- Секвенирование ДНК по Сэнгеру. Курс 3 ЦИОП Медицина будущего

Содержание
- 2. Секвенирование геномов
- 3. Стратегии секвенирования геномов Метод дробовика (небольшие геномы) Метод сборки контигов из клонов
- 4. Клонирование в векторную последовательность
- 5. Задачи, решаемые методом секвенирования ДНК Секвенирование – это определение последовательности нуклеотидов в молекуле ДНК Зачем нужно
- 6. Кто и когда разработал методы секвенирования ДНК 1953 г. – двойная спираль ДНК Д. Уотсона и
- 7. Плюс-минус метод Сэнгера-Коулсона Разработан к 1975 г. Сэнгером и Коулсоном В "плюс" системе проводили четыре реакции
- 8. Метод химической деградации Максама-Гилберта Метод основан на химической деградации ДНК.
- 9. Метод основан на химической деградации ДНК. Суть метода сводится к следующему: один из концов фрагмента ДНК
- 10. Оригинальный ферментативный метод Сэнгера В основе метода лежало ферментативное копирование с помощью фрагмента Кленова ДНК полимеразы
- 11. Развитие метода дидезокситерминаторов Эволюция метода Сэнгера Чтение нуклеотидной последовательности
- 12. Автоматическое секвенирование Преимущества флуоресцентных меток над изотопными: Отсутствует радиоактивное загрязнение Меньшая трудоемкость Экономичность Автоматизация секвенирования ACCTGATGCCAGTTGCAAGT
- 13. Пиросеквенирование Принцип метода довольно прост и основан на (+/-)-секвенировании, предложенном ещё в 60-х годах. При последовательном
- 14. Развитие сопутствующих технологий для автоматизации метода Сэнегра: флуоресцентные красители Флуоресценция – излучение, возникающее при переходе электрона
- 15. Развитие сопутствующих технологий для автоматизации метода Сэнегра: полимераза и электрофорез Полимеразы для автоматического секвенирования ДНК В
- 16. Рутинные задачи, решаемые на капиллярных секвенаторах Высокоразрешающий капиллярный электрофорез с детекцией флуоресценции Секвенирование Фрагментный анализ Плазмида
- 17. Обзор современных капиллярных секвенаторов 310 Genetic Analyzer (1 капилляр) 3100 Avant и 3130 Genetic analyzer (4
- 18. Прямая ДНК-диагностика наследственных заболеваний ACA CTG CCA GTG ACA CTG CTA GTG T L P V
- 19. Последовательность действий при секвенировании по Сэнгеру Получение матрицы для секвенирования (ПЦР-продукт, плазмида) Очистка матрицы от невключившихся
- 20. Очистка ПЦР-продукта перед секвенированием Неспецифические продукты амплификации Да Нет Далеко от целевого ПЦР-продукта Близко к целевому
- 21. Очистка ПЦР-продукта перед секвенированием в отсутствие неспецифических продуктов амплификации Обработка аликвоты ПЦР-продуктов смесью экзонуклеазы I из
- 22. Очистка целевого ПЦР-продукта от неспецифических фрагментов с помощью электрофореза Агарозный гель-электрофорез (1,5% агарозы, 0,5х-буфер ТВЕ, 150-200
- 23. Методы эллюции и очистки ПЦР-продукта 1. Растворение LMP-агарозы 1. Вырезать полосу геля, содержащую анализируемый ПЦР-продукт, взвесить.
- 24. Проведение реакции наработки меченых фрагментов Требования некоторых лабораторий к образцам для секвенирования «Синтол»: не менее 10
- 25. Очистка образцов после реакции секвенирования Необходимо избавиться от: невключившихся меченых ddNTPs ионов, которые могут влиять на
- 26. Какие бывают результаты? Сиквенс высокого качества (плазмидная ДНК)
- 27. Не прошла (или прошла с очень низкой эффективностью) реакция секвенирования
- 28. В образце присутствует неспецифическая матричная последовательность ДНК, или продукты мечения комплементарной цепи, или аллель с делецией
- 29. 1. Мало ДНК в образце, 2 и 3 – недостаточная очистка от ddNTPs после реакции секвенирования
- 30. Димеры праймеров или короткая неспецифика в образце
- 31. ABI Prizm® Sequencing Analysis Software – коммерческая программа для получения и редактирования хроматограмм секвенирования, поставляется вместе
- 33. Скачать презентацию
Секвенирование геномов
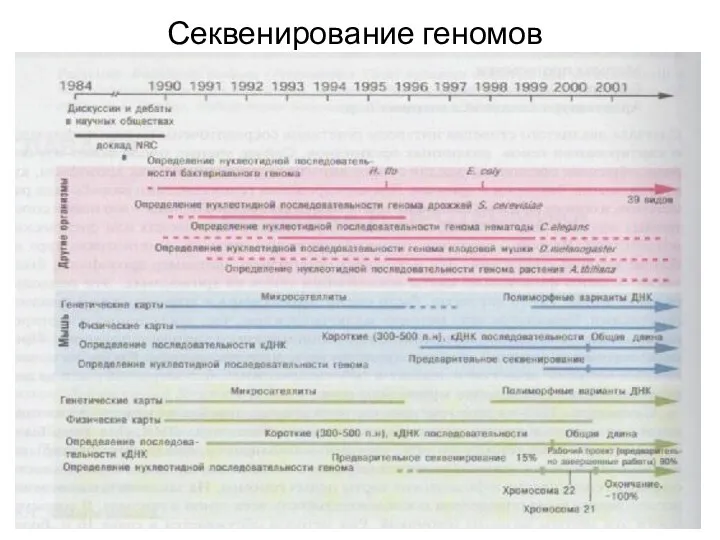
Секвенирование геномов
Стратегии секвенирования геномов
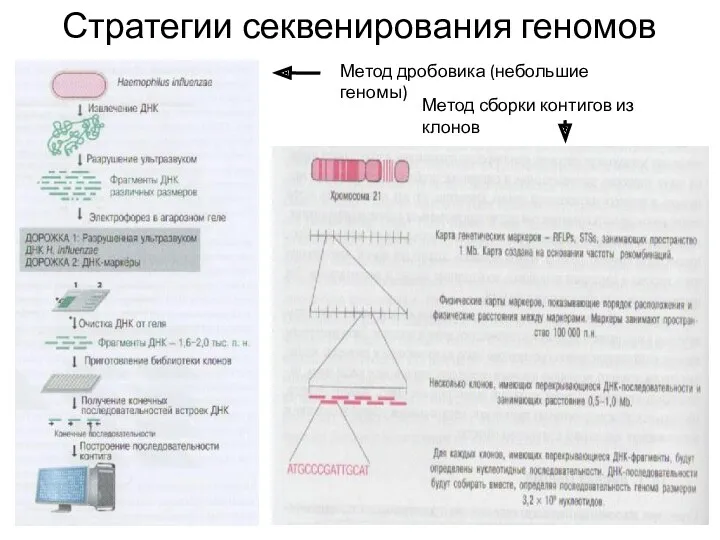
Метод дробовика (небольшие геномы)
Метод сборки контигов из клонов
Стратегии секвенирования геномов
Метод дробовика (небольшие геномы)
Метод сборки контигов из клонов
Клонирование в векторную последовательность
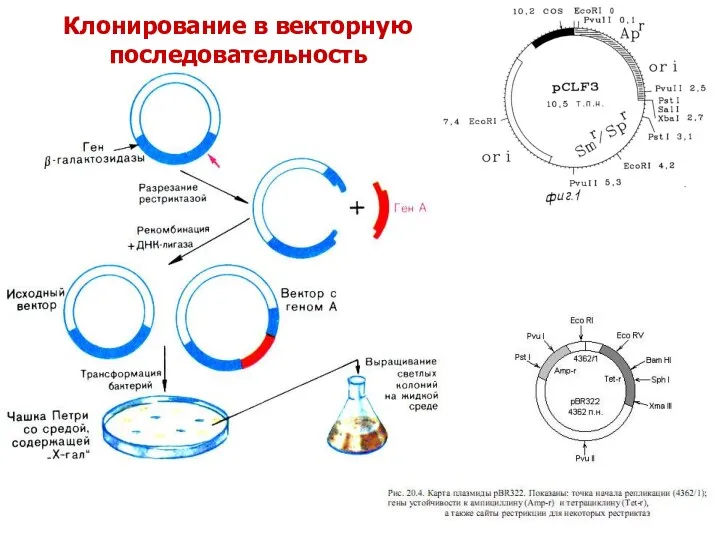
Клонирование в векторную последовательность

Задачи, решаемые методом секвенирования ДНК
Секвенирование – это определение последовательности нуклеотидов в
Задачи, решаемые методом секвенирования ДНК
Секвенирование – это определение последовательности нуклеотидов в

Кто и когда разработал методы секвенирования ДНК
1953 г. – двойная спираль
Кто и когда разработал методы секвенирования ДНК
1953 г. – двойная спираль
Плюс-минус метод Сэнгера-Коулсона
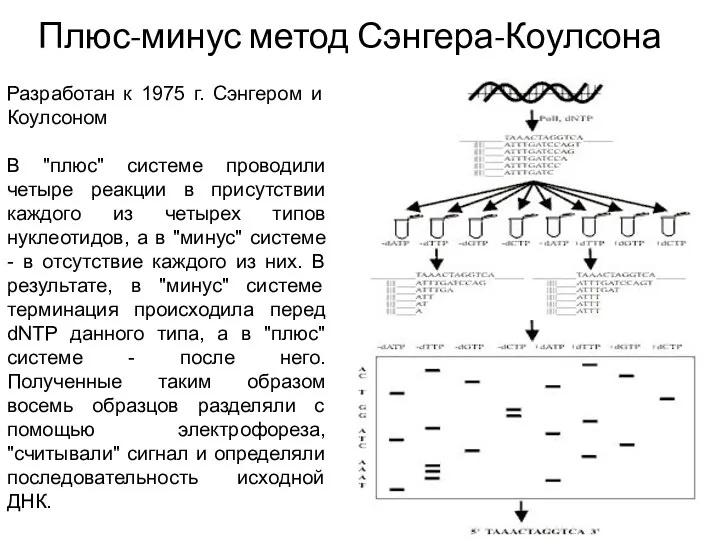
Разработан к 1975 г. Сэнгером и Коулсоном
В "плюс" системе
Плюс-минус метод Сэнгера-Коулсона
Разработан к 1975 г. Сэнгером и Коулсоном
В "плюс" системе
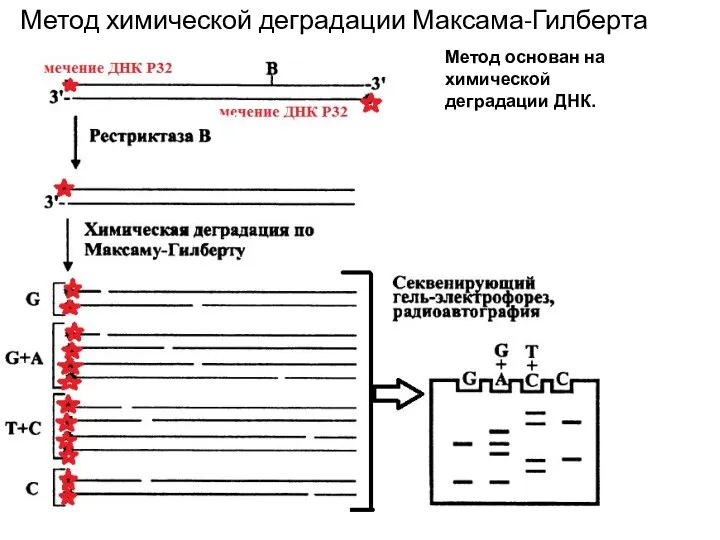
Метод химической деградации Максама-Гилберта
Метод основан на химической деградации ДНК.
Метод химической деградации Максама-Гилберта
Метод основан на химической деградации ДНК.

Метод основан на химической деградации ДНК.
Суть метода сводится к следующему:
Метод основан на химической деградации ДНК.
Суть метода сводится к следующему:
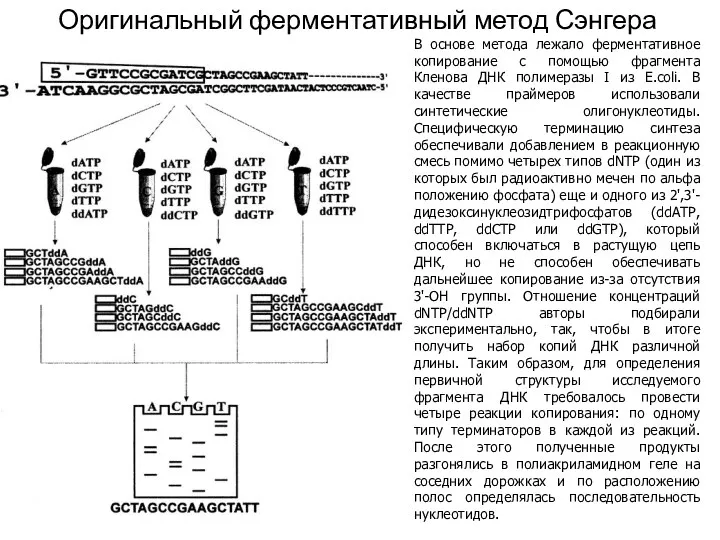
Оригинальный ферментативный метод Сэнгера
В основе метода лежало ферментативное копирование с помощью
Оригинальный ферментативный метод Сэнгера
В основе метода лежало ферментативное копирование с помощью

Развитие метода дидезокситерминаторов
Эволюция метода Сэнгера
Чтение нуклеотидной последовательности
Развитие метода дидезокситерминаторов
Эволюция метода Сэнгера
Чтение нуклеотидной последовательности
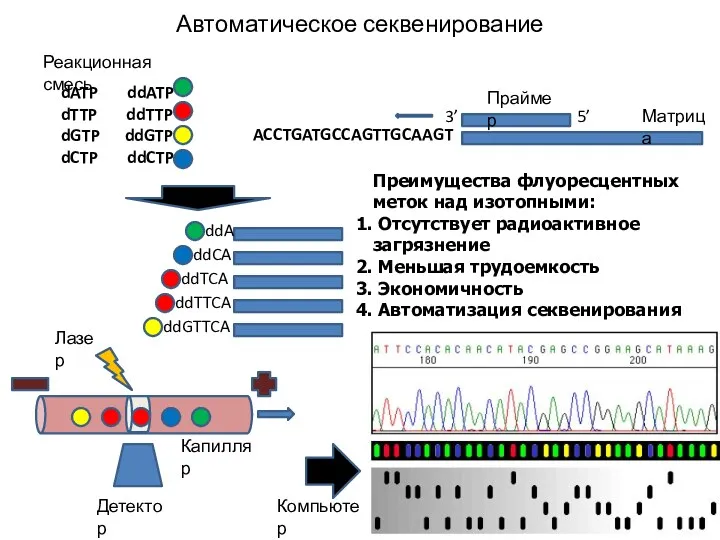
Автоматическое секвенирование
Преимущества флуоресцентных меток над изотопными:
Отсутствует радиоактивное загрязнение
Меньшая трудоемкость
Автоматическое секвенирование
Преимущества флуоресцентных меток над изотопными:
Отсутствует радиоактивное загрязнение
Меньшая трудоемкость
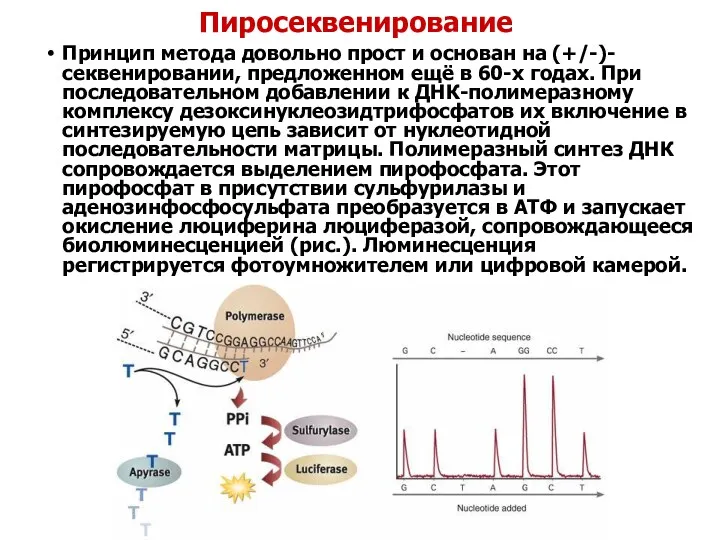
Пиросеквенирование
Принцип метода довольно прост и основан на (+/-)-секвенировании, предложенном ещё
Пиросеквенирование
Принцип метода довольно прост и основан на (+/-)-секвенировании, предложенном ещё

Развитие сопутствующих технологий для автоматизации метода Сэнегра: флуоресцентные красители
Флуоресценция – излучение,
Развитие сопутствующих технологий для автоматизации метода Сэнегра: флуоресцентные красители
Флуоресценция – излучение,

Развитие сопутствующих технологий для автоматизации метода Сэнегра: полимераза и электрофорез
Полимеразы для
Развитие сопутствующих технологий для автоматизации метода Сэнегра: полимераза и электрофорез
Полимеразы для
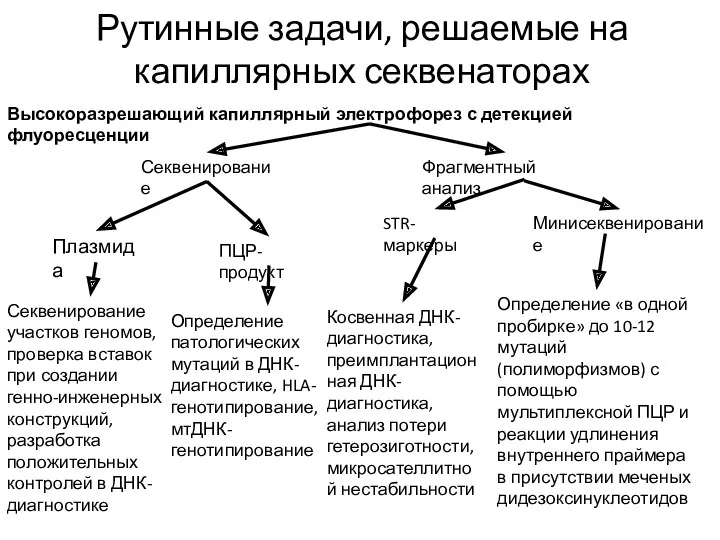
Рутинные задачи, решаемые на капиллярных секвенаторах
Высокоразрешающий капиллярный электрофорез с детекцией флуоресценции
Секвенирование
Фрагментный
Рутинные задачи, решаемые на капиллярных секвенаторах
Высокоразрешающий капиллярный электрофорез с детекцией флуоресценции
Секвенирование
Фрагментный
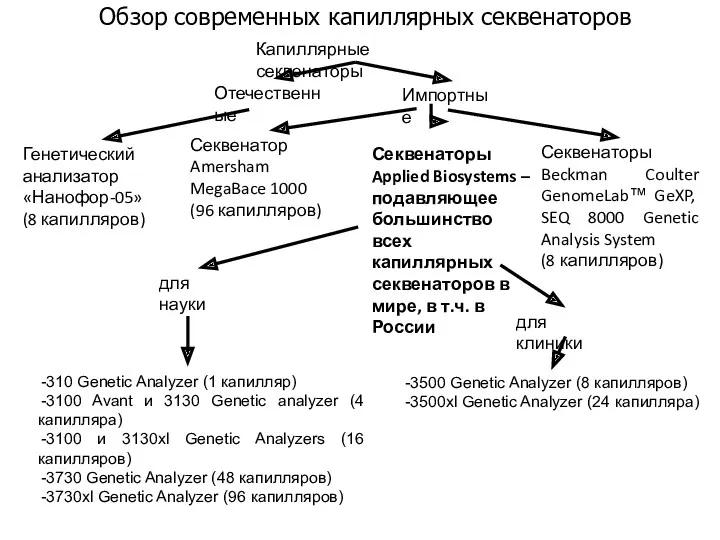
Обзор современных капиллярных секвенаторов
310 Genetic Analyzer (1 капилляр)
3100 Avant и 3130
Обзор современных капиллярных секвенаторов
310 Genetic Analyzer (1 капилляр)
3100 Avant и 3130
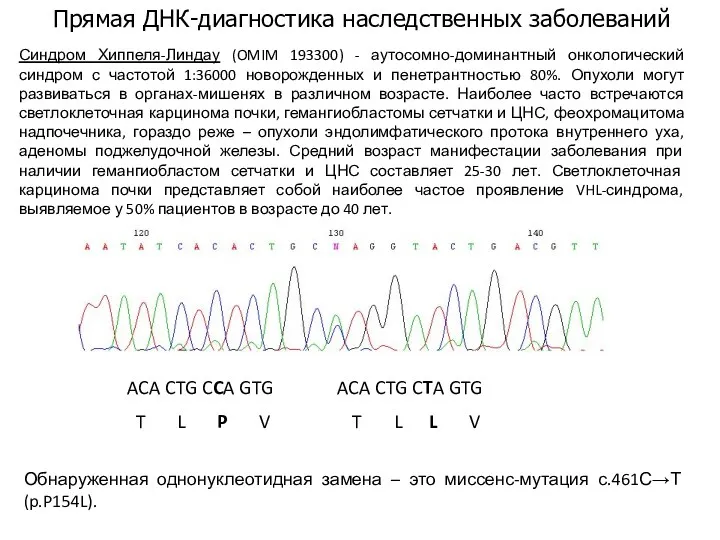
Прямая ДНК-диагностика наследственных заболеваний
ACA CTG CCA GTG ACA CTG CTA GTG
Прямая ДНК-диагностика наследственных заболеваний
ACA CTG CCA GTG ACA CTG CTA GTG

Последовательность действий при секвенировании по Сэнгеру
Получение матрицы для секвенирования (ПЦР-продукт, плазмида)
Очистка
Последовательность действий при секвенировании по Сэнгеру
Получение матрицы для секвенирования (ПЦР-продукт, плазмида)
Очистка
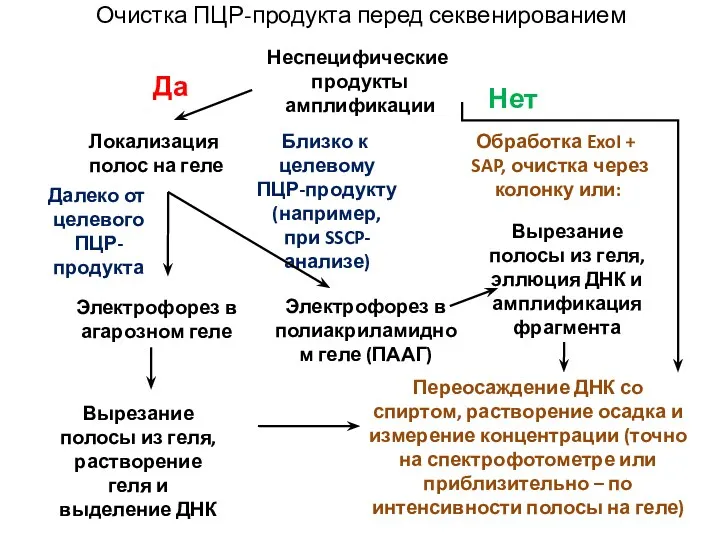
Очистка ПЦР-продукта перед секвенированием
Неспецифические продукты амплификации
Да
Нет
Далеко от целевого ПЦР-продукта
Очистка ПЦР-продукта перед секвенированием
Неспецифические продукты амплификации
Да
Нет
Далеко от целевого ПЦР-продукта

Очистка ПЦР-продукта перед секвенированием в отсутствие неспецифических продуктов амплификации
Обработка аликвоты
Очистка ПЦР-продукта перед секвенированием в отсутствие неспецифических продуктов амплификации
Обработка аликвоты

Очистка целевого ПЦР-продукта от неспецифических фрагментов с помощью электрофореза
Агарозный гель-электрофорез
(1,5%
Очистка целевого ПЦР-продукта от неспецифических фрагментов с помощью электрофореза
Агарозный гель-электрофорез
(1,5%

Методы эллюции и очистки ПЦР-продукта
1. Растворение LMP-агарозы
1. Вырезать полосу геля,
Методы эллюции и очистки ПЦР-продукта
1. Растворение LMP-агарозы
1. Вырезать полосу геля,

Проведение реакции наработки меченых фрагментов
Требования некоторых лабораторий к образцам для
Проведение реакции наработки меченых фрагментов
Требования некоторых лабораторий к образцам для

Очистка образцов после реакции секвенирования
Необходимо избавиться от:
невключившихся меченых ddNTPs
ионов, которые
Очистка образцов после реакции секвенирования
Необходимо избавиться от:
невключившихся меченых ddNTPs
ионов, которые
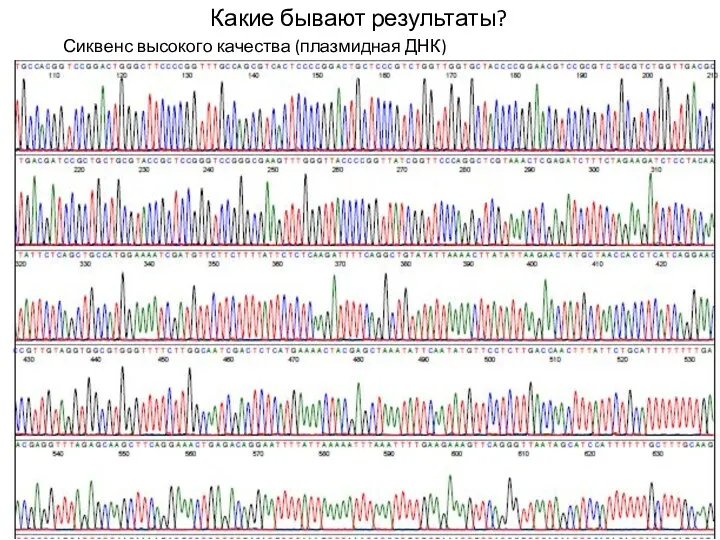
Какие бывают результаты?
Сиквенс высокого качества (плазмидная ДНК)
Какие бывают результаты?
Сиквенс высокого качества (плазмидная ДНК)
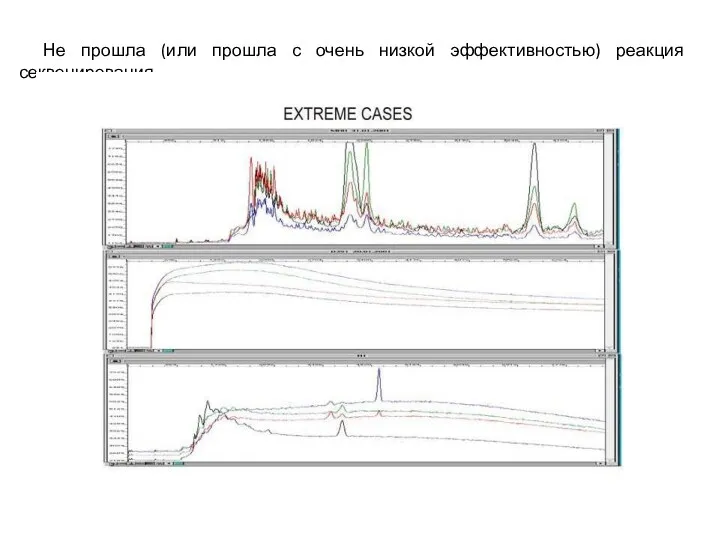
Не прошла (или прошла с очень низкой эффективностью) реакция секвенирования
Не прошла (или прошла с очень низкой эффективностью) реакция секвенирования
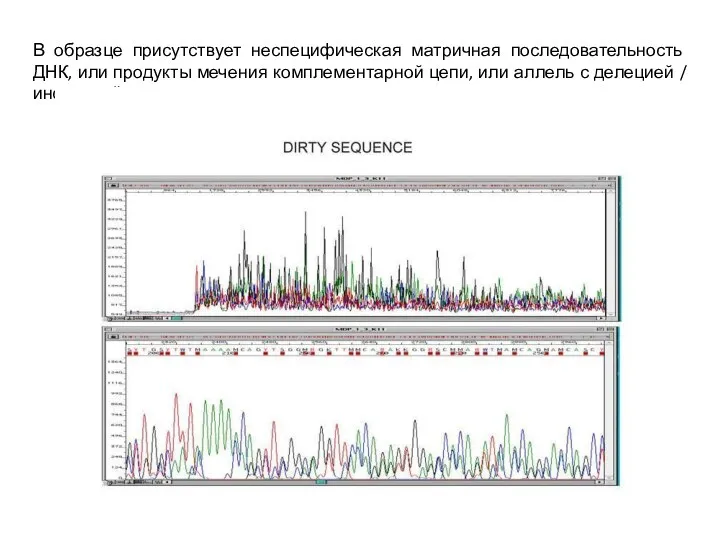
В образце присутствует неспецифическая матричная последовательность ДНК, или продукты мечения комплементарной
В образце присутствует неспецифическая матричная последовательность ДНК, или продукты мечения комплементарной
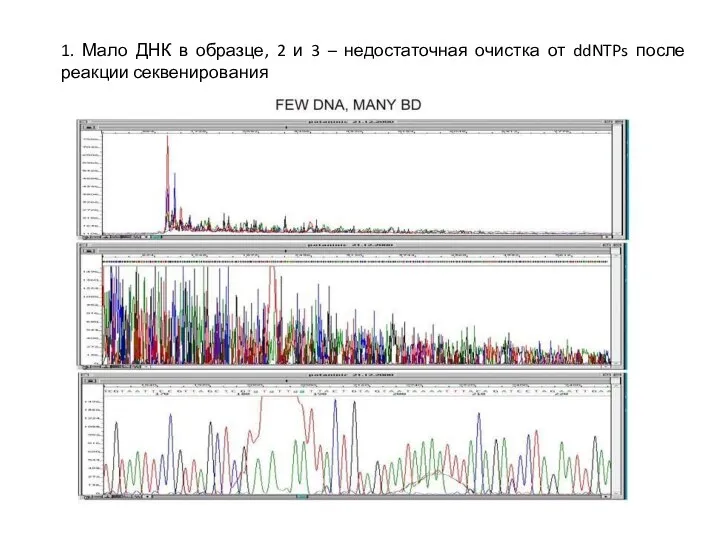
1. Мало ДНК в образце, 2 и 3 – недостаточная очистка
1. Мало ДНК в образце, 2 и 3 – недостаточная очистка
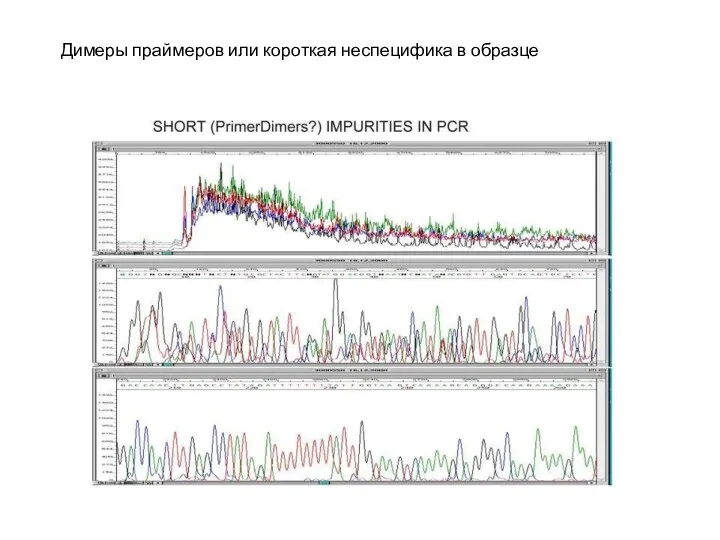
Димеры праймеров или короткая неспецифика в образце
Димеры праймеров или короткая неспецифика в образце

ABI Prizm® Sequencing Analysis Software – коммерческая программа для получения и
ABI Prizm® Sequencing Analysis Software – коммерческая программа для получения и
 Балалардағы гельминтоздар (аскаридоз, энтеробиоз, трихинеллез)
Балалардағы гельминтоздар (аскаридоз, энтеробиоз, трихинеллез) Врожденные пороки сердца. Недостаточность кровообращения
Врожденные пороки сердца. Недостаточность кровообращения Туберкулез костей и суставов
Туберкулез костей и суставов Реанимация. Методы реанимации. Реанимация и интенсивная терапия при внезапной остановке кровообращения
Реанимация. Методы реанимации. Реанимация и интенсивная терапия при внезапной остановке кровообращения Системная склеродермия
Системная склеродермия Дефицит массы тела
Дефицит массы тела Екстракорпоральне запліднення
Екстракорпоральне запліднення Гигиенические требования к земельному участку, зданию ДОУ и планировке помещений
Гигиенические требования к земельному участку, зданию ДОУ и планировке помещений Омыртқа жотасының қисаюы
Омыртқа жотасының қисаюы Возбудители бактериальных кишечных инфекций
Возбудители бактериальных кишечных инфекций Анаэробная инфекция
Анаэробная инфекция Сестринский процесс при бронхиальной астме
Сестринский процесс при бронхиальной астме Сердечный насос. Ветеринарная клиника ВИТА
Сердечный насос. Ветеринарная клиника ВИТА Догляд за недоношеними дітьми після народження
Догляд за недоношеними дітьми після народження БМСК ұйымдарында тіркеу және есеп беру құжаттарын рәсімдеу
БМСК ұйымдарында тіркеу және есеп беру құжаттарын рәсімдеу Заболевание гипоталамо-гипофизарной системы. Ее участие в корегировании функциональной активности желез внутренней секреции
Заболевание гипоталамо-гипофизарной системы. Ее участие в корегировании функциональной активности желез внутренней секреции Защитная функция крови
Защитная функция крови Опухоли головного мозга
Опухоли головного мозга Фармакологические средства, влияющие на передачу возбуждения в холинэргических синапсах
Фармакологические средства, влияющие на передачу возбуждения в холинэргических синапсах The Functional Histology of Respiratory System
The Functional Histology of Respiratory System Особенности сестринского ухода за пациентами с панкреатитом
Особенности сестринского ухода за пациентами с панкреатитом Бронхиальная астма
Бронхиальная астма Анатомия пәні және мақсаты. Тіндерге, ағзаларға және ағзалар жүйесіне жалпы сипаттама. Адам эмбриогенезінің алғашқы сатысы
Анатомия пәні және мақсаты. Тіндерге, ағзаларға және ағзалар жүйесіне жалпы сипаттама. Адам эмбриогенезінің алғашқы сатысы 40-55 жас аралығындағы ішек өткізбеушілігі диагнозымен ауыратын науқастарда перитониттің алдын алу үшін энтеростомияға
40-55 жас аралығындағы ішек өткізбеушілігі диагнозымен ауыратын науқастарда перитониттің алдын алу үшін энтеростомияға Анемии. Классификация анемий
Анемии. Классификация анемий Сау және науқас адамдарға терең веналарының өткізгіштігін тексеру әдістерін жүргізу
Сау және науқас адамдарға терең веналарының өткізгіштігін тексеру әдістерін жүргізу Расстройства аутистического спектра (искажённый вид психического дизонтогенеза). Лекция №7
Расстройства аутистического спектра (искажённый вид психического дизонтогенеза). Лекция №7 Иппотерапия
Иппотерапия