место битвы.

Слайд 191
Адгезия
Вовлечение тромбоцитов в процесс гемостаза невозможно без их адгезии
Слайд 191
Адгезия
Вовлечение тромбоцитов в процесс гемостаза невозможно без их адгезии

Слайд 195
Слайд 195
 Гидроцефалия, гипотензивтік синдром, менингеальдық миндром
Гидроцефалия, гипотензивтік синдром, менингеальдық миндром Осложнения лекарственной терапии. Побочные действия и токсические свойства лекарств
Осложнения лекарственной терапии. Побочные действия и токсические свойства лекарств Иммунотропные средства
Иммунотропные средства Спинномозговые нервы
Спинномозговые нервы Нейролептики (антипсихотики)
Нейролептики (антипсихотики) Электрокардиография (ЭКГ)
Электрокардиография (ЭКГ) Заболевание половых органов у девочек и их профилактика. Вульвагинит
Заболевание половых органов у девочек и их профилактика. Вульвагинит Органосохраняющее лечение рака почки в РКОД
Органосохраняющее лечение рака почки в РКОД Переливание крови
Переливание крови Проблема биосовместимости (лекция 3)
Проблема биосовместимости (лекция 3) Внутричерепные отогенные осложнения
Внутричерепные отогенные осложнения Сегментарное строение спинного мозга. Простая и сложная рефлекторные дуги
Сегментарное строение спинного мозга. Простая и сложная рефлекторные дуги Целиакия. Глютеновая энтеропатия
Целиакия. Глютеновая энтеропатия Организация деятельности архива медицинской организации
Организация деятельности архива медицинской организации Лекарственная лихорадка
Лекарственная лихорадка Пациент және оның отбасына реабилитация шараларына үйрету
Пациент және оның отбасына реабилитация шараларына үйрету Индивидуальді жазылым бойынша ұнтақ тәрізді формада шығарылатын косметикалық препараттар
Индивидуальді жазылым бойынша ұнтақ тәрізді формада шығарылатын косметикалық препараттар Герпетическая инфекция
Герпетическая инфекция Вскармливание ребенка одного года
Вскармливание ребенка одного года Доброякісні пухлини ЖСО: класифікація, етіологія, патогенез, клініка, лікування
Доброякісні пухлини ЖСО: класифікація, етіологія, патогенез, клініка, лікування Железодефицитное состояние при беременности: тоже лечим
Железодефицитное состояние при беременности: тоже лечим Коррекция нарушений в сенсорной сфере у детей с РАС
Коррекция нарушений в сенсорной сфере у детей с РАС System of pharmacovigilance in ukraine. Concept of side effects of drugs
System of pharmacovigilance in ukraine. Concept of side effects of drugs Лайм-боррелиоз. Факторы патогенности
Лайм-боррелиоз. Факторы патогенности Пути и способы введение лекарственных средств
Пути и способы введение лекарственных средств Гостра дихальна недостатність
Гостра дихальна недостатність Менструальді циклдің нейроэндокринді реттелісі
Менструальді циклдің нейроэндокринді реттелісі Миксовирусы. Вирусологическая диагностика гриппа, парагриппа, кори
Миксовирусы. Вирусологическая диагностика гриппа, парагриппа, кори