- Транскрипция. Гендерді әртүрлі тиімділікпен экспрессиялануы мүмкін

Содержание
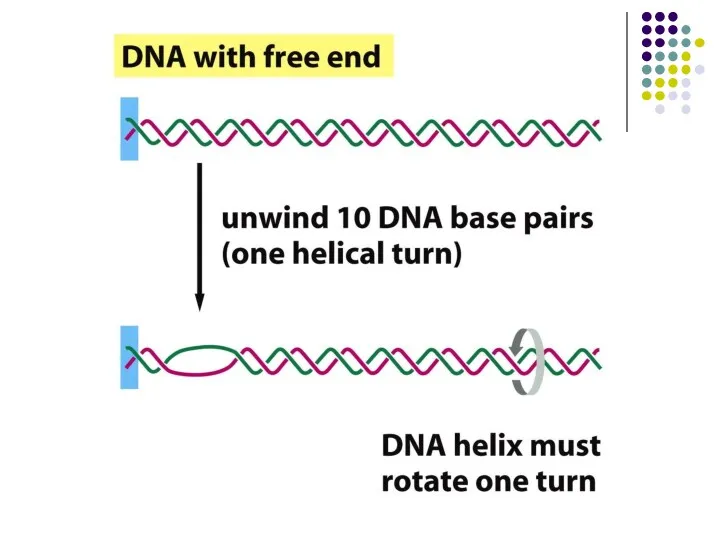
- 2. Ақпараттың екі тізбекті ДНҚ -дан бір тізбекті РНҚ- ға РНК-транскриптаза немесе РНК-полимераза деп аталатын ферменттер көмегімен
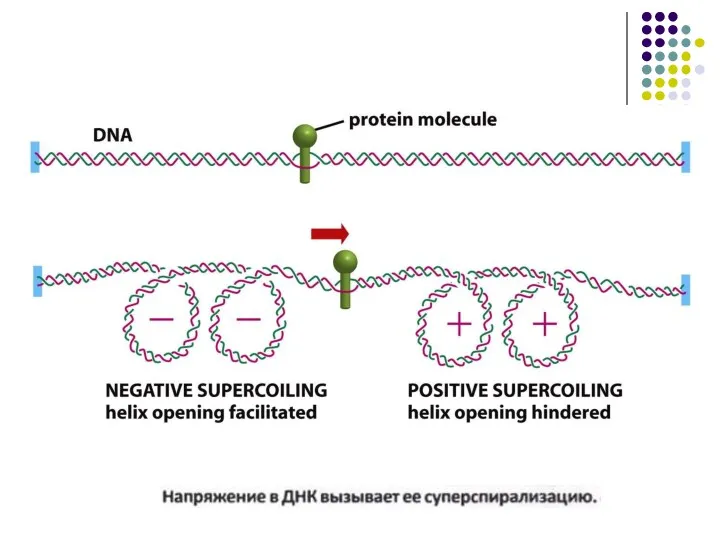
- 3. А.ДНК матрицадағы РНК транскрипция процесінің схематикалық бейнеленуі. Транскрипцияға қатысатын матрицалық тізбекті білу үшін ДНҚ спиралі екі
- 4. Гендерді әртүрлі тиімділікпен экспрессиялануы мүмкін
- 7. Урацил аденинмен комплементарлы тізбек түзеді
- 12. Е.СоІі-дің ДНҚ полимеразасыРНҚның барлық 3 классының: рибосомалық рРНҚ, транспорттық тРНҚ, ақпараттық немесе матрицалық мРНҚ молекулаларының реті
- 13. Эукариоттардың РНК-полимеразасы және оның транскрипттері
- 14. РНҚ полимераза І ядрошықта орналасып, 18S, 5,8S, 28S гендерін транскрипциялайды. Бұл РНҚ тізбегі тұтас ізашар ретінде
- 15. Ашытқыларда 5S рРНҚ гені рРНҚның қайталама ізашар гендерінің арасында қарама – қарсы бағытта орналасады. Эукариоттарда 5S
- 16. Ашытқылар мен эукариоттардың рРНК гендері
- 17. РНҚ полимеразаның суббірліктері І,ІІ,ІІІ түріне қатысына байланысты А,В,С әріптерімен белгіленеді. Кейбір ақуыздар бірнеше РНҚ полимеразаның құрамына
- 19. РНҚ полимераза І –ді тек бір транскрипт синтездейді. РНҚ полимераза І –дің транскрипциондық белсенділігі жасушалық циклдың
- 20. рРНҚның ізашар гендік реттеуші элементтерінің құрылымы. Транскрипцияны реттеуші РНҚ полимераза І мен ақуыздар
- 21. РНҚ полимераза ІІІ рРНҚның транспорттық РНҚның және бірнеше кіші ядролы РНҚның 5S генін транскрипциялайды. Гендердің бұл
- 22. РНК полимераза III –пен транскрипцияланатын гендер. Транскрипцияны қамтамасыз ететін ақуыздар
- 23. Кіші ядролы РНҚ генінің промоторы мүлдем өзгеше құрылған (РНҚ полимераза ІІІ –пен транскрипцияланады). Бұл гендердің барлық
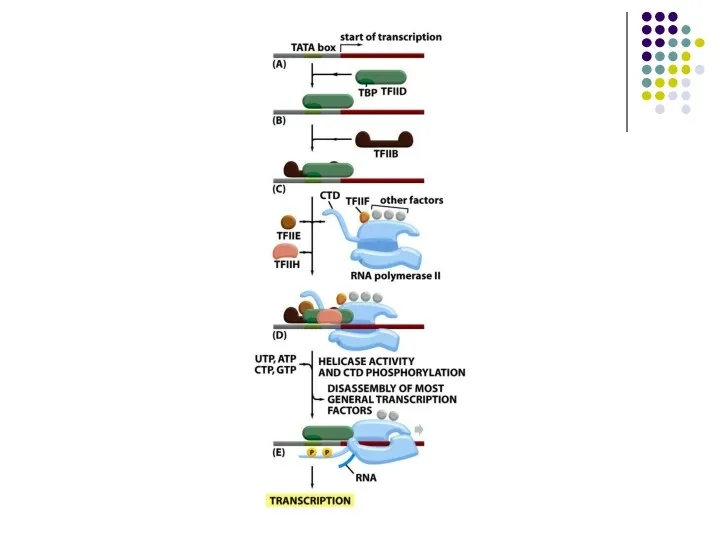
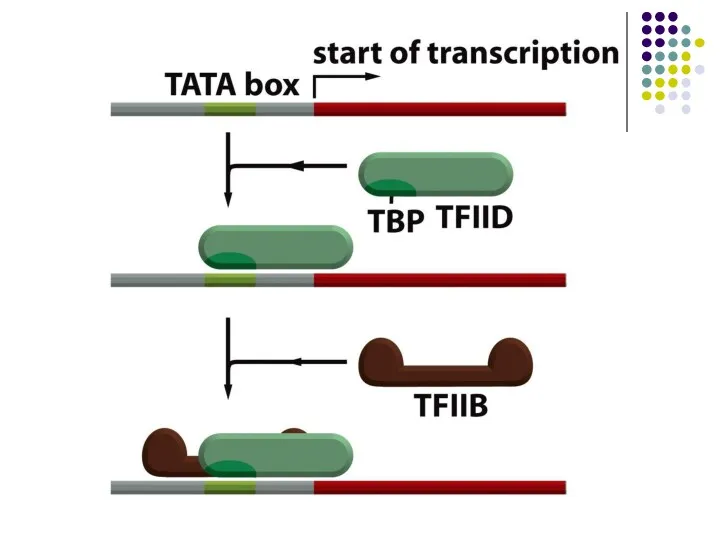
- 24. Барлық 3 типтегі промоторларға ортақ механизм – басқа ДНҚ байланыстырушы белсендіруші ақуыздар көмегімен ТҒІІІВ –ның промоторға
- 25. РНК полимеразы III жұмысының реттелуі
- 26. ТВІІІВ компоненттері тікелей фосфорилдену көмегімен де, репрессор ақуыз Rb және p53 арқылы да реттеледі. La ақуызы
- 27. РНҚ полимераза ІІ РНҚның мРНҚ және кіші РНҚның түрлі класстарын транскрипцилайды. Бұл фермент полимеразаның 3 классының
- 28. РНК полимераза I және ІІІ сияқты соңғы транскрипция инициация стадиясы РНК полимераза ІІ промоторға тігілуі,атап айтқанда
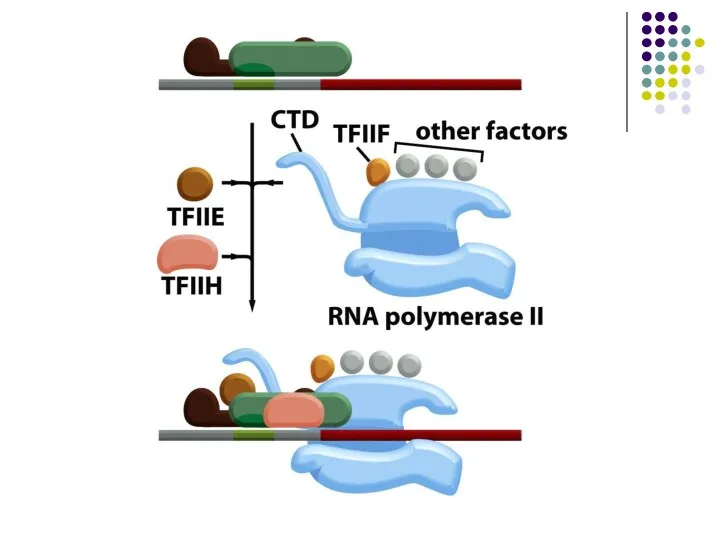
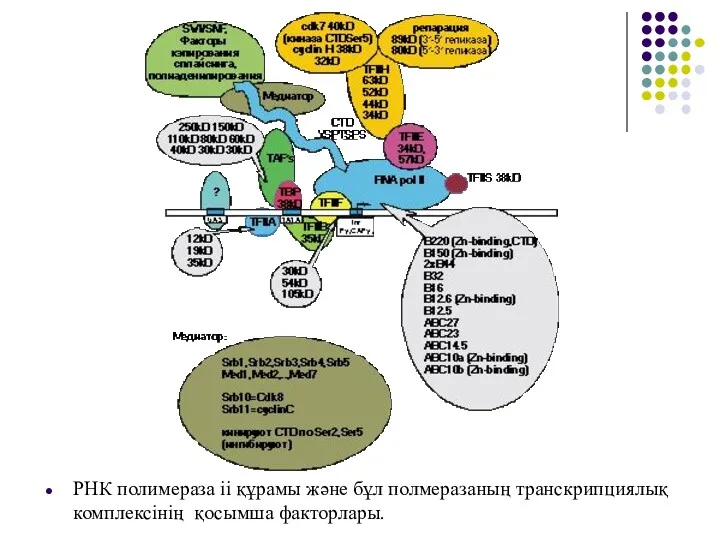
- 31. РНК полимераза іі құрамы және бұл полмеразаның транскрипциялық комплексінің қосымша факторлары.
- 32. Жазылған факторлардан басқа РНК полимераза ІІ транскрипция инициациясында басқа да көптегендері қатысады. Олар әртүрлі гендердің транскрипциасының
- 33. Фосфорленбеген (инициация стадиясында) мен РНК полимераза С-сонды домені ақуызды комплекспен әрекеттеседі, ол медиатор деп аталады. Кейбір
- 34. РНК полимераза ІІ барлық белсенділігін реттеу фосфорленген оның С-соңды домені Ser2 және Ser5 көмегімен іске асуы
- 35. Медиатор – бұл ақуызды комплекс, РНК полимераза ІІ С-соңды доменмен ассоцирленген. Алғаш ол ашытқыдан табылған және
- 36. Медиатор: құрамы және РНК полимераза ІІ-мен әрекеттесуі
- 37. Медиаторда 3 модулді айыру мүмкін болды. Gal11 модулі ДНК байланыстырушы активаторымен немесе транскрипция ингибиторымен жеке бірлесіп
- 38. Жоғарғы эукариоттарда медиатордың гомологтары табылған болатын. Күтуге болатындай, ұқсас комплекстердің бірнешеуі табылды, сонымен қатар олардың құрамы
- 39. РНК полимераза ІІ-нің элонгационды күйінің басты маркері - фосфорленген СTD. Элонгация процесінде фосфорленген күйінің ұсталуына Сdk9
- 40. РНК полимераза ІІ транскрипциясының элонгациясы
- 41. Элонгация процессінде РНК полимеразасы прокариоттың РНК полимеразасы сияқты мәселесімен кездесуі мүмкін: ДНК зақымдалуы, пауза туғызатын және
- 46. Скачать презентацию

Ақпараттың екі тізбекті ДНҚ -дан бір тізбекті РНҚ- ға РНК-транскриптаза немесе
Ақпараттың екі тізбекті ДНҚ -дан бір тізбекті РНҚ- ға РНК-транскриптаза немесе
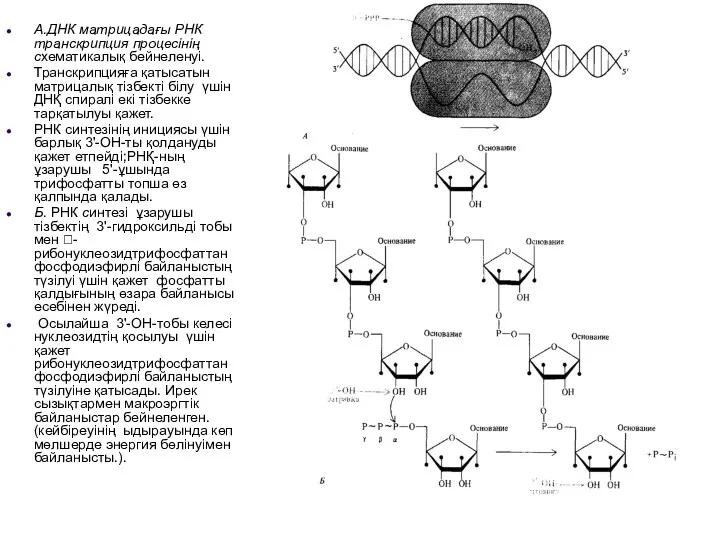
А.ДНК матрицадағы РНК транскрипция процесінің схематикалық бейнеленуі.
Транскрипцияға қатысатын матрицалық тізбекті
А.ДНК матрицадағы РНК транскрипция процесінің схематикалық бейнеленуі.
Транскрипцияға қатысатын матрицалық тізбекті
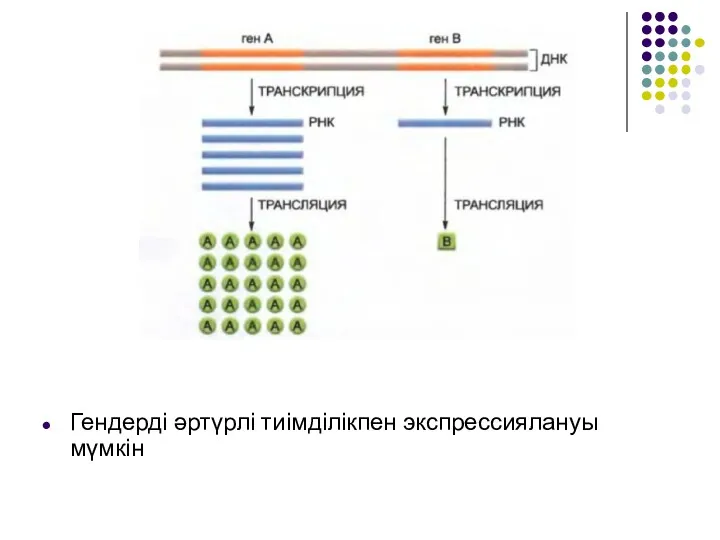
Гендерді әртүрлі тиімділікпен экспрессиялануы мүмкін
Гендерді әртүрлі тиімділікпен экспрессиялануы мүмкін
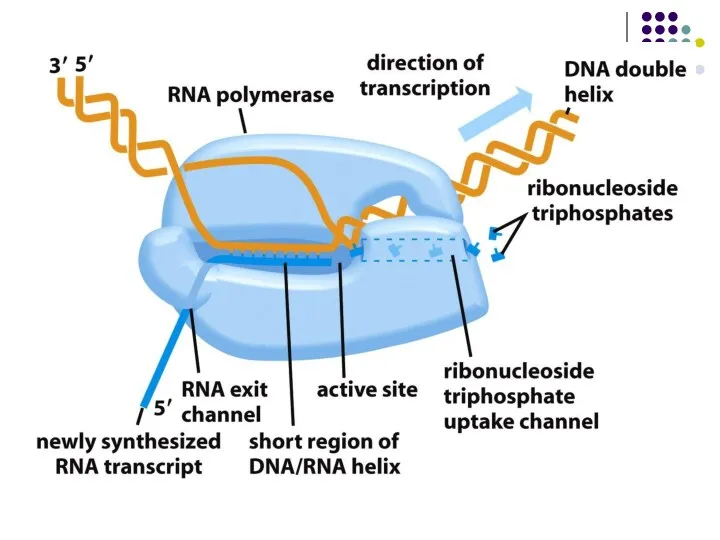
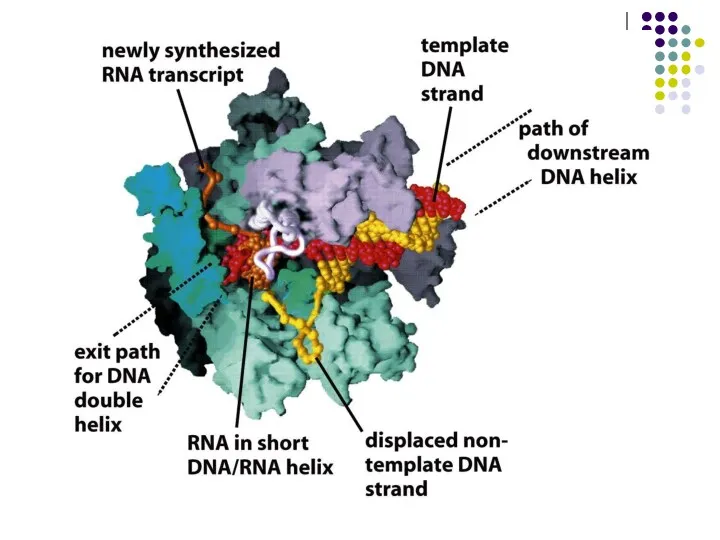
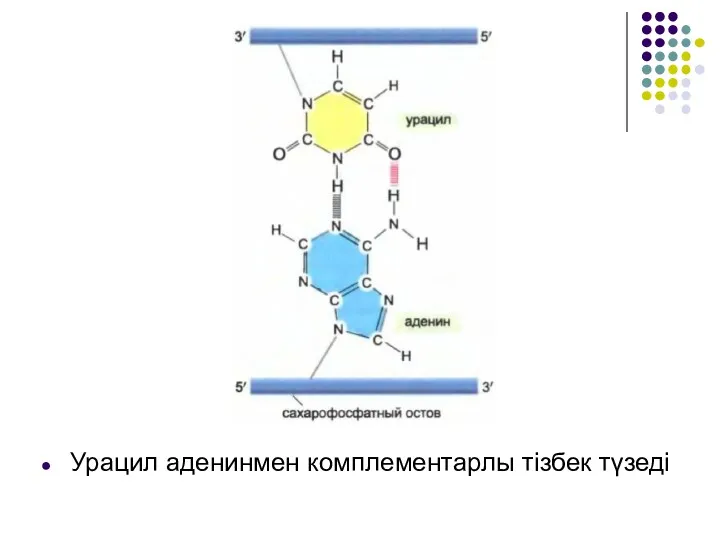
Урацил аденинмен комплементарлы тізбек түзеді
Урацил аденинмен комплементарлы тізбек түзеді



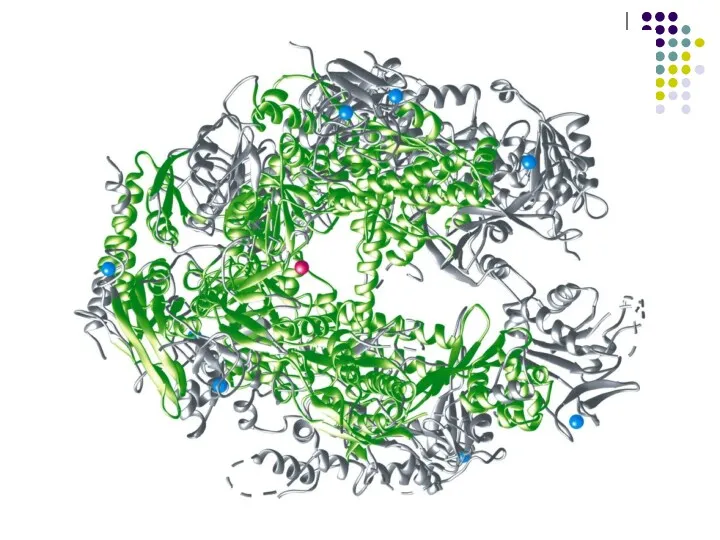

Е.СоІі-дің ДНҚ полимеразасыРНҚның барлық 3 классының: рибосомалық рРНҚ, транспорттық тРНҚ, ақпараттық
Е.СоІі-дің ДНҚ полимеразасыРНҚның барлық 3 классының: рибосомалық рРНҚ, транспорттық тРНҚ, ақпараттық

Эукариоттардың РНК-полимеразасы және оның транскрипттері
Эукариоттардың РНК-полимеразасы және оның транскрипттері

РНҚ полимераза І ядрошықта орналасып, 18S, 5,8S, 28S гендерін транскрипциялайды.
Бұл РНҚ
РНҚ полимераза І ядрошықта орналасып, 18S, 5,8S, 28S гендерін транскрипциялайды.
Бұл РНҚ

Ашытқыларда 5S рРНҚ гені рРНҚның қайталама ізашар гендерінің арасында қарама –
Ашытқыларда 5S рРНҚ гені рРНҚның қайталама ізашар гендерінің арасында қарама –
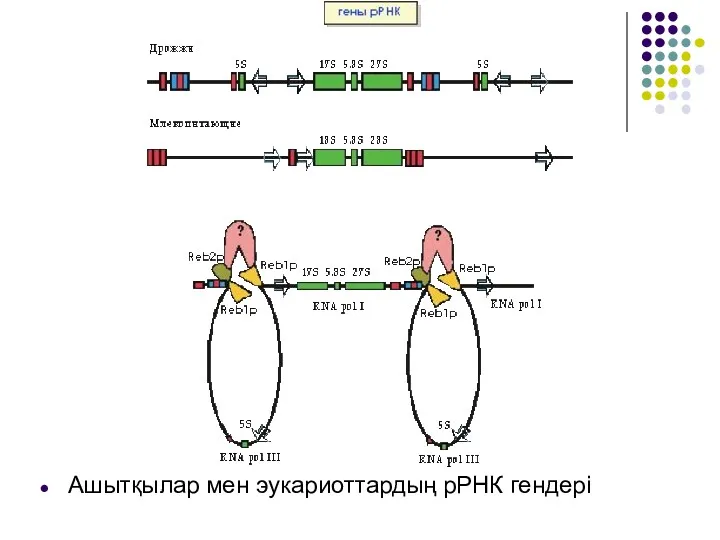
Ашытқылар мен эукариоттардың рРНК гендері
Ашытқылар мен эукариоттардың рРНК гендері

РНҚ полимеразаның суббірліктері І,ІІ,ІІІ түріне қатысына байланысты А,В,С әріптерімен белгіленеді.
Кейбір ақуыздар
РНҚ полимеразаның суббірліктері І,ІІ,ІІІ түріне қатысына байланысты А,В,С әріптерімен белгіленеді.
Кейбір ақуыздар

РНҚ полимераза І –ді тек бір транскрипт синтездейді.
РНҚ полимераза І –дің
РНҚ полимераза І –ді тек бір транскрипт синтездейді.
РНҚ полимераза І –дің
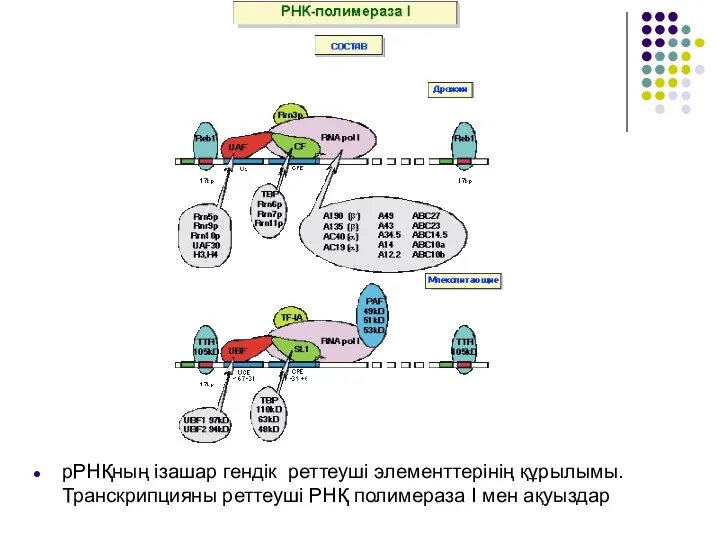
рРНҚның ізашар гендік реттеуші элементтерінің құрылымы. Транскрипцияны реттеуші РНҚ полимераза І
рРНҚның ізашар гендік реттеуші элементтерінің құрылымы. Транскрипцияны реттеуші РНҚ полимераза І

РНҚ полимераза ІІІ рРНҚның транспорттық РНҚның және бірнеше кіші ядролы РНҚның
РНҚ полимераза ІІІ рРНҚның транспорттық РНҚның және бірнеше кіші ядролы РНҚның
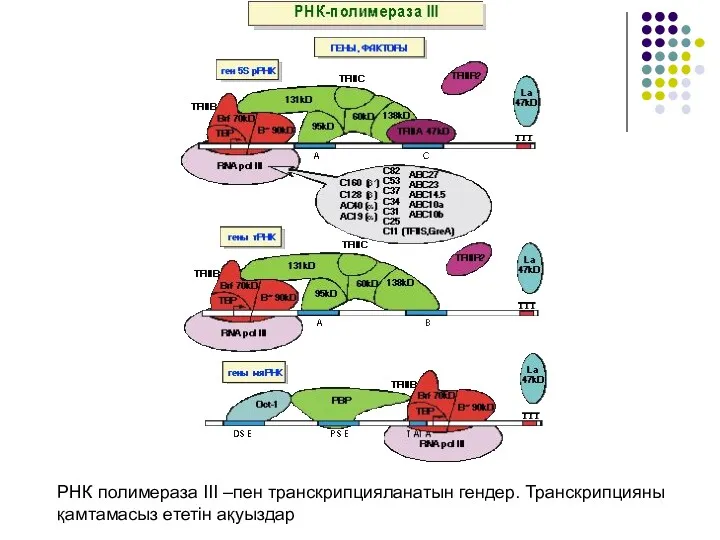
РНК полимераза III –пен транскрипцияланатын гендер. Транскрипцияны қамтамасыз ететін ақуыздар
РНК полимераза III –пен транскрипцияланатын гендер. Транскрипцияны қамтамасыз ететін ақуыздар

Кіші ядролы РНҚ генінің промоторы мүлдем өзгеше құрылған (РНҚ полимераза ІІІ
Кіші ядролы РНҚ генінің промоторы мүлдем өзгеше құрылған (РНҚ полимераза ІІІ

Барлық 3 типтегі промоторларға ортақ механизм – басқа ДНҚ байланыстырушы белсендіруші
Барлық 3 типтегі промоторларға ортақ механизм – басқа ДНҚ байланыстырушы белсендіруші
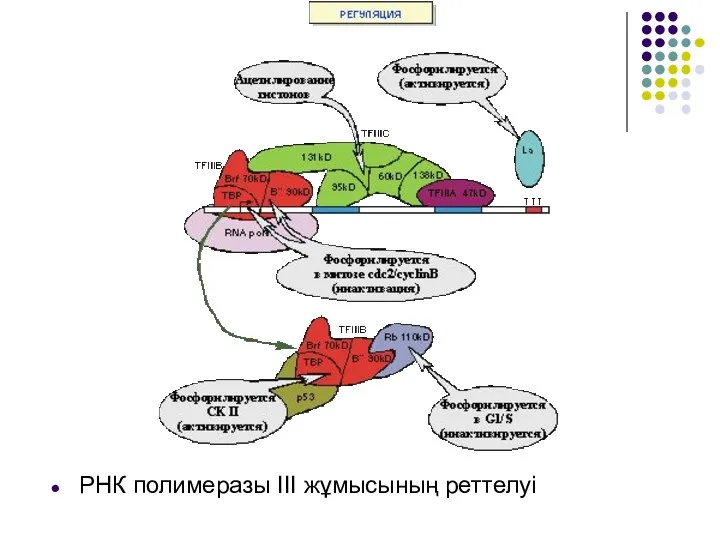
РНК полимеразы III жұмысының реттелуі
РНК полимеразы III жұмысының реттелуі

ТВІІІВ компоненттері тікелей фосфорилдену көмегімен де, репрессор ақуыз Rb және p53
ТВІІІВ компоненттері тікелей фосфорилдену көмегімен де, репрессор ақуыз Rb және p53

РНҚ полимераза ІІ РНҚның мРНҚ және кіші РНҚның түрлі класстарын транскрипцилайды.
РНҚ полимераза ІІ РНҚның мРНҚ және кіші РНҚның түрлі класстарын транскрипцилайды.

РНК полимераза I және ІІІ сияқты соңғы транскрипция инициация стадиясы РНК
РНК полимераза I және ІІІ сияқты соңғы транскрипция инициация стадиясы РНК
РНК полимераза іі құрамы және бұл полмеразаның транскрипциялық комплексінің қосымша факторлары.
РНК полимераза іі құрамы және бұл полмеразаның транскрипциялық комплексінің қосымша факторлары.

Жазылған факторлардан басқа РНК полимераза ІІ транскрипция инициациясында басқа да көптегендері
Жазылған факторлардан басқа РНК полимераза ІІ транскрипция инициациясында басқа да көптегендері

Фосфорленбеген (инициация стадиясында) мен РНК полимераза С-сонды домені ақуызды комплекспен әрекеттеседі,
Фосфорленбеген (инициация стадиясында) мен РНК полимераза С-сонды домені ақуызды комплекспен әрекеттеседі,

РНК полимераза ІІ барлық белсенділігін реттеу фосфорленген оның С-соңды домені Ser2
РНК полимераза ІІ барлық белсенділігін реттеу фосфорленген оның С-соңды домені Ser2

Медиатор – бұл ақуызды комплекс, РНК полимераза ІІ С-соңды доменмен ассоцирленген.
Медиатор – бұл ақуызды комплекс, РНК полимераза ІІ С-соңды доменмен ассоцирленген.
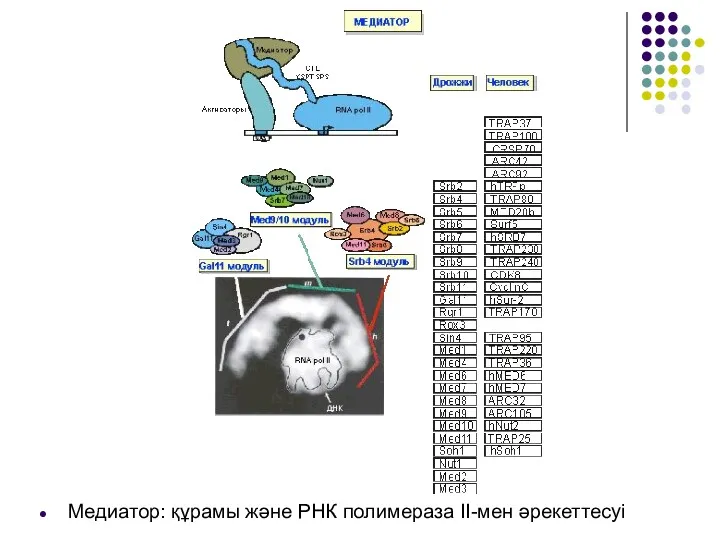
Медиатор: құрамы және РНК полимераза ІІ-мен әрекеттесуі
Медиатор: құрамы және РНК полимераза ІІ-мен әрекеттесуі

Медиаторда 3 модулді айыру мүмкін болды.
Gal11 модулі ДНК байланыстырушы активаторымен немесе
Медиаторда 3 модулді айыру мүмкін болды.
Gal11 модулі ДНК байланыстырушы активаторымен немесе

Жоғарғы эукариоттарда медиатордың гомологтары табылған болатын. Күтуге болатындай, ұқсас комплекстердің бірнешеуі
Жоғарғы эукариоттарда медиатордың гомологтары табылған болатын. Күтуге болатындай, ұқсас комплекстердің бірнешеуі

РНК полимераза ІІ-нің элонгационды күйінің басты маркері - фосфорленген СTD.
Элонгация
РНК полимераза ІІ-нің элонгационды күйінің басты маркері - фосфорленген СTD.
Элонгация
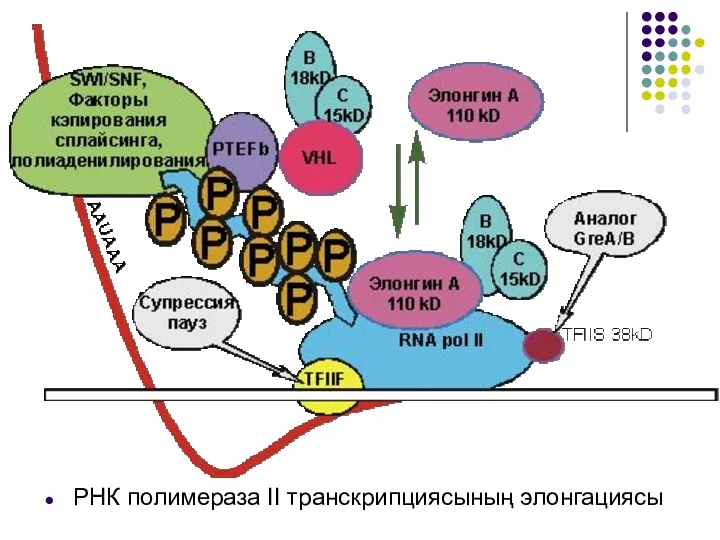
РНК полимераза ІІ транскрипциясының элонгациясы
РНК полимераза ІІ транскрипциясының элонгациясы

Элонгация процессінде РНК полимеразасы прокариоттың РНК полимеразасы сияқты мәселесімен кездесуі мүмкін:
Элонгация процессінде РНК полимеразасы прокариоттың РНК полимеразасы сияқты мәселесімен кездесуі мүмкін:

 Этические проблемы генной инженерии
Этические проблемы генной инженерии Виды прикусов
Виды прикусов Гастроэнтерологиядағы стандартты диагностикалау және емдеу әдістері
Гастроэнтерологиядағы стандартты диагностикалау және емдеу әдістері Шаншуға арналған дәрілік препараттар
Шаншуға арналған дәрілік препараттар Неврологический осмотр больного в коматозном состоянии
Неврологический осмотр больного в коматозном состоянии Бронхиальная астма
Бронхиальная астма ЭКСПЕРТИЗА ПРОФПРИГОДНОСТИ
ЭКСПЕРТИЗА ПРОФПРИГОДНОСТИ Реабилизация неврологических больных
Реабилизация неврологических больных Кандидоз слизистой оболочки полости рта
Кандидоз слизистой оболочки полости рта Endocrine system
Endocrine system Вступ до фармацевтичної біоетики
Вступ до фармацевтичної біоетики Патогенез лейкозов
Патогенез лейкозов Послеродовый период. Уход за родильницей
Послеродовый период. Уход за родильницей Тіндердің шығу тегі мен дамуының заңдылықтары. Тіндердің жіктелуі. Жүйе түзетін факторлар және оның тіндердің тұрақтылығын
Тіндердің шығу тегі мен дамуының заңдылықтары. Тіндердің жіктелуі. Жүйе түзетін факторлар және оның тіндердің тұрақтылығын Задержка роста (внутриутробного развития) плода
Задержка роста (внутриутробного развития) плода Определение центрального соотношения челюстей при полном отсутствии зубов
Определение центрального соотношения челюстей при полном отсутствии зубов Влияние алкоголя на организм человека
Влияние алкоголя на организм человека Правила обработки рук в соответствии с СанПиН. Методы и средства
Правила обработки рук в соответствии с СанПиН. Методы и средства Ультразвуковая диагностика заболеваний органов мошонки
Ультразвуковая диагностика заболеваний органов мошонки Дәрігерлік деонтология, семиология
Дәрігерлік деонтология, семиология Артериальная гипертензия
Артериальная гипертензия СПИД-ассоцированные инфекции. Хронические неинфекционные заболевания при ВИЧ-инфекции
СПИД-ассоцированные инфекции. Хронические неинфекционные заболевания при ВИЧ-инфекции Дитяче зубне протезування
Дитяче зубне протезування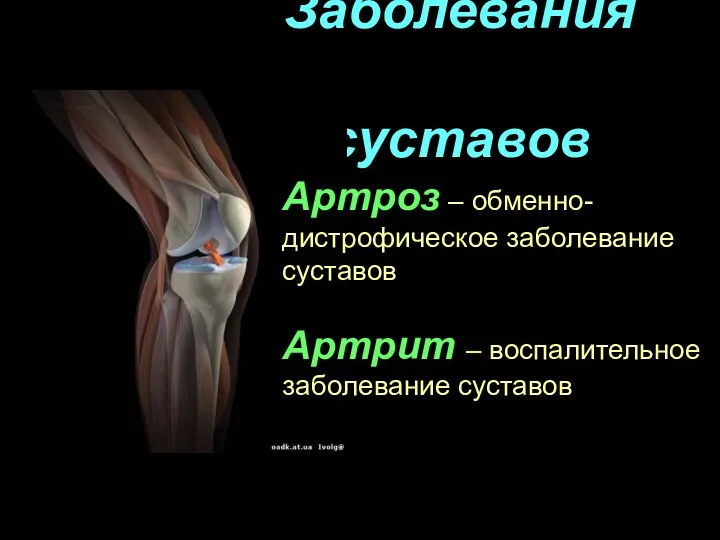 Заболевания суставов
Заболевания суставов Повреждение костей предплечья
Повреждение костей предплечья Растворы для обеспечения инфекционной безопасности
Растворы для обеспечения инфекционной безопасности Патофизиология сердечной недостаточности
Патофизиология сердечной недостаточности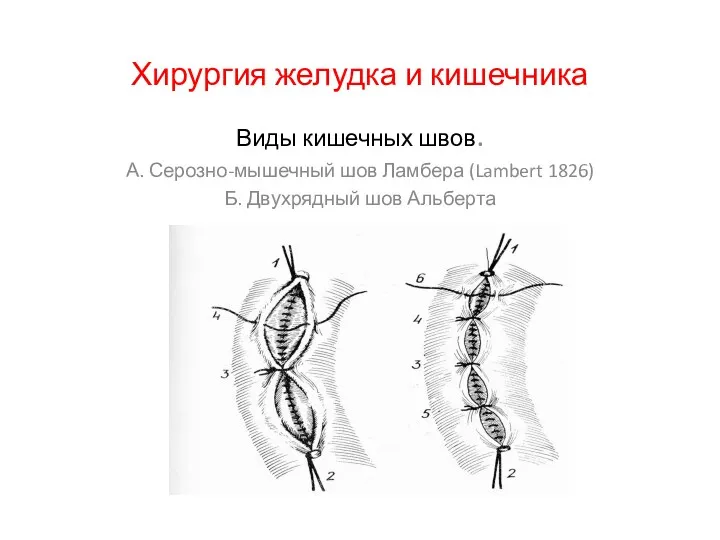 Хирургия желудка и кишечника
Хирургия желудка и кишечника