- Центральная нервная система. Торможение. Координация

Содержание
- 2. Гомосинаптическая модуляция частое (тетаническое) раздражение пресинаптической мембраны ↓ повышение [Са2+ ] в синаптическом окончании ↓ увеличение
- 3. Гомосинаптическая модуляция На постсинаптической мембране возрастает концентрация рецепторов, поэтому повышается чувствительность к медиатору и, как следствие,
- 4. Гетеросинаптическая модуляция Этапы: Выделение медиатора →↑ цАМФ → инактивация S- К+-каналов → удлинение фазы деполяризации ПД
- 5. ТОРМОЖЕНИЕ в ЦНС, КООРДИНАЦИОННЫЕ ПРИНЦИПЫ
- 6. Торможение и его функции ТОРМОЖЕНИЕ - это самостоятельный нервный процесс, вызываемый возбуждением и проявляющийся в подавлении
- 7. Виды торможения ЦЕНТРАЛЬНОЕ (Сеченовское) ВОЗВРАТНОЕ а) собственно возвратное б) латеральное в) медиальная зона РЕЦИПРОКНОЕ ТОРМОЖЕНИЕ ВСЛЕД
- 8. Тормозные нейроны В ЦНС имются нейроны со специфической функцией торможения: клетки Реншоу спинного мозга ; клетки
- 9. ТИПЫ ТОРМОЖЕНИЯ П Е Р В И Ч Н О Е: А) ЦЕНТРАЛЬНОЕ (СЕЧЕНОВСКОЕ) Б) ВОЗВРАТНОЕ
- 10. Торможение в ЦНС (опыт центрального торможения И.М. Сеченова)
- 11. Центральное торможение
- 12. Периферическое торможение по Гольцу
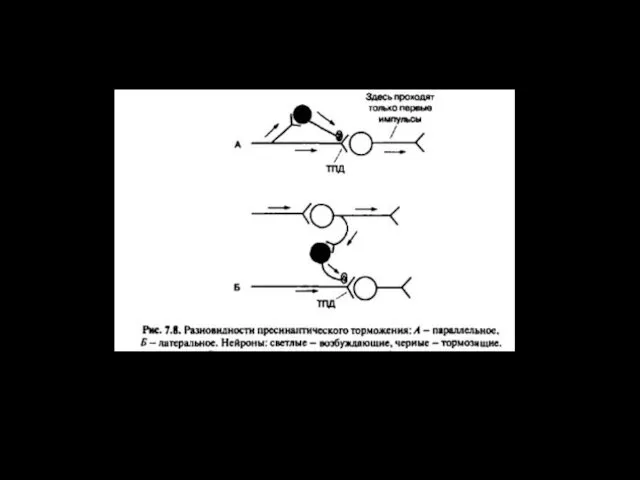
- 13. Окклюзия 1 2 3 4 5 6 А В При раздражении А возбуждаются 4 нейрона (1,2,3,4)
- 14. а) Собственно возвратное торможение (по Реншоу) б) Латеральное торможение ТОРМОЖЕНИЕ В ЦНС
- 15. Медиальная тормозная зона
- 16. Сопряженное (реципркное) торможение
- 17. Торможение вслед за возбуждением Суммация следовой гиперполяризации после серии возбуждений
- 18. Пессимальное торможение 1. При частом раздражении постсинаптические потенциалы суммируются, что приводит к стойкой деполяризации постсинаптической мембраны
- 19. Механизмы торможения: Пресинаптическое торможение Постсинаптическое торможение
- 20. Связан с воздействием тормозного медиатора (глицина) на постсинаптическую мембрану и развитием ТПСП, как следствие – снижением
- 21. постсинаптическое торможение
- 22. Тормозной постсинаптический потенциал ( ТПСП ) - 90 - 94 0 4 6 8 мв мс
- 23. СТРИХНИН – КОНКУРИРУЕТ С ГЛИЦИНОМ ЗА СВЯЗЫВАНИЕ С РЕЦЕПТОРАМИ НА ПОСТСИНАПТИЧЕСОЙ МЕМБРАНЕ СТОЛБНЯЧНЫЙ ТОКСИН - НАРУШАЕТ
- 24. МЕХАНИЗМ ПРЕСИНАПТИЧЕСКОГО ТОРМОЖЕНИЯ Возникает в области аксо-аксональных синапсов, на пресинаптических терминалях. Сопровождается медленной и длительной деполяризацией
- 25. ПРЕСИНАПТИЧЕСКОЕ ТОРМОЖЕНИЕ 1 - аксон тормозного нейрона; 2 - аксон возбуждающего нейрона; 3 - постсинаптическая мембрана
- 27. ГАМК-рецептор Агонист ГАМК-рецепторов – мусцимол . Антогонист ГАМК-рецепторов - бикукулин
- 28. Блокаторы и агонисты ГАМК-рецепторов Основным ядом, блокирующим пресинаптическое торможение является бикукулин. Вещества, усиливающие ингибиторный эффект ГАМК,
- 29. Растения семейства дымянковых Fumariaceae (хохлатка полая, хохлатка Горчакова, хохлатка Маршалла) содержат ядовитые алкалоиды группы бульбокапнина -
- 30. Антиноцицептивная система выделяет биологически активные эндогенные опиоидные вещества – это «внутренние наркотики». Они называются эндорфины, энкефалины,
- 32. Координация нервных процессов в ЦНС
- 33. Функции координированной деятельности: 1) обеспечивает четкое выполнение определенных функций, рефлексов; 2) обеспечивает последовательное включение в работу
- 34. ПРИНЦИПЫ КООРДИНАЦИИ РЕФЛЕКТОРНОЙ ДЕЯТЕЛЬНОСТИ В ЦНС 1. КОНВЕРГЕНЦИИ ИЛИ ОБЩЕГО КОНЕЧНОГО ПУТИ (по Шеррингтону) 2. ДИВЕРГЕНТНОСТИ
- 35. ПРИНЦИП ОБЩЕГО КОНЕЧНОГО ПУТИ КОРА ПОДКОРКА СТВОЛ СПИННОЙ МОЗГ
- 36. Принцип общего конечного пути (по Шеррингтону) в спинном мозге:
- 37. ДИВЕРГЕНЦИЯ НЕРВНЫХ ИМПУЛЬСОВ В ЦНС
- 40. Обратные связи Положительные Отрицательные импульсы с периферии, импульсы с периферии, возникающие в возникающие в результате какой-либо
- 41. РЕФЛЕКТОРНАЯ ДУГА И РЕФЛЕКТОРНОЕ КОЛЬЦО
- 42. ПРИНЦИП ОБРАТНОЙ АФФЕРЕНТАЦИИ
- 43. ОПРЕДЕЛЕНИЕ ДОМИНАНТЫ ( ПО А.А.Ухтомскому, 1931) Доминанта - временно господствующий рефлекс или поведенческий акт, которым трансформируется
- 44. Собака – с переполненым кишечником Укол конечности Акт дефекации ? Ухтомский А.А. в 1902-1903 г.г. думает
- 45. Основные cвойства доминанты ( по А.А.Ухтомскому) 1. Повышенная возбудимость доминантного центра 2. Способность к суммации (подкреплять
- 46. Виды доминант Внутренние доминанты – ведущую роль играет гормональный фон. Внешние доминанты – ведущую роль играет
- 47. Причины прекращения существования доминанты Достижение результата Появление новой, более значимой доминанты Запредельное торможение
- 48. ПРИНЦИП СУБОРДИНАЦИИ НЕРВНЫХ ЦЕНТРОВ
- 50. ПРИНЦИП РЕЦИПРОКНОСТИ ( СОПРЯЖЕННОГО ТОРМОЖЕНИЯ ) + + - - +
- 51. Динамика основных нервных процессов: Процессы иррадиации и концентрации составляют основу индукционных отношений в ЦНС. Индукция –
- 52. Возникновение торможения при последовательной индукции называется - отрицательной последовательной индукцией. Возникновение возбуждения при последовательной индукции называется
- 53. 9. Принцип индукции Индукция ограничивает распространение нервных процессов и обеспечивает концентрацию. одновременная отрицательная индукция одновременная положительная
- 54. последовательная положительная индукция
- 55. От степени развития индукции зависит подвижность нервных процессов, возможность выполнения движений скоростного характера, требующих быстрой смены
- 56. Частная ЦНС СПИННОЙ МОЗГ. ФИЗИОЛОГИЯ СТВОЛА МОЗГА.
- 57. Сегменты спинного мозга 8 шейных (C1 - C8) 12 грудных (Th1 - Th12) 5 поясничных (L1
- 58. ПРИНЦИП МЕТАМЕРИИ
- 59. Принципы иннервации спинного мозга: Сегментарный принцип Морфологических границ между сегментами нет, поэтому деление является – функциональным.
- 60. Закон Белла - Мажанди Вентральные (передние) корешки содержат эфферентные двигательные (выходящие) волокна, а дорсальные (задние) корешки
- 61. Нейроны спинного мозга (13 млн.): 1. Двигательные или мотонейроны (3%): - альфа-мотонейроны - гамма-мотонейроны 2. Вставочные
- 62. Нейроны. В сером веществе спинного мозга находятся тела двигательных, вставочных и вегетативных нейронов. 1) Мотонейроны. Различают
- 63. По эффекторному действию нейроны разделяются на: Возбуждающие (3%) Тормозные
- 64. ФУНКЦИИ СПИННОГО МОЗГА: АФФЕРЕНТНАЯ ПРОВОДНИКОВАЯ РЕФЛЕКТОРНАЯ
- 65. Основные рефлексы спинного мозга Рефлексы растяжения (миотатические) - в основном разгибательные - рефлексы позы, толчковые (прыжок,
- 66. Рефлекторные функции спинного мозга Регуляция фазной активности: Сгибательные рефлексы Локомоции (автоматизмы) Регуляция тонуса: Миотатические рефлексы Позно-тонические
- 67. Строение мышечных веретен и сухожильных органов. А. Схема мышечного веретена. Б. Рисунок сухожильного органа Гольджи В.
- 68. Миотатические рефлексы (сухожильные, Т-рефлексы) – рефлексы на растяжение мышцы. Вызываются ударом по сухожилию мышцы. При таком
- 69. Т (tendon – сухожилие)-рефлекс Рефлекторная дуга моносинаптического рефлекса растяжения
- 70. Дуги рефлекса растяжения и реципрокного торможения мышц–антагонистов С–мотонейроны сгибателей коленного сустава; Р–мотонейроны разгибателей коленного сустава.
- 71. Облегчение Т–рефлексов. Если коленный и другие Т–рефлексы нижней конечности ослаблены, их можно усилить, если больной, сцепив
- 72. ТОНИЧЕСКАЯ ФУНКЦИЯ СПИННОГО МОЗГА. ГАММА-МОТОРНАЯ ПЕТЛЯ
- 73. Коактивация альфа– и гамма–мотонейронов при движениях. α– и γ–мотонейроны возбуждаются одновременно; однако начало импульсации афферентов мышечных
- 74. Функции сухожильных органов. Рефлекторная дуга сухожильных органов служит для поддержания постоянства напряжения мышцы. У каждой мышцы
- 75. Схема состояния и характера импульсации мышечных веретен и сухожильных органов Гольджи А - в покое, Б
- 76. Внутрисегментарные связи волокон Ib от сухожильных органов
- 77. Сгибательные рефлексы Осуществляются при раздражении кожных рецепторов, рецепторов суставов, рецепторов давления и боли скелетных мышц. Например,
- 79. СГИБАТЕЛЬНЫЙ РЕФЛЕКС Внутрисегментарные связи афферентного волокна от болевого рецептора кожи стопы Красным показаны афферентное волокно группы
- 80. Возвратное торможение Реншоу и пресинаптическое торможение в спинальных двигательных рефлекторных дугах Торможение клеток Реншоу служит для
- 81. Сухожильные (миотатические) рефлексы спинного мозга
- 82. Сухожильные (миотатические) рефлексы спинного мозга
- 84. Скачать презентацию

Гомосинаптическая
модуляция
частое (тетаническое) раздражение пресинаптической мембраны
↓
повышение [Са2+ ] в
Гомосинаптическая
модуляция
частое (тетаническое) раздражение пресинаптической мембраны
↓
повышение [Са2+ ] в

Гомосинаптическая
модуляция
На постсинаптической мембране возрастает концентрация рецепторов, поэтому повышается чувствительность к
Гомосинаптическая
модуляция
На постсинаптической мембране возрастает концентрация рецепторов, поэтому повышается чувствительность к
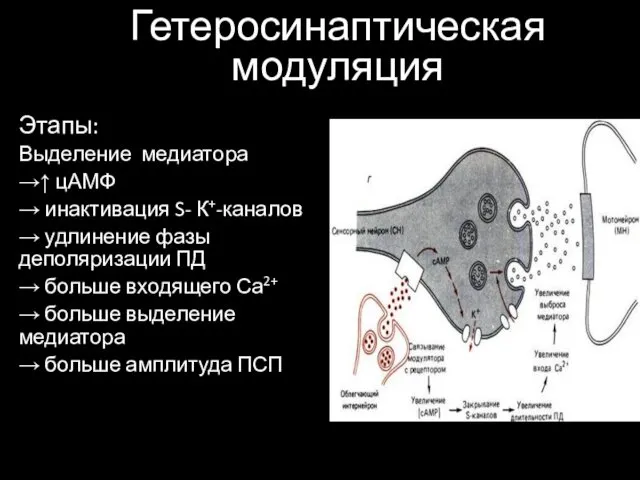
Гетеросинаптическая модуляция
Этапы:
Выделение медиатора
→↑ цАМФ
→ инактивация S- К+-каналов
→ удлинение фазы
Гетеросинаптическая модуляция
Этапы:
Выделение медиатора
→↑ цАМФ
→ инактивация S- К+-каналов
→ удлинение фазы
ТОРМОЖЕНИЕ в ЦНС,
КООРДИНАЦИОННЫЕ ПРИНЦИПЫ
ТОРМОЖЕНИЕ в ЦНС,
КООРДИНАЦИОННЫЕ ПРИНЦИПЫ

Торможение и его функции
ТОРМОЖЕНИЕ - это самостоятельный нервный процесс, вызываемый возбуждением
Торможение и его функции
ТОРМОЖЕНИЕ - это самостоятельный нервный процесс, вызываемый возбуждением

Виды торможения
ЦЕНТРАЛЬНОЕ (Сеченовское)
ВОЗВРАТНОЕ
а) собственно возвратное
б) латеральное
в) медиальная
Виды торможения
ЦЕНТРАЛЬНОЕ (Сеченовское)
ВОЗВРАТНОЕ
а) собственно возвратное
б) латеральное
в) медиальная

Тормозные нейроны
В ЦНС имются нейроны со специфической функцией торможения: клетки Реншоу
Тормозные нейроны
В ЦНС имются нейроны со специфической функцией торможения: клетки Реншоу

ТИПЫ ТОРМОЖЕНИЯ
П Е Р В И Ч Н О Е:
А)
ТИПЫ ТОРМОЖЕНИЯ
П Е Р В И Ч Н О Е:
А)
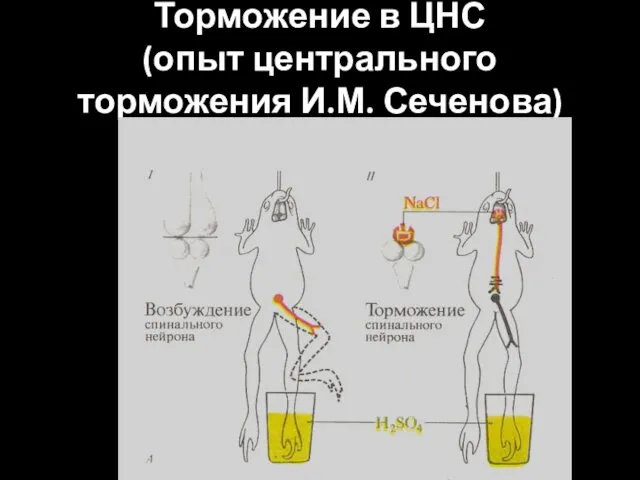
Торможение в ЦНС
(опыт центрального торможения И.М. Сеченова)
Торможение в ЦНС
(опыт центрального торможения И.М. Сеченова)
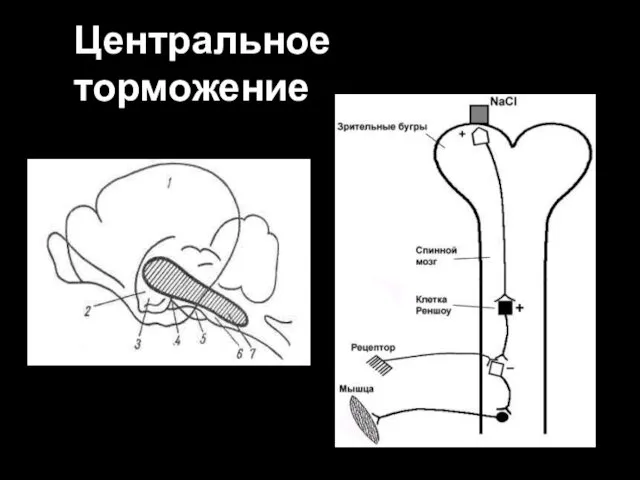
Центральное торможение
Центральное торможение

Периферическое торможение по Гольцу
Периферическое торможение по Гольцу
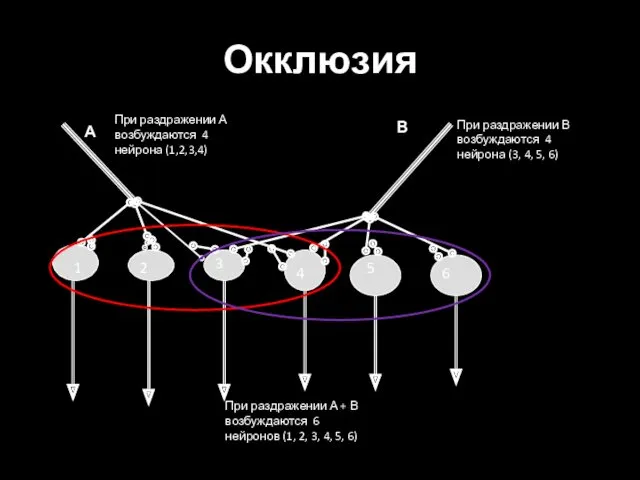
Окклюзия
1
2
3
4
5
6
А
В
При раздражении А возбуждаются 4 нейрона (1,2,3,4)
При раздражении В возбуждаются 4
Окклюзия
1
2
3
4
5
6
А
В
При раздражении А возбуждаются 4 нейрона (1,2,3,4)
При раздражении В возбуждаются 4
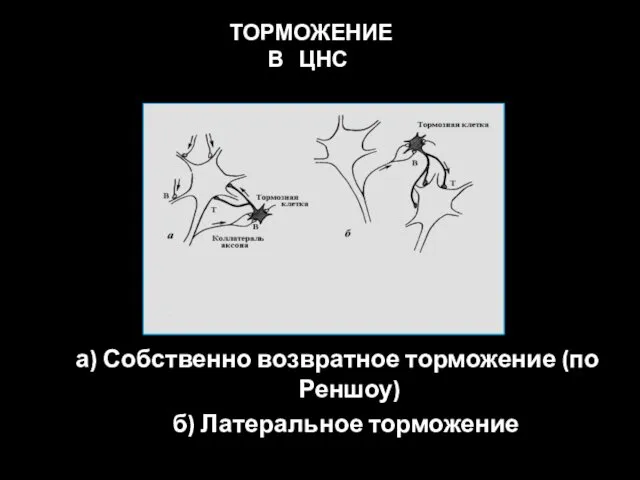
а) Собственно возвратное торможение (по Реншоу)
б) Латеральное торможение
ТОРМОЖЕНИЕ В
а) Собственно возвратное торможение (по Реншоу)
б) Латеральное торможение
ТОРМОЖЕНИЕ В

Медиальная тормозная зона
Медиальная тормозная зона
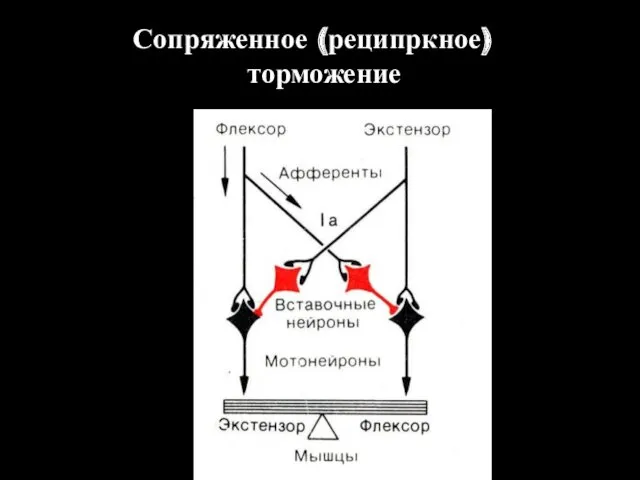
Сопряженное (реципркное) торможение
Сопряженное (реципркное) торможение
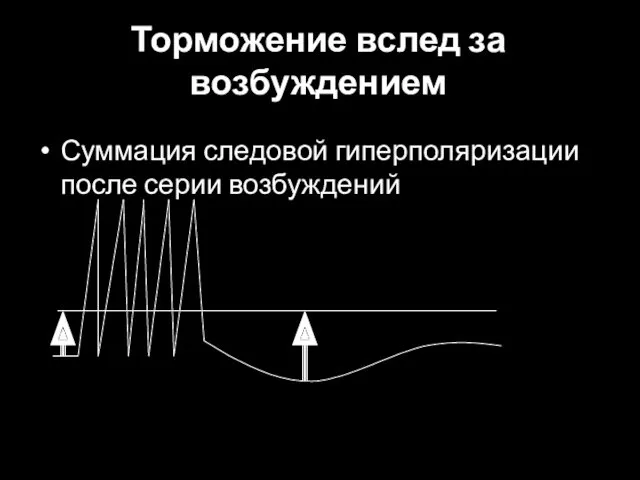
Торможение вслед за возбуждением
Суммация следовой гиперполяризации после серии возбуждений
Торможение вслед за возбуждением
Суммация следовой гиперполяризации после серии возбуждений

Пессимальное торможение
1. При частом раздражении постсинаптические потенциалы суммируются, что приводит к
Пессимальное торможение
1. При частом раздражении постсинаптические потенциалы суммируются, что приводит к

Механизмы торможения:
Пресинаптическое торможение
Постсинаптическое торможение
Механизмы торможения:
Пресинаптическое торможение
Постсинаптическое торможение

Связан с воздействием тормозного медиатора (глицина) на постсинаптическую мембрану и развитием
Связан с воздействием тормозного медиатора (глицина) на постсинаптическую мембрану и развитием
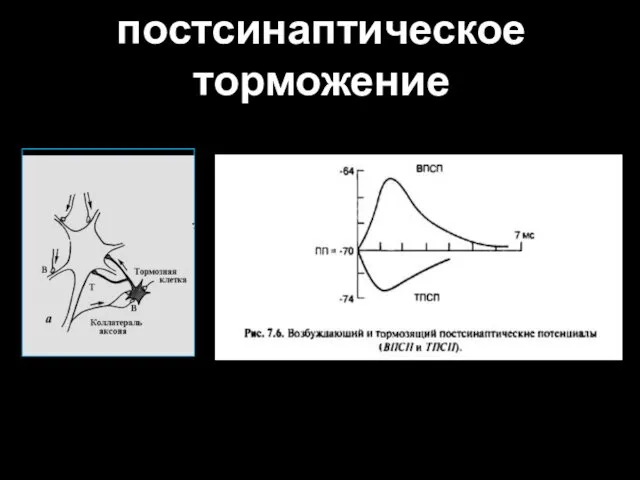
постсинаптическое торможение
постсинаптическое торможение
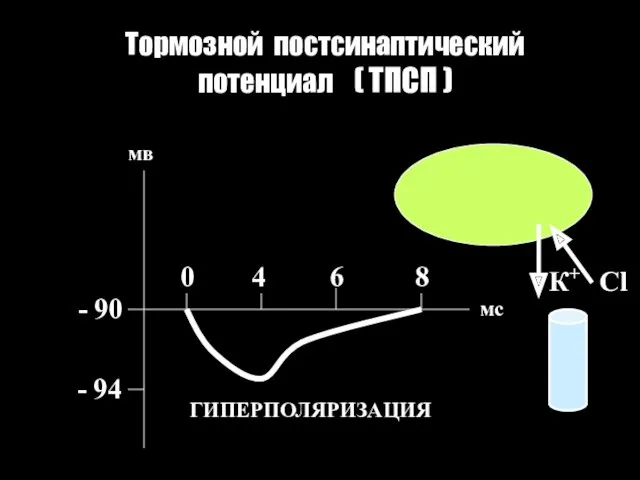
Тормозной постсинаптический потенциал ( ТПСП )
- 90
- 94
0 4
Тормозной постсинаптический потенциал ( ТПСП )
- 90
- 94
0 4

СТРИХНИН – КОНКУРИРУЕТ С ГЛИЦИНОМ ЗА СВЯЗЫВАНИЕ С РЕЦЕПТОРАМИ НА ПОСТСИНАПТИЧЕСОЙ

МЕХАНИЗМ ПРЕСИНАПТИЧЕСКОГО ТОРМОЖЕНИЯ
Возникает в области аксо-аксональных синапсов, на пресинаптических терминалях. Сопровождается
МЕХАНИЗМ ПРЕСИНАПТИЧЕСКОГО ТОРМОЖЕНИЯ
Возникает в области аксо-аксональных синапсов, на пресинаптических терминалях. Сопровождается
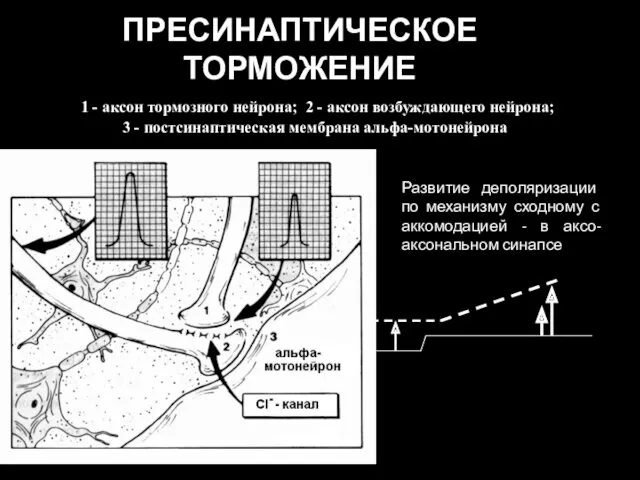
ПРЕСИНАПТИЧЕСКОЕ ТОРМОЖЕНИЕ
1 - аксон тормозного нейрона; 2 - аксон возбуждающего
ПРЕСИНАПТИЧЕСКОЕ ТОРМОЖЕНИЕ
1 - аксон тормозного нейрона; 2 - аксон возбуждающего
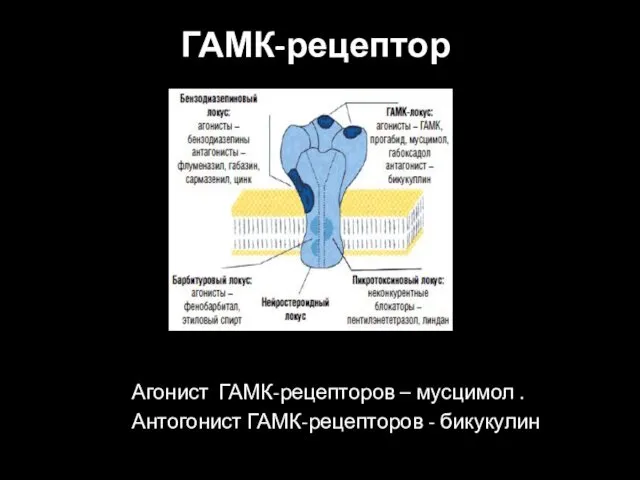
ГАМК-рецептор
Агонист ГАМК-рецепторов – мусцимол .
Антогонист ГАМК-рецепторов - бикукулин
ГАМК-рецептор
Агонист ГАМК-рецепторов – мусцимол .
Антогонист ГАМК-рецепторов - бикукулин

Блокаторы и агонисты ГАМК-рецепторов
Основным ядом, блокирующим пресинаптическое торможение является бикукулин.
Вещества, усиливающие
Блокаторы и агонисты ГАМК-рецепторов
Основным ядом, блокирующим пресинаптическое торможение является бикукулин.
Вещества, усиливающие

Растения семейства дымянковых Fumariaceae (хохлатка полая, хохлатка Горчакова, хохлатка Маршалла) содержат
Растения семейства дымянковых Fumariaceae (хохлатка полая, хохлатка Горчакова, хохлатка Маршалла) содержат
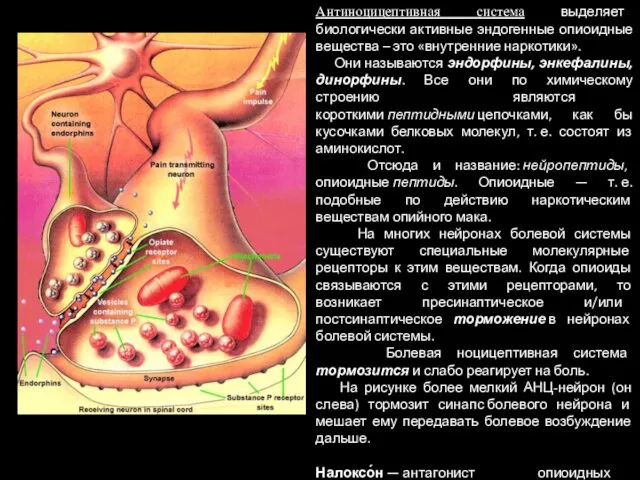
Антиноцицептивная система выделяет биологически активные эндогенные опиоидные вещества – это «внутренние
Антиноцицептивная система выделяет биологически активные эндогенные опиоидные вещества – это «внутренние

Координация нервных процессов в ЦНС
Координация нервных процессов в ЦНС

Функции координированной деятельности:
1) обеспечивает четкое выполнение определенных функций, рефлексов;
2) обеспечивает последовательное включение в
Функции координированной деятельности:
1) обеспечивает четкое выполнение определенных функций, рефлексов;
2) обеспечивает последовательное включение в

ПРИНЦИПЫ КООРДИНАЦИИ РЕФЛЕКТОРНОЙ ДЕЯТЕЛЬНОСТИ В ЦНС
1. КОНВЕРГЕНЦИИ ИЛИ ОБЩЕГО КОНЕЧНОГО ПУТИ
ПРИНЦИПЫ КООРДИНАЦИИ РЕФЛЕКТОРНОЙ ДЕЯТЕЛЬНОСТИ В ЦНС
1. КОНВЕРГЕНЦИИ ИЛИ ОБЩЕГО КОНЕЧНОГО ПУТИ
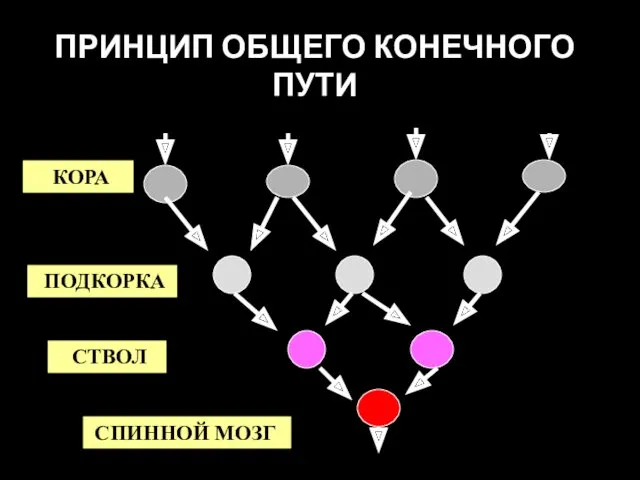
ПРИНЦИП ОБЩЕГО КОНЕЧНОГО ПУТИ
КОРА
ПОДКОРКА
СТВОЛ
СПИННОЙ МОЗГ
ПРИНЦИП ОБЩЕГО КОНЕЧНОГО ПУТИ
КОРА
ПОДКОРКА
СТВОЛ
СПИННОЙ МОЗГ
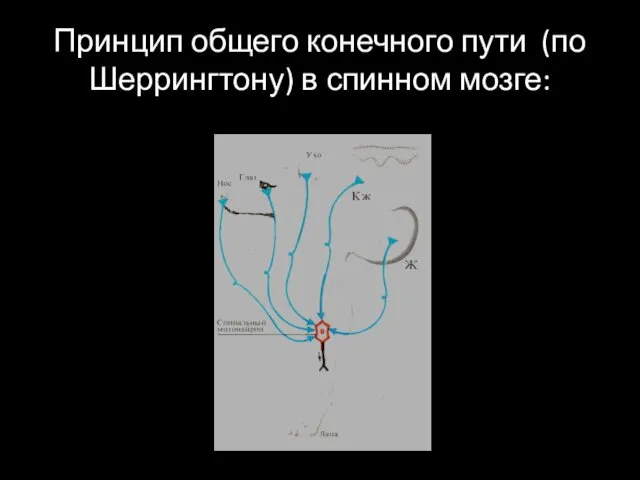
Принцип общего конечного пути (по Шеррингтону) в спинном мозге:
Принцип общего конечного пути (по Шеррингтону) в спинном мозге:

ДИВЕРГЕНЦИЯ НЕРВНЫХ ИМПУЛЬСОВ В ЦНС
ДИВЕРГЕНЦИЯ НЕРВНЫХ ИМПУЛЬСОВ В ЦНС

Обратные связи
Положительные Отрицательные
импульсы с периферии, импульсы с периферии,
возникающие в возникающие
Обратные связи
Положительные Отрицательные
импульсы с периферии, импульсы с периферии,
возникающие в возникающие

РЕФЛЕКТОРНАЯ ДУГА И РЕФЛЕКТОРНОЕ КОЛЬЦО
РЕФЛЕКТОРНАЯ ДУГА И РЕФЛЕКТОРНОЕ КОЛЬЦО
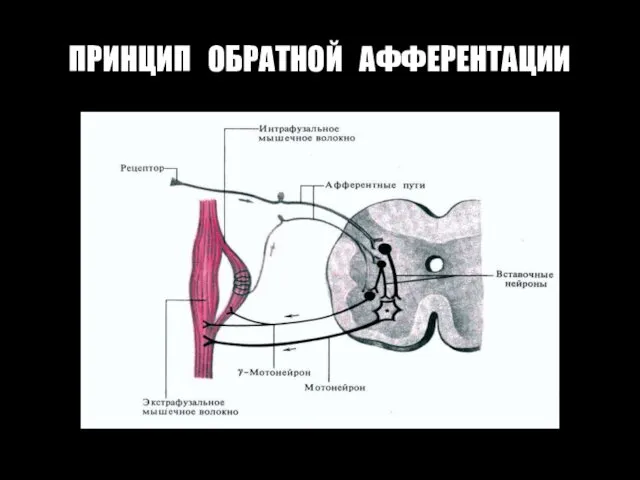
ПРИНЦИП ОБРАТНОЙ АФФЕРЕНТАЦИИ
ПРИНЦИП ОБРАТНОЙ АФФЕРЕНТАЦИИ

ОПРЕДЕЛЕНИЕ ДОМИНАНТЫ
( ПО А.А.Ухтомскому, 1931)
Доминанта - временно господствующий рефлекс или поведенческий
ОПРЕДЕЛЕНИЕ ДОМИНАНТЫ
( ПО А.А.Ухтомскому, 1931)
Доминанта - временно господствующий рефлекс или поведенческий
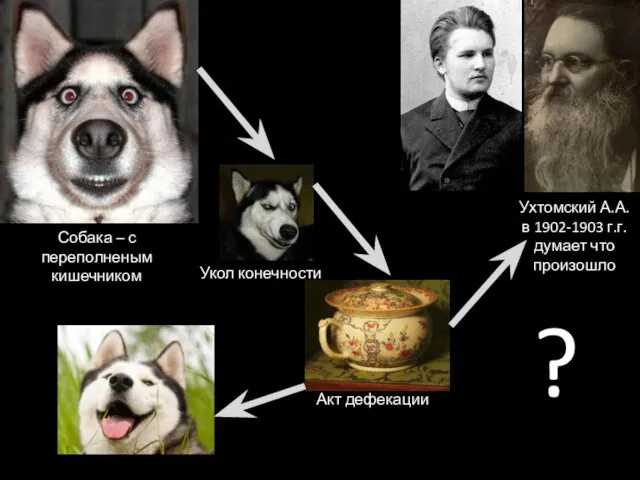
Собака – с переполненым кишечником
Укол конечности
Акт дефекации
?
Ухтомский А.А. в 1902-1903 г.г.
Собака – с переполненым кишечником
Укол конечности
Акт дефекации
?
Ухтомский А.А. в 1902-1903 г.г.

Основные cвойства доминанты
( по А.А.Ухтомскому)
1. Повышенная возбудимость доминантного центра
2. Способность к
Основные cвойства доминанты
( по А.А.Ухтомскому)
1. Повышенная возбудимость доминантного центра
2. Способность к

Виды доминант
Внутренние доминанты – ведущую роль играет гормональный фон.
Внешние доминанты –
Виды доминант
Внутренние доминанты – ведущую роль играет гормональный фон.
Внешние доминанты –

Причины прекращения существования доминанты
Достижение результата
Появление новой, более значимой доминанты
Запредельное торможение
Причины прекращения существования доминанты
Достижение результата
Появление новой, более значимой доминанты
Запредельное торможение
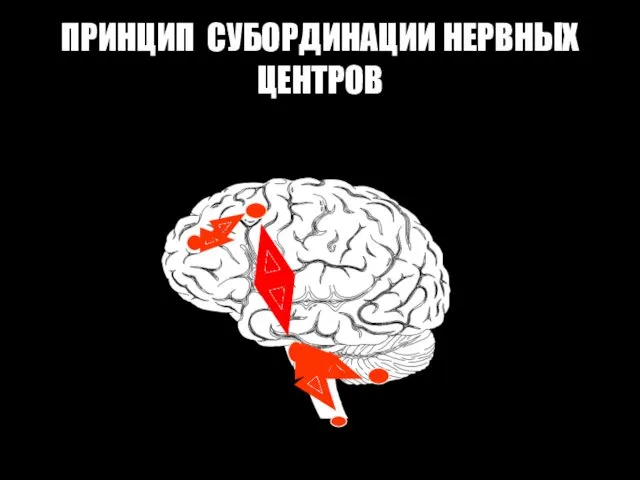
ПРИНЦИП СУБОРДИНАЦИИ НЕРВНЫХ ЦЕНТРОВ
ПРИНЦИП СУБОРДИНАЦИИ НЕРВНЫХ ЦЕНТРОВ
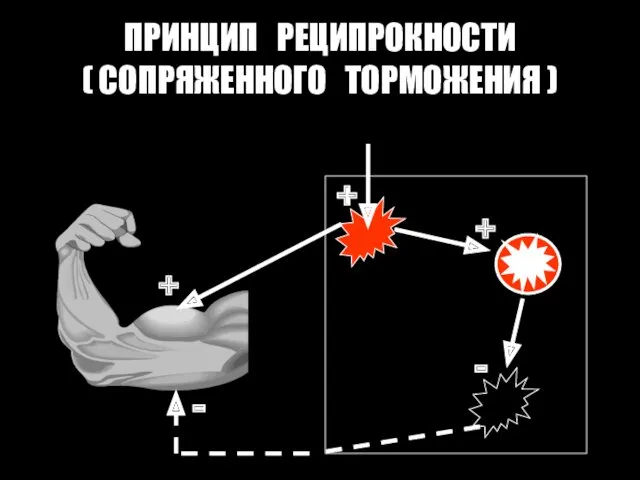
ПРИНЦИП РЕЦИПРОКНОСТИ
( СОПРЯЖЕННОГО ТОРМОЖЕНИЯ )
+
+
-
-
+
ПРИНЦИП РЕЦИПРОКНОСТИ
( СОПРЯЖЕННОГО ТОРМОЖЕНИЯ )
+
+
-
-
+

Динамика основных нервных процессов:
Процессы иррадиации и концентрации составляют основу индукционных отношений
Динамика основных нервных процессов:
Процессы иррадиации и концентрации составляют основу индукционных отношений

Возникновение торможения при последовательной индукции называется -
отрицательной последовательной индукцией.
Возникновение возбуждения при
Возникновение торможения при последовательной индукции называется -
отрицательной последовательной индукцией.
Возникновение возбуждения при
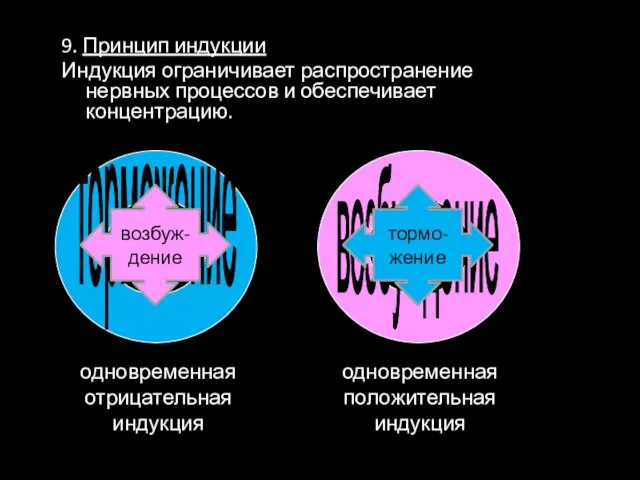
9. Принцип индукции
Индукция ограничивает распространение нервных процессов и обеспечивает концентрацию.
одновременная
отрицательная
9. Принцип индукции
Индукция ограничивает распространение нервных процессов и обеспечивает концентрацию.
одновременная
отрицательная
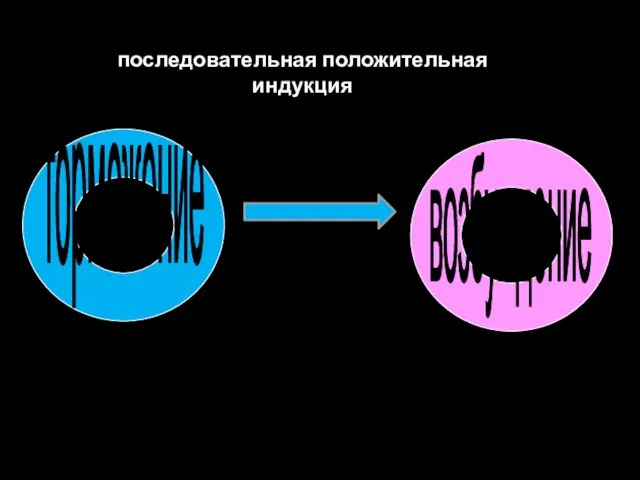
последовательная положительная индукция
последовательная положительная индукция
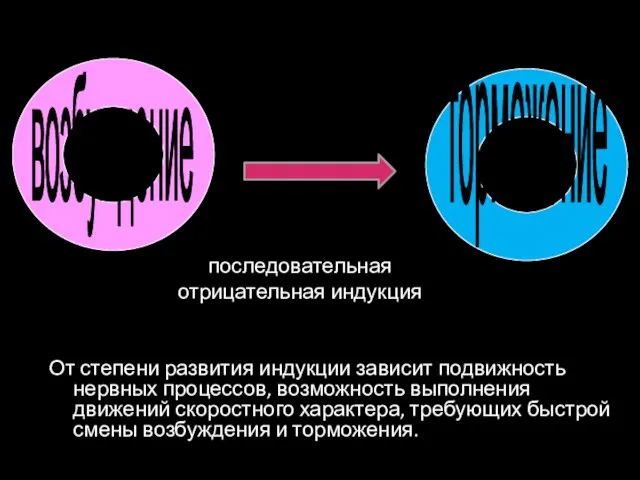
От степени развития индукции зависит подвижность нервных процессов, возможность выполнения движений
От степени развития индукции зависит подвижность нервных процессов, возможность выполнения движений

Частная ЦНС
СПИННОЙ МОЗГ.
ФИЗИОЛОГИЯ
СТВОЛА
МОЗГА.
Частная ЦНС
СПИННОЙ МОЗГ.
ФИЗИОЛОГИЯ
СТВОЛА
МОЗГА.
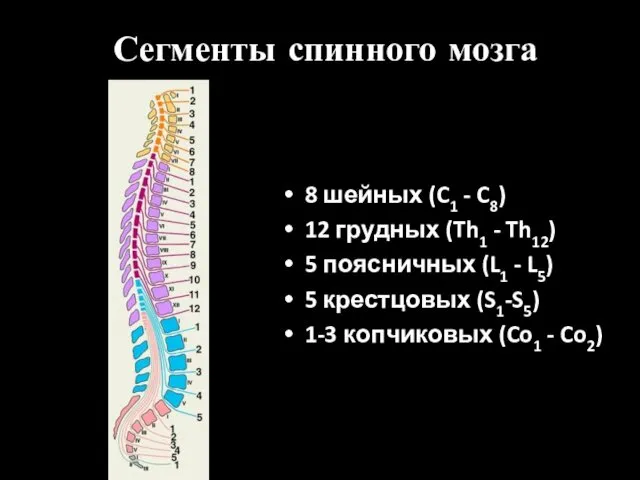
Сегменты спинного мозга
8 шейных (C1 - C8)
12 грудных (Th1 - Th12)
5
Сегменты спинного мозга
8 шейных (C1 - C8)
12 грудных (Th1 - Th12)
5
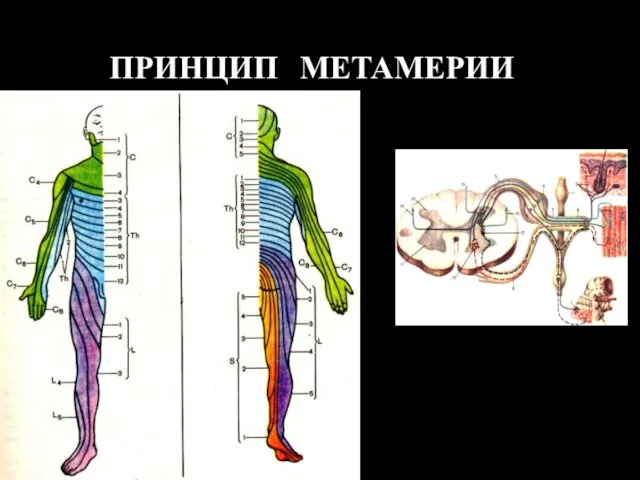
ПРИНЦИП МЕТАМЕРИИ
ПРИНЦИП МЕТАМЕРИИ

Принципы иннервации спинного мозга:
Сегментарный принцип
Морфологических границ между сегментами нет, поэтому
Принципы иннервации спинного мозга:
Сегментарный принцип
Морфологических границ между сегментами нет, поэтому

Закон Белла - Мажанди
Вентральные (передние) корешки содержат эфферентные двигательные (выходящие) волокна,
Закон Белла - Мажанди
Вентральные (передние) корешки содержат эфферентные двигательные (выходящие) волокна,

Нейроны спинного мозга
(13 млн.):
1. Двигательные или мотонейроны (3%):
- альфа-мотонейроны
Нейроны спинного мозга
(13 млн.):
1. Двигательные или мотонейроны (3%):
- альфа-мотонейроны

Нейроны. В сером веществе спинного мозга находятся тела двигательных, вставочных и
Нейроны. В сером веществе спинного мозга находятся тела двигательных, вставочных и

По эффекторному действию нейроны разделяются на:
Возбуждающие (3%)
Тормозные
По эффекторному действию нейроны разделяются на:
Возбуждающие (3%)
Тормозные

ФУНКЦИИ СПИННОГО МОЗГА:
АФФЕРЕНТНАЯ
ПРОВОДНИКОВАЯ
РЕФЛЕКТОРНАЯ
ФУНКЦИИ СПИННОГО МОЗГА:
АФФЕРЕНТНАЯ
ПРОВОДНИКОВАЯ
РЕФЛЕКТОРНАЯ

Основные рефлексы спинного мозга
Рефлексы растяжения (миотатические) - в основном разгибательные -
Основные рефлексы спинного мозга
Рефлексы растяжения (миотатические) - в основном разгибательные -

Рефлекторные функции спинного мозга
Регуляция фазной активности:
Сгибательные рефлексы
Локомоции (автоматизмы)
Регуляция тонуса:
Миотатические рефлексы
Позно-тонические рефлексы
Рефлекторные функции спинного мозга
Регуляция фазной активности:
Сгибательные рефлексы
Локомоции (автоматизмы)
Регуляция тонуса:
Миотатические рефлексы
Позно-тонические рефлексы
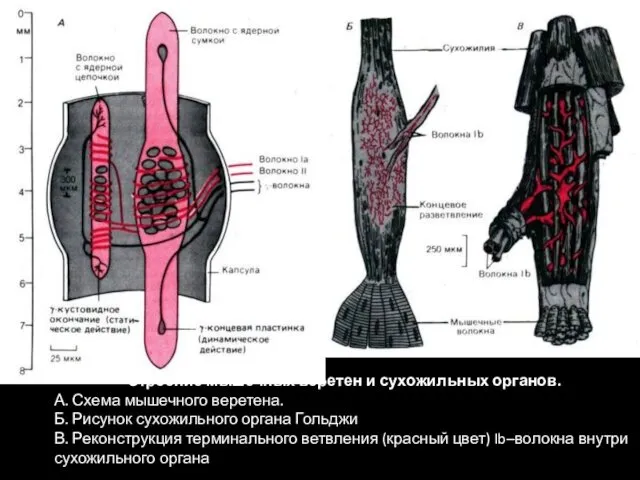
Строение мышечных веретен и сухожильных органов.
А. Схема мышечного веретена.
Б. Рисунок
Строение мышечных веретен и сухожильных органов.
А. Схема мышечного веретена.
Б. Рисунок
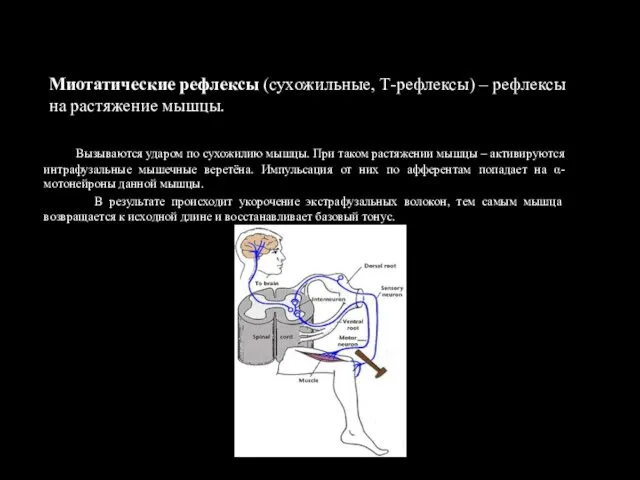
Миотатические рефлексы (сухожильные, Т-рефлексы) – рефлексы на растяжение мышцы.
Вызываются
Миотатические рефлексы (сухожильные, Т-рефлексы) – рефлексы на растяжение мышцы.
Вызываются
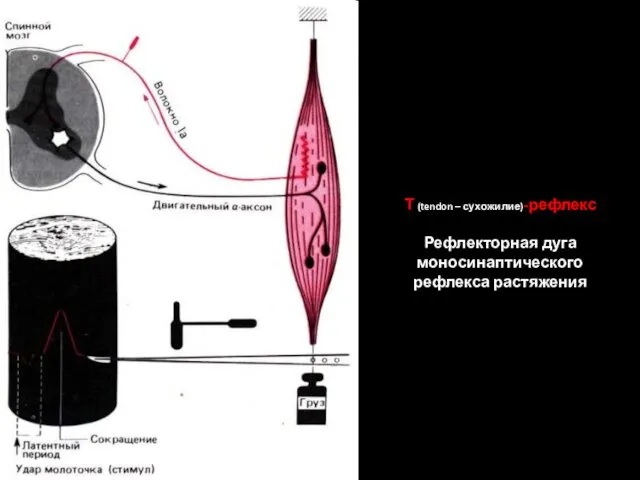
Т (tendon – сухожилие)-рефлекс
Рефлекторная дуга моносинаптического рефлекса растяжения
Т (tendon – сухожилие)-рефлекс
Рефлекторная дуга моносинаптического рефлекса растяжения
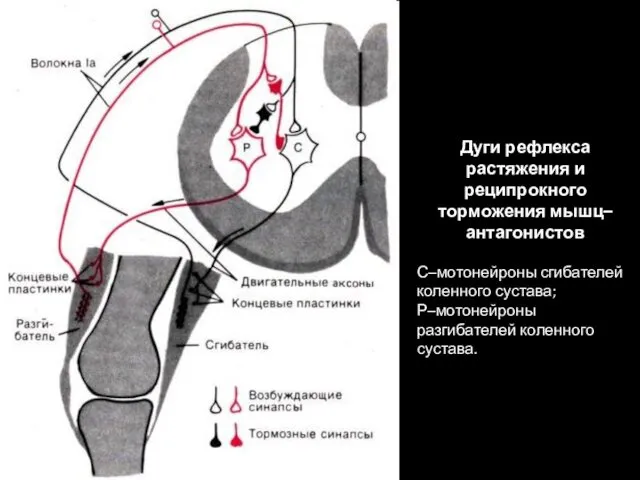
Дуги рефлекса растяжения и реципрокного торможения мышц–антагонистов
С–мотонейроны сгибателей коленного сустава;
Р–мотонейроны
Дуги рефлекса растяжения и реципрокного торможения мышц–антагонистов
С–мотонейроны сгибателей коленного сустава;
Р–мотонейроны

Облегчение Т–рефлексов.
Если коленный и другие Т–рефлексы нижней конечности ослаблены, их можно
Облегчение Т–рефлексов.
Если коленный и другие Т–рефлексы нижней конечности ослаблены, их можно
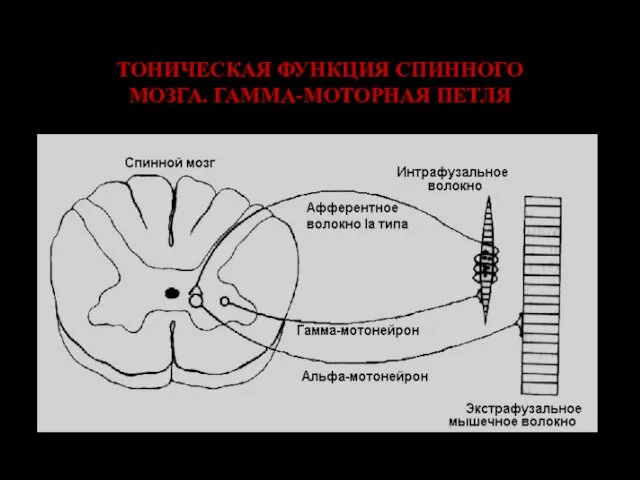
ТОНИЧЕСКАЯ ФУНКЦИЯ СПИННОГО МОЗГА. ГАММА-МОТОРНАЯ ПЕТЛЯ
ТОНИЧЕСКАЯ ФУНКЦИЯ СПИННОГО МОЗГА. ГАММА-МОТОРНАЯ ПЕТЛЯ

Коактивация альфа– и гамма–мотонейронов при движениях.
α– и γ–мотонейроны возбуждаются одновременно; однако начало импульсации афферентов мышечных веретен отстает
Коактивация альфа– и гамма–мотонейронов при движениях.
α– и γ–мотонейроны возбуждаются одновременно; однако начало импульсации афферентов мышечных веретен отстает

Функции сухожильных органов.
Рефлекторная дуга сухожильных органов служит для поддержания постоянства напряжения мышцы.
У
Функции сухожильных органов.
Рефлекторная дуга сухожильных органов служит для поддержания постоянства напряжения мышцы.
У
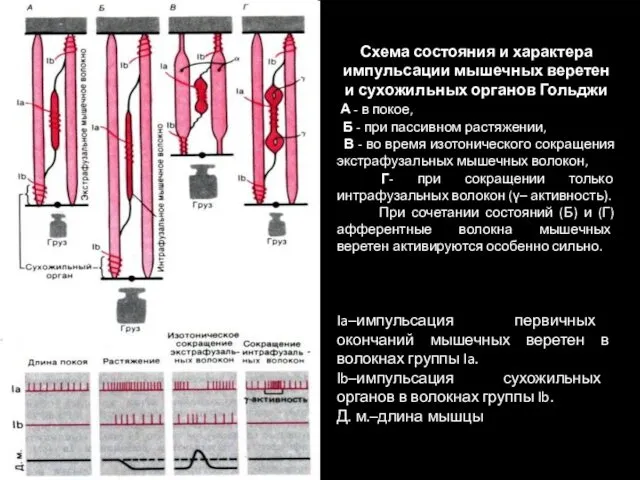
Схема состояния и характера импульсации мышечных веретен и сухожильных органов Гольджи
Схема состояния и характера импульсации мышечных веретен и сухожильных органов Гольджи
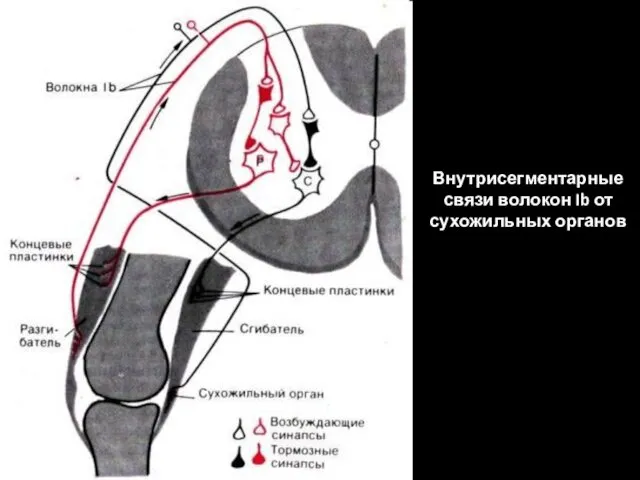
Внутрисегментарные связи волокон Ib от сухожильных органов
Внутрисегментарные связи волокон Ib от сухожильных органов

Сгибательные рефлексы
Осуществляются при раздражении кожных рецепторов, рецепторов суставов, рецепторов давления и
Сгибательные рефлексы
Осуществляются при раздражении кожных рецепторов, рецепторов суставов, рецепторов давления и
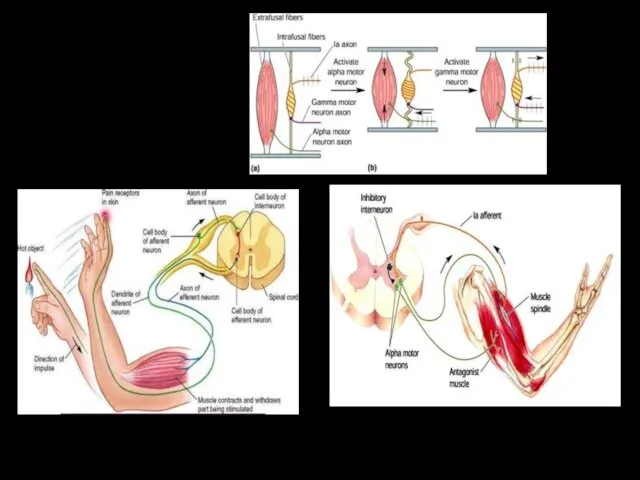
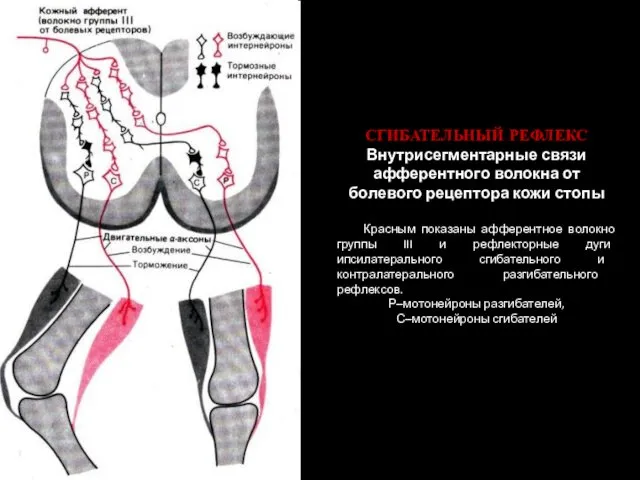
СГИБАТЕЛЬНЫЙ РЕФЛЕКС
Внутрисегментарные связи афферентного волокна от болевого рецептора кожи стопы
Красным
СГИБАТЕЛЬНЫЙ РЕФЛЕКС
Внутрисегментарные связи афферентного волокна от болевого рецептора кожи стопы
Красным
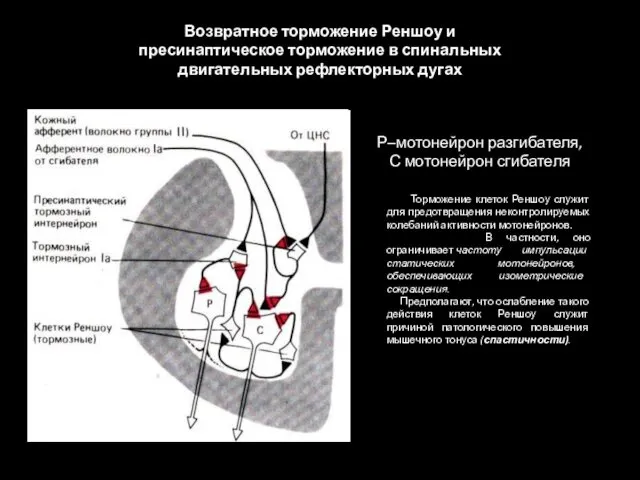
Возвратное торможение Реншоу и пресинаптическое торможение в спинальных двигательных рефлекторных дугах
Возвратное торможение Реншоу и пресинаптическое торможение в спинальных двигательных рефлекторных дугах
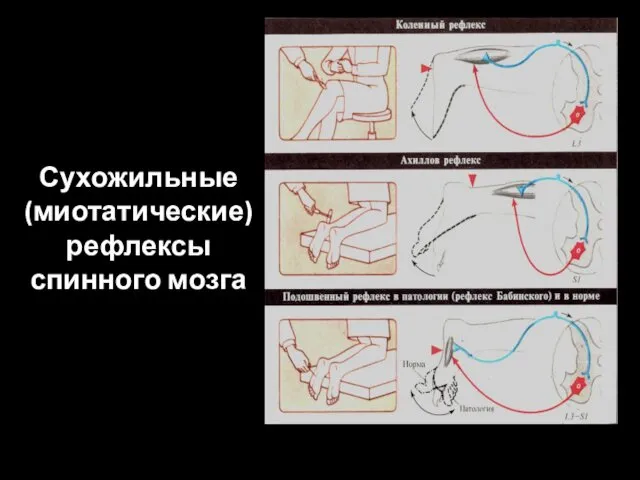
Сухожильные
(миотатические)
рефлексы спинного мозга
Сухожильные
(миотатические)
рефлексы спинного мозга
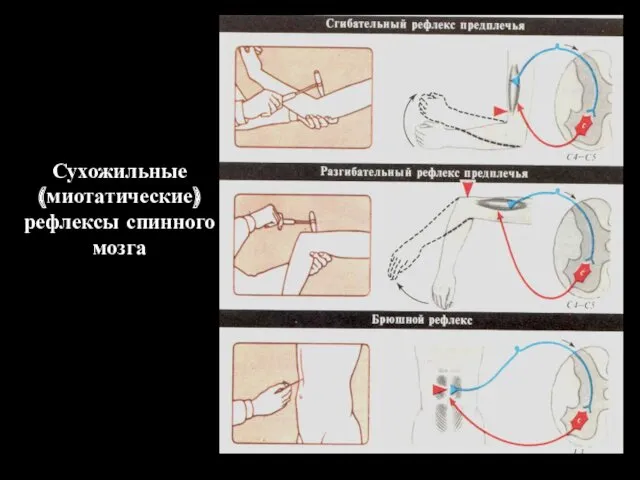
Сухожильные
(миотатические)
рефлексы спинного мозга
Сухожильные
(миотатические)
рефлексы спинного мозга
 Лечение миомы матки
Лечение миомы матки Болезни органов дыхания у детей
Болезни органов дыхания у детей Патология вагинального отростка брюшины
Патология вагинального отростка брюшины Микробиологическая диагностика гриппа и заболеваний полости рта, вызываемых вирусом герпеса
Микробиологическая диагностика гриппа и заболеваний полости рта, вызываемых вирусом герпеса Острые вирусные гепатиты
Острые вирусные гепатиты Преэклампсия кезіндегі дәрігерге дейінгі күттірмейтін көмекті ұйымдастыру
Преэклампсия кезіндегі дәрігерге дейінгі күттірмейтін көмекті ұйымдастыру Патогенез мигрени
Патогенез мигрени Комплексное лечение зубочелюстных аномалий и деформаций
Комплексное лечение зубочелюстных аномалий и деформаций Рентгеноанатомия пищеварительной системы
Рентгеноанатомия пищеварительной системы Қазақстан Республикасында сібір жарасы ауруының таралу корсеткіштері
Қазақстан Республикасында сібір жарасы ауруының таралу корсеткіштері Фармацевтическая микробиология
Фармацевтическая микробиология Трезвость – норма жизни!
Трезвость – норма жизни! Профилактические и противоэпидемические мероприятия в детских учреждениях. Оказание помощи детям при ОРВИ
Профилактические и противоэпидемические мероприятия в детских учреждениях. Оказание помощи детям при ОРВИ Закономерности распределения лекарственных веществ в организме мелких домашних животных
Закономерности распределения лекарственных веществ в организме мелких домашних животных Тіс қатарының жартылай кетіктерін ЖАПП, доғаалы протездермен емдеу көрсеткіштері
Тіс қатарының жартылай кетіктерін ЖАПП, доғаалы протездермен емдеу көрсеткіштері Жизнь и медицинские заслуги Павла Захаровича Кондоиди
Жизнь и медицинские заслуги Павла Захаровича Кондоиди Дисфункциональные маточные кровотечения
Дисфункциональные маточные кровотечения Современные подходы к диагностике и лечению вегетососудистой дистонии у пациентов детского возраста
Современные подходы к диагностике и лечению вегетососудистой дистонии у пациентов детского возраста Врачевание в древней Греции
Врачевание в древней Греции Дифференциальная диагностика острой хирургической патологии у детей
Дифференциальная диагностика острой хирургической патологии у детей Функциональная автономия щитовидной железы
Функциональная автономия щитовидной железы Medical Education in Canada
Medical Education in Canada Міокардити. Ревматична гарячка
Міокардити. Ревматична гарячка Сестринский процесс в реабилитации больных с заболеваниями дыхательной системы
Сестринский процесс в реабилитации больных с заболеваниями дыхательной системы Синдром приобретённого иммунного дефицита (СПИД)
Синдром приобретённого иммунного дефицита (СПИД) The cardiac cycle and ECG
The cardiac cycle and ECG Нефротический синдром
Нефротический синдром Миокард инфаркты
Миокард инфаркты